
Abstract
Previous research has demonstrated that emotional information processing can be modulated by what is being held in working memory (WM). Here, we showed that such content-based WM effects can occur even when the emotional information is suppressed from conscious awareness. Using the delayed-match-to-sample paradigm in conjunction with continuous flash suppression, we found that suppressed threatening (fearful and angry) faces emerged from suppression faster when they matched the emotional valence of WM contents than when they did not. This effect cannot be explained by perceptual priming, as it disappeared when the faces were only passively viewed and not held in WM. Crucially, such an effect is highly specific to threatening faces but not to happy or neutral faces. Our findings together suggest that WM can modulate nonconscious emotion processing, which highlights the functional association between nonconsciously triggered emotional processes and conscious emotion representation.
Owing to their adaptive survival value and evolutionary salience, emotionally charged stimuli often hold a special place in working memory (WM). For example, faces can be better remembered or maintained in WM when their expressions are angry or fearful rather than happy or neutral (Jackson, Wu, Linden, & Raymond, 2009; Sessa, Luria, Gotler, Jolicoeur, & Dell’acqua, 2011). Although a plethora of research has demonstrated that emotional salience can exert a powerful modulatory effect on WM (Jackson, Linden, & Raymond, 2014; Jackson et al., 2009; Sessa et al., 2011), only a few studies have investigated the effect of WM content on emotional processing (Grecucci, Soto, Rumiati, Humphreys, & Rotshtein, 2010; Moriya, Koster, & De Raedt, 2014). It has been found that emotional faces trigger much stronger responses in the fronto-thalamic occipital network when they match an emotional word held in WM than when they do not (Grecucci et al., 2010). WM contents can also modulate the anger-superiority effect in a visual search task (Moriya et al., 2014). These studies have indicated that the processing of emotional information is susceptible to WM modulations; however, it remains to be delineated whether such content-based WM effects can take place even when the emotional information is prevented from reaching visual awareness.
WM modulation effects have been observed in the nonconscious processing of nonemotional stimuli (Gayet, Paffen, & Van der Stigchel, 2013; Pan, Cheng, & Luo, 2012; Pan, Lin, Zhao, & Soto, 2014). It has been demonstrated that WM can facilitate the processing of relevant low-level visual features (e.g., color) even when stimuli with those features are suppressed from visual awareness (Gayet et al., 2013; Pan et al., 2012). Furthermore, a recent study found that when a face in WM and a suppressed face have matching identities, WM can bias nonconscious visual processing in favor of the suppressed face, which indicates that WM modulation effects on nonconscious processing could be extended to biologically significant stimuli (Pan et al., 2014). Here, we asked whether and to what extent the processing of facial emotion can also be modulated by the content of WM in the absence of conscious awareness. There is evidence that brain regions executing top-down WM functions have reciprocal connections with those dedicated to emotional processing (Curtis & D’Esposito, 2003; Ghashghaei, Hilgetag, & Barbas, 2007; McDonald, 1998; Sah, Faber, Lopez De Armentia, & Power, 2003). Moreover, the amygdala, which is central to emotional processing, can be activated even when emotional stimuli are perceptually inaccessible to conscious awareness (Jiang & He, 2006; Morris, Öhman, & Dolan, 1999; Tamietto & de Gelder, 2010; Whalen et al., 1998). These findings raise the possibility that WM tasks could influence nonconscious processing of emotional signals.
To directly probe this issue, we used a modified version of the delayed-match-to-sample paradigm. Specifically, participants were instructed to keep a face (either fearful or neutral) in WM while performing a target-detection task. The target, another face with a new identity (fearful or neutral), was suppressed from awareness utilizing continuous flash suppression (CFS; Tsuchiya & Koch, 2005). In this technique, the target is monocularly presented and hidden from visual awareness by simultaneously presenting dynamic noise to the other eye. We measured the time it took for the suppressed face to emerge from suppression (the breaking-CFS, or b-CFS, paradigm; Jiang, Costello, & He, 2007; Stein, Senju, Peelen, & Sterzer, 2011; Yang, Zald, & Blake, 2007). We specifically tested whether faces would emerge from suppression more quickly if they matched the emotional valence of WM contents than if they did not.
Method
Participants
A total of 50 participants (mean age = 24.34 years) took part in the study. 1 Twelve (9 female, 3 male) participated in Experiment 1, 12 (8 female, 4 male) participated in Experiment 2, 12 (8 female, 4 male) participated in Experiment 3, and the remaining 14 (9 female, 5 male) participated in Experiment 4. All participants had normal or corrected-to-normal eyesight and provided informed consent. The study was approved by the institutional review board of the Institute of Psychology, Chinese Academy of Sciences. All participants were naive to the purpose of the experiments.
Stimuli
Stimuli were displayed using MATLAB (The MathWorks, Natick, MA) together with the Psychophysics Toolbox extensions (Brainard, 1997; Pelli, 1997). Face images were photographs of 36 actors (18 male and 18 female) selected from the NimStim face-stimulus set (Tottenham et al., 2009) for having the most recognizable fearful and neutral expressions. All hair and nonfacial features were removed, and only the central face area was left. The CFS display consisted of high-contrast, colored noise patterns that changed at a rate of 10 Hz. All stimuli were presented at a viewing distance of 58.5 cm against a gray background. Images presented to the two eyes were displayed side by side on the monitor and fused using a mirror stereoscope. Fusion frames (9.78° × 9.16°) and fixation crosses (0.98° × 0.98°) facilitated stable convergence of the two images.
Procedure and data analysis
Experiment 1
Each trial began with a central fixation cross presented to both eyes. In addition, a sound was played for 500 ms to alert the participant that the trial had begun. Each trial consisted of three successive stages (see Fig. 1). In the first stage (the sample period), a face subtending 3.06° × 3.72° of visual angle was presented to both eyes for 1 s, and participants were asked to memorize the face for a recognition test at the end of the trial. The second stage was a delay period in which participants performed the b-CFS task. In this stage, which lasted for 5 s, participants were asked to maintain fixation on a centrally placed cross that was continuously presented to both eyes. A CFS display was presented to one of the participant’s eyes with full contrast, and a face image (1.53° × 1.86°) was presented to the other eye at a random location within the region corresponding to the location of the noise. The contrast of the face image was increased gradually from 0 to 100% across the first 1 s of each trial and then kept constant until a response was made or 5 s elapsed. Participants were instructed to press the left or right arrow key to indicate the side of fixation on which the face appeared. If the participant pressed a key within 5 s, the noise pattern and the image disappeared, and only the fixation cross remained on the screen for the remaining time. Note that on each trial, the identity of the face images presented in the delay and sample periods was different. The final stage was a test period. In this stage, a face was displayed, and participants were asked to press the left or right arrow key to indicate whether it had the same identity as the memory cue in the sample period.

Sample trial sequence from Experiments 1, 2, and 3. In Experiment 1, participants were first asked to remember a face (the sample period), and then they were asked to detect a new face (the delay period). The target face in the delay period was presented to one eye but was suppressed from awareness by a colored noise mask presented to the other eye. Participants had to indicate whether the face (if it became visible) was on the left or right of the screen. Finally, in the test period, they indicated whether a test face had the same identity as the face they had memorized at the start of the trial. Experiment 2 followed the same procedure as Experiment 1 except that the face in the sample period was only passively viewed, and there was no test period. The procedure of Experiment 3 was also identical to that of Experiment 1, except that the face in the delay period was blended into the dynamic noise pattern, which was presented to both eyes.
Each participant completed 144 trials: 36 trials in each of four conditions created by crossing the emotional valence of the suppressed face (fearful vs. neutral) and the congruency between the emotional valence of the suppressed face and that of the face in WM (matching vs. nonmatching). The gender of the face stimuli was counterbalanced across the four conditions, the eye to which the face image was shown was counterbalanced across trials in each condition, and all test trials were presented in a new random order for each participant. The memory-test item and the memory cue shared the same identity in half of the trials, and they had the same emotional valence and gender across all trials in each condition. Response time (RT) was a measure of suppression time, that is, the time it took for participants to indicate that they saw the suppressed stimulus.
Experiment 2
Experiment 2 followed the same procedure as in Experiment 1, except that participants were asked only to passively view the faces in the sample period, and there was no test period (see Fig. 1).
Experiment 3
The procedure for Experiment 3 was the same as that for Experiment 1, with two exceptions. First, in the delay period of each trial, the suppression faces were blended into the dynamic noise patterns, and participants viewed the blended images through both eyes without binocular competition (see Fig. 1). Participants were required to perform the same task as in Experiment 1, and their perceptual experience was similar. Second, to ensure that RTs in this experiment (measured by detection time, i.e., the time it took participants to indicate that they had detected the stimulus) would be within the same range as the suppression-time RTs in Experiment 1, we modified the time course of contrast ramping so that the stimuli could reach full contrast within 5 s (as opposed to 1 s) at a ramp rate of 20% per second.
Experiment 4
In Experiment 4, the delay and test periods were the same as in Experiment 1, but the sample phase differed. In the sample period, two randomly chosen faces with different identities and expressions (one with a fearful expression and the other with a neutral expression) were sequentially presented for 1 s each in the center of the screen (see Fig. 2), with a blank interval (1 s) between them. Each face was numbered onscreen (“1” or “2”) to make the faces easier for participants to identify later. After the second face presentation, there was a 1-s interstimulus interval, in which only the fixation cross was displayed, followed by a cue (“1” or “2”) that instructed participants whether the first or the second face was the real sample face, which they would need to recall on a later recognition test. The presentation order of the two faces (fearful vs. neutral) was randomized across trials. For half of the trials, the fearful face was selected as the sample face, while for the other half of the trials, the neutral face was selected.

Sample trial sequence from Experiment 4. In the sample period, two faces were sequentially presented, followed by a digit cue (“1” or “2”) instructing participants which face to remember. The delay and test periods were exactly the same as in Experiment 1 (see Fig. 1).
Results
The proportion of correct responses on the WM task (i.e., in the test period) was .89 in Experiment 1, .93 in Experiment 3, and .85 in Experiment 4. There were no significant differences between accuracy in the fearful-face and the neutral-face conditions in these experiments, Experiment 1: t(11) = −1.74, p = .110; Experiment 3: t(11) = −1.62, p = .133; Experiment 4: t(13) = −1.37, p = .195.
For all four experiments, we collected RTs from trials with correct responses in the b-CFS task (i.e., in the delay period). We excluded trials in which RTs were more than 3 standard deviations from the mean (< 1% of all trials). RTs from Experiments 1 through 4 are shown in Figure 3. RTs from Experiment 1 were entered into a 2 × 2 repeated measures analysis of variance with the emotional valence of the suppressed face (fearful vs. neutral) and the congruency between the emotional valence of the suppressed face and that of the face in WM (matching vs. nonmatching) as within-subjects factors. Results revealed a significant main effect of emotional valence, F(1, 11) = 5.06, p = .046, η p 2 = .32, and a significant main effect of congruency, F(1, 11) = 11.86, p = .005, η p 2 = .52. Importantly, there was a marginally significant interaction between emotional valence and congruency, F(1, 11) = 4.36, p = .061, η p 2 = .28. Further analyses showed that the suppressed fearful faces emerged from suppression significantly faster if they matched WM contents in emotional valence (M = 1.56 s) than if they did not (M = 1.80 s, mean difference = −0.24, 95% confidence interval, or CI = [−0.38, −0.09]), t(11) = −3.59, p = .004, d = 0.66. This suggests that emotional information retained in WM has a direct impact on subsequent nonconscious processing of emotional signals (in this case, fear). This WM facilitation effect, however, was not found when neutral faces were involved in the b-CFS task (matching faces: M = 1.79 s, nonmatching faces: M = 1.82 s, mean difference = −0.03, 95% CI = [−0.16, 0.11]), t(11) = −0.45, p > .250, d = 0.07. Furthermore, in two additional experiments (see the Supplemental Material available online), we found that this WM facilitation effect extended to angry faces but not happy faces.
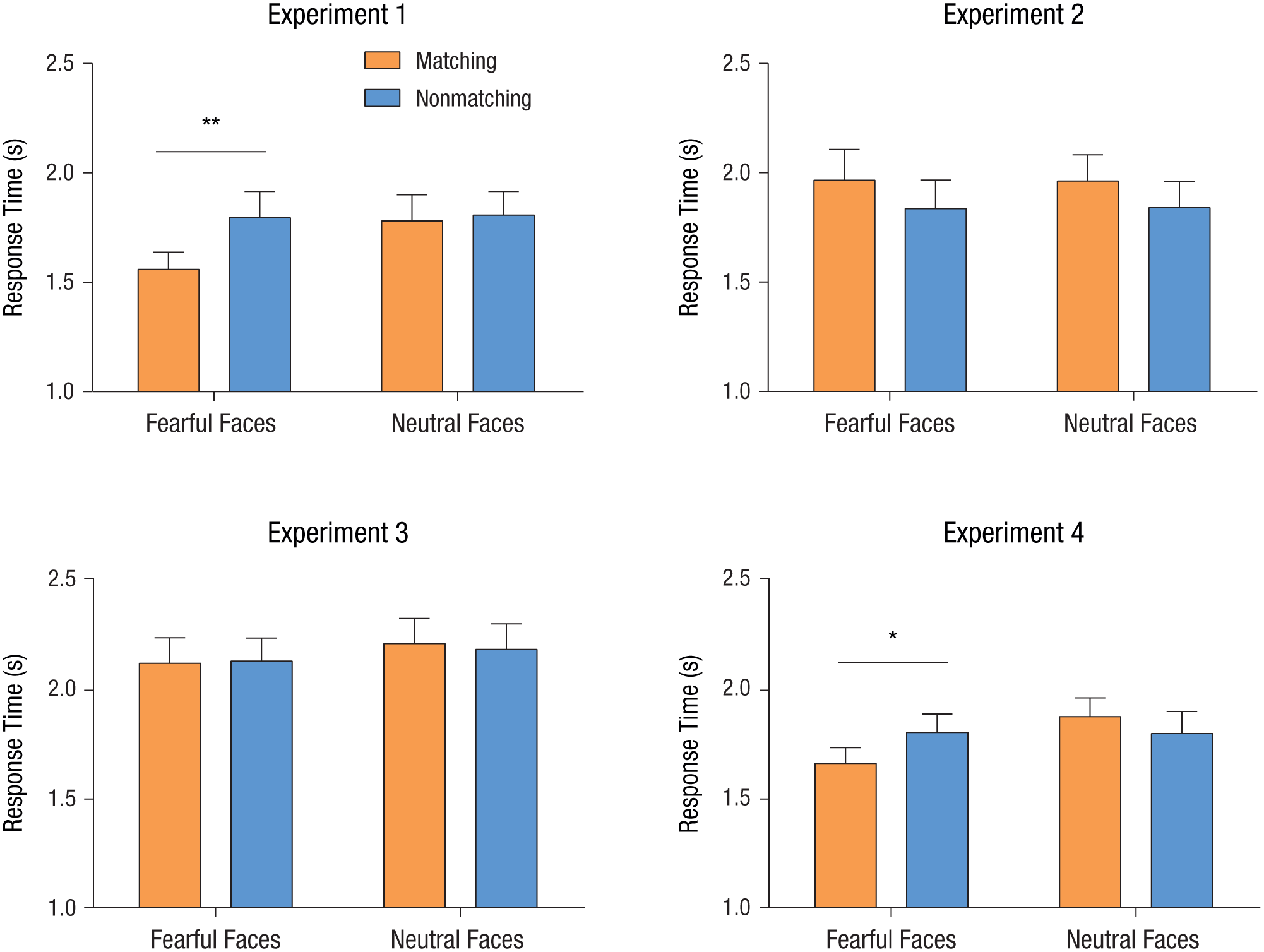
Results from Experiments 1 through 4: mean response time in the delay period as a function of suppressed faces’ emotional valence and whether that valence matched or did not match that of a face being held in working memory (Experiments 1, 3, and 4) or that had been passively viewed (Experiment 2). Error bars show +1 SE. Asterisks indicate significant differences between responses for matching and nonmatching faces (*p < .05, **p < .01).
In Experiment 2, in which participants were asked to passively view rather than memorize the faces, there were no significant effects—main effect of emotional valence, F(1, 11) = 0.00, p > .250, η p 2 = .00; main effect of congruency, F(1, 11) = 2.00, p = .185, η p 2 = .15; interaction between emotional valence and congruency, F(1, 11) = 0.01, p > .250, η p 2 = .00. Subsequent analyses showed that there were no effects of matching when fearful faces were used in the b-CFS task (matching faces: M = 1.96 s, nonmatching faces: M = 1.85 s, mean difference = 0.11, 95% CI = [−0.05, 0.31]), t(11) = 1.57, p = .144, d = 0.27; this was also true for neutral faces (matching faces: M = 1.97 s, nonmatching faces: M = 1.85 s, mean difference = 0.12, 95% CI = [−0.14, 0.38]), t(11) = 1.03, p > .250, d = 0.29. These results revealed that RTs were no longer affected by the match in emotional valence when participants were required to passively view rather than memorize faces, which indicates that the facilitation effect found in Experiment 1 cannot be accounted for by a priming mechanism but indeed reflects influences from WM. Collectively, these findings demonstrate that WM specifically modulates nonconscious processing of fearful but not neutral facial emotion.
To ensure that the results obtained in Experiment 1 reflected the differences in nonconscious visual processing rather than the differences in detection criteria or recognition speeds in the b-CFS task, we conducted an additional control experiment (Experiment 3). In Experiment 3, the target was blended into the dynamic noise pattern, and these noise patterns were presented binocularly rather than dichoptically so that the target did not interocularly compete with the suppression noise. Results from this control experiment showed that there was a significant main effect of emotional valence, F(1, 11) = 19.12, p = .001, η p 2 = .64, whereas the main effect of congruency, F(1, 11) = 2.06, p = .179, η p 2 = .16, and the interaction between emotional valence and congruency, F(1, 11) = 0.50, p > .250, η p 2 = .04, were not significant. Further analyses revealed that there were no significant differences in RTs (i.e., detection times) between fearful faces that matched and those that did not match the sample faces in emotional valence (matching faces: M = 2.12 s, nonmatching faces: M = 2.13 s, mean difference = −0.01, 95% CI = [−0.05, 0.03]), t(11) = −0.49, p > .250, d = 0.02; likewise, there were no significant RT differences for neutral faces (matching faces: M = 2.19 s, nonmatching faces: M = 2.22 s, mean difference = −0.03, 95% CI = [−0.06, 0.01]), t(11) = 1.72, p = .113, d = 0.07. This finding confirms that the WM-induced effect observed in Experiment 1 reflects an advantage in conscious access of a WM-matched face, rather than a response bias in detecting a WM-matched face compared with a non-WM-matched face.
However, it could be argued that participants initially processed the sample target more intensively in the WM task (Experiment 1) than in the passive-viewing task (Experiment 2), which might partially account for the different observations between these two experiments. In other words, the facilitation effect on visual awareness found in the WM task might simply reflect stronger initial processing of the face in the sample period rather than the maintenance of this target in WM. To test this possibility, we controlled in Experiment 4 for the initial processing level of the memorized and passively viewed items and further examined whether WM could still influence nonconscious emotion processing. We found a significant main effect of emotional valence, F(1, 13) = 6.82, p = .022, η p 2 = .34, and a significant interaction between emotional valence and congruency, F(1, 13) = 8.54, p = .012, η p 2 = .40, whereas the main effect of congruency was not significant, F(1, 13) = 0.52, p > .250, η p 2 = .04. Further analyses revealed significantly faster RTs for fearful faces that matched the emotional valence of the sample face (M = 1.67 s) than for fearful faces that matched the emotional valence of the discarded face (M = 1.81 s, mean difference = −0.14, 95% CI = [−0.26, −0.02]), t(13) = −2.59, p = .022, d = 0.46. In contrast, RTs for neutral faces that matched the emotional valence of the sample face (M = 1.88 s) did not differ significantly from the RTs for neutral faces that did not (M = 1.80 s, mean difference = 0.08, 95% CI = [−0.06, 0.21]), t(13) = 1.26, p = .231, d = 0.22. In other words, despite the fact that the sample faces and the discarded faces were initially processed at a similar level, only those maintained in WM (i.e., the sample faces) could exert a significant influence on nonconscious emotion processing.
Discussion
Recent research has demonstrated that the content of WM can prioritize relevant emotional information (Grecucci et al., 2010; Moriya et al., 2014), but whether the contents of WM could also exert an influence on emotional information that is not yet consciously processed remains unknown. In the current study, we employed emotional faces in a WM task in conjunction with the b-CFS method and found a robust modulation effect of WM on nonconscious emotion processing (Experiment 1). Specifically, suppressed fearful faces gained access into awareness more quickly when their emotional valence matched that of a face kept in WM than when they did not. The observed WM facilitation effect is in line with the accessibility hypothesis, which suggests that information retained in WM is more accessible and thus can facilitate conscious access to matching information (Baddeley, 2010). Moreover, this WM effect cannot be accounted for by a priming mechanism (Experiment 2), a difference in detection criterion (Experiment 3), or stronger initial processing of the sample face (Experiment 4). Crucially, no evidence of a WM facilitation effect was observed when neutral faces were engaged. Furthermore, the WM facilitation effect was also observed with angry faces but not with happy faces (see the Supplemental Material). In sum, the present study extends previous findings by demonstrating that the content of WM can affect emotional processing in the absence of conscious awareness, and such WM modulation effects on nonconscious processing seem to be tuned to threat-related signals (e.g., fear and anger).
The facilitation effects on nonconscious information processing observed in previous work and in the current study were evident only in the WM task (Experiment 1) but not in the passive-viewing task (Experiment 2), which reflects the critical role of WM in generating these modulation effects (Gayet et al., 2013; Pan et al., 2012). With regard to the neural mechanisms underlying WM-induced effects, it is reasonable to postulate that the cortical regions implicated in WM (e.g., the prefrontal cortex) might exert influences over different visual-processing areas associated with the perceptual awareness of relevant stimuli. Recent brain-imaging studies have shown that visual feature representations held in WM can evoke similar activity patterns in the early visual cortex as those associated with actual visual perception (Harrison & Tong, 2009; Serences, Ester, Vogel, & Awh, 2009). It has been proposed that the early visual cortex is involved in WM modulation effects of nonconscious low-level feature processing (Gayet et al., 2013; Pan et al., 2012). In a similar manner, the effect observed in the present study is likely supported by higher-level visual cortex, which is related to the conscious representation of face stimuli. Indeed, it has been demonstrated that neural activity in face-selective cortical areas (e.g., the fusiform face area, or FFA; Kanwisher, McDermott, & Chun, 1997) is greater when participants are asked to memorize a face than when they are asked to passively view a face (Chadick & Gazzaley, 2011), and this enhancement is specifically associated with the modulation effect from the frontal-parietal WM network (Chadick & Gazzaley, 2011; Ranganath, Cohen, Dam, & D’Esposito, 2004). Hence, the influence of WM on nonconscious threatening-face processing found in the present study might be the result of the feedback modulation that WM exerts over these higher-level visual areas dedicated to face-stimulus representation (e.g., FFA).
Furthermore, the WM facilitation effect found in the current study is highly specific to threatening (i.e., fearful and angry) faces but not happy or neutral faces. From a functional perspective, threatening information generally signals a need for immediate action, whereas nonthreatening information can often be safely ignored. This threat-related specificity resonates well with a wealth of previous research showing that threatening stimuli hold greater sway over perceptual experience than their nonthreatening counterparts do (Ito, Larsen, Smith, & Cacioppo, 1998; Rozin & Royzman, 2001). Our findings also add to the growing body of evidence that there is a primitive mechanism in the human brain that preferentially processes threatening stimuli for early threat detection and defense mobilization, which provides adaptive advantages in many situations (Adolphs, 2013; Öhman, Carlsson, Lundqvist, & Ingvar, 2007; Öhman & Mineka, 2001). It is likely that the amygdala, a central neural node for processing threatening signals (Dalgleish, 2004), also participates in the WM modulation. Indeed, it has been shown that the amygdala shares reciprocal projections with the prefrontal cortex, which is involved in WM, as well as the fusiform gyrus, which is related to visual perception of face stimuli (Ghashghaei et al., 2007; McDonald, 1998; Pessoa, 2008; Sah et al., 2003). Moreover, robust amygdala activity can be found even when threatening faces are presented outside of visual awareness (Jiang & He, 2006; Morris et al., 1999; Tamietto & de Gelder, 2010; Whalen et al., 1998). Although happy expressions can also activate the amygdala, the response profile of such expressions is qualitatively different from that of threatening expressions (Tamietto & de Gelder, 2010), and the results are more complicated when happy expressions are nonconsciously presented (Juruena et al., 2010; Whalen et al., 1998). It is plausible that the amygdala, which responds to both conscious and nonconscious threatening information, can work with the prefrontal cortex to modulate the neural activity that codes face stimuli in the visual ventral pathway, thereby leading to the observed effects. Yet the exact neural mechanisms mediating the WM modulation effect on nonconscious threatening-face processing remain an important question for future investigations.
Previous studies have shown that the suppression duration of visual stimuli under CFS is sensitive to various stimulus properties (Gayet, Van der Stigchel, & Paffen, 2014; Jiang et al., 2007; Stein, Senju, et al., 2011; Yang et al., 2007). For instance, it has been found that fearful faces emerge from suppression into awareness faster than neutral ones (Yang et al., 2007). Here, we extended this phenomenon and demonstrated that the suppression duration of fearful but not neutral faces can be significantly modulated by what is actively held in WM. However, we did not observe the superiority effect of fearful faces in the passive-viewing condition. Note that, differently from previous work, a face stimulus was always presented before the b-CFS task in the current experiments. We speculate that the brief presentation of a face stimulus, even though task-irrelevant, may have distracted participants’ attention. Given findings that attention allocation can modulate the behavioral and brain responses to stimuli under CFS (Hsieh, Colas, & Kanwisher, 2011; Yuval-Greenberg & Heeger, 2013), it is possible that the superiority effect of the fearful faces observed in previous studies is also susceptible to attentional modulation, which might explain the absence of the effect in the passive-viewing condition.
It should also be noted that while nonrivalrous control tasks are commonly used to examine the effects arising from conscious detection (e.g., response criterion; Alsius & Munhall, 2013; Jiang et al., 2007; Mudrik, Breska, Lamy, & Deouell, 2011), their usefulness is still a matter of debate in b-CFS studies (Gayet et al., 2014; Stein, Hebart, & Sterzer, 2011). Nevertheless, the b-CFS method is regarded as a valid and sensitive measure of access to awareness (Gayet et al., 2014; Stein, Hebart, & Sterzer, 2011). Moreover, the sharp contrasts in our experiments between results for the WM and the passive-viewing tasks and between RTs for the threatening (i.e., fearful and angry) and the nonthreatening (i.e., happy and neutral) faces provide compelling evidence that WM can prioritize the conscious access of subconscious threatening signals in the environment.
In conclusion, the present experiments provide strong evidence that WM can specifically facilitate nonconscious processing of threatening faces, which challenges a traditional view that nonconscious processing of emotional information is automatic and inflexible (Hommel, 2007; Tamietto & de Gelder, 2010). Our findings clearly demonstrate that nonconscious perception of threatening signals can be significantly modulated by one’s current conscious state (i.e., one may volitionally retain information) and thus contribute to the growing evidence that nonconscious processes can involve some nonautomatic processes (Bargh, 1994; Hassin, 2013; Soto & Silvanto, 2014). Nonconsciously mediated emotional processing may be fast and unintentional, but people can nevertheless exert conscious control over it to guide behaviors.
Footnotes
Action Editor
Eddie Harmon-Jones served as action editor for this article.
Declaration of Conflicting Interests
The authors declared that they had no conflicts of interest with respect to their authorship or the publication of this article.
Funding
This study was supported by grants from the Strategic Priority Research Program of the Chinese Academy of Sciences (No. XDB02010003), the National Natural Science Foundation of China (Nos. 31525011, 31671137, and 31100733), and the Youth Innovation Promotion Association of the Chinese Academy of Sciences (No. 2014073).
Notes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.