
Abstract
Attention switching is a crucial ability required in everyday life, from toddlerhood to adulthood. In adults, shifting attention from one word (e.g., dog) to another (e.g., sea) results in backward semantic inhibition, that is, the inhibition of the initial word (dog). In this study, we used the preferential-looking paradigm to examine whether attention switching is accompanied by backward semantic inhibition in toddlers. We found that 24-month-olds can indeed refocus their attention to a new item by selectively inhibiting attention to the old item. The consequence of backward inhibition is that subsequent attention to a word semantically related to the old item is impaired. These findings have important implications for understanding the underlying mechanism of backward semantic inhibition and the development of lexical-semantic inhibition in early childhood.
Attention switching is a crucial ability required in everyday life: A writer may have to switch from writing from a hero’s perspective to a villain’s perspective, or a politician may have to switch from contemplating health-care policies to war strategies. A typical day for a toddler also involves attention switching—from an old toy to a new toy or from learning about furniture names to animal names. Performing these actions requires the toddler to disengage attention from the no-longer-relevant item to switch attention to the currently relevant item. What mechanisms are involved in the flexible reassignment of mental resources during attention switching? Do very young children possess the same attention-switching mechanisms as adults?
Backward Inhibition
Adult studies have suggested that the ability to shift attention from one task to another is accompanied by backward inhibition, or the inhibition of an initial stimulus when attention is switched to a new stimulus. In a typical task-switching study, participants are shown a written cue at the beginning of each trial to indicate the relevant task (e.g., attending to color, orientation, or movement), followed by a four-stimulus display on a screen. Participants have to indicate which one of the four stimuli is different from the others in the dimension indicated by the cue. Afterward, each trial is categorized as an “ABA” sequence or a “CBA” sequence, on the basis of the two preceding trials: In an ABA sequence, the current trial (A) is a repetition of the trial that occurred two trials previously (with a different trial, B, intervening), whereas in a CBA sequence, new tasks or stimuli are introduced on the current A trial (making it distinct from the C and B trials that preceded it). Participants’ performance is typically impaired when they have to return to a previously attended task (as in the ABA sequence) compared with a new task (as in the CBA sequence), which indicates that attention to a new task is accompanied by persisting inhibition of the old task. Backward inhibition has been widely replicated in adult studies (Mayr & Keele, 2000; see Kiesel et al., 2010, for a review).
Backward Semantic Inhibition
While backward inhibition is mostly reported in the context of task switching, Fuentes, Vivas, and Humphreys (1999, Experiment 2) demonstrated that backward inhibition can also occur when attention is switched between different categories of semantic representations. In Fuentes et al.’s modified lexical-decision paradigm (see Fig. 1), adult participants were shown a prime word (e.g., “DOG”) followed by an intervening stimulus, which was either a word belonging to a different semantic category from the prime word (e.g., “SEA”) or a neutral string of letters (“XXX”). In the test phase, participants were shown either a word or nonword target, and they had to press a key to make a lexical decision. Importantly, the word targets in the test phase were either semantically related to the prime (e.g., “cat,” in trials analogous to the ABA sequence) or semantically unrelated to the prime (e.g., “finger,” in trials analogous to the CBA sequence). Fuentes and colleagues found that when the intervening stimulus was a word, lexical decisions were significantly slower for targets that were semantically related to the prime than for targets that were semantically unrelated—that is, backward semantic inhibition was observed. In contrast, when the intervening stimulus was a neutral string of letters (“XXX”), response times did not differ significantly regardless of whether the prime was related or unrelated to the target.

Example trial sequence and results from a lexical decision task conducted by Fuentes, Vivas, and Humphreys (1999, Experiment 2). In each trial (left), a prime word was presented at the center of the screen. This was followed by an intervening stimulus, which was either a semantically unrelated word or a neutral letter string, in the same position. A target then appeared at either the left or the right side of the screen, and participants had to make a lexical decision. Mean response time (right) for this task is shown as a function of whether the intervening stimulus was a word or a letter string and whether the prime was semantically related or semantically unrelated to the target.
Fuentes et al.’s (1999) results demonstrate that attending to a new item requires backward semantic inhibition of the previously attended item and that backward semantic inhibition can spread to new, semantically related items. Moreover, backward semantic inhibition takes place only when there is a shift of attention within semantic space (e.g., “dog-sea-cat”). There was no semantic inhibition for the intervening-stimulus condition (e.g., “dog-XXX-cat”), because no semantic processing of the neutral “XXX” stimulus was required.
Semantic Inhibition in Toddlers
To date, no study has directly examined backward semantic inhibition in toddlers. Forward semantic inhibition, however, has been demonstrated in toddler semantic-priming studies using a variation of the intermodal-preferential-looking (IPL) task. In a typical IPL study, toddlers are shown two pictures (e.g., a cat and a finger) and hear a spoken target word (e.g., “cat”). Provided that toddlers understand the target word, they spend more time fixating the named target than the distractor picture, which demonstrates a preference for the target. In a semantic-priming IPL study, the spoken target word is preceded by a prime either semantically related to the target (e.g., “dog”) or semantically unrelated to the target (e.g., “door”). For example, “Yesterday, I saw a dog [prime]. Cat [target]!” In adults, a facilitatory priming effect is typically observed in semantic-priming studies (e.g., lexical decision response time for “cat” is faster if preceded by “dog” than if preceded by “door”; Meyer & Schvaneveldt, 1971). However, no facilitatory priming effect has been observed in toddlers. Toddlers at 18 months show the same amount of target preference in the related-prime and unrelated-prime conditions. Toddlers at 24 months, in contrast, show an inhibitory semantic-priming effect: For these infants, baseline-level target recognition has been found in the related-prime condition (similarly to 18-month-olds), but impaired target recognition was found in the unrelated-prime condition (i.e., looking times to the target and distractor were at chance; Arias-Trejo & Plunkett, 2013; Styles & Plunkett, 2009; see also Arias-Trejo & Plunkett, 2009). These results in 24-month-old toddlers provide evidence of forward inhibitory effects of an unrelated prime on a target. Inhibition is inferred from the absence of systematic target looking in the unrelated-prime condition compared with the related-prime condition.
The Current Study
In the current study, we investigated whether attention switching is accompanied by backward semantic inhibition in 24-month-old toddlers by adapting the adult backward-semantic-inhibition (lexical-decision) paradigm (Fuentes et al., 1999) to an infant IPL eye-tracking paradigm (see Fig. 2). The task was simply to fixate pictures on the screen. Because labeling a picture leads to the automatic activation of the relevant semantic representations in the toddler’s lexicon (i.e., spoken-word recognition), the bimodal presentation of stimuli in this study ensured that participants engaged in attention switching between different items at the lexical-semantic level.
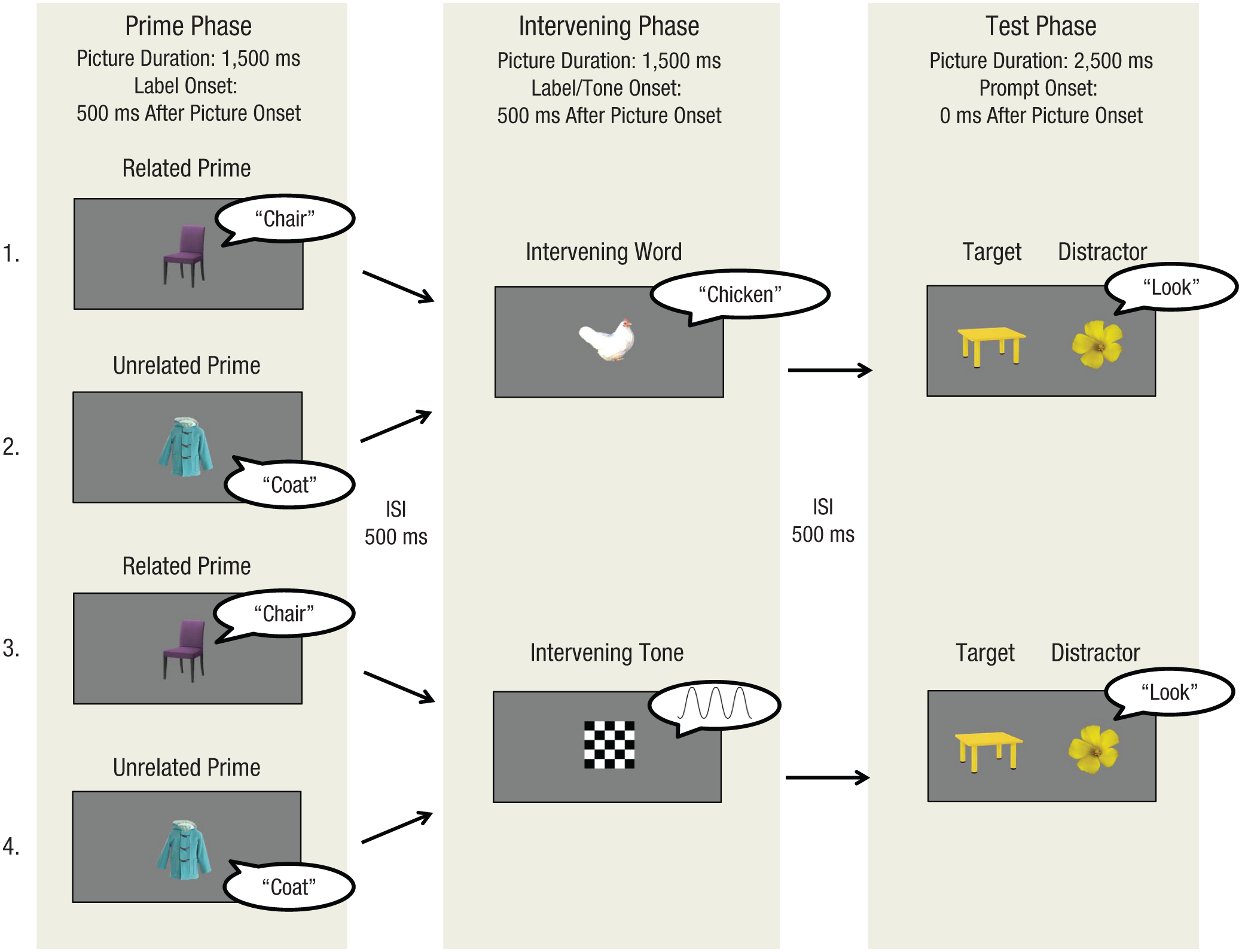
Example timeline for one trial in each of the four experimental conditions. In the prime phase, participants saw and heard a description of an object (a chair or coat) that would be related or unrelated to the target during the test phase. In the intervening phase, they saw and heard a description of a chicken or a car on half of the trials, and they saw a diamond or square checkerboard and heard a sine wave tone on the other half. In the test phase, they saw a target and distractor accompanied by an attention-getting word, and their looking times toward each object were recorded. ISI = interstimulus interval.
On the basis of the adult study (Fuentes et al., 1999), we predicted that shifting attention from the prime (e.g., “chair” or “coat”) to the intervening word (e.g., “chicken”) would result in a shift of attention within semantic space and consequent backward semantic inhibition of the prime representation. Subsequently, in the test phase, participants’ responses to the target picture would be impaired in the related-prime condition (e.g., “chair-chicken-table”) because of the inhibitory connections between the related prime (e.g., “chair”) and the target picture (e.g., “table”). In contrast, responses to the target picture would not be impaired in the unrelated-prime condition (e.g., “coat-chicken-table”). Shifting attention from the prime to an intervening tone instead of an intervening word (the tone being analogous to the letter string in the adult study) would not result in a shift of attention within semantic space. As a result, there would be no backward semantic inhibition of the prime representation, and responses to the target picture would be facilitated in the related-prime condition in comparison with the unrelated-prime condition.
Method
Participants
Thirty-five toddlers (mean age = 24.31 months, age range = 23.34−26.70; 19 males, 16 females) were recruited from British English monolingual households. Five additional participants did not complete the experiment because of fussiness. On average, 12 out of 16 trials from each participant were available for analysis. Because the effect we were investigating has not been previously studied with infants, we targeted a sample size of 30 to 40 on the basis of our previous experience conducting similar work.
Materials
The prime-phase stimuli were a picture and an audio label of either a chair (50% of trials) or a coat (50%). In the intervening phase, the stimuli were a picture and an audio label of either a chicken (25%) or a car (25%) for half of the trials, and a square (25%) or a diamond checkerboard (25%) accompanied with a tone for the other half of the trials. In the test phase, the target pictures were either a table (50%) or a hat (50%), and the distractor pictures were either a balloon (50%) or a flower (50%). The prime and target test stimuli were chosen for their semantic relatedness: chair-table and coat-hat are both taxonomically related and associated according to the Edinburgh Associative Thesaurus (Kiss, Armstrong, & Milroy, 1972). The intervening words were chosen for their semantic unrelatedness with the prime and the target. There were four experimental conditions (consisting of four trials each). 1 In two of the conditions, the primes were semantically related to the targets (e.g., chair-table), whereas in the other two conditions, the primes were semantically unrelated to the targets (e.g., chair-hat). Within each semantic-relatedness condition, four trials featured an audio label as part of the intervening stimulus, and the other four trials featured a tone as part of the intervening stimulus.
Each participant’s parent filled in the Oxford Communicative Development Inventory (Hamilton, Plunkett, & Schafer, 2000). These surveys indicated that all but 6 participants understood the labels for all eight pictures used in the current study. These 6 participants understood only four to seven words used in the study. 2
Visual stimuli
Realistic photographic representations were used. Objects were edited out of their original background and placed in the center of a 19.59 × 19.59 cm (16.8° × 16.8°) gray background using Adobe Photoshop. During the prime and intervening phases, the picture was located at the center of the screen. During the test phase, the two pictures were located in the middle left and middle right side of the screen, separated from each other by a visual angle of 19.8°. To reduce boredom, we used four different picture tokens for each prime, intervening stimulus, target, and distractor. To control for stimulus saliency during the test phase, we matched the color and luminance (given in parentheses) of the target and distractor pictures in each trial: red (120), blue (121), yellow (126), or white (140). The prime, intervening, and test pictures were never in the same color within a trial, to avoid color cuing. Within each trial, the prime and intervening pictures, if not presented in frontal views, faced in opposite directions. The prime and intervening pictures faced both the left and right side for an equal number of trials. In the test phase, the target and distractor pictures appeared on the left and right sides of the screen for an equal number of trials.
Auditory stimuli
A female Southern British English speaker recorded the auditory stimuli in child-directed speech. The duration of the prime words was 683 ms (“chair”) and 739 ms (“coat”). The duration of the intervening words was 700 ms (“chicken”) and 679 ms (“car”). The tone accompanying the square checkerboard was a sine wave tone in C, with the same duration as the word “chicken.” The tone accompanying the diamond checkerboard was a sine wave tone in D, with the same duration as the word “car.” The duration of the attention-getting words was 617 ms (“look”) and 781 ms (“wow”). Background noise and head and tail clicks were removed using GoldWave software (St. John’s, Newfoundland and Labrador, Canada).
Apparatus and procedure
Toddlers sat on their caregiver’s lap approximately 65 cm from the Tobii TX300 eye tracker (Tobii Technology, Danderyd, Sweden) and a 23-in. screen (1,920 × 1,080 resolution). The sampling rate was 120 Hz. Caregivers were instructed to keep their eyes shut, remain quiet, and refrain from any interactions with the infant during the experiment. A nine-point eye calibration was performed. The calibration attention getter was a colorful beach ball. The background color of the screen remained 50% gray throughout the experiment. Auditory stimuli were presented through a centrally located loudspeaker. In an adjacent room, the experimenter monitored the infant’s eye movements through a centrally located video camera above the screen. To achieve good calibration, we recalibrated individual locations when necessary. After calibration, toddlers were shown 16 trials, which were presented in a random order generated by the testing software. The experimenter initiated all trials by pressing a computer key when the infant’s attention was on the screen. Eye movements were recorded using an in-house eye-tracking software, Presentmate.
Each trial began with a 1,000-ms attention getter. In the 1,500-ms prime phase, participants were shown a prime picture and its audio label. This prime was semantically either related or unrelated to the target picture. The prime was followed by a 1,500-ms intervening phase, in which participants either saw a picture and heard its audio label (the word condition) or saw a checkerboard and heard a sine wave tone (the tone condition). Finally, there was a 2,500-ms test phase in which a target and a distractor picture were presented and an attention-getting word (“look” or “wow”) was heard. We did not name the picture in the test phase to avoid explicitly directing the toddlers’ attention to either picture. There was a 500-ms interstimulus interval (a blank screen in silence) between the prime and intervening phases and between the intervening and test phases. The stimulus onset asynchrony between the prime and test pictures was 4,000 ms.
Data processing
Custom code written in MATLAB (The MathWorks, Natick, MA) was used to extract fixation information from raw gaze data. Minimum fixation duration was set to 100 ms within a radius of 35 pixels. Fixation data between 0 and 2,500 ms after picture onset in the test phase was aggregated into fifty 50-ms time bins by frame (the 120 Hz eye tracker recorded six frames within 50 ms) for each condition and each participant. Probability of target fixation in each time bin was determined from the number of frames in which the participant fixated the target picture in the test phase, divided by the total number of frames in which the participant fixated the target and distractor pictures in the test phase.
Results
The overall pattern of the results is depicted in Figure 3. 3 From 300 ms to 1,400 ms after picture onset in the test phase, participants showed a similar amount of target preference in all four conditions. However, different patterns were observed from approximately 1,401 ms to the end of the trial. When the intervening stimulus was a word (as opposed to a tone), participants’ preference for the target picture was weaker in the related-prime condition than in the unrelated-prime condition. In contrast, when the intervening stimulus was a tone, participants’ preference for the target picture was stronger in the related-prime condition than in the unrelated-prime condition.

Probability of fixating targets in the test phase that were semantically related and semantically unrelated to primes, collapsed across the (a) intervening-word conditions and (b) intervening-tone conditions. Data for the period starting at 300 ms were divided into two windows of equal duration for analysis. Points indicate fixation data aggregated by 50-ms time bins, lines indicate smoothing splines, and shaded ribbons indicate 95% confidence intervals based on the smoothing splines.
The differing response patterns before and after 1,400 ms following picture onset in the test phase suggested two windows of interest. For ease of analysis, we divided the time course from 300 ms to 2,500 ms in the test phase into two equal halves: Window 1 was 300 to 1,400 ms after picture onset, and Window 2 was 1,401 to 2,500 ms after picture onset. In each window of interest, we analyzed the fixation data using growth curve and binomial logistic mixed-effects models with the R package glmmPQL (Package MASS; R Development Core Team, 2008). The overall time course of fixations was captured by third-order orthogonal polynomials (Mirman, 2014) with Level 2 fixed effects of prime type (related vs. unrelated) and intervening-stimulus type (word vs. tone) on the intercept, linear, quadratic, and cubic terms, and participant random effects on the intercept, linear, and quadratic terms. Unrelated prime and intervening tone were treated as the baseline in the model, and relative parameters were estimated for related prime and intervening word.
Table 1 summarizes the results of the growth curve model for Window 1. Collapsed across intervening-stimulus type, there was no significant effect of prime type, which suggests that there was no systematic difference between preferences in the related-prime and unrelated-prime conditions. Collapsed across prime type, there was no significant effect of intervening-stimulus type, which suggests that there was no systematic difference between the intervening-word and intervening-tone conditions. There was also no significant interaction of prime type and intervening-stimulus type. In summary, no significant effects were observed in Window 1.
Results of the Growth Curve Analysis for Window 1 (300–1,400 ms After Picture Onset in the Test Phase)

Note: The unrelated-prime and intervening-tone conditions were treated as the baseline in the model, and relative parameters were estimated for the related-prime and intervening-word conditions.
Table 2 summarizes the results of the growth curve model for Window 2. In this window, a significant effect of prime type was found after we collapsed across intervening-stimulus type. The significant intercept term indicates that participants looked more at the target in the related-prime condition than in the unrelated-prime condition. Moreover, the significant quadratic term indicates that the rate of change in the growth of target fixations differed significantly between the related- and unrelated-prime conditions. Similarly, there was a significant effect of intervening-stimulus type after we collapsed across prime type. The significant intercept term indicates that participants looked less at the target in the intervening-word condition than in the intervening-tone condition. The significant linear and quadratic terms indicate that the rate of change in the growth of target fixations differed significantly in the intervening-word and intervening-tone conditions.
Results of the Growth Curve Analysis for Window 2 (1,401–2,500 ms After Picture Onset in the Test Phase)

Note: The unrelated-prime and intervening-tone conditions were treated as the baseline in the model, and relative parameters were estimated for the related-prime and intervening-word conditions.
More important, there was a significant interaction of prime type and intervening-stimulus type on the intercept and quadratic terms. Post hoc comparisons with Bonferroni-adjusted p values were carried out to interpret the significant terms. In the intervening-word condition, participants fixated the target picture significantly less in the related-prime condition than in the unrelated-prime condition (intercept estimate = −0.23, SE = 0.05, z = −4.26, p < .001), and the curve of target fixations in the related-prime condition reached a significantly lower peak than in the unrelated-prime condition (quadratic estimate = 1.11, SE = 0.26, z = 4.21, p < .001). In the intervening-tone condition, participants fixated the target picture significantly more in the related-prime condition than in the unrelated-prime condition (intercept estimate = 0.29, SE = 0.05, z = 5.35, p < .001), and the curve of target fixations in the related-prime condition reached a significantly higher peak than in the unrelated-prime condition (quadratic estimate = −0.82, SE = 0.26, z = −3.18, p = .0175). In summary, in Window 2, we observed an effect of backward semantic inhibition when the intervening stimulus was a word, but a facilitatory semantic-priming effect when the intervening stimulus was a tone.
Discussion
These findings provide direct evidence of backward semantic inhibition in 24-month-olds and indicate that attention switching in toddlers involves backward semantic inhibition. When a shift of attention occurred from the prime to the intervening word, the prime representation was inhibited. This conclusion is substantiated by the finding that subsequent attention to the target picture was impaired in the related-prime condition, which suggests a spread of inhibition from the related-prime representation to the target representation, either via their shared semantic features or direct links between them. In contrast, when the intervening stimulus was a tone, we observed facilitatory semantic priming: Participants showed a significantly stronger preference for the target pictures in the related-prime condition. This finding echoes what was reported by Fuentes et al. (1999) in their study of backward semantic inhibition. Backward semantic inhibition did not take place when there was no shift of attention within semantic space. This is likely because processing of the intervening tone did not require refocusing of attention in the semantic domain.
Lateral inhibition versus self-inhibitory mechanisms
Two possible explanations for backward inhibition have been proposed. The first invokes lateral inhibition. Neural lateral inhibition is a well-established concept: When a target is activated, nontarget representations become inhibited to support fine-tuned selection of the target (Blakemore & Tobin, 1972; Sillito, 1975). In adult task-switching studies, the old task set and response are inhibited when they interfere with the activation of the new task set (Arbuthnott & Frank, 2000; Koch, Gade, Schuch, & Philipp, 2010; Philipp & Koch, 2006). The second explanation, a self-inhibitory mechanism, assumes that representations of a task set have inhibitory self-feedback connections. As soon as the task set is no longer required, the representations receive negative feedback and become inhibited (Grange, Juvina, & Houghton, 2013; Houghton & Tipper, 1994, 1996). An argument against a self-inhibitory mechanism is that if backward inhibition were purely “self-inflicted” in the ABA task sequence, one would expect to observe backward inhibition of task A regardless of the characteristics of task B. For example, studies using a go/no-go version of the task-switching paradigm have found no backward inhibition when task B did not require a response (e.g., Schuch & Koch, 2003).
The current study, and that of Fuentes et al. (1999), examined the switching of attention between semantic categories, rather than the switching of attention between tasks. 4 Nonetheless, our findings of a facilitatory rather than an inhibitory effect in the neutral intervening-tone condition suggest that the source of backward semantic inhibition in toddlers is unlikely to be a purely self-inhibitory mechanism, but instead involves lateral inhibition to resolve conflict between attending to old and new items.
Development of an inhibitory mechanism
The finding of backward semantic inhibition in 24-month-olds not only demonstrates that a semantic inhibitory mechanism is in place during early development to ensure efficient selective attention but also provides valuable insights into the development and mechanisms of the infant lexical-semantic system. Connectionist models of language processing (McClelland & Elman, 1986; Plaut & Booth, 2000) assume that words, concepts, and their features are connected by both excitatory and inhibitory links. As with selective attention, an inhibitory mechanism in the adult lexical-semantic system ensures the efficiency and accuracy of language processing by inhibiting nonoptimal responses. Prior to the current study, there was only limited evidence that an inhibitory mechanism exists in the infant lexical-semantic system. For example, it has been shown that 18- and 19-month-olds show a graded sensitivity to the severity of mispronunciation (e.g., target looking decreased when the number of mispronounced phonetic features of “shoe” increased from one to three: “foo,” “voo,” and “goo”; Bailey & Plunkett, 2002; White & Morgan, 2008), while 24-month-olds treat these mispronunciations as equivalent (Bailey & Plunkett, 2002). These developmental changes have been simulated using the TRACE model (Mayor & Plunkett, 2014). Mayor and Plunkett showed that the toddler data can be modeled when lexical inhibition is “switched off” at 18 months and “switched on” again at 24 months to allow interference from cohort competitors in the lexicon, which points to developmental shifts in the operation of inhibitory processes. Future experiments should examine whether semantic and lexical inhibitory mechanisms stem from a general inhibitory mechanism (e.g., inhibition of attention to a previously visually attended location has been observed in 4- to 6-month-old infants; Clohessy, Posner, Rothbart, & Vecera, 1991; Hood, 1993) or emerge separately with the acquisition of language.
Conclusion
This study demonstrates that in 24-month-old toddlers, a semantic inhibitory mechanism is in place and operates during attentional selection. As with adults, semantic processing of a new word (or concept) leads to backward semantic inhibition of an old word (or concept) in the toddler’s lexical-semantic system, and such inhibition can spread to new semantically related items. Furthermore, backward semantic inhibition is instigated only when the old and new items require attention to competing representations. We might therefore expect that 24-month-olds can refocus their attention to a new item by selectively inhibiting attention to an old item.
Footnotes
Acknowledgements
We thank Mihaela Duta, who developed the eye-tracking software Presentmate and the gaze-processing codes. We also thank Amanda Griffins and Irina Lepadatu for participant recruitment, and all the infants and parents who took part in the study.
Action Editor
Brian Ackerman served as action editor for this article.
Declaration of Conflicting Interests
The authors declared that they had no conflicts of interest with respect to their authorship or the publication of this article.
Funding
L. J. Fuentes is funded in part by the Spanish Ministry of Economy and Competitiveness (Project PSI2014-53427-P, FEDER funds), Fundación Séneca (Project 19267/PI/14), and the Leverhulme Trust.
Notes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.