
Abstract
Individuals differ in their ability to initiate self- and emotional-control mechanisms. These differences have been explicitly described in Kuhl’s action-control theory. Although interindividual differences in action control make a major contribution to our everyday life, their neural foundation remains unknown. Here, we measured action control in a sample of 264 healthy adults and related interindividual differences in action control to variations in brain structure and resting-state connectivity. Our results demonstrate a significant negative correlation between decision-related action orientation (AOD) and amygdala volume. Further, we showed that the functional resting-state connectivity between the amygdala and the dorsal anterior cingulate cortex was significantly associated with AOD. Specifically, stronger functional connectivity was associated with higher AOD scores. These findings are the first to show that interindividual differences in action control, namely AOD, are based on the anatomical architecture and functional network of the amygdala.
Keywords
No matter whether it is in sports or academics, we need to overcome hurdles to reach our goals. In this context, we often ask ourselves why some people get along more successfully than others and where the differences are between those who stumble and those who do not. Self- and emotional-control mechanisms have been related to personal and professional success repeatedly (Tangney, Baumeister, & Boone, 2004). Former research has shown that individuals differ in their ability to efficiently use psychological control processes such as executing an intention, defending it from competing alternatives, and inhibiting negative thoughts or unwanted negative feelings (Kuhl & Beckmann, 1994). The interindividual differences regarding the various mechanisms that are needed to obtain a predefined goal have been explicitly described in Kuhl’s (1992) action-control theory. This theory classifies individuals as either action or state oriented, depending on their ability to use one of these mechanisms. Action orientation is associated with the efficient use of self- and emotional-control processes and goal-directed behavior, whereas state orientation is related to the difficulty of recruiting sufficient self- and emotional-control processes, which hinders goal-directed behavior (Kuhl, 1992). This results in interindividual differences in how consequences and uncertainties are processed (Koole, Kuhl, Jostmann, & Vohs, 2005). On average, action-oriented individuals are less influenced by the possible consequences of an event (Koole et al., 2005). Remarkably, Quirin, Kuhl, and Düsing (2011) demonstrated that these differences also show in the physiological response to a social stressor. Their results indicate that individuals who are more action oriented appear to be more resistant to cues that induce negative affect or stress.
The propensity to be action or state oriented at a given moment is thought to depend on situations in which action control is needed (Kuhl, 1994). This leads to three scales of action control, namely (a) action orientation subsequent to failure versus preoccupation (the AOF scale), (b) prospective and decision-related action Orientation Versus Hesitation (the AOD scale), and (c) Action Orientation During (Successful) performance of Activities Versus Volatility (the AOP scale). AOF describes the way an individual copes with drawbacks and failures. Whereas action-oriented persons manage to leave the adverse incident behind to start something new (Kuhl & Beckmann, 1994), state-oriented persons tend to remain in the undesirable emotional state and ruminate about the aversive event. AOD describes an individual’s ability to initiate actions. State-oriented persons commonly struggle to begin an intentional action. This is consistent with the postponement of decisions and with procrastination (Blunt & Pychyl, 1998). AOP relates to maintaining an action-oriented mind-set for as long as it is necessary to complete a task successfully. Whereas action-oriented individuals can firmly focus on the task without being distracted by alternative actions, state-oriented persons are prone to switch between different activities without any good reason (Kuhl & Beckmann, 1994).
Up to this point, there have been various studies on the effects of interindividual differences in action control. This research has shown that, among other things, interindividual differences in action control moderate the interaction between executive functioning and self-controlled behavior (Wolff et al., 2016). Additionally, previous research has shown that differences in action control are associated with physical and mental health (Baumann, Kaschel, & Kuhl, 2005) as well as academic (Schlüter et al., 2017) and occupational (Diefendorff, 2004) performance.
By definition, being action oriented appears to be beneficial in various situations, whereas being state oriented seems to go along with some challenges. However, there is also research indicating that a state-oriented personality might be useful from time to time (Koole et al., 2005; Schlüter et al., 2017). This is especially the case in situations that make it necessary to temporarily suppress one’s needs to improve performance (Baumann et al., 2005) or in situations in which “ruminating about the possibility of failure may result in more cautious and deliberative goal-directed behavior” (Diefendorff, 2004, p. 379).
Although interindividual differences in action control make a significant contribution to our everyday life and have a major social impact, their neural basis is mostly unknown. The only evidence so far comes from electroencephalographic (EEG) studies. For instance, Pinnow, Laskowski, Wascher, and Schulz (2015) showed that action-oriented individuals show signatures of a more efficient inhibition of irrelevant information at the prefrontal level. Further, an EEG study on the role of action orientation under social stress discovered that the relative asymmetry of alpha waves toward the left frontal cortex was related to preoccupation with a stressful task (Düsing, Tops, Radtke, Kuhl, & Quirin, 2016). These studies support the assumption that individual variances in action control are related to interindividual differences in frontal activity. However, because the spatial resolution of EEG is quite limited (Jackson & Bolger, 2014), specific cortical areas are hardly detected and cannot be associated with action orientation. Furthermore, EEG signals reflect the synaptic activity of cortical neurons (Jackson & Bolger, 2014), whereas the functional activity of subcortical brain regions is barely reflected. Therefore, these studies provide only partial information on the entire networks that constitute interindividual differences in action control.
Neuroimaging techniques allow us to specifically connect interindividual differences in brain structure with those of cognitive abilities or personality (Kanai & Rees, 2011). Especially the recent shift toward a more network-driven perspective enables a novel view on the neural mechanisms that generate cognition and behavior. As part of this endeavor, analyses of functional MRI (fMRI) resting-state connectivity can be exploited to gain insight into the strengths of functional connectivities underlying interindividual differences (Fox & Raichle, 2007). For instance, studies indicated that interindividual differences in resting-state functional connectivity are correlated with cognitive abilities (Deary, Penke, & Johnson, 2010) and differences in personality (Allen & DeYoung, 2017).
None of these methods have been used to investigate the structural and functional architecture of action control. Therefore, in this study, we used a multimodal approach to examine whether interindividual differences in the structure and functional resting-state connectivity of specific brain regions can predict the interindividual variability of action control in humans.
Method
Sample-size estimation
Because this is the first study investigating the structural and functional architecture of action control, a literature-based a priori sample-size estimation could not be performed. Thus, a post hoc test was performed using G*Power software (Faul, Erdfelder, Buchner, & Lang, 2009) to compute achieved power. The analysis was based on a bivariate normal model with a small to medium effect size (r) of .20 1 (α = .05, two-tailed) in a sample of 264 participants. The analysis computed an achieved power of .91.
Participants
A total of 266 participants between 18 and 35 years old (mean age = 23.85 years; 137 males) took part in the study. The sample was composed mainly of university students with different majors, who received either a financial reward or course credit for their participation. All participants had normal or corrected-to-normal vision and hearing and no history of psychiatric or neurological disorders, and all met the standard inclusion criteria for MRI examinations. Information on the state of health was part of the demographic questionnaire and was therefore self-reported by the subjects. Data from 2 participants had to be excluded because of imaging artifacts from excessive head motion. Thus, the results are based on data from 264 participants. The study protocol was approved by the local ethics committee of the Ruhr University Bochum Faculty of Psychology (Vote 165). All participants gave written informed consent and were treated in accordance with the Declaration of Helsinki.
Acquisition and analysis of behavioral data
Action control
The Action Control Scale by Kuhl (1990; German version: HAKEMP 90) was used to capture interindividual differences in action control. This measure consists of a questionnaire that records the participants’ degree of action control under three different circumstances: (a) AOF, (b) AOD, and (c) AOP. Each of the subscales has 12 items.
For each item, a situation is briefly described, and the participant has to choose one of two possible behaviors. Each response is coded as either action or state oriented (Table 1). The individual’s degree of action control is calculated by summing the action-oriented answers of each scale, which leads to a total value between 0 and 12. High values in action control indicate action orientation and therefore advanced emotional- and self-control mechanisms.
Sample Items From Each of the Action Control Scales (Adapted from Kuhl, 1994)

Note: The three scales assessed (a) Action Orientation Subsequent to Failure Versus Preoccupation (the AOF scale), (b) Prospective and Decision-Related Action Orientation Versus Hesitation (the AOD scale), and (c) Action Orientation During (Successful) Performance of Activities Versus Volatility (the AOP scale). Action-oriented answers are highlighted in bold.
Neuroticism
To measure neuroticism as a potential confound, we used the German version of the Revised NEO Personality Inventory (Ostendorf & Angleitner, 2004). Individual neuroticism values were computed as a sum of the 48 items that constitute the Neuroticism scale. The individual score for each item depends on the answer given by the participant (−2 = strong disagreement, −1 = disagreement, 0 = neutral, 1 = agreement, 2 = strong agreement).
Acquisition of imaging data
All imaging data were acquired at the Bergmannsheil Hospital in Bochum, Germany, using a 3T Philips Achieva scanner (Best, The Netherlands) with a 32-channel head coil.
Anatomical imaging
For the segmentation of brain scans into gray- and white-matter sections, as well as for the identification of anatomical landmarks for the connectivity analyses, a T1-weighted high-resolution anatomical image was acquired—magnetization-prepared rapid-acquisition gradient echo, repetition time (TR) = 8,179 ms, echo time (TE) = 3.7 ms, flip angle = 8°, 220 slices, matrix size = 240 × 240, resolution = 1 mm × 1 mm × 1 mm). The acquisition time of the anatomical image was 6 min.
Resting-state imaging
For the analysis of functional connectivity, fMRI resting-state images were acquired using echo-planar imaging (EPI; TR = 2,000 ms, TE = 30 ms, flip angle = 90°, 37 slices, matrix size = 80 × 80, resolution = 3 mm × 3 mm × 3 mm). Participants were instructed to lie still with their eyes closed during the resting-state sequence. The acquisition time of the resting-state images was 7 min.
Analysis of imaging data
Analysis of anatomical data
We used published surface-based methods in FreeSurfer (Version 5.3.0; Fischl, 2012) to reconstruct the cortical surfaces of the T1-weighted images. The details of this procedure have been described elsewhere (Fischl, Sereno, & Dale, 1999). The automatic reconstruction steps included skull stripping, gray- and white-matter segmentation, as well as reconstruction and inflation of the cortical surface. These processing steps were performed for each participant individually. After preprocessing, each segmentation was quality controlled slice by slice, and inaccuracies were corrected manually, if necessary. After segmentation into gray and white matter, the brain was further segmented into 42 specific brain regions per hemisphere (Fig. 1a). These regions were extracted by an automatic segmentation procedure in FreeSurfer. Thirty-four of the 42 regions were extracted as cortical gray-matter regions following a gyral/sulcal-based parcellation procedure on the reconstructed cortical surface (Desikan et al., 2006). The remaining 8 regions were extracted as subcortical regions (Fischl et al., 2002). Also, the cortical white matter was segmented into 34 distinct regions per hemisphere. Each white-matter voxel was labeled according to the nearest cortical gray-matter voxel within a distance limit of 5 mm. This resulted in 34 white-matter regions corresponding to the 34 gyral-labeled gray-matter regions (Klein et al., 2014). Subsequently, the gray-matter volume (GMV) and white-matter volume (WMV) of each cortical region, and the GMV of each subcortical region, were assessed. Moreover, 6 regions representing the four ventricles of the brain were extracted to serve as a reference for later blood-oxygen-level-dependent signal analyses.
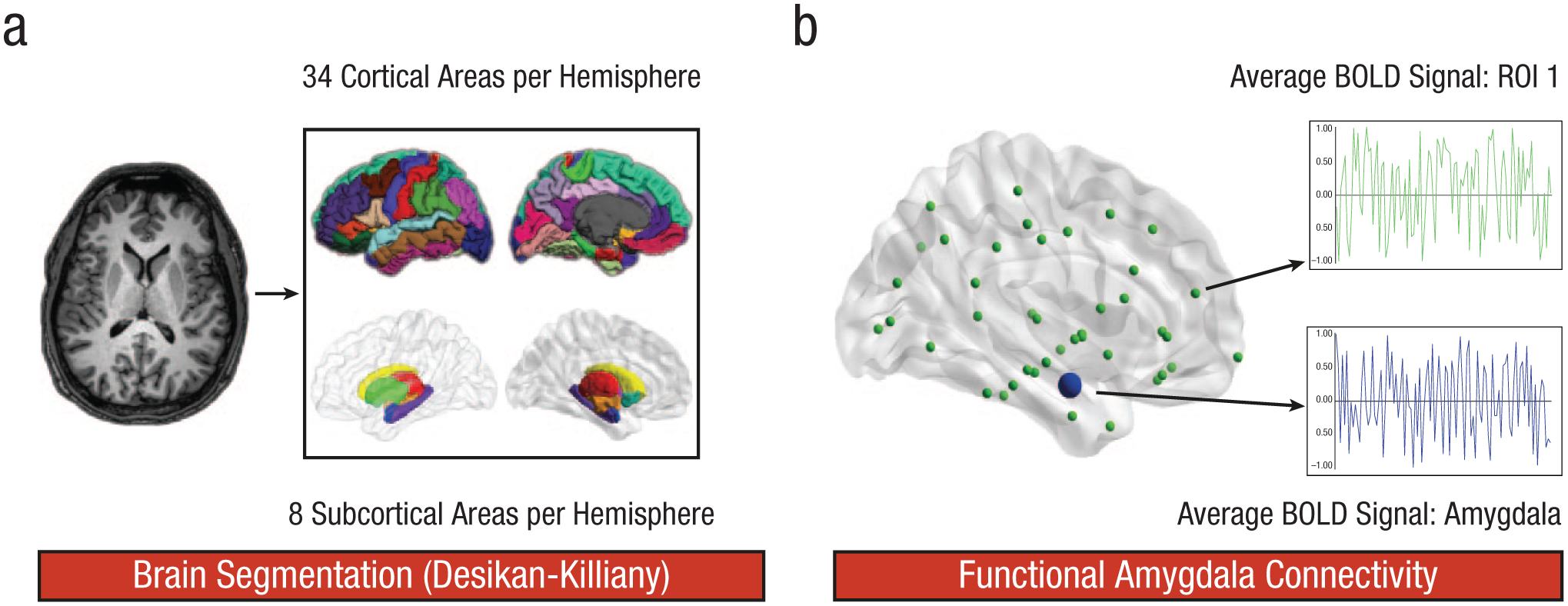
Methodological sequence for the parcellation of the brain and the analysis of amygdala resting-state connectivity. First (a), T1-weighted images were segmented into 34 cortical and 8 subcortical brain regions per hemisphere according to the Desikan-Killiany atlas. For each of these brain regions, gray-matter and white-matter volume were computed. Second, brain regions were linearly transformed into the native space of the resting-state images. Third (b), the functional connectivity between the amygdala and every other brain region of interest (ROI) was investigated. Thus, we obtained the correlation between the subcortical and cortical regions of both hemispheres and the left or right amygdala, respectively. All correlation coefficients were transformed using Fisher’s r-to-z′ transformation. These z-transformed connectivity values were averaged to determine the mean correlation of the amygdala and each cortical and subcortical region. BOLD = blood oxygen level dependent.
Finally, we examined structure–function relationships on the level of single brain regions and correlated action control (AOF, AOD, AOP) with the volume of each of the gray- and white-matter regions. To do so, we averaged the properties of each brain region across both hemispheres, which resulted in 42 regions for GMV and 34 regions for WMV (no subcortical regions were available). In the last step, all 42 subcortical and cortical regions from each hemisphere, as well as ventricular regions, were linearly transformed into the native space of the resting-state images using “mri_label2vol” in FreeSurfer, so we could continue the analysis in FSL (www.fmrib.ox.ac.uk/fsl). Data from each participant were visually inspected to confirm that the transformation procedure was successful.
Analysis of resting-state data
Resting-state data were preprocessed using MELODIC, which is also a part of the FSL toolbox. Images were preprocessed in three steps. We discarded the first two EPI volumes from each resting-state scan to allow for signal equilibration, applied motion and slice-timing correction, and filtered high-pass temporal frequencies (0.005 Hz cutoff). Spatial smoothing was not applied to avoid introducing spurious correlations in neighboring voxels. For each of the subcortical and cortical regions acquired from the automatic segmentation described above, we calculated a mean resting-state time course by averaging the preprocessed time courses of corresponding voxels.
Functional amygdala connectivity
The structural analysis of the imaging data yielded a significant association between amygdala volume and action orientation. Because the amygdala is functionally connected to various cortical and subcortical brain regions (Roy et al., 2009), the influence of interindividual differences in the amygdala’s functional connectivity on interindividual differences in action orientation was analyzed (Fig. 1b). For this purpose, we used the amygdala as a seed and computed partial correlations between the average time courses of the left and right amygdala and all remaining 41 cortical and subcortical regions while controlling for several nuisance variables. We regressed out the trajectories of all six motion parameters as well as the mean time courses averaged across all voxels representing white matter or cerebrospinal fluid, which were also obtained from the automatic FreeSurfer segmentation mentioned above. Thus, we obtained the correlation between the subcortical and cortical regions of both hemispheres and the left or right amygdala, respectively. All correlation coefficients were transformed using Fisher’s r-to-z′ transformation. These z-transformed connectivity values were averaged to get the mean correlation of the amygdala and each cortical and subcortical region. Finally, we tested whether the interindividual differences in the functional network connectivity between the amygdala and other brain regions was correlated with interindividual differences in action or state orientation. For this purpose, the individual’s z′-transformed connectivity values between the amygdala and each of the brain regions were correlated with the individual’s action-orientation score (AOF, AOD, AOP).
Statistical analysis
Statistical analyses were performed using MATLAB (Version 7.14.0.739; The MathWorks, Natick, MA) and SPSS (Version 20; SPSS, Chicago, IL). For all analyses, linear parametric methods were used. Testing was two-tailed, with an alpha level of .05.
Analysis of sex differences
We analyzed our behavioral data with respect to potential sex differences. To this end, we compared males and females regarding action orientation (AOF, AOD, AOP) by using two-sample t tests.
Correlation analysis
Because we found significant sex differences regarding action and state orientation (see Fig. S1 in the Supplemental Material available online) and because previous research indicates a consistent link between age and brain volume as well as between sex and brain volume (Pakkenberg & Gundersen, 1997), we used age and sex (male = 0, female = 1) as nuisance variables for our correlation analyses. Therefore partial correlation coefficients were computed between action orientation and the structure or the functional connectivity of single brain regions. These analyses were controlled for multiple comparisons by correcting the alpha level using a Bonferroni factor of 42 (α = .05/42 = .001).
Results
Our analyses yielded significant sex differences regarding action control. AOF scores showed that females (M = 5.37, SD = 3.05) tended to be significantly less action oriented than males (M = 6.91, SD = 2.97), t(262) = 4.19, p < .001, when it comes to handling drawbacks and failures (Fig. S1a). This means that men are more capable of leaving negative emotions behind to start something new (Kuhl & Beckmann, 1994), whereas women tend to remain in the undesirable emotional state and ruminate about the aversive event. On the contrary, AOD scores revealed that males (M = 5.58, SD = 3.08) turned out to be significantly less action oriented when it comes to initiating actions than females were (M = 7.35, SD = 2.92), t(262) = −4.80, p < .001 (Fig. S1b). Thus, men, who in this case tend to be more state oriented, struggle to begin an intentional action. There was no significant sex difference regarding AOP scores, t(262) = −1.38, p = .168 (Fig. S1c), which means that males (M = 9.46, SD = 2.12) and females (M = 9.83, SD = 2.20) are equally able to maintain an action-oriented mind-set as long as it is necessary to complete a task successfully without being distracted by alternative actions (Kuhl & Beckmann, 1994).
Subsequently, we analyzed the neural signatures of action control. First, we tested whether differences in structural brain properties can predict interindividual differences in action control. For all of the 42 brain regions (Fig. 1), GMV and WMV were extracted and correlated by controlling for multiple comparisons with the AOF, AOD, and AOP scales. Interestingly we found a significant negative association only between decision-related action orientation (AOD) and GMV of the amygdala (r = −.24, p < .001; Table 2, Fig. 2). This result indicates that individuals who are state oriented when it comes to initiating actions and therefore tend to hesitate or procrastinate show higher amygdala volume. Because previous research indicates that both amygdala volume and action orientation are associated with other personality constructs, such as neuroticism (Allen & DeYoung, 2017; Kuhl & Beckmann, 1994), we repeated the previous analysis adding neuroticism as a nuisance variable. However, the pattern of association between AOD and amygdala volume remained the same (r = −.20, p = .001).
Correlations Between AOD Score and Brain Volume in Each of the 42 Regions of Interest
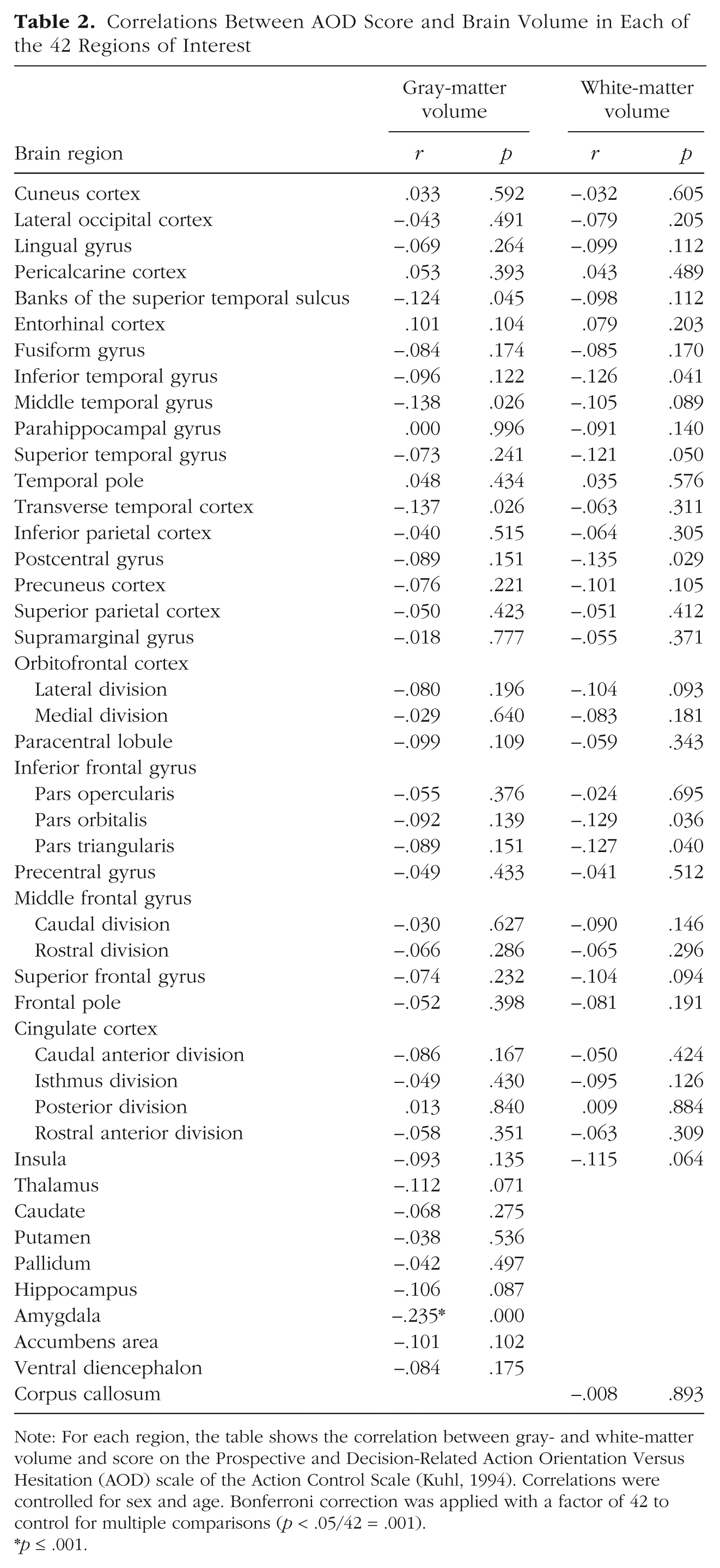
Note: For each region, the table shows the correlation between gray- and white-matter volume and score on the Prospective and Decision-Related Action Orientation Versus Hesitation (AOD) scale of the Action Control Scale (Kuhl, 1994). Correlations were controlled for sex and age. Bonferroni correction was applied with a factor of 42 to control for multiple comparisons (p < .05/42 = .001).
p ≤ .001.
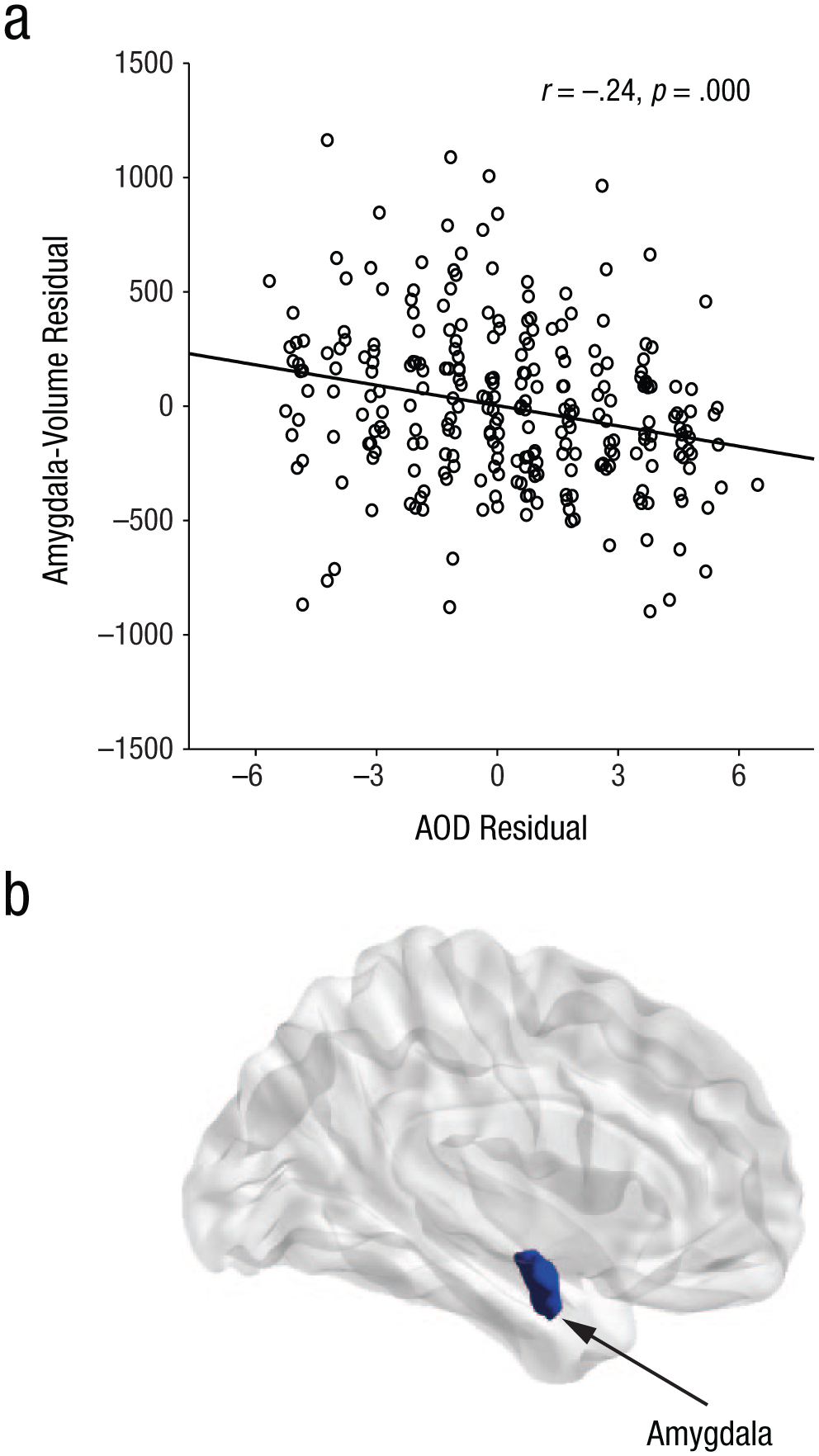
Results of the correlation analysis between amygdala volume and score on the Prospective and Decision-Related Action Orientation Versus Hesitation (AOD) scale of the Action Control Scale (Kuhl, 1994). The scatterplot (a; with best-fitting regression line) illustrates the statistically significant relationship (N = 264, p < .001) between AOD scores and gray-matter volume of the amygdala. The brain image (b) shows the location of the amygdala within the cortex.
There was no significant correlation between AOD and WMV of any region (Table 2). Moreover, there was no significant correlation between GMV of any brain region and the scales of (a) action orientation after failure (AOF) or (b) action orientation during (successful) performance of activities (AOP), nor between WMV of any region and either of these scales (see Tables S2 and S3 in the Supplemental Material).
Because the amygdala is structurally and functionally connected with a large number of subcortical and cortical structures (Roy et al., 2009), we tested whether differences in the amygdala’s functional resting-state connectivity were associated with interindividual differences in action control. To do so, we used the amygdala as a seed region and analyzed the resting-state connectivity between the amygdala and the remaining 41 cortical and subcortical brain regions. These values for resting-state connectivity were in turn correlated by controlling for multiple comparisons with the AOF, AOD, and AOP scores of each individual. This analysis indicated that interindividual differences in AOD were predicted by interindividual differences in resting-state connectivity between the amygdala and the dorsal anterior cingulate cortex (dACC; FreeSurfer nomenclature: caudal anterior division of the cingulate cortex; Desikan et al., 2006; r = .20, p = .001; see Table 3 and Fig. 3). This structure–function relationship showed a significant positive association, indicating that high functional resting-state connectivity between the amygdala and the dACC was related to higher action orientation in decision-related contexts. As stated earlier, this connection could also be the result of other psychological factors (Allen & DeYoung, 2017). Thus, we also controlled this analysis for neuroticism. Nonetheless, the correlation pattern between AOD and the functional connectivity between the amygdala and the dACC remained the same (r = .20, p = .001). In addition, there was no significant correlation between any of the other action-control scales and the resting-state connectivity seeded from the amygdala (see Tables S3 and S4 in the Supplemental Material).
Correlations Between AOD Score and Functional Amygdala Connectivity With Each of the 42 Regions of Interest
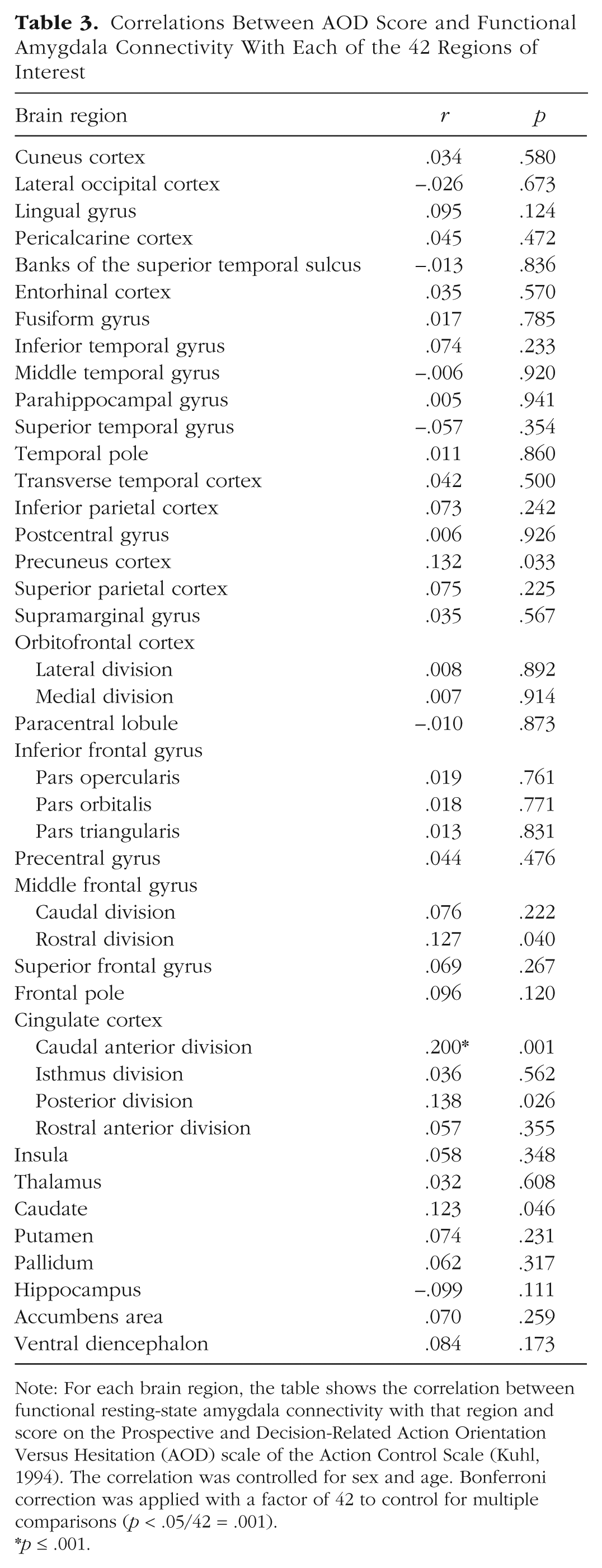
Note: For each brain region, the table shows the correlation between functional resting-state amygdala connectivity with that region and score on the Prospective and Decision-Related Action Orientation Versus Hesitation (AOD) scale of the Action Control Scale (Kuhl, 1994). The correlation was controlled for sex and age. Bonferroni correction was applied with a factor of 42 to control for multiple comparisons (p < .05/42 = .001).
p ≤ .001.
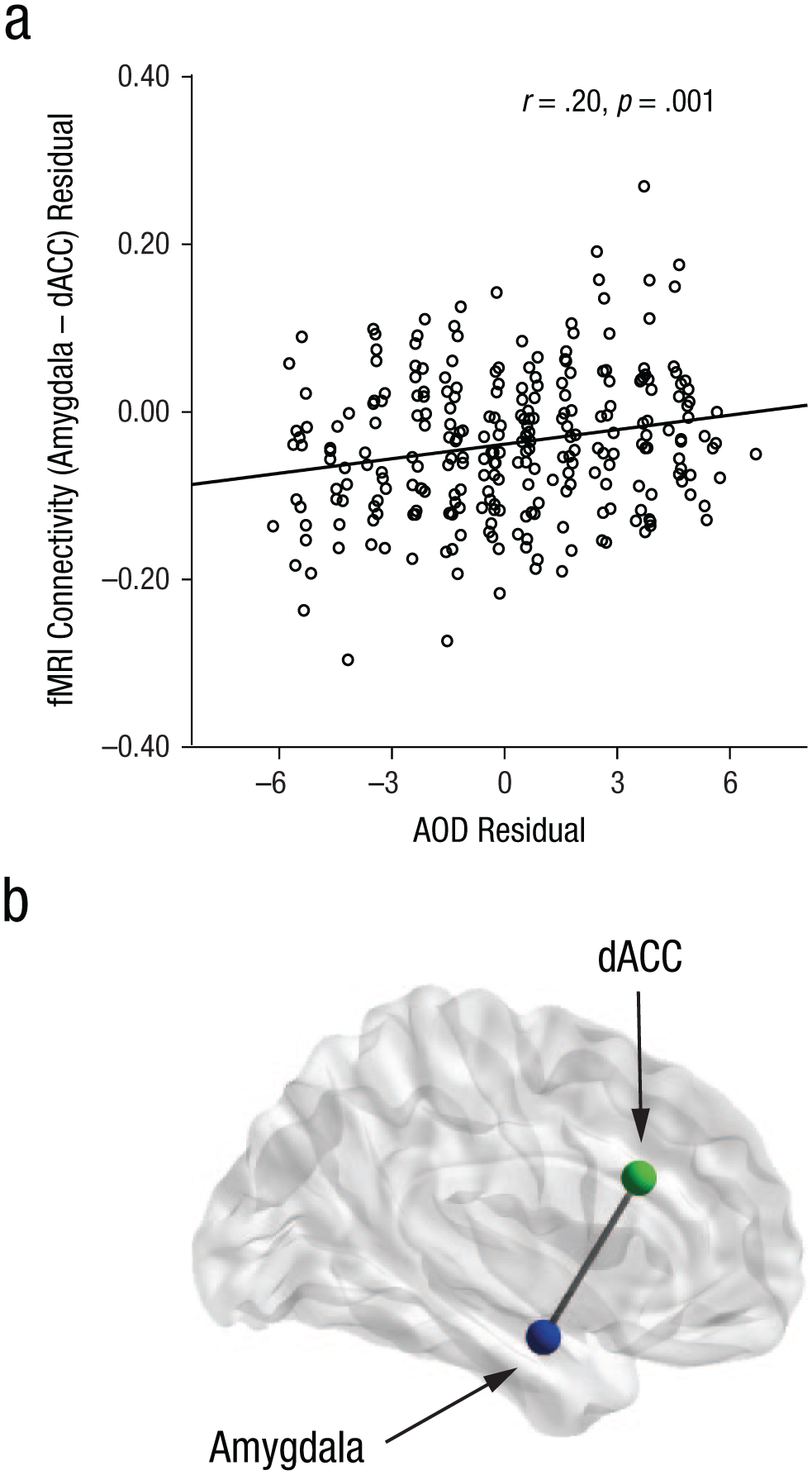
Results of the correlation analysis between functional MRI (fMRI) amygdala connectivity and score on the Prospective and Decision-Related Action Orientation Versus Hesitation (AOD) scale of the Action Control Scale (Kuhl, 1994). The scatterplot (a; with best-fitting regression line) illustrates the statistically significant relationship (N = 264, p = .001) between AOD scores and resting-state connectivity between the amygdala and the dorsal anterior cingulate cortex (dACC). The brain image (b) shows the location of the amygdala and dACC.
Discussion
The current study is the first to investigate the neural basis of interindividual differences in action control, which is the ability to orchestrate self- and emotion-control mechanisms. For this purpose, we examined the volume of single brain regions and utilized a resting-state fMRI protocol to analyze the resting-state functional connectivity within these brain regions. Our results showed that individuals with lower amygdala volume tend to be more action oriented in decision-related contexts than individuals with higher amygdala volume. Thus, people with higher amygdala volume appear to be more state oriented and therefore tend to hesitate to initiate an intention and tend to delay the beginning of tasks without any good reason (Blunt & Pychyl, 1998).
The amygdala is known to be a neuroanatomical hub for fear-motivated behavior (Davis & Whalen, 2001). During fear conditioning, the synaptic plasticity of the amygdala transforms sensory and somatosensory information of thalamic and cortical regions into a distinct fear memory (Davis & Whalen, 2001). This memory becomes especially important in decision-making situations, when the affective significance of a given behavior, stimulus, threat, or reward needs to be evaluated (Davis & Whalen, 2001). Numerous studies have investigated the role of the amygdala in risk evaluation and decision making (Bechara, Damasio, Damasio, & Lee, 1999; Hsu, Bhatt, Adolphs, Tranel, & Camerer, 2005). For instance, Bechara et al. (1999) showed that patients with selective amygdala lesions make disadvantageous decisions and appear not to draw any consequences from them to improve subsequent decision-making processes. Thus, the amygdala guides the selection of actions by selecting desirable behaviors (Davis & Whalen, 2001) and inhibiting actions that lead to potentially unfavorable outcomes (De Martino, Camerer, & Adolphs, 2010), on the basis of previous experiences.
Regarding action control, this could mean that individuals with a larger amygdala volume have learned from past mistakes and evaluate future actions and their possible consequences more extensively. This, in turn, might lead to greater concern and hesitation, as observed in individuals with low AOD scores (Blunt & Pychyl, 1998).
Interestingly, it seems as if the amygdala is especially important when the results of a particular behavior are uncertain (De Martino et al., 2010; Hsu et al., 2005; Whalen, 2007). This might explain why state-oriented individuals are especially vulnerable to the effects of uncontrollable outcomes and failures (Koole et al., 2005), whereas action-oriented individuals seem less affected by possible negative consequences (Quirin et al., 2011). This can also be seen in everyday situations. For instance, state-oriented (low AOD) individuals perform equally well as action-oriented (high AOD) individuals in sports as long as there is no direct threat or pressure on them. However, under pressure (e.g., when attempting to set a record), their performance drops significantly (Heckhausen & Strang, 1988). The reason for this might be that state-oriented individuals emphasize possible adverse consequences so excessively (e.g., not achieving a record) that action initiation is inhibited.
Regarding functional resting-state connectivity, we showed that interindividual differences in the functional connectivity of the amygdala and the dACC are associated with differences in decision-related action control (AOD). Specifically, higher connectivity between the amygdala and the dACC was associated with more action-oriented behavior, such as initiative taking.
Previous studies indicate that the dACC has reciprocal structural and functional connections with the amygdala (Feng, Feng, Chen, & Lei, 2014). The synergy between the dACC and the amygdala is assumed to play a significant role in purposive behavior (Feng et al., 2014) and self-control mechanisms. For instance, studies dealing with the neural basis of self-control failure support a model of top-down regulation of the amygdala by frontal and anterior cingulate structures (Ochsner, Bunge, Gross, & Gabrieli, 2002). In this context, the anterior cingulate cortex is usually associated with different self-regulatory processes (Heatherton, 2011), such as the detection of conflicts in information processing (Botvinick, Nystrom, Fissell, Carter, & Cohen, 1999), action-outcome evaluation, and reward-related selection of an action (Shenhav, Botvinick, & Cohen, 2013), as well as strategic adjustment of behavior and emotion (Botvinick, Cohen, & Carter, 2004). For instance, Hermann, Bieber, Keck, Vaitl, and Stark (2014) showed that the dACC is associated with expressive suppression, a major emotion regulation strategy. For this, the anterior cingulate cortex receives information from cortical and subcortical brain regions, such as the amygdala (Shenhav et al., 2013), and in turn regulates them by top-down projections (Ochsner et al., 2002). Unlike the findings mentioned above, our results indicate that interindividual differences in the often-reported functional connectivity between the amygdala and the dACC are associated with interindividual differences in the shaping of decision-related action orientation (AOD).
Thus, a permanent imbalance between both brain regions, in favor of the amygdala, correlates with a persistent inability to efficiently use psychological control processes, as it is seen in state orientation (low AOD). Our results go along with this assumption, as they showed that larger amygdala volume and less functional resting-state connectivity of the amygdala and dACC are associated with state orientation. One possible mechanism could be that the higher amygdala volume leads to behavior that is more strongly motivated by fear. This, in turn, might lead to an increased worry about negative consequences, which again provokes hesitancy and postponement. Furthermore, the low functional coupling between the amygdala and the dACC might result in insufficient top-down regulation of negative emotions, which could result in an inadequate selection and implementation of actions in state-oriented individuals (low AOD).
The results of our study are based on correlations between variables representing structural and functional brain features and the participants’ action control and therefore do not directly support causal inferences. Nevertheless, our results are consistent with the literature on the function of the amygdala and the dACC (Davis & Whalen, 2001; Feng et al., 2014). We found that a substantial amount of variance in AOD was explained by amygdala volume and network connectivity. However, there might be other factors that could contribute to the remaining variance in AOD. Even though the concurrent determination of both our neural and behavioral variables by a third factor cannot be entirely excluded, we demonstrated the specificity of our effects by comparison to neighboring brain areas, as well as considering various confounding variables (age, sex, and neuroticism) that might mediate or generate such a relationship.
In conclusion, our results are the first to show that variations in the anatomical architecture and network function of the amygdala are related to interindividual differences in action orientation in decision-related contexts. Because AOD is central to various aspects of life, the insight into its neurobiological basis is of high relevance. Thus, future studies should examine the hypotheses derived from this study and investigate whether the proposed neurobiological mechanisms are fixed or whether brain stimulation or specific training can lead to changes at both the behavioral and the neurobiological levels. Such findings would not only provide a causal explanation for the structure–function relationships we uncovered here but also offer a possibility to shift individuals with low AOD toward more desirable self-regulating behavior.
Supplemental Material
SchlueterSupplementalMaterial – Supplemental material for The Structural and Functional Signature of Action Control
Supplemental material, SchlueterSupplementalMaterial for The Structural and Functional Signature of Action Control by Caroline Schlüter, Christoph Fraenz, Marlies Pinnow, Patrick Friedrich, Onur Güntürkün and Erhan Genç in Psychological Science
Footnotes
Acknowledgements
We thank Katharina Berger for her support during the behavioral measurements. We also thank Burkhard Mädler of Philips Germany for scientific support with the MRI measurements as well as Tobias Otto for technical assistance.
Action Editor
Eddie Harmon-Jones served as action editor for this article.
Author Contributions
E. Genç conceived the project and supervised the experiments. C. Schlüter, M. Pinnow, O. Güntürkün, and E. Genç designed the project. C. Schlüter, C. Fraenz, and P. Friedrich collected the data. C. Schlüter, C. Fraenz, and E. Genç analyzed the data. C. Schlüter and E. Genç wrote the manuscript. All authors discussed the results and edited the manuscript. All the authors approved the final manuscript for submission.
Declaration of Conflicting Interests
The author(s) declared that there were no conflicts of interest with respect to the authorship or the publication of this article.
Funding
This work was supported by Deutsche Forschungsgemeinschaft Grants GU 227/16-1 and GE 2777/2-1, by SFB 1280 Project A03, and by Mercur Foundation Grant An-2015-0044.
Open Practices
Data and materials for this study have not been made publicly available, and the design and analysis plans were not preregistered.
Notes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.