
Abstract
Large-scale cooperation is a hallmark of our species and appears to be unique among primates. Yet the evolutionary mechanisms that drove the emergence of humanlike patterns of cooperation remain unclear. Studying the cognitive processes underlying cooperative behavior in apes, our closest living relatives, can help identify these mechanisms. Accordingly, we employed a novel test battery to assess the willingness of 40 chimpanzees to donate resources, instrumentally help others, and punish a culpable thief. We found that chimpanzees were faster to make prosocial than selfish choices and that more prosocial individuals made the fastest responses. Further, two measures of self-control did not predict variation in prosocial responding, and individual performance across cooperative tasks did not covary. These results show that chimpanzees and humans share key cognitive processes for cooperation, despite differences in the scope of their cooperative behaviors.
Humans are a hypercooperative species; we regularly engage in prosocial behavior, acting to benefit other people by giving our time, effort, and other valuable resources. The cognitive mechanisms that enable this human-specific pattern of cooperation are a subject of intense interest for both social scientists and biologists. Comparative studies of other species can provide important insights into the emergence of these mechanisms by identifying cognitive processes that are unique to humans and may therefore support human-specific forms of cooperation. In the current study, we assessed patterns of cooperative decision making in chimpanzees (Pan troglodytes) to probe the psychological processes that shape prosocial behavior and illuminate its evolutionary roots.
Chimpanzees are an important comparative model for human cooperation. They are our closest living relatives and exhibit a suite of complex cooperative behaviors in the wild. For example, chimpanzees hunt in groups, participate in joint attacks of neighboring communities, and form long-term alliances (Muller & Mitani, 2005). Experimental studies further show that chimpanzees help others in need (Melis & Warneken, 2016; Warneken, Hare, Melis, Hanus, & Tomasello, 2007; Warneken & Tomasello, 2009) and punish others for intentional harms (Jensen, Call, & Tomasello, 2007). However, chimpanzees do not routinely provide valued resources to others (Jensen, Hare, Call, & Tomasello, 2006; Silk et al., 2005), a key feature of human prosociality, or intervene on behalf of third parties (Riedl, Jensen, Call, & Tomasello, 2012), an important stabilizing mechanism for human cooperation. These major differences in the scope of chimpanzee cooperation are central to recent theories about human uniqueness (Hare, 2017; Melis & Warneken, 2016). Yet the cognitive mechanisms underlying these differences remain unclear.
Here, we leveraged recent developments in the study of human cooperation to probe the cognitive basis of cooperation in chimpanzees. First, we examined chimpanzees’ response latency and choice behavior during cooperative decisions to identify individual- and group-level prosocial preferences. Response times (RTs) have long been recognized for their utility in revealing the psychological processes underlying human behavior (for a review, see Kahneman, 2011). The prevailing view is that faster RTs reflect the engagement of more automatic processes, whereas slower responding reflects the engagement of deliberative cognitive processes, such as the active inhibition of a prepotent response (Aron, Robbins, & Poldrack, 2014; Verbruggen & Logan, 2008). Similarly, work in primate neuroscience shows that longer response latencies occur when switching from automatic to controlled behaviors (Hikosaka & Isoda, 2010). More recently, the use of RT data has been extended to identify the cognitive processes shaping human social decision making (Nosek, Hawkins, & Frazier, 2011), including cooperation.
In particular, recent proposals have used RT data to argue that humans exhibit extreme cooperation because prosocial responses are relatively automatic. Prosocial responding can be promoted by directly manipulating decision speed, and evidence shows that prosocial decisions are executed more rapidly than selfish ones in some contexts (Lotito, Migheli, & Ortona, 2013; Lotz, 2015; Rand, 2016; Rand, Greene, & Nowak, 2012; Rand et al., 2014; Yamagishi et al., 2017). Indeed, behavioral heuristics for automatic prosocial responding are proposed to have played a key role in the evolution of human-unique forms of cooperation (Bear, Kagan, & Rand, 2017; Bear & Rand, 2016; Rand et al., 2014). However, the interpretation of response latency data in such studies is a source of considerable debate. According to one view, faster prosocial responding, as well as increased prosocial responding under time constraints, stems from an intrinsic prosocial bias (Rand et al., 2012; Rand et al., 2014; Zaki & Mitchell, 2013). Another proposal holds that faster prosocial responding rather reflects reduced decision conflict: Individuals with stronger prosocial preferences experience less decision conflict when choosing the prosocial option and therefore make prosocial responses more quickly (Evans, Dillon, & Rand, 2015; Krajbich, Bartling, Hare, & Fehr, 2015). Thus, decision speed may reflect the strength of an individual’s preference. Importantly, neither of these views predicts that chimpanzees should exhibit fast prosocial responding in contexts in which they do not exhibit strong preferences for prosociality overall, so comparative work provides a strong test of these proposals.
Second, we measured the relationship between chimpanzees’ propensity to cooperate and their capacities for motoric and temporal self-control. Many theories of cooperation assume that individuals are inherently self-interested and that prosocial behavior requires the active inhibition of selfish impulses (Stevens & Hauser, 2004). In fact, some evidence shows that measures of self-control—such as temporal discounting—can predict an individual’s propensity to cooperate, and experimental manipulations of self-control can shift cooperative preferences in adults (Ainsworth, Baumeister, Ariely, & Vohs, 2014; Curry, Price, & Price, 2008; Espin, Brañas-Garza, Herrmann, & Gamella, 2012). However, other authors have suggested that costly decisions to help other people are facilitated by the high subjective value of providing benefits, rather than through the active inhibition of selfish actions. This alternate view is supported by neuroimaging and behavioral work (Buckholtz, 2015; Crockett, Kurth-Nelson, Siegel, Dayan, & Dolan, 2014; Zaki & Mitchell, 2011) and suggests that the evolutionary origins of human hypercooperation and self-control may be independent. Studies of chimpanzees can therefore test the evolutionary importance of self-control for human cooperation (Hare, 2017).
Finally, we characterized the latent structure of chimpanzee prosociality. Rates of altruistic donation and helping, but not punishment, are correlated in adult humans (Böckler, Tusche, & Singer, 2016; Peysakhovich, Nowak, & Rand, 2014). These findings suggest that costly behaviors that benefit other people access a single cognitive domain, whereas punishment or norm-based decisions index a distinct domain. Studying the structure of prosocial behavior in chimpanzees can therefore provide insights into the origins of these relationships across cooperation domains. If the structure of adult prosocial cognition facilitates human hypercooperation, chimpanzees should not exhibit this pattern. One possibility is that apes more closely resemble toddlers, who—like chimpanzees—engage in robust instrumental helping but do not yet share resources readily (Warneken & Tomasello, 2009). Indeed, the earliest emerging forms of helping and sharing are not correlated in young children (Dunfield & Kuhlmeier, 2013; Paulus, 2014), unlike in adults. To date, however, no work has directly compared independent measures of donation, helping, and punishment across the same individual chimpanzees to parse these relationships.
To answer these questions, we developed a task battery to examine patterns of prosocial decision making in 40 semi-free-ranging chimpanzees, the largest experimental sample of chimpanzee cooperation to date. The battery comprised measures of resource donation, helping, and punishment that have been extensively validated in prior work with chimpanzees (Jensen et al., 2007; Jensen et al., 2006; Silk et al., 2005; Warneken et al., 2007; Warneken & Tomasello, 2009) and that represent key components of human prosociality. We also included two self-control tasks to index variation in individuals’ abilities to delay gratification and inhibit prepotent motor responses, as well as a social-responsivity task to assess individual differences in chimpanzees’ interest in social stimuli. We used this battery to answer several questions about the cognitive mechanisms underlying chimpanzee cooperation. First, do chimpanzees exhibit a fast response bias, and do more cooperative individuals respond faster than selfish individuals? Second, do individuals with less self-control show lower rates of prosocial responding across the three social tasks? Finally, do distinct measures of chimpanzee prosociality covary? Answering these questions offers important insights into the evolution of humanlike cooperation.
Method
Subjects
We tested 40 wild-born, semi-free-ranging chimpanzees from Tchimpounga Chimpanzee Sanctuary in Republic of Congo (21 males, 19 females; age: M = 15 years, range = 6–25 years; for individual characteristics, see Table S1 in the Supplemental Material available online). This sample size, exceeding those used in prior experimental work on chimpanzee cooperation, allowed us to examine individual variation in responses. All chimpanzees were naive to the social decision-making tasks. All were socially housed, had ad libitum access to water, were never food deprived for testing, and were tested in familiar night dormitory buildings; most chimpanzees freely ranged in tropical rainforest during the day. Behavioral tests were approved by Harvard University’s Institution Animal Care and Use Committee (IACUC Protocol 11-12) and adhered to local laws (Republic of Congo’s Ministry of Scientific Research and Technological Innovation Permit No. 013/MRS/DGRST/DMAST).
Procedure
We analyzed six tasks assessing cooperation and self-control from a decision-making task battery. These included a resource-donation task, in which chimpanzees could provide food to a partner at no cost (Silk et al., 2005); an instrumental-helping task, in which they could give a partner an out-of-reach object (Warneken et al., 2007); and a punishment task, in which they could stop a thief from accessing a stolen resource (Jensen et al., 2007). Because chimpanzees engage in second-party (but not third-party) punishment in response to resource theft (Jensen et al., 2007; Riedl et al., 2012), we used a second-party-punishment task. The same chimpanzees also completed a temporal-discounting task assessing delay of gratification (Rosati & Hare, 2013), a go/no-go task indexing motor response inhibition (Verbruggen & Logan, 2008), and an independent measure of social interest (following Herrmann, Call, Hernàndez-Lloreda, Hare, & Tomasello, 2007).
We included only social tasks that have been validated in previous chimpanzee studies (Jensen et al., 2007; Jensen et al., 2006; Melis et al., 2011; Riedl et al., 2012; Silk et al., 2005; Warneken et al., 2007), focusing on critical test conditions for those tasks in order to examine individual variation in responses. Our goal was not to provide evidence for the existence of these behaviors in chimpanzees but, rather, to measure individual variation in responses. We therefore did not include all of the control conditions from prior work that have been used to validate these tasks, such as how chimpanzees respond to the donation task when no recipient is present (Silk et al., 2005) or whether they retrieve out-of-reach objects in the instrumental-helping task when the experimenter does not indicate their need (Warneken et al., 2007). Importantly, we did include initial exposure and comprehension-check trials, as relevant, to ensure that individuals understood the basic setups of the tasks. This approach allowed us to omit several time-consuming controls that have been extensively validated in prior work, enabling development of an appropriately rich battery of tasks. In doing so, we followed the same approach as in recent studies examining individual variation in primate cognitive abilities (Beran & Hopkins, 2018; Herrmann et al., 2007; Herrmann, Hernández-Lloreda, Call, Hare, & Tomasello, 2010).
Resource-donation task
In the resource-donation task, apes could choose between a prosocial option that provided food to both the subject and a human recipient and a selfish option that provided food only to the subject (see Video S1 in the Supplemental Material). The ape and recipient (Experimenter 1) sat across from each other at a table with a sliding top. A second experimenter (Experimenter 2) sat to one side to distribute food across trials. Each option consisted of two plates (one closer on the table to the subject, which the subject would receive, and the other closer to the recipient, which the recipient would receive). Both plates were baited with banana slices for the prosocial option, whereas only the plate closest to the chimpanzee was baited for the selfish option (following methods from Silk et al., 2005); side assignment of the two options was counterbalanced across trials.
Experimenter 1 reached toward the prosocial option to indicate a desire for the food, as prosocial helping is sensitive to such expressions of need (Melis & Warneken, 2016; Warneken et al., 2007). A grate served as a barrier between Experimenter 1 and the table to block direct access to the food. Apes first completed 20 exposure trials, with only one option available at a time, to ensure that each ape had experienced the outcome of both options. They then made choices on 20 test trials. Prior work has demonstrated that chimpanzees do not show a strong preference to donate food to others, exhibiting chance-level responses in social contexts and in nonsocial control conditions with no recipient (Jensen et al., 2006; Silk et al., 2005). We measured the chimpanzees’ choices for the prosocial option as well as their latency to respond by pointing or extruding their fingers toward one of the options. Latencies were coded frame by frame from when the table was pushed within reach to when the chimpanzee extended his or her finger to indicate a choice (see the Supplemental Material). One chimpanzee would not participate (would not choose over multiple trials) and was excluded from relevant analyses of this task.
Instrumental-helping task
In the instrumental-helping task, apes could decide whether to hand an out-of-reach item to a human experimenter who was actively reaching for it (see Video S2 in the Supplemental Material). On each trial, Experimenter 1 sat at the table, playing with a stick. After 5 s, Experimenter 2 approached, struggled with Experimenter 1 for the stick, and then tossed the stick into the chimpanzee’s room approximately 2 m away. Experimenter 1 then reached for the stick while making effortful vocalizations for 30 s, after which she switched to calling the chimpanzee’s name and alternating gaze between the ape and the stick (following methods from Warneken et al., 2007). Chimpanzees completed 10 test trials, each lasting up to a maximum of 60 s or until the chimpanzee gave the stick to Experimenter 1; they were never rewarded for helping. Chimpanzees robustly help in this situation when recipients signal their need, but they rarely pick up and return objects in control conditions in which recipients do not signal their goal (Melis et al., 2011; Warneken et al., 2007; Warneken & Tomasello, 2009). This suggests that the vast majority of object returns are prosocially motivated. We measured whether the chimpanzee handed over the stick as well as the latency to do so after Experimenter 1 started reaching for it.
Punishment task
In the punishment task, chimpanzees could punish a culpable thief by preventing the thief from profiting from the act (see Video S3 in the Supplemental Material). Chimpanzees initially had access to a food resource, which was then “stolen” by an experimenter who pulled it away and appeared to consume it. On each trial, Experimenter 1 sat across from the chimpanzee at a table with a collapsible wing (on the experimenter’s side). This wing was propped up by a false leg; a rope tied to the leg was threaded into the chimpanzee’s room so he or she could pull the leg out and collapse the table. Experimenter 2 first placed a large tray of food on the chimpanzee’s side. After 5 s, Experimenter 1 stole the tray and pretended to eat the food (following methods from Jensen et al., 2007). Chimpanzees first completed two initial nonsocial exposure trials in which the collapsible platform held a tray of rocks, so they could experience the physical effects of pulling the rope; this ensured that the apes understood the affordances of the task. Each chimpanzee then completed 10 test trials lasting a maximum of 60 s or until the ape pulled the rope to collapse the table. Chimpanzees are most likely to collapse a table so its contents fall out of reach of the “thief” specifically following intentional theft, compared with when an experimenter moves food over to the recipient (Jensen et al., 2007). We measured choices and latency to punish by collapsing the table.
Temporal-discounting task
In the temporal-discounting task, apes made decisions between a smaller option (one banana slice), available immediately, and a larger option (three banana slices), available after a 1-min delay (see Video S4 in the Supplemental Material). The experimenter sat across from the chimpanzee at the sliding table and placed the rewards on the table, with side assignment counterbalanced across trials (following methods in Rosati & Hare, 2013). Subjects first completed a number pretest with 4 trials (involving a different food type and no delays) to ensure that they could discriminate these quantities and preferred the larger amount when there were no time costs. They then completed 8 exposure trials (only one option available at a time) to introduce the rewards and delay contingencies. They finally completed 14 test trials, in which they made choices between the smaller, immediate reward and a larger, delayed reward. We measured the chimpanzees’ choices for the larger reward.
Go/no-go task
For the go/no-go task, we adapted a response-inhibition paradigm commonly used to measure motoric self-control in humans (Verbruggen & Logan, 2008). If chimpanzees responded to a “go” option (by touching it), they received a piece of food, but responses to a “no-go” option resulted in the experimenter throwing the food away (see Video S5 in the Supplemental Material). The options were two overturned plastic containers of different shapes and colors (counterbalanced across subjects). Apes first completed 20 exposure trials (in blocks of 5 trials per cue), so they could experience both situations. On each trial, the experimenter placed one option (either the go or no-go cue) and a piece of food on the table. The container was closer to the chimpanzee, and the food was out of reach. The experimenter then pushed the table forward so the chimpanzee could touch the container and experience the resultant outcome. Apes then completed learning trials, in which they made direct choices between the go and no-go options, and they had to choose the go cue option in 10 of the prior 12 trials to meet the criterion. Finally, they completed 40 test trials. The majority (80%) of test trials involved the go cue, with no-go cue trials randomly interspersed, to examine the chimpanzees’ ability to withhold the prepotent motor response they emitted on the majority of trials. Each trial lasted until the chimpanzee touched the object or up to 10 s. Chimpanzees rarely showed complete inhibition of responding during the 10-s trial period, as this testing context necessitated using a longer trial duration relative to typical human studies administered with a computer (Verbruggen & Logan, 2008). Our analyses therefore focused on the chimpanzee’s relative latency to respond to the go versus no-go options after the table was pushed forward. Latencies were coded frame by frame from when the table was pushed within reach to when the chimpanzee extended his or her finger to indicate a choice (see the Supplemental Material); chimpanzees who refrained for the entire trial duration were coded as having a 10-s latency. One chimpanzee would not participate (would not choose over multiple trials) and was excluded from relevant analyses of this task.
Social-responsivity task
In the social-responsivity task, chimpanzees could approach a series of social and nonsocial stimuli, allowing us to selectively assess social-approach motivation, while controlling for differences in general exploratory behavior (see Video S6 in the Supplemental Material). On each trial, Experimenter 2 centered the chimpanzee approximately 2 m from the table, and Experimenter 1 placed a novel item on the table (following the basic procedure in Herrmann et al., 2007). Each subject completed four trials with social stimuli (a person only, a person making an emotional vocalization, a photograph of an unfamiliar male chimpanzee, and a photograph of an unfamiliar female swelling) and four nonsocial trials (baseline with table only, food item, novel stationary object, and novel moving object). We measured the chimpanzees’ latency to approach the table during the 30-s trial.
Data coding and analysis approach
All tests were videotaped, and a coder blind to the hypotheses coded choices and response latencies for all sessions. Reliability for choices was compared with the live coding of the experimenter, with perfect agreement (κ = 1.0 in all cases). For latency coding, a second coder scored 20% of sessions for RTs, which were highly reliable (rp > .92 for all task latencies; for all reliability details, see the Supplemental Material). We analyzed data in the R programming environment (Version 3.4.1; R Core Team, 2017; for all analysis details, see the Supplemental Material). To analyze trial-by-trial choices and latencies, we implemented generalized linear mixed models (GLMMs) using the glmer function or linear mixed models using the lmer function, both from the lme4 package. Models included subject as a random intercept to account for repeated measurements within subjects. To examine the relationship between individual variation in mean choices and variation in mean latencies, we used linear regressions implemented with the lm function. Across analyses, we compared the fit of different models using likelihood-ratio tests (LRTs) and examined model contrasts using the lsmeans package. We report unstandardized model parameter estimates throughout. Finally, we also used the BayesMed package to interrogate null findings because evaluating support for null hypotheses is difficult within a frequentist statistical framework (for details, see the Supplemental Material).
Results
Prosocial choices are faster than selfish choices
We first examined the speed of prosocial versus self-interested decisions in the donation task, the design of which allowed measurement of response latency for both prosocial and selfish choices. Chimpanzees chose the prosocial option on a mean of 56.54% of the 20 test trials, 95% confidence interval (CI) = [52.30, 60.78], showing a slight preference for the prosocial option—one-sample, two-tailed t test on the proportion of prosocial choices: t(38) = 3.12, p = .0034, Cohen’s d = 0.50. Our large sample size likely permitted us to detect this modest effect. Choice frequency for the prosocial option ranged from 30% to 90% across individual chimpanzees (for individual chimpanzee responses, see Table S1). Overall, this aligns with prior work showing that chimpanzees do not have a strong overall preference for donating food to others, even at no personal cost (Jensen et al., 2006; Silk et al., 2005). However, it is noteworthy that despite the absence of a strong preference on the whole, we did observe interindividual variation in prosocial responding.
Our main question was whether prosocial responses were made more rapidly than selfish ones. Across test trials, chimpanzees chose the prosocial option after a mean of 0.30 s and the selfish option after a mean of 0.36 s—paired-samples t test: t(38) = −2.10, p = .04, 95% CI for the mean difference = [−0.13, −0.002], Cohen’s d = 0.34 (see Fig. 1a). To model choices between the prosocial and selfish options, we then fitted binomial GLMMs with a logit link function. A base model accounted for age, sex, and test-trial number (1–20), with subject as a random factor. A second model added response latency to test the hypothesis that prosocial decisions were made more rapidly than self-interested decisions. In fact, the inclusion of response latency improved model fit—LRT: χ2(1) = 7.52, p = .006. In particular, faster decision latencies significantly predicted more prosocial responding (estimate = −0.47, SE = 0.018, z = −2.60, p = .009; for all model parameters, see Table 1). Whereas chimpanzees showed a preference for the prosocial option at short latencies, longer latencies were associated with increasingly more frequent selfish choices (see Fig. 1b). On the whole, this analysis confirms that prosocial choices were made more rapidly than selfish ones in chimpanzees.
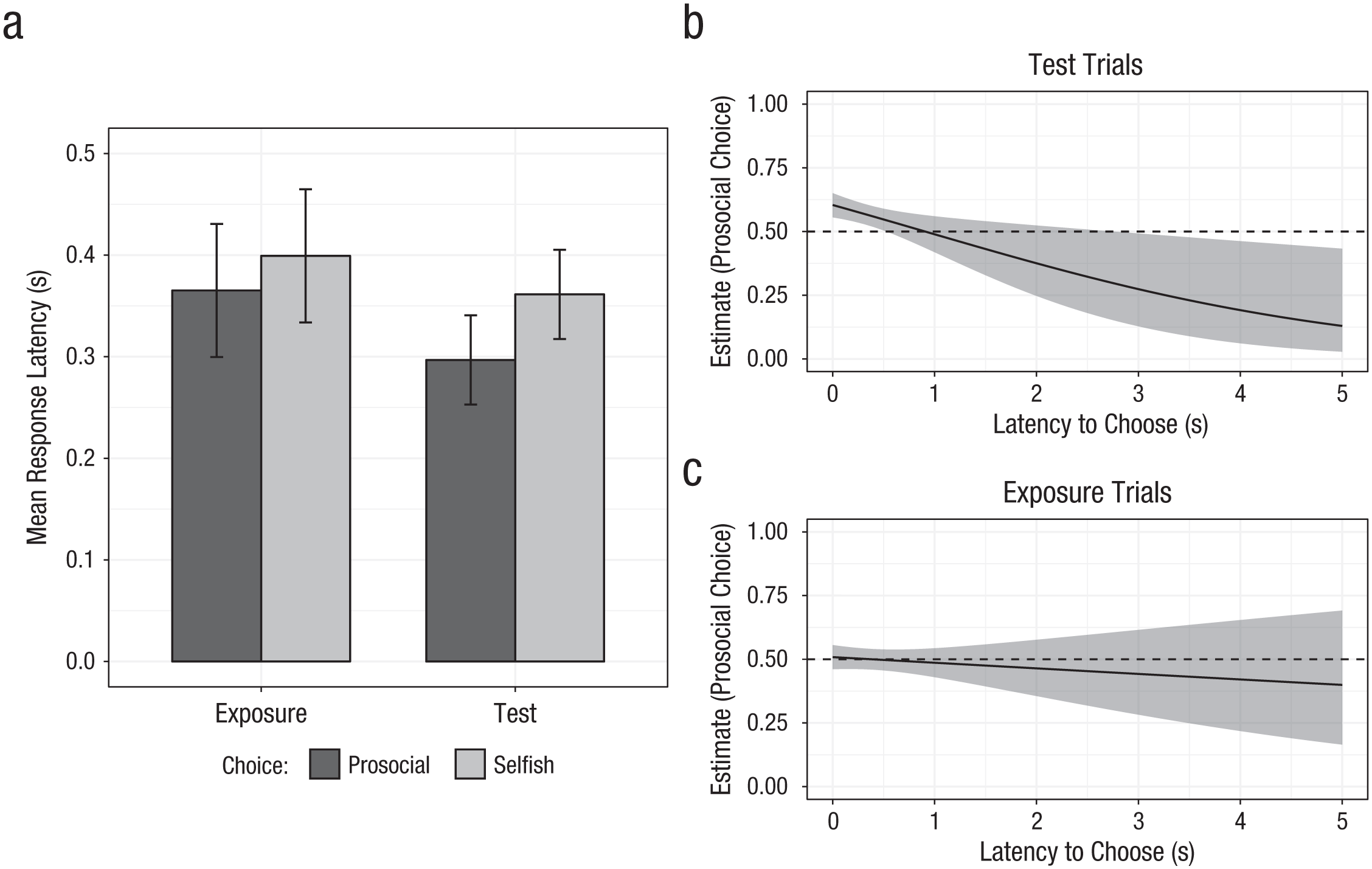
Choices and latencies in the resource-donation task. Chimpanzees chose between a prosocial option (providing food to the subject and an experimenter) and a selfish option (providing food only to the subject). The graph in (a) shows mean response latency for prosocial and selfish options in exposure trials (only one option available at a time) and test trials (active choices between options). Error bars indicate within-subjects 95% confidence intervals (CIs). Estimated values for prosocial (vs. selfish) choices are shown for decision latency (b) on test trials and (c) on exposure trials. Estimates were derived from generalized linear mixed models controlling for age, sex, and trial number. Dashed lines indicate chance preference between the prosocial and selfish options; error bands indicate 95% CIs for choice estimates.
Factors Influencing Propensity to Choose the Prosocial Option in Donation Test Trials
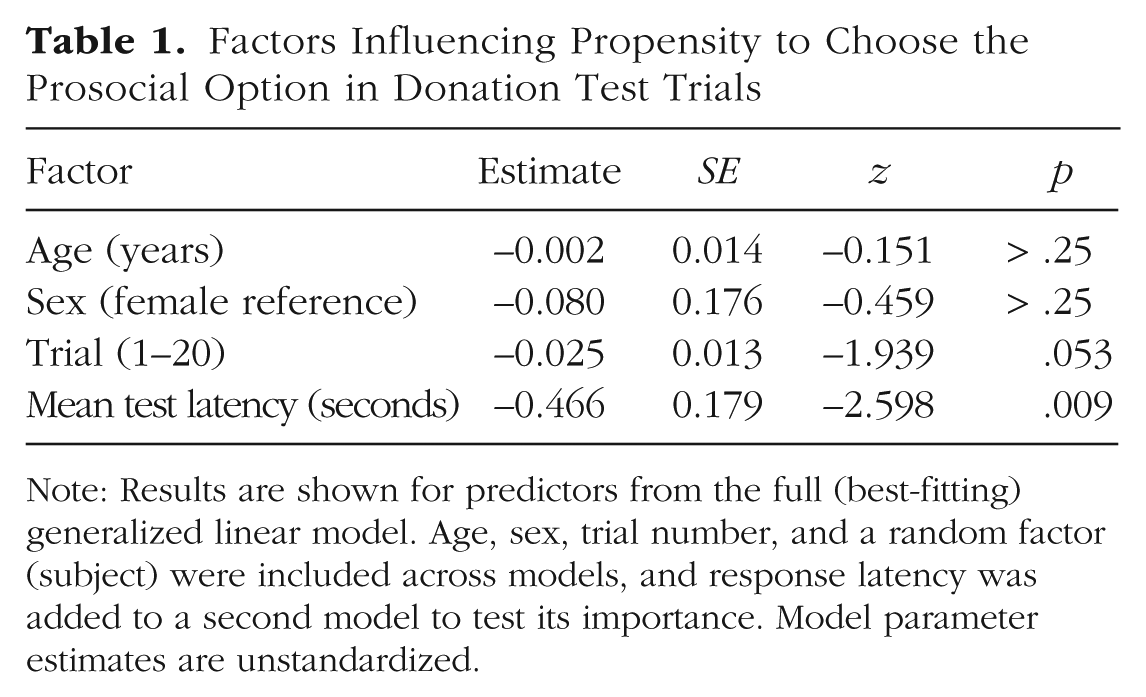
Note: Results are shown for predictors from the full (best-fitting) generalized linear model. Age, sex, trial number, and a random factor (subject) were included across models, and response latency was added to a second model to test its importance. Model parameter estimates are unstandardized.
As a further check, we examined responses on initial forced-choice exposure trials, in which only one option was available at a time. Here, apes responded to the prosocial option after a mean of 0.37 s and the selfish option after a mean of 0.40 s—paired-samples t test: t(38) = −0.74, p > .25, 95% CI for the mean difference = [−0.13, 0.06], Cohen’s d = 0.12. Using the same analysis procedure as for test trials, we found that response latency was not a significant predictor and did not improve model fit—LRT: χ2(1) = 0.58, p > .25 (for the parameters, see the Supplemental Material). That is, latency was not related to trial-by-trial choices in exposure trials (see Fig. 1c). Thus, the latency effect was observed only when chimpanzees were making an active choice on test trials, but not on exposure trials, ruling out the possibility that one option was more intrinsically salient. Additional checks confirmed that these findings were not driven by more extreme latency values, as results for both test and exposure trials held when trials with extreme latencies were removed (see the Supplemental Material). Thus, chimpanzees’ fast prosocial response bias was specific to test trials on which they had to make a consequential decision, despite not showing a robust preference to donate overall on such trials.
Work with adult humans suggests that response latencies can be driven by decision conflict, such that individuals with stronger prosocial preferences experience less decision conflict when choosing the prosocial option and therefore make prosocial responses more quickly (Evans et al., 2015; Krajbich et al., 2015). It is therefore possible that the present findings could reflect the fact that some individuals have stronger prosocial preferences and thus make prosocial responses more rapidly than selfish ones, rather than indicating a general prosocial response bias in chimpanzees. If this is the case, individuals who show an overall behavioral pattern of indifference to prosociality should not respond more rapidly when making prosocial choices. To test this, we examined chimpanzees’ trial-by-trial latencies as a function of their particular choice on a given trial, as well as their overall preference for prosociality. We first set up a base linear mixed model accounting for age, sex, and test-trial number (1–20), with subject as a random factor. A second model added choice (prosocial or selfish choice on that trial) to test whether prosocial decisions were made more rapidly than self-interested decisions on choice trials; this improved model fit—LRT: χ2(1) = 5.98, p = .015.
In a third model, we then added each individual’s prosocial preference (i.e., his or her mean frequency of prosocial choices across all test trials) to examine whether the relationship between prosocial decision making and response latencies depended on an individual’s overall strength of preference. This did not improve model fit—LRT: χ2(1) = 1.52, p = .22 (for the parameters, see the Supplemental Material). We then estimated predicted marginal means from this model and examined choice-latency relationships under the assumption of behavioral indifference (i.e., at a prosocial preference of 50% with choices equally distributed between the prosocial and selfish options). In fact, contrasts indicated significantly faster predicted latencies for the prosocial option than for the selfish option, even when individuals did not show an overall preference for prosocial choices—contrast estimate for selfish versus prosocial choice latencies = 0.07, 95% CI for the mean difference = [0.01, 0.13], t(739) = 2.31, p = .021. In contrast, the same analysis of choices on exposure trials indicated no relationship between choices and latencies (see the Supplemental Material). Taken together, this further supports the view that chimpanzee social decision making is characterized by a fast prosocial response bias, even in the absence of an overall preference for prosocial actions or outcomes.
More prosocial individuals make faster responses
We next examined whether cooperative individuals make faster responses than selfish individuals during social decision making. To test this, we examined chimpanzees’ choices to engage in instrumental helping and punishment, two domains in which chimpanzees do exhibit more robust preferences. In particular, prior work indicates that chimpanzees show more helping (Melis & Warneken, 2016; Melis et al., 2011; Warneken et al., 2007; Warneken & Tomasello, 2009) and punishment (Jensen et al., 2007; Riedl et al., 2012) in test conditions, compared with their low rates of responding in control conditions. This is in contrast to their relative indifference to prosociality in resource-donation contexts (Jensen et al., 2006; Silk et al., 2005).
In the instrumental-helping task, chimpanzees could choose to hand an out-of-reach object to a person reaching for it. Chimpanzees helped on an average of 5.83 out of 10 total test trials, 95% CI = [4.57, 7.08], range = 0% to 100% of trials across individuals. GLMMs of trial-by-trial choices to help indicated that there was no effect of age, sex, or changes in helping responses across trials (see the Supplemental Material). Our main question concerned the relationship between helping frequency and helping latency across individuals (note that, unlike in the donation task, selfishness here involved not producing a cooperative response at all, necessitating a different analytical approach). Decisions to offer help were made after an average of 20.72 s, 95% CI = [16.40, 25.04]. Examining the 33 subjects who helped at least one time, we found that mean helping rates and mean latency to help were significantly negatively correlated (rp = −.64, p < .001): Individuals who helped more often did so more quickly (see Fig. 2a). We found the same result when limiting analyses to helping acts that occurred only in the first half of sessions, before the actor actively recruited the chimpanzees’ help (n = 29, rp = −.66, p < .001).
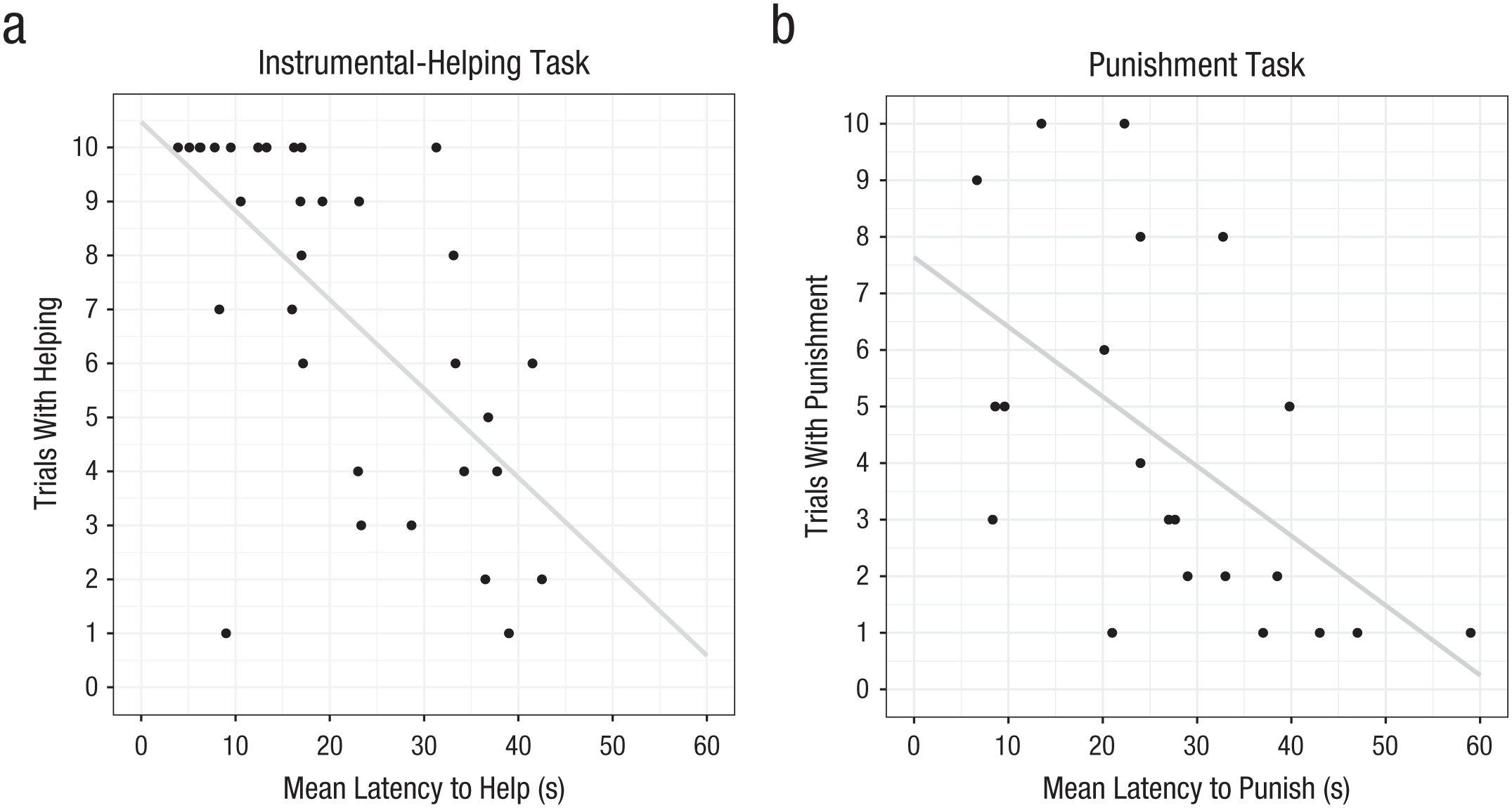
Scatterplots (with best-fitting regression lines) depicting associations between (a) each subject’s mean latency to help and the number of trials in which the subject helped and (b) each subject’s mean latency to punish and the number of trials in which the subject made a decision to punish.
We then used multiple linear regression to first fit a model of mean helping responses accounting for age and sex; in a second model, we included mean response latency to test whether faster individuals were more prosocial overall. This improved model fit—LRT: χ2(1) = 15.08, p < .001; adjusted r2 = .39—indicating that response latency is a significant predictor of helping behavior. In particular, individuals who were the quickest to help also helped the most frequently—estimate = −0.154, SE = 0.037, t(29) = −4.098, p < .001 (for the parameters, see Table 2). This supports the conclusion that more cooperative individuals tend to make prosocial choices faster than do more selfish individuals.
Factors Influencing Mean Rate of Helping
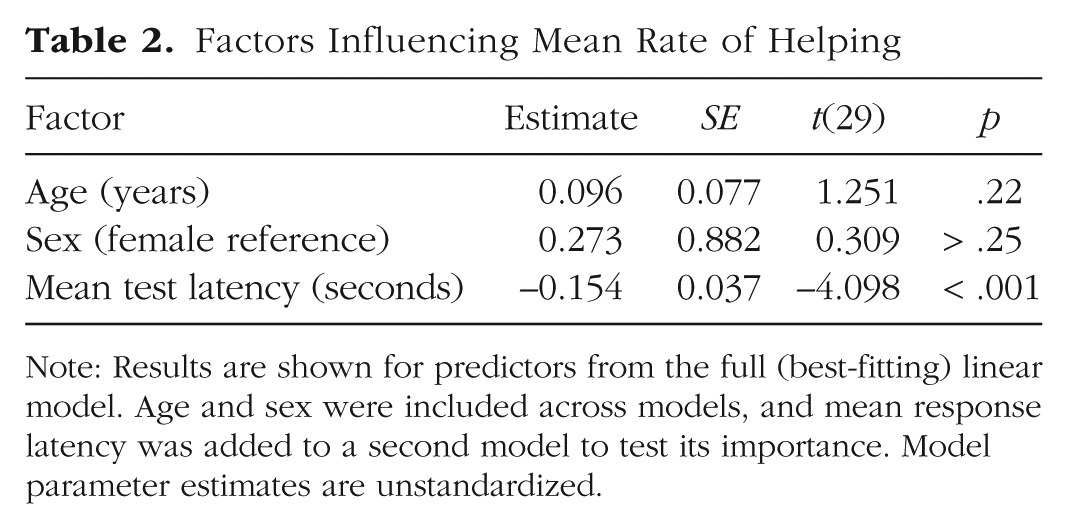
Note: Results are shown for predictors from the full (best-fitting) linear model. Age and sex were included across models, and mean response latency was added to a second model to test its importance. Model parameter estimates are unstandardized.
We used the same approach to examine decisions in the punishment task, which had the same basic structure as the instrumental-helping task. Chimpanzees could punish a thief who had stolen their food by collapsing the table holding the food. They collapsed the table on an average of 2.25 out of 10 test trials, 95% CI = [1.25, 3.25], range = 0% to 100% across individuals. GLMMs examining trial-by-trial choices to punish revealed that there was no effect of age, sex, or changes over trials in punishment responses (see the Supplemental Material). Our main question again concerned the relationship between punishment frequency and punishment latency across individuals. Chimpanzees’ mean latency to collapse the table was 27.23 s, 95% CI = [20.91, 33.55], after the theft event. Examining the 21 subjects who punished at least one time, we found that overall mean punishment rates and mean latency to punish were correlated (rp = −.55, p = .01): Individuals who punished more often did so more quickly (see Fig. 2b). We then fitted a linear model accounting for age, sex, and mean exposure response latency (to account for each individual’s speed to collapse the table in nonsocial exposure trials). In a second model, we then added a mean test-response-latency predictor, which improved model fit—LRT: χ2(1) = 8.58, p = .003; adjusted r2 = .30. Indeed, test-response latency significantly predicted responses—estimate = −0.155, SE = 0.054, t(16) = −2.841, p = .012—but exposure latency did not (for the parameters, see Table 3). Thus, individuals who engaged in more punishment were also the quickest to punish, even when analyses controlled for individual chimpanzees’ latency to respond on nonsocial exposure trials.
Factors Influencing Mean Rate of Punishment
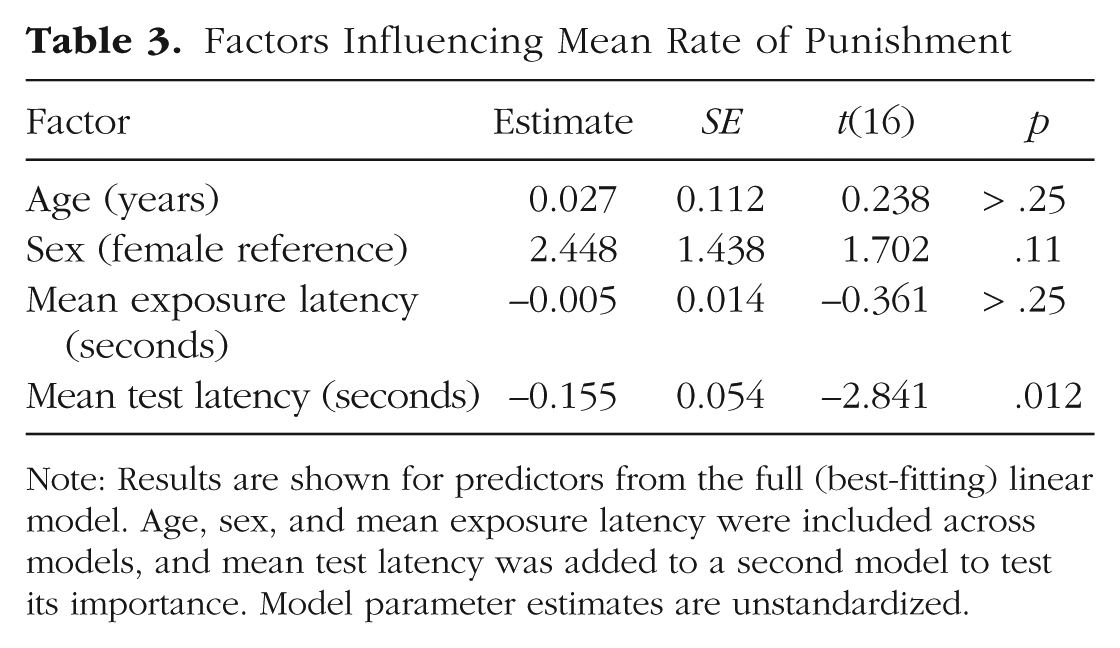
Note: Results are shown for predictors from the full (best-fitting) linear model. Age, sex, and mean exposure latency were included across models, and mean test latency was added to a second model to test its importance. Model parameter estimates are unstandardized.
Self-control does not predict prosocial responding in chimpanzees
We next examined whether individual variation in self-control predicted chimpanzees’ cooperative tendencies. To do so, we first assessed their performance on the temporal-discounting task measuring willingness to delay gratification. In the number pretest without delays, chimpanzees chose the larger reward on 80.00% of trials, a rate above chance, t(39) = 9.22, p < .001, 95% CI = [73.42, 86.58], Cohen’s d = 1.46. However, they chose the larger reward on only 65.18% of test trials in which they had to wait for it, which was also above chance, t(39) = 5.68, p < .001, 95% CI = [59.77, 70.58], Cohen’s d = 0.90. This was significantly less often than on number trials, t(39) = −3.95, p < .001, 95% CI of the mean difference = [−22.40, −7.24], Cohen’s d = 0.62 (see Fig. 3a). GLMMs of trial-by-trial choices for the larger reward confirmed that including trial type (number vs. test) improved model fit—LRT: χ2(1) = 6.69, p < .01, but there were no effects of age, sex, or changes in preferences across trials (see the Supplemental Material). Thus, chimpanzees were significantly less likely to choose the larger reward in test trials (i.e., when they had to wait for it) compared with the number pretest trials, confirming their sensitivity to delay costs in this task.
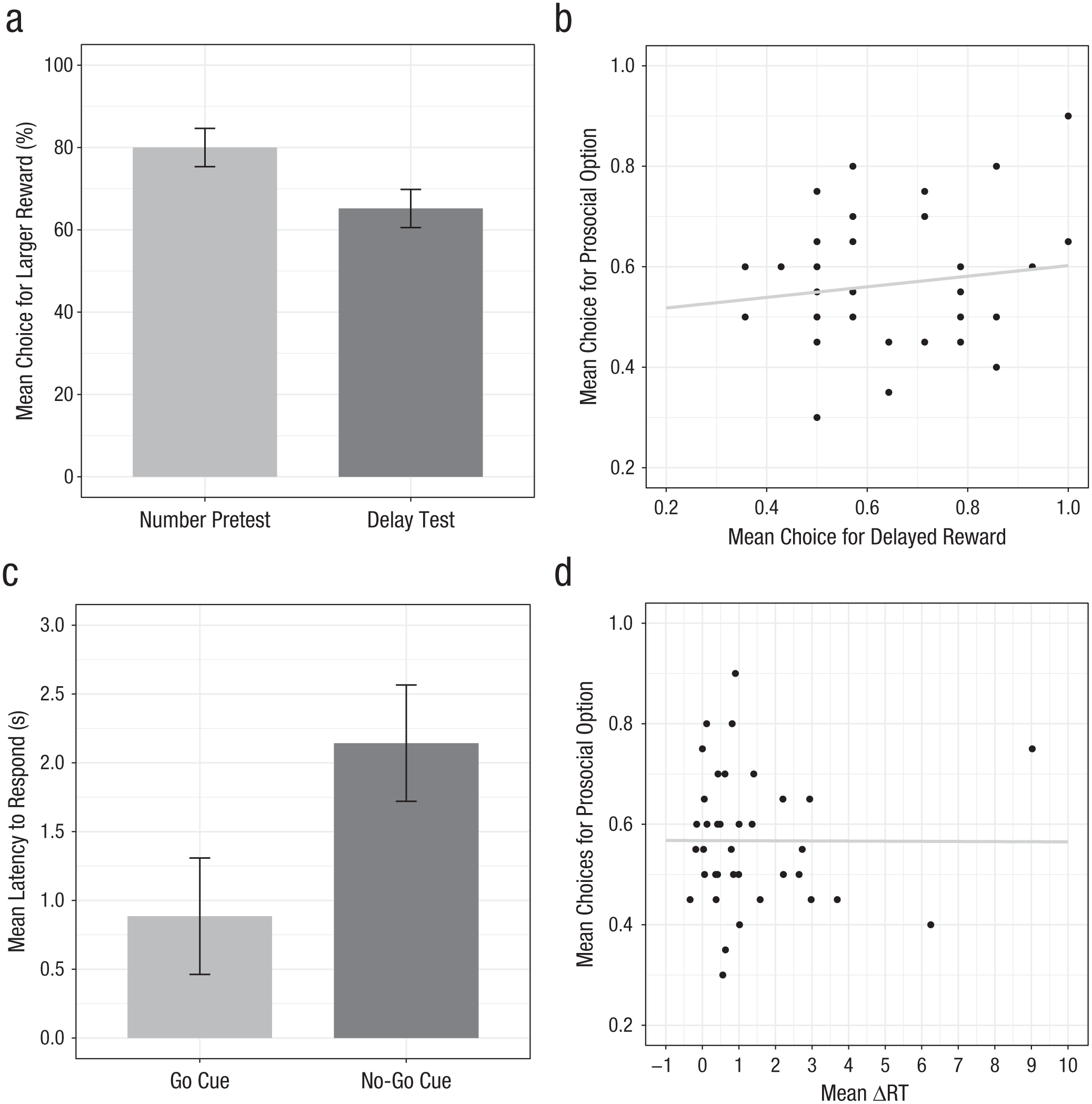
Relationships between temporal and motoric inhibitory control and prosocial behavior. The mean percentage of choices for the larger reward in the temporal-discounting task (a) is shown for both the number pretest (without temporal delays) and the delay test; error bars indicate within-subjects 95% confidence intervals (CIs). The scatterplot (with best-fitting regression line) in (b) shows the relation between mean proportion of choices for the larger, delayed rewards in the temporal-discounting task and mean proportion of prosocial donation frequency. Mean latency to respond (c) is shown for both the no-go and go cues in the go/no-go task; error bars indicate within-subjects 95% CIs. The scatterplot (with best-fitting regression line) in (d) shows the relation between mean change in response time (ΔRT) in the go/no-go task and mean proportion of prosocial donation frequency.
Our main question was whether individual variation in willingness to wait for the delayed reward was related to cooperative tendencies. As frequentist models are unable to provide positive evidence for a null hypothesis (H0), we further used Bayes factor (BF) analysis to calculate the strength of evidence for the H0 (no association between self-control and social-task performance; for details, see the Supplemental Material). Individuals varied widely in their ability to delay gratification (36%–100% of trials across individuals), but there was no relationship between capacity to delay gratification and individual differences in the frequency of cooperative behavior in the resource-donation task (n = 39, rp = .14, p > .25, BF01 = 5.67), the instrumental-helping task (n = 40, rp = −.19, p = .25, n.s., BF01 = 4.15), or the punishment task (n = 40, rp = .14, p > .25, n.s., BF01 = 5.55). BF01 values for these correlations indicated moderate support for H0 (e.g., the donation data were 5.67 times more likely under the null hypothesis than under the alternative that there is a relationship between prosocial donation and temporal discounting). Finally, regression models accounting for subject’s sex, age, and performance in the number pretest were also not improved by including the cooperation measures as predictors (for details, see the Supplemental Material). Thus, we found no evidence that individual variation in intertemporal choices was related to cooperative tendencies.
We found similar results with our measure of motoric inhibitory control. The go/no-go task assessed chimpanzees’ ability to withhold a prepotent motor response when it was not appropriate. All chimpanzees demonstrated comprehension of the basic contingencies of the task, passing the criterion for preferring the go cue over the no-go cue after an average of 17.67 learning trials (67% passed in their first 12 trials). In subsequent test trials, mean RT for responses to no-go cues was 2.14 s, compared with 0.89 s for trials with go cues, t(38) = 4.26, p < .001, 95% CI for the mean difference = [0.65, 1.86], Cohen’s d = 0.68 (see Fig. 3c). GLMM analyses of trial-by-trial response latencies confirmed that including cue type (go cue vs. no-go cue) improved model fit—LRT: χ2(1) = 204.1, p < .001. That is, chimpanzees were slower to respond to the no-go cues, as expected for a task tapping into motor response inhibition. There were no effects of age or sex on responses (see the Supplemental Material).
As our main question was whether go/no-go task performance was related to cooperative tendencies, we used each individual’s average difference in RT between no-go trials and go trials (ΔRT) as our individual-difference measure. Here, larger (positive) ΔRTs indicated greater inhibitory control over prepotent motor responses. Mean ΔRT was 1.26, 95% CI = [0.66, 1.86], with individual chimpanzees’ difference scores ranging from −0.74 s to 9.02 s. However, individual variation in ΔRT was not related to frequency of prosocial behavior in the donation task (n = 38, rp = −.003, p > .25, BF01 = 7.92), the instrumental-helping task (n = 39, rp = .19, p = .25, BF01 = 4.18), or the punishment task (n = 39, rp = −.10, p > .25, n.s., BF01 = 5.77). As with discounting, these BFs provide moderate support for the null hypothesis that there is no relationship between self-control and cooperation. Additional checks accounting for the nonnormal distribution of ΔRT data, as well as regression models accounting for sex and age, found similar results (see the Supplemental Material).
Relationship between cooperative measures
Our final set of analyses examined the relations between the chimpanzees’ performance across the three cooperation tasks. We examined the structure of individual variation in chimpanzee cooperation using intertask correlations, similar to prior work in adults (Böckler et al., 2016; Peysakhovich et al., 2014) and children (Dunfield & Kuhlmeier, 2013; Paulus, 2014). In fact, we found that chimpanzee performance on one task did not correlate with performance on any of the other tasks when we compared donation with helping (n = 39, rp = .05, p > .25, BF01 = 7.73), donation with punishment (n = 39, rp = .19, p = .24, BF01 = 4.08), or helping with punishment (n = 40, rp = −.03, p > .25, BF01 = 7.96). These BF values provide moderate support for the null hypothesis that there is no relationship between chimpanzees’ performance across these cooperative tasks.
Our social-responsivity task further supports the conclusion that these social decision-making tasks index distinct components of cooperation in chimpanzees. This task provided an independent measure of social motivation, while controlling for differences in general approach or exploratory behavior toward novel situations. We found that individuals who were faster to approach social stimuli exhibited more instrumental helping but less frequent resource donation; punishment was unrelated (see the Supplemental Material). That is, our three indices of chimpanzee prosocial decision making were uncorrelated, and each differed in its relationship to an independent measure of social motivation, supporting the conclusion that donation, helping, and punishment represent distinct facets of chimpanzee cooperation. This result stands in contrast to work in adult humans, which shows that rates of altruistic donation and helping tend to be correlated (Böckler et al., 2016; Peysakhovich et al., 2014), but is more similar to findings from young children, in which early forms of prosociality do not share variance (Dunfield & Kuhlmeier, 2013; Paulus, 2014).
Discussion
We developed a novel decision-making battery comprising measures of prosociality and self-control and used it to examine the cognitive mechanisms underlying cooperation in chimpanzees. We found that chimpanzees make prosocial donation choices more rapidly than self-interested ones and that more cooperative individuals respond more rapidly in both helping and punishment contexts. Second, we found that chimpanzees’ prosocial behavior is not related to individual variation in either temporal or motoric self-control, suggesting a dissociation of self-control capacity and cooperation in chimpanzees in these contexts. Finally, we found that chimpanzees’ responses across the donation, helping, and punishment tasks were unrelated but that each task exhibited different relationships with an independent measure of social motivation. A key feature of our approach was a focus on individual variation in cooperative performance, using tasks that have been previously validated with experimental controls (Jensen et al., 2007; Jensen et al., 2006; Riedl et al., 2012; Silk et al., 2005; Warneken et al., 2007). The performance of these chimpanzees at the group level broadly conformed with the results of prior studies, providing additional support for the validity of this approach.
Our finding that chimpanzees make prosocial decisions faster than selfish ones has important implications for understanding the evolution of human-unique forms of cooperation. A recent model explicitly linked a bias for fast cooperative responding to the emergence of robust human cooperation (Bear et al., 2017; Bear & Rand, 2016; Rand et al., 2014). In particular, a fast prosocial bias is proposed to emerge in evolution only when cooperation is a highly successful strategy; self-interested responses otherwise become the default. Whereas earlier work in humans inferred automatic or intuitive modes of cognition from RT data (Rand et al., 2012; Rand et al., 2014), more recent work suggests that inferences about modes of cognition cannot be made from endogenous RT measurements alone. In particular, faster cooperative decisions may also occur when people feel less decision conflict (Evans et al., 2015; Krajbich et al., 2015). Yet our results conflict with both views. Chimpanzees exhibited faster prosocial responding even though they did not exhibit a strong preference for prosocial donation (as would be suggested by the decision-conflict view) and are not a hypercooperative species overall. Furthermore, individual variation in RTs predicted preferences to help and punish to a similar degree, even though chimpanzees, on the whole, engaged in more helping than punishment. As this work used endogenous RTs, future studies should employ external time-pressure manipulations to definitively test the role of intuitive versus deliberative processes in ape cooperation. Nevertheless, most interpretations of RT data hinge on the idea that faster responses are more automatic or prepotent, whereas slower responses require more effortful control (Aron et al., 2014; Hikosaka & Isoda, 2010; Kahneman, 2011; Nosek et al., 2011; Verbruggen & Logan, 2008).
We also found that individual variation in temporal and motoric self-control did not predict chimpanzee cooperation. This is especially striking given recent work showing that delay of gratification does predict individual variation in chimpanzee cognition in other cognitive domains (Beran & Hopkins, 2018). However, no prior work examined the relationship between self-control and cooperation in chimpanzees. Recent work in humans suggests that costly decisions to help other people can be linked to intrinsic valuation of prosocial outcomes (Crockett et al., 2014; Zaki & Mitchell, 2011). The fact that chimpanzee prosociality was unrelated to self-control offers some support for this value-based account in chimpanzees as well. Our BF analysis also supports a lack of relationship, although this inference is based on a null effect and should be interpreted with caution until replicated in larger populations.
Finally, our results reveal some important differences in the latent cognitive structures of chimpanzee and human cooperation. In adults, rates of altruistic donation and helping are correlated across individuals, whereas punishment is not (Böckler et al., 2016; Peysakhovich et al., 2014). This implies that the former two tasks index a latent cognitive process that is distinct from that accessed by punishment. In contrast, the earliest emerging forms of cooperation are all uncorrelated in young children (Dunfield & Kuhlmeier, 2013; Paulus, 2014). We found that chimpanzees more closely resemble children, with uncorrelated responses across the donation, helping, and punishment tasks. This also aligns with the finding that both chimpanzees and young children exhibit more robust instrumental helping than resource sharing (Warneken & Tomasello, 2009). The fact that each task exhibited a different pattern of association with an independent measure of social motivation is further evidence that the lack of intertask correlation is meaningful rather than artifactual. This suggests that the latent structure of adult human cooperation may be different from that seen in other species and, further, that human developmental processes may build on cognitive structures that more closely resemble our ape relatives.
Ultimately, our results show that chimpanzee cooperation involves several cognitive mechanisms that parallel those seen in humans, despite chimpanzees being markedly less cooperative than humans overall. In particular, chimpanzees exhibit a bias for fast cooperation that is independent from both intertemporal and motor self-control. This has important implications for the evolution of human ultrasociality, as our results suggest that the presence of fast cooperative responding cannot account for the extreme nature of human prosociality and may occur even in the absence of an overall preference for prosociality. An important aim for future research will therefore be to investigate whether these signatures of human cooperation are even more widely shared across other primate species who vary in their tendency to share resources, such as in more tolerant bonobos (Hare, 2017). Comparative data examining the relationships between value-related decisional biases, self-control, and cooperative behaviors across species will be critical for understanding the evolution of human hypercooperation.
Supplemental Material
RosatiOpenPracticesDisclosure – Supplemental material for Chimpanzee Cooperation Is Fast and Independent From Self-Control
Supplemental material, RosatiOpenPracticesDisclosure for Chimpanzee Cooperation Is Fast and Independent From Self-Control by Alexandra G. Rosati, Lauren DiNicola and Joshua W. Buckholtz in Psychological Science
Supplemental Material
RosatiSupplementalMaterial – Supplemental material for Chimpanzee Cooperation Is Fast and Independent From Self-Control
Supplemental material, RosatiSupplementalMaterial for Chimpanzee Cooperation Is Fast and Independent From Self-Control by Alexandra G. Rosati, Lauren DiNicola and Joshua W. Buckholtz in Psychological Science
Footnotes
Acknowledgements
We thank Randy Buckner, Felix Warneken, and Richard Wrangham for comments; Steve Worthington and Simo Goshev at Harvard University’s Institute for Quantitative Social Science for statistical advice; and Ronan Donovan for chimpanzee photo stimuli. At Tchimpounga Chimpanzee Sanctuary, we thank Rebeca Atencia, Debby Cox, the chimpanzee caretakers, the Jane Goodall Institute USA, and the Congolese Ministry of Research.
Action Editor
Steven W. Gangestad served as action editor for this article.
Author Contributions
A. G. Rosati and J. W. Buckholtz designed the study. A. G. Rosati collected the data, and A. G. Rosati and L. M. DiNicola coded the data. A. G. Rosati analyzed the data with input from J. W. Buckholtz. All the authors wrote the manuscript and approved the final manuscript for submission.
Declaration of Conflicting Interests
The author(s) declared that there were no conflicts of interest with respect to the authorship or the publication of this article.
Funding
This research was supported by a Leakey Foundation grant to A. G. Rosati. J. W. Buckholtz was supported by the Sloan Foundation, the Brain and Behavior Research Foundation, the Harvard Foundations of Human Behavior Initiative, and the Massachusetts General Hospital Center for Law, Brain and Behavior.
Open Practices
All data and materials have been made publicly available via the Dryad repository and can be accessed at https://doi.org/10.5061/dryad.cn20jv8. The complete Open Practices Disclosure for this article can be found at https://journals-sagepub-com.web.bisu.edu.cn/doi/suppl/10.1177/0956797618800042. This article has received the badges for Open Data and Open Materials. More information about the Open Practices badges can be found at ![]() .
.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.