
Abstract
The “5-to-7-year shift” refers to the remarkable improvements observed in children’s cognitive abilities during this age range, particularly in their ability to exert control over their attention and behavior—that is, their executive functioning. As this shift coincides with school entry, the extent to which it is driven by brain maturation or by exposure to formal schooling is unclear. In this longitudinal study, we followed 5-year-olds born close to the official cutoff date for entry into first grade and compared those who subsequently entered first grade that year with those who remained in kindergarten, which is more play oriented. The first graders made larger improvements in accuracy on an executive-function test over the year than did the kindergartners. In an independent functional MRI task, we found that the first graders, compared with the kindergartners, exhibited a greater increase in activation of right posterior parietal cortex, a region previously implicated in sustained attention; increased activation in this region was correlated with the improvement in accuracy. These results reveal how the environmental context of formal schooling shapes brain mechanisms underlying improved focus on cognitively demanding tasks.
Keywords
The “5-to-7-year shift” refers to the remarkable improvements observed in children’s cognitive abilities during this age range, particularly in their ability to exert control over their own behavior. The sources of the 5-to-7 shift are proposed to lie not only within children themselves, but also in concurrent changes in their environment (Sameroff & Haith, 1996).
Across many cultures, children start school during this period. In school, they must learn to sit still and pay attention in the classroom, while avoiding being distracted by peers or other thoughts (e.g., Burrage et al., 2008; Roebers, Röthlisberger, Cimeli, Michel, & Neuenschwander, 2011). Thus, children who were previously in a less structured environment, such as a play-oriented kindergarten, experience an increase in demand on their ability to control their attention, thoughts, and behavior—a set of cognitive abilities often referred to as executive functions (EFs) or cognitive control. Converging evidence suggests that executive functioning in childhood is positively associated with later academic achievement, including math and reading abilities (e.g., Blair & Razza, 2007; Gawrilow et al., 2014) as well as social and cognitive competencies in adolescence (e.g., Zelazo & Müller, 2010). However, researchers do not know how much the experience of schooling, as compared with age-related changes in brain maturation, drives these developmental changes (Galván, 2010).
Studies exploring the effects of schooling on EFs have typically employed what is called the cutoff design, which involves comparing children who are of similar age but enrolled in school in different school years because of fixed entry dates (Morrison, Smith, & Dow-Ehrensberger, 1995). Although some studies have found beneficial effects of schooling on various aspects of executive functioning (e.g., Burrage et al., 2008; McCrea, Mueller, & Parrila, 1999; Roebers et al., 2011), the findings have been inconsistent (see Roebers et al., 2011), and the effect sizes have been small.
Despite the fact that the 5-to-7 shift has been an important topic in developmental psychology research, the emerging field of developmental cognitive neuroscience has thus far been silent about it. Although functional MRI (fMRI) studies have demonstrated a close link between development of cognitive control during middle childhood and adolescence and the maturation of frontoparietal circuitry (e.g., Luna et al., 2001; Satterthwaite et al., 2013), fMRI studies focusing specifically on the age range of 5 to 7 have been sparse (but see Sheridan, Kharitonova, Martin, Chatterjee, & Gabrieli, 2014). Electroencephalography (EEG) studies that have looked at cognitive control during this age range have revealed an age-related increase in the error positivity (Grammer, Carrasco, Gehring, & Morrison, 2014) and a positive association between the error positivity and early academic achievement (Kim et al., 2016). However, nothing is known so far about the direct effect of schooling on the development of the brain network underlying cognitive control.
To fill this knowledge gap, we conducted the current study, the first neuroscientific inquiry of changes in cognitive control and their neural correlates during the 5-to-7 shift, and how these changes relate to school entrance. By combining task-based fMRI and behavioral measures of EFs, we aimed to delineate more specifically which, if any, aspects of EF change are due to entering formal schooling. Beyond providing insights regarding mechanisms of change, examining effects of formal schooling on brain activation could be particularly helpful when neural changes precede changes in behavior and predict later academic performance (Gabrieli, Ghosh, & Whitfield-Gabrieli, 2015).
EFs are commonly subdivided into three main, separate components: inhibitory control, working memory, and cognitive flexibility (Miyake et al., 2000). These components are assumed to drive performance in different EF tasks to varying degrees. In a revised model of EFs, inhibitory control has been viewed as a general component that runs through all EF tasks (Miyake & Friedman, 2012). Inhibitory control has been further subdivided into interference control, or the ability to sustain selective attention to something while ignoring distracting information, and response inhibition, or the ability to override strong internal predispositions or external lures (Diamond, 2013). All of these EF components, including the subcomponents of inhibitory control, were required to varying degrees for successful performance of the tasks used in this study.
Interindividual differences in adults’ EFs have been found to be largely heritable (Engelhardt, Briley, Mann, Harden, & Tucker-Drob, 2015; Friedman et al., 2008), but this does not imply that individuals’ experiences do not shape EFs. Almost all individuals in Western societies undergo schooling, which makes it difficult to determine the causal effects that schooling might have on EFs.
Using a longitudinal cutoff design and two cognitive-control tasks, we tested the change in EF components across a year in two groups of similar-age children: a group who received formal schooling (first grade) and a group who attended more play-oriented kindergarten. We were thus able to assess several cognitive-control abilities behaviorally and relate any performance changes to brain activation. We predicted that EFs, and inhibitory control in particular, would improve over the year for both groups as a result of brain maturation, along with increased engagement of frontoparietal control regions, namely, ventrolateral and dorsolateral prefrontal cortex (vlPFC and dlPFC) and posterior parietal cortex (PPC). In addition, we tested three alternative hypotheses regarding the relation between formal school and changes in EFs: (a) that formal schooling magnifies these behavioral and neural changes, (b) that its effects vary across different aspects of the task and different brain regions, and (c) that it has no additional influence over and above the effects of age-related changes.
Method
Participants and general procedure
This experiment was carried out as part of a large-scale longitudinal study called the HippoKID study, which took place at the Max Planck Institute for Human Development in Berlin. In this larger study, we measured the impact of schooling on the cognitive development of 5- to 6-year-old children using a wide variety of cognitive assessments as well as functional and structural MRI measurements. Five-year-old kindergarten children born between October and March were recruited through advertisements in kindergartens, in newspapers, and on Internet forums for parents.
This age group was chosen because October falls shortly before the official cutoff date for formal schooling in Berlin (the 31st of December) and March falls shortly afterward. For example, kindergartners born during the calendar year 2008 were supposed to attend the first grade in fall 2014, whereas those born during 2009 were supposed to stay in kindergarten for 1 more year. However, because the cutoff is not definitive, parents of children born around the cutoff date can request to send their children to first grade earlier or later; most parents prefer delaying school entry. As a result, the age distributions of first graders and kindergartners in the current study overlapped (see Fig. 1, left panel).

Histograms showing the distribution of the first graders’ and kindergartners’ ages at pretest (left panel) and their pretest and posttest performance on the hearts-and-flowers task (HFT; middle and right panels).
In order to have an appropriate sample size in the larger study given the expected high dropout rates, we tested two cohorts of children: one starting in 2013 and the other in 2014. All settings were identical for the two cohorts. Because of the expected difficulties in data collection, we took a pragmatic approach toward a priori power calculation. We estimated that in order to detect an interaction effect at the lower end of the medium-size range (Cohen’s f = .20) in a Group × Time mixed-design analysis of variance (ANOVA), we would need 60 participants, given α = .05 and β = .85.
Testing for the larger study took place over two consecutive summers. Each child took part in three testing sessions per year. These sessions took place on different days; the number of days between sessions varied, depending on parents’ availability. At the pretest measurement, all the children were attending kindergarten. At the posttest measurement 1 year later, some had attended almost a full year of first grade, whereas others had continued with kindergarten. The children attended various schools and kindergartens in Berlin, but, in contrast to kindergarten classes, all first-grade classes followed the same curriculum, which was determined at the federal level. Critically, the classroom setting was more structured and goal oriented in first grade than in kindergarten, which was more play oriented.
Each session took approximately 90 min, and included a behavioral and a neuroimaging component. The latter took place in the MRI scanner and lasted about 20 min (excluding preparation). One task included in the present study, the cats-and-dogs task (CDT), was performed in the scanner; the other task included in the present study, the hearts-and-flowers task (HFT), was not (see the next section for task descriptions). The children were paid €10 per hour for their participation and additionally received a small gift after each testing session. All participants were native German speakers and were screened for psychiatric and neurological disorders through parental report. Ethics approval was obtained from the ethics committee of the German Psychological Society. The children’s parents or legal guardians gave written informed consent.
Sixty-two children (mean age at pretest = 5.40 years, range = 5.1–5.8 years) completed at least one of the two EF tasks included in the present study at both time points. Two of these children did not follow the instructions for the HFT. This resulted in a final sample of 60 children for analyses involving this task. Of these 60, 21 attended first grade in between the two measurement occasions (mean age at pretest = 5.50 years, SD = 0.15; 12 female) and 39 did not (mean age at pretest = 5.36 years, SD = 0.15; 20 female). The age of these two groups differed significantly, t(58) = 3.48, p = .001. Three of the 62 children did not complete the CDT at both pretest and posttest, and an additional 4 children did not provide valid CDT data because of problems with the MRI button box at either time point. Thus, the final sample for the behavioral analyses involving this task consisted of 55 children, of whom 19 attended first grade between the two measurement occasions (mean age at pretest = 5.50 years, SD = 0.15; 11 female) and 36 did not (mean age at pretest = 5.34 years, SD = 0.15; 20 female). The age of the two groups differed significantly, t(53) = 3.59, p = .001.
Testing 5- and 6-year-olds in an MRI scanner is challenging, particularly when they are required to perform tasks. Thus, attrition was expected to be substantial for the fMRI component of the study. However, we took several measures to reduce sample attrition as much as possible. First, to increase the children’s’ motivation, we told them that they were little astronauts embarking on a journey into their own brain with a machine that is similar to a space shuttle. Second, the children were accustomed to the MRI scanner via a mock scanner session during the first testing session. Third, while the children were lying inside the MRI scanner, they were accompanied by an experimenter who was standing next to them. Despite these measures, 11 children still had to be excluded from the fMRI analyses reported here because of excessive movement (> 3 mm). This led to a final sample of 44 children for the fMRI analyses of the CDT. Of these, 15 attended school between the two measurement occasions (mean age at pretest = 5.48 years, SD = 0.14; 10 female) and 29 did not (mean age at pretest = 5.33 years, SD = 0.16; 16 female). The age of these two groups differed significantly, t(42) = 3.15, p = .003.
Almost all of the children in the sample for this study came from families with high socioeconomic status. Parents’ mean number of years of education was high for both the first-grade group (M = 17.4, SD = 1.7, range = 14.5–20.5) and the kindergarten group (M = 16.7, SD = 2.4, range = 11.5–21.5), and did not differ between these groups, t(51) = 1.11, p = .27. Net monthly household income was also high for both the first-grade group (M = €4,310, SD = 1,347, range = €2,250–€9,000) and the kindergarten group (M = €4,629, SD = 2,100, range = €2,250–€9,000), and again there was no difference between the groups, t(44) = −0.56, p = .577. We also asked the parents to report the kind of school that their child attended (e.g., public, private, religious). All but 3 children in our sample attended public schools.
Paradigms
The HFT (see Diamond, Barnett, Thomas, & Munro, 2007) includes three conditions, all of which require sustained attention and maintenance of task rules in working memory; the conditions vary in requirements for inhibitory control and cognitive flexibility. The CDT, a go/no-go task adopted from Durston et al. (2002), was optimized for our age group to assess changes in the neural mechanisms supporting inhibitory control. As does the HFT, this task additionally requires sustained attention and maintenance of task rules in working memory.
The HFT was performed at a desktop computer during the first of the three testing sessions each year. On each trial, a red heart or flower appeared on the right or left side of the screen for 1,500 ms, and the children had to press a button with their left or right index finger during the presentation of the stimulus or during the following 500-ms fixation-cross display (i.e., response window = 2,000 ms). The task consisted of three blocks, with 20 trials per block, and the children were given instructions and practice trials prior to each block. The task parameters were nearly identical to those used by Davidson, Amso, Anderson, and Diamond (2006) except for the stimulus presentation time, which was shortened from 2,500 ms to 1,500 ms on the basis of a pilot test.
In the first block (congruent condition), a heart was presented on every trial, and the children were instructed to press the button on the same side on which the heart appeared. In the second block (incongruent condition), a flower was presented on every trial, and the children were instructed to press the button on the side opposite to the one on which the flower appeared. In the third block (mixed condition), heart and flower trials were intermixed, and the children had to continue to follow the rules learned previously, switching between the heart condition (respond on the same side) and the flower condition (respond on the opposite side). Successful performance in all three conditions of the HFT require sustained attention to the sequence of trials and maintenance of task rules in working memory. The incongruent condition additionally required inhibitory control, as it was necessary to override a prepotent response tendency, and the mixed condition additionally required cognitive flexibility, as it was necessary to switch between rules.
The CDT was performed in the MR scanner during the third of the three test sessions at each time point. Pictures of cats and dogs were presented for 500 ms each, followed by a fixation-cross display of variable duration. If the children saw a picture of a dog, they had to press a button with their right index finger (go condition), whereas if they saw a picture of a cat, they had to withhold this response (no-go condition). Button presses were counted even if they occurred during the subsequent fixation period. To optimize the statistical efficiency of our rapid event-related design, we used Optseq2 (Dale, 1999) to determine the jittered fixation periods, which ranged from 1.5 to 11.5 s (M = 3 s; the distribution followed an exponential function).
Go trials were presented 3 times as often as no-go trials. The order of presentation of go and no-go trials was pseudorandom, with the constraint that no-go trials were preceded equally often by 1, 2, 4, or 5 go trials. Sustained attention was needed to encode the appearance of stimuli in this task, and working memory was needed to remember to press a button in response to dogs but not cats. On the rare occasion when a cat was presented, response inhibition was needed to withhold the button press. Because we used different cat and dog stimuli both within and across the blocks, our task was more challenging than the typical go/no-go task, which requires remembering only a single no-go stimulus.
Before the children entered the MRI scanner, the task was explained to them by saying that they should press a button when they saw a dog but not when they saw a cat because people take dogs, but not cats, for a walk. The task consisted of three blocks and took place entirely in the scanner. To increase the children’s motivation to perform the task well, we used different pictures of cats and dogs for each of the three blocks. Two pictures of cats and four pictures of dogs were used per block. The children were familiarized with the images by presenting the six pictures to be used during a particular block together on one screen during the instructions before that block commenced.
The first block served as a training block. No scans were performed during this block, so that the experimenter could talk to the children and provide feedback. The experimenter terminated the first block manually when the children performed in accordance with the instructions for roughly 10 consecutive trials. Blocks 2 and 3 were similar except that scanning was performed. Also, these blocks consisted of 82 and 86 continuous trials, respectively, without feedback. Between blocks, the children were given a short rest and were encouraged to perform well on the task.
MRI data acquisition
T2*-weighted echo-planar images were acquired using a 3-T Siemens TIM Trio MRI scanner with a 12-channel head coil (transverse slice orientation, interleaved ascending scanning direction), field of view = 216 mm, repetition time = 2,000 ms, echo time = 30 ms, 36 slices, slice thickness = 3 mm, matrix = 72 × 72, voxel size = 3 × 3 × 3 mm, distance factor = 10%, 281 volumes each block. The first four scans of each run were discarded to ensure that a steady state of tissue magnetization was reached. Structural data were acquired using a T1-weighted 3-D magnetization-prepared rapid gradient-echo sequence (repetition time = 2,500 ms, echo time = 2,500 ms, sagittal slice orientation, spatial resolution = 1 × 1 × 1 mm).
Behavioral data analyses
It has been suggested that for children in the age range of this study, accuracy is a more sensitive measure of performance than is response time (RT; Diamond et al., 2007; see Diamond & Kirkham, 2005). However, we report both measures here. Performance was analyzed using R (R Core Team, 2014). For all ANOVAs, we used Type III sums-of-squares calculations along with effect coding, as recommended for mixed-effects designs with unequal group sizes. We set our α level at .05.
For the HFT, mean accuracy (button press on the correct side) and RT were calculated for each of the three conditions (congruent, incongruent, mixed). Statistical significance was evaluated using mixed-design ANOVAs with time (pretest, posttest) and condition (congruent, incongruent, mixed) as within-subjects factors and group (first graders, kindergartners) as a between-subjects factor.
For the CDT, mean accuracy was calculated separately for go trials (button press on trials with dogs) and no-go trials (no button press on trials with cats). In addition, mean RT was calculated for correct go trials. Statistical significance was evaluated using mixed-design ANOVAs with time (pretest, posttest) and condition (congruent, incongruent; only for accuracy) as within-subjects factors and group (first graders, kindergartners) as a between-subjects factor.
To exclude potential cohort effects that might have affected our results, we also performed all the ANOVAs with cohort as an additional between-subjects factor. To examine potential gender effects, we also ran all the ANOVAs with gender as an additional between-subjects factor.
fMRI data analyses
The fMRI data from the CDT were preprocessed and analyzed using FEAT in FSL (FMRIB’s Software Library; 1 Smith, Jenkinson, & Woolrich, 2004). Functional data were corrected for motion (MCFLIRT in FSL) and slice acquisition times (interleaved), then high-pass filtered (80 Hz), and spatially smoothed using a 5-mm full-width half-maximum Gaussian filter. Data for each child were first coregistered with the individual’s structural image and then spatially normalized into a common space. Given the young age of our sample, age-specific brain templates were created from participants’ T1 images using the nonlinear-registration ANTS program (Avants et al., 2011), following the iterative procedures of Sanchez, Richards, and Almli (2012). To allow comparison of our sample-specific activation maps with the results reported in the literature, we subsequently transferred the local maxima of the sample-specific maps into Montreal Neurological Institute (MNI) space.
Given that the existing cognitive-control literature highlights the role of frontoparietal regions, and given our a priori predictions about changes in these regions, we focused on these regions in our analyses. An anatomical bilateral frontoparietal mask was created using FSL’s Harvard-Oxford Cortical Structural Atlas. The mask encompassed dlPFC and vlPFC (i.e., middle and inferior frontal gyri), as well as posterior parietal lobe and neighboring lateral superior occipital cortex. We created z-statistic images with a voxel-wise threshold of z > 2.3 and a cluster threshold of p = .05 (corrected for family-wise error), using FLAME1 in FSL, which provides a rather conservative cluster-wise inference estimation (cf. Eklund, Nichols, & Knutsson, 2016).
After preprocessing, first-level analyses were conducted using general linear modeling. A separate model was built for each combination of task block (1 or 2; i.e., excluding the practice block) and measurement occasion (pretest or posttest). Individual time series were modeled with a gamma hemodynamic response function (500-ms boxcar function, linked to the event onsets). Four types of events were modeled with separate regressors (correct go trial, incorrect go trial, correct no-go trial, incorrect no-go trial). The regressors for incorrect trials were included as regressors of no interest.
Results from the two task blocks were combined using a within-subjects fixed-effects analysis and were normalized to the study-specific brain template. To assess whether there were changes in frontoparietal activation across time that differed by group (first graders, kindergartners), we first tested for differences between the groups at pretest. Between-subjects mixed-effects analyses were performed with FLAME in FSL to compare the two groups’ activation separately for correct go trials and no-go trials, using the variable-duration fixation periods as an implicit baseline. Next, we tested for differences between the groups 1 year later (posttest), performing the same analyses as for the pretest. To ensure that any group differences detected at posttest were not driven by group differences at pretest, we extracted percentage signal change from the contrasts at both assessments using the clusters identified at posttest. Statistical significance was evaluated using mixed-design ANOVAs with time (pretest, posttest) as a within-subjects factor and group (first graders, kindergartners) as a between-subjects factor. As in the case of the behavioral analyses, all ANOVAs were also performed with the additional between-subjects factors of cohort and gender, and Type III sums-of-squares calculations were used.
We also examined differences between the two conditions with a no-go > go contrast. No significant clusters were detected either at pretest or at posttest. The lack of control-related activation for this contrast would be of concern in a study of adults performing a typical go/no-go task, but we used a more demanding task (with a larger number of stimuli to respond to), and the young children likely found both the go and the no-go trials challenging. Given the result for this contrast, we focus here entirely on group differences separately for correct go trials and correct no-go trials.
In a parallel analysis, we sought to determine, first, whether there were frontoparietal areas displaying mean changes in the neural correlates of successful performance across participants, and, second, whether the two groups differed in the magnitude of change. Within-subjects fixed-effects analyses were performed to test for increases or decreases between pretest and posttest for the go and no-go conditions separately. Subsequent across-subjects analyses were carried out using mixed-effects models. Clusters identified in these analyses were subjected to follow-up analyses of percentage signal change, which enabled a direct comparison of the two groups’ change. As in the case of the behavioral analyses, we evaluated statistical significance using mixed-design ANOVAs with time (pretest, posttest) as a within-subjects factor and group (first graders, kindergartners) as a between-subjects factor. In addition, to explore the relationship between changes in brain activation and changes in performance in the two EF tasks, we calculated the correlation between individual change in parietal activation and mean change in accuracy in the CDT and in the HFT.
Results
Behavioral performance
A mixed-design ANOVA on HFT accuracy (see Table 1) revealed main effects of group, F(1, 58) = 4.10, p = .047, generalized η2 (ηG2) = .02; time, F(1, 58) = 19.96, p < .001, ηG2 = .05; and condition, F(2, 116) = 61.06, p < .001, ηG2 = .24, as well as a Group × Time interaction, F(1, 58) = 4.38, p = .041, ηG2 = .01, and a Condition × Time interaction, F(2, 116) = 3.33, p = .039, ηG2 = .02. Neither a Group × Condition interaction, F(2, 116) = 1.35, p = .26, nor a Group × Time × Condition interaction, F(2, 116) = 0.07, p = .93, was observed. Taken together, these results and associated post hoc analyses indicate that (a) the first graders performed better overall than the kindergartners; (b) the children’s overall task performance improved from pretest to posttest; (c) the children performed best on congruent trials and worst on mixed trials; (d) the first graders improved more on the task than the kindergartners did (see Fig. 1, middle and right panels); and (e) improvement was greater in the incongruent and mixed conditions than in the congruent condition. The Group × Time interaction, together with the lack of a Group × Time × Condition interaction, indicates that the first-graders improved more than the kindergartners in all three task conditions.
Mean Accuracy and Response Times (RTs) for the Hearts-and-Flowers Task
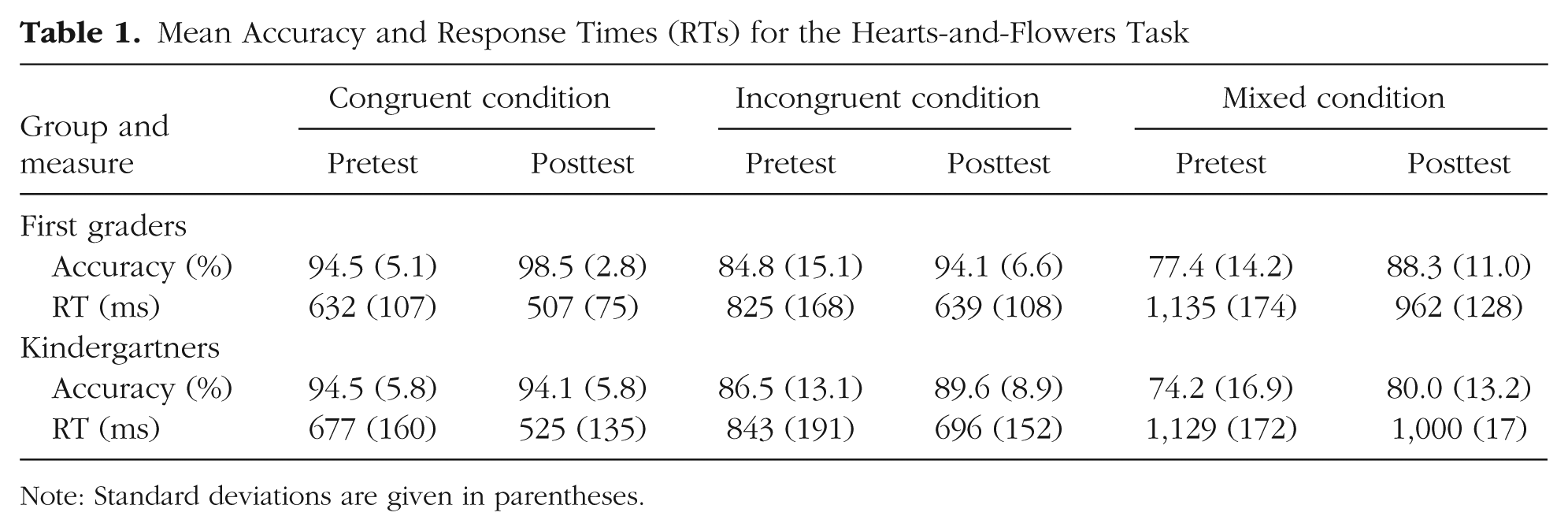
Note: Standard deviations are given in parentheses.
The mixed-design ANOVA on the HFT RTs (see Table 1) revealed main effects of condition, F(2, 116) = 536.74, p < .001, ηG2 = .60, and time, F(1, 58) = 107.23, p < .001, ηG2 = .19, but no main effect of group, F(1, 58) = 0.78, p = .38, and no significant interactions (all ps > .25). RTs were longer for the more difficult conditions and sped up over time in both groups and all conditions—that is, the improvement in RTs was independent of schooling. Overall, then, schooling led to a general improvement in accuracy in all conditions of this EF test, without a concomitant change in RTs, as in a prior intervention study involving children of this age range (Diamond et al., 2007). The lack of improvement in RTs was possibly due to the large variability in RTs at this young age (Diamond & Kirkham, 2005).
The mixed-design ANOVA on CDT accuracy (see Table 2) revealed main effects of condition, F(1, 53) = 92.49, p < .001, ηG2 = .38, and time, F(1, 53) = 7.06, p = .010, ηG2 = .02, but not of group, F(1, 53) = 0.17, p = .68, and no significant interactions (all ps > .25). Accuracy was lower overall for no-go than for go trials, and the children’s performance improved over the year, albeit similarly for the two groups and two conditions. RTs for correct go trials (see Table 2) showed a main effect of time, F(1, 53) = 14.21, p < .001, ηG2 = .05, but no main effect of group, F(1, 53) = 1.42, p = .24, and no interaction, F(1, 53) = 0.69, p = .41. In sum, the CDT results indicate that performance was improved at posttest, and that this improvement was independent of schooling.
Mean Accuracy and Response Times (RTs) for the Cats-and-Dogs Task
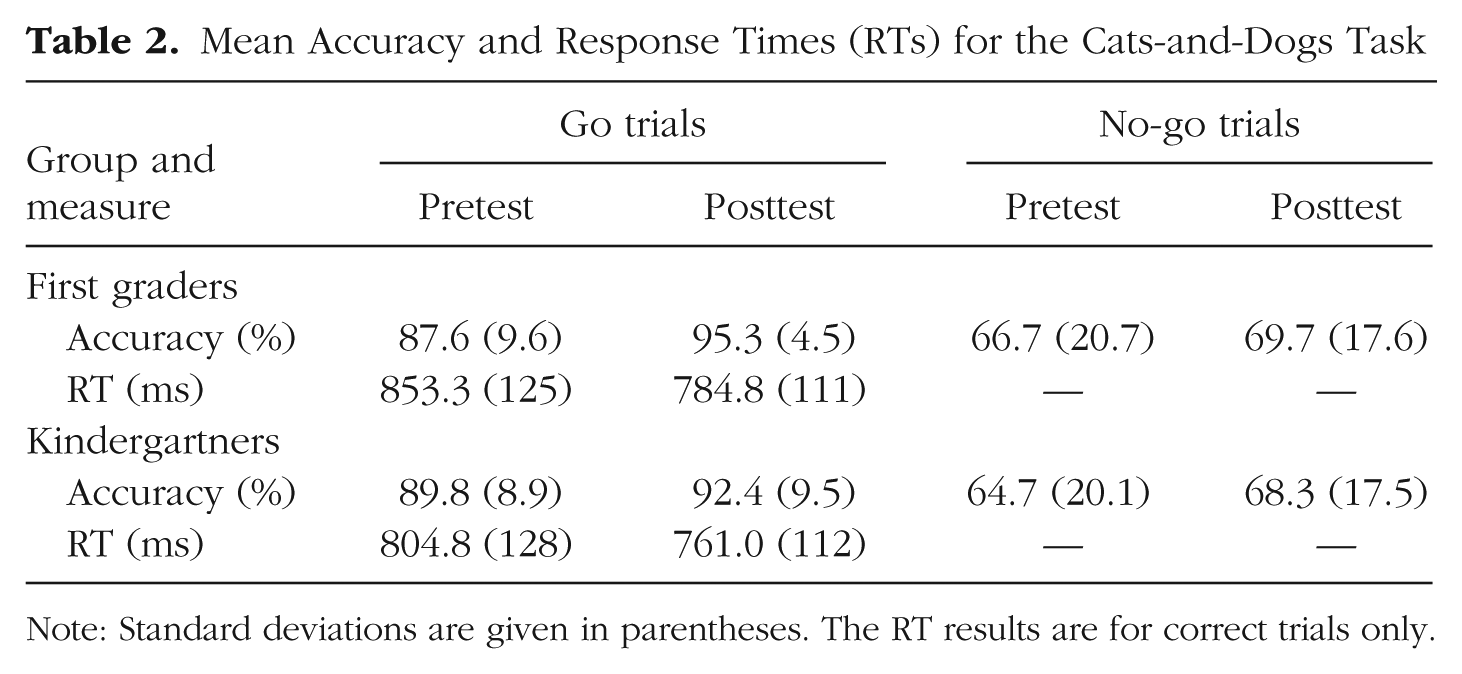
Note: Standard deviations are given in parentheses. The RT results are for correct trials only.
Children’s cohort (whether they were tested during the first or second wave of data acquisition in our study) and gender did not affect any of the reported behavioral results. There were no significant main effects or interactions involving cohort or gender (all ps > .25).
fMRI results
We first aimed to test for differences between the groups in frontoparietal activation at pretest and posttest. At pretest, no differences were detected between the two groups’ activation during either correct go or correct no-go trials. At posttest, there was again no difference between the two groups’ activation during correct no-go trials. However, for correct go trials, there were significant group differences; compared with the kindergartners, the first graders showed enhanced activation in bilateral superior PPC (peak at x = 30, y = −50, z = 50: z = 3.6; peak at x = −30, y = −44, z = 46: z = 3.4) as well as bilateral dlPFC (peak at x = 30, y = 10, z = 56: z = 3.4; peak at x = −38, y = 6, z = 46: z = 3.5; see Fig. 2, left panel).
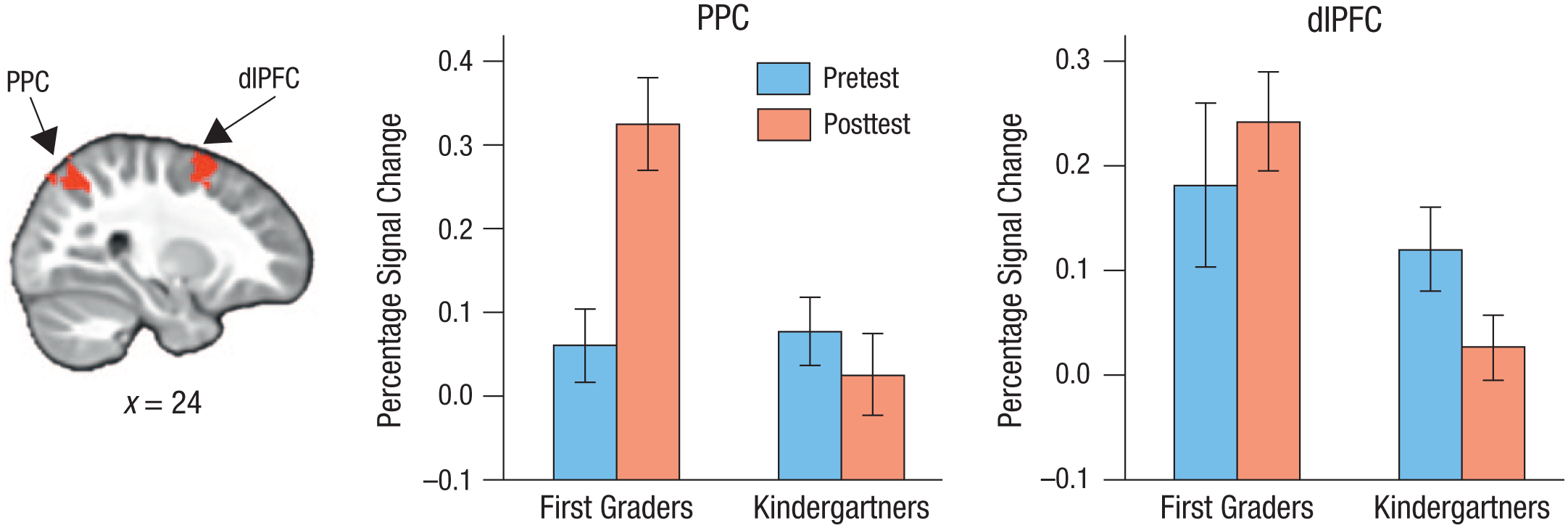
Results from the region-of-interest analysis. The image at the left shows the location of the right-hemisphere areas in posterior parietal cortex (PPC) and dorsolateral prefrontal cortex (dlPFC) where the first graders showed enhanced activation, relative to the kindergartners, during the cats-and-dogs task at posttest. The graphs show mean percentage signal change in these clusters separately for each group at pretest and posttest. Error bars indicate ±1 SE (between subjects).
To ensure that these group differences were not driven by group differences at pretest, we extracted percentage signal change in the superior PPC and dlPFC clusters from both the pretest and the posttest contrasts. For the right PPC cluster (see Fig. 2, middle panel), the ANOVA revealed a main effect of group, F(1, 42) = 6.35, p = .016, ηG2 = .08; a main effect of time, F(1, 42) = 4.72, p = .036, ηG2 = .046; and a significant Group × Time interaction, F(1, 42) = 10.51, p = .002, ηG2 = .10. Similarly, for the left PPC cluster, the ANOVA revealed a main effect of group, F(1, 42) = 6.01, p = .018, ηG2 = .09; a trend toward a main effect of time, F(1, 42) = 2.78, p = .103, ηG2 = .02; and a significant Group × Time interaction, F(1, 42) = 7.97, p = .007, ηG2 = .06. Thus, right and, to a lesser extent, left superior PPC exhibited a schooling effect that was not driven by group differences at pretest.
For the right dlPFC cluster (see Fig. 2), an ANOVA revealed a main effect of group, F(1, 42) = 6.45, p = .015, ηG2 = .08, but no main effect of time, F(1, 42) = 0.12, p = .73, and no interaction, F(1, 42) = 2.72, p = .107. Similarly, for the left DLPFC cluster, an ANOVA revealed a main effect of group, F(1, 42) = 12.06, p = .001, ηG2 = .13, but no main effect of time, F(1, 42) = 0.23, p = .633, and no interaction, F(1, 42) = 1.59, p = .215. As is apparent in Figure 2, the first graders had higher dlPFC activation than the kindergartners at pretest and displayed virtually no change at posttest, whereas the kindergartners exhibited a decrease in dlPFC activation at posttest, which contributed to the observed group difference at that assessment. Taken together, these region-of-interest analyses revealed a specific effect of schooling in the superior PPC during correct go trials.
In a follow-up analysis, we directly searched for frontoparietal regions that changed from pretest to posttest across participants and then tested whether these effects were larger for the first graders than for the kindergartners. Across participants, an increase in activation was detected for go trials in a right superior PPC cluster (peak: x = 22, y = −74, z = 46; 194 voxels; see Fig. 3, left panel), which overlapped strongly with the right superior PPC cluster that exhibited stronger activation for first graders than kindergartners at posttest. To explore potential group differences in this cluster, we extracted percentage signal change for go trials (see Fig. 3, middle panel). An ANOVA revealed a significant main effect of time, F(1, 42) = 11.75, p = .001, ηG2 = .10, and a significant Group × Time interaction, F(1, 42) = 5.24, p = .027, ηG2 = .05, but no main effect of group, F(1, 42) = 2.33, p = .134. Therefore, this analysis confirmed that the first graders showed a larger increase in the engagement of right superior PPC than the kindergartners after attending 1 year of school. No decreases in activation were observed for go trials, and no change over the year was detected for no-go trials.
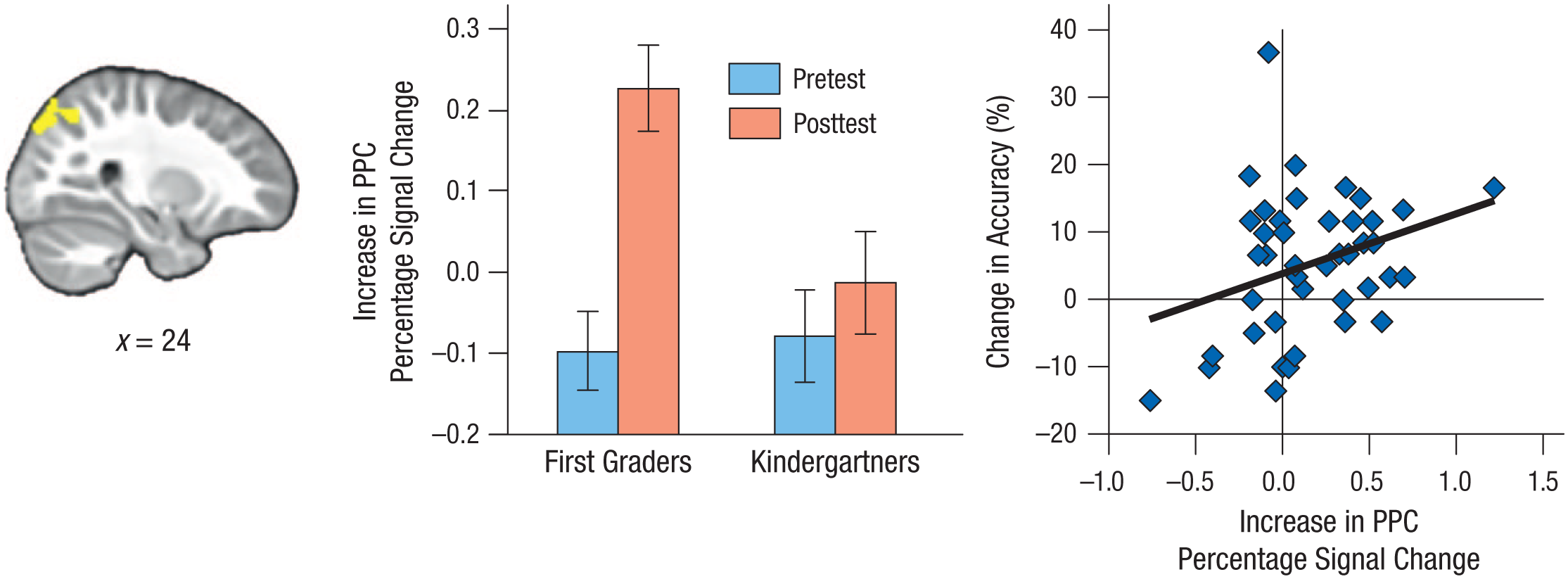
Results from the follow-up analysis of change in frontoparietal activation. The image at the left shows the location of the area in right posterior parietal cortex (PPC) where activation during correct go trials increased across participants from pretest to posttest. The middle panel shows the increase in mean percentage signal change in this cluster separately for each group at pretest and posttest. Error bars indicate ±1 SE (between subjects). The scatterplot (with best-fitting regression line) on the right shows the association between change in accuracy on the hearts-and-flowers task from pretest to posttest and increase in percentage signal change in this cluster.
Children’s cohort and gender did not affect any of the reported fMRI results; nor were there any significant main effects or interactions involving cohort or gender (all ps > .25).
To examine whether the observed increase in activation for correct go trials was related to individual differences in performance improvement from pretest to posttest, we extracted percentage signal change from the PPC cluster and correlated these data with the change in accuracy for the CDT and HFT. These analyses revealed a trend toward a positive correlation between increase in PPC activation and CDT performance (r = .22, p = .075) and a significant positive correlation between increase in PPC activation and HFT performance (r = .31, p = .023; see Fig. 3); when outliers (> 2.5 SD from the mean) were removed from the analysis, the correlation remained significant (r = .26, p = .047). The strengths of the two correlations did not differ from one another (z = 0.47, p = .32).
Discussion
This study demonstrates the impact that schooling has on the development of EFs and their neural correlates in 5- to 6-year-old children. Children exposed to formal schooling, compared with kindergartners of similar age, demonstrated greater improvements in EFs, as indicated by accuracy on the HFT. Brain activation patterns during an independent task (CDT) showed that both the first graders and the kindergartners displayed an increase in right superior PPC activation during correct go trials across 1 year. However, the increase in activation was larger for the first graders. Finally, the increase in PPC activation during the CDT was correlated with the improvement in performance on the HFT. Thus, our findings suggest that formal education contributes to age-related increases in EFs.
In the first grade, children must learn to sit still and to pay attention to teachers over a sustained amount of time, which is challenging at this age (Harnishfeger & Bjorklund, 1993). The finding that schooling led to increased engagement of superior PPC fits with this observation, as this region is key for sustained attention (Corbetta & Shulman, 2002). Our findings of changes in the dlPFC, which is also associated with cognitive control, could not be interpreted clearly because of group differences at pretest. Our findings resonate well with a prior study showing that EF training in adults led to improved attentional control, along with increased electrophysiological activity that was localized to the parietal cortex (Oelhafen et al., 2013). Thus, we conclude that, because of increased demands on sustained attention, early schooling leads to improved accuracy on an attentionally demanding task, as well as to increased engagement of the PPC.
We observed a general effect of schooling on both behavioral and neural indices of attention, rather than a particular improvement in inhibitory control. Both groups improved on the go/no-go task over the year, but there was a differential effect of schooling only on brain activation. If neural measures are generally more sensitive to change than behavioral assessments are (Gabrieli et al., 2015), the fMRI finding could be a harbinger of even larger effects of schooling on cognition over time.
A potential weakness of this study is that the no-go condition may not have selectively taxed inhibitory control in our participants. Contrary to what has been found in prior fMRI studies involving older children and adults, the activation maps for these 5- and 6-year-olds looked highly similar for no-go and go trials. This difference from prior studies may be related to the age of the participants, as maintaining the task rules in mind would be quite taxing for 5-year-olds, and our version of the task provided the additional challenge that it included several different go and no-go stimuli. The equivalent patterns of activation for go and no-go trials suggest that the children did not treat the two conditions as two separate tasks, but rather as one task that required maintenance of two rules. There is, in fact, evidence from latent-factor modeling that rule maintenance and inhibitory control are not separable functions in children of this age (Shing, Lindenberger, Diamond, Li, & Davidson, 2010).
A related point is that although right vlPFC has been implicated in inhibitory control (Aron, Robbins, & Poldrack, 2014; Bunge, Dudukovic, Thomason, Vaidya, & Gabrieli, 2002), we did not detect changes in right vlPFC activation for no-go trials across time, or a Group × Time interaction. In fact, we found no evidence of significant recruitment of vlPFC in our age group at all. To the best of our knowledge, no fMRI study has specifically looked at activation during a go/no-go task in a sample as young as ours, although Sheridan et al. (2014) also did not find vlPFC activation in a sample of 5- to 10-year-olds performing a related task, which suggests that the engagement of vlPFC for inhibitory control has a late onset.
Our results provide evidence for the notion that, despite their high heritability, EFs can be shaped by experience in a structured learning environment (see Miyake & Friedman, 2012). An important avenue for future research is to test the extent to which the heritability of EFs is attenuated or magnified by schooling. Unlike the manipulations in typical cognitive-training studies (see Simons et al., 2016), formal education is a multifaceted and fully immersive experience, which might be a prerequisite for far transfer (i.e., transfer across settings and tasks). The first graders in our study experienced a qualitative shift in modes of instruction when they left kindergarten, and thus our study is different from other studies that have examined quantitative effects of more versus less kindergarten (i.e., dosage effects; Burrage et al., 2008; Skibbe, Connor, Morrison, & Jewkes, 2011). It would additionally be interesting to test whether cumulative effects emerge across elementary school. Tentative support for a cumulative effect comes from a cross-sectional cutoff study that revealed a stronger effect of age at school entry on inhibitory control in older compared with younger children (McCrea et al., 1999).
Finally, there is a clear need for future studies to determine which classroom variables contribute to the observed EF improvements. As long as these factors remain unclear, we caution against interpreting our results to favor early schooling over curricula that emphasize playful learning (Lillard & Else-Quest, 2006). It is important to note that the first graders did not receive explicit training in EFs; nor does working with computers form part of the first-grade curriculum in Berlin, where the study took place. Further, our tasks also did not involve any school-related content. Thus, our findings suggest that formal education contributes to age-related increases in EFs. It will be important to determine whether these effects generalize across a broader range of socioeconomic status, as well as across school systems in different countries.
These findings highlight the contributions that developmental cognitive neuroscience can make to pinpointing the mechanisms of change that underlie cognitive development (see also Amso & Casey, 2006). Furthermore, they demonstrate the potential of cognitive neuroscience for identifying changes due to an intervention before those changes are fully evident in behavior. This is an important message for policymakers who evaluate educational effectiveness (Baker, Salinas, & Eslinger, 2012; Weiland & Yoshikawa, 2013). To conclude, our results reveal for the first time the strong impact that formal education exerts on normal brain development.
Footnotes
Acknowledgements
We thank Kristina Günther, our student assistants, and the courageous young participants for their invaluable help in this study.
Action Editor
Brian P. Ackerman served as action editor for this article.
Declaration of Conflicting Interests
The authors declared that they had no conflicts of interest with respect to their authorship or the publication of this article.
Funding
The HippoKID study was supported by a Minerva Research Group to Y. L. Shing from the Max Planck Society. G. Brod was supported by a Ph.D. fellowship from the International Max Planck Research School LIFE. S. A. Bunge was supported by a Jacobs Foundation Advanced Career Research Fellowship.
Open Practices
All data and materials, along with analysis scripts, have been made publicly available via the Open Science Framework and can be accessed at https://osf.io/c8gbj/. In addition, unthresholded statistical maps as well as more detailed information regarding parameters of the functional MRI analyses are available via NeuroVault and can be accessed at http://neurovault.org/collections/YYMVWBOF/. Individual-level imaging data are available upon request. The complete Open Practices Disclosure for this article can be found at http://journals.sagepub.com/doi/suppl/10.1177/0956797617699838. This article has received badges for Open Data and Open Materials. More information about the Open Practices badges can be found at ![]() .
.
Notes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.