
Abstract
Deficient self-control leads to shortsighted decisions and incurs severe personal and societal costs. Although neuroimaging has advanced our understanding of neural mechanisms underlying self-control, the ecological validity of laboratory tasks used to assess self-control remains largely unknown. To increase ecological validity and to test a specific hypothesis about the mechanisms underlying real-life self-control, we combined functional MRI during value-based decision-making with smartphone-based assessment of real-life self-control in a large community sample (N = 194). Results showed that an increased propensity to make shortsighted decisions and commit self-control failures, both in the laboratory task as well as during real-life conflicts, was associated with a reduced modulation of neural value signals in the ventromedial prefrontal cortex in response to anticipated long-term consequences. These results constitute the first evidence that neural mechanisms mediating anticipations of future consequences not only account for self-control in laboratory tasks but also predict real-life self-control, thereby bridging the gap between laboratory research and real-life behavior.
Keywords
Humans frequently face conflicts between social norms, moral values, or long-term goals (e.g., avoiding lung cancer, staying slim) and competing desires for tempting immediate rewards with often adverse long-term consequences (e.g., to smoke, to eat tasty but unhealthy food; Hofmann, Baumeister, Förster, & Vohs, 2012; Hofmann, Vohs, & Baumeister, 2012; Wolff et al., 2016). Self-control denotes the ability to resist such temptations and align behavior with superordinate goals (Baumeister, Tice, & Vohs, 2018; Goschke, 2014; Inzlicht, Legault, & Teper, 2014). Deficient self-control is associated with a wide range of harmful behaviors, such as substance abuse, overeating, problem gambling, and impulsive aggression (Bühringer, Wittchen, Gottlebe, Kufeld, & Goschke, 2008; Goschke, 2014; Heatherton & Wagner, 2011), which incur immense societal costs due to poor health, disability, and premature death (Schroeder, 2007; Wittchen et al., 2011).
In recent years, behavioral and neuroimaging studies of decision-making and cognitive control have substantially advanced our understanding of the psychological and neural processes underlying self-control (Inzlicht et al., 2014; Kotabe & Hofmann, 2015; Turner et al., 2018). However, concerns have recently been raised that task-based measures of self-control are not meaningfully associated with self-reported self-control, lack reliability, and have poor predictive power (Eisenberg et al., 2019; Enkavi et al., 2019; Saunders, Milyavskaya, Etz, Randles, & Inzlicht, 2018). Thus, a major challenge in self-control research is to assess the ecological validity of the highly artificial laboratory tasks that are typically used to assess self-control during value-based decisions. This is a particular concern regarding neuroimaging studies, which impose tight constraints on the paradigms that can be used in a scanner environment (e.g., asking participants to repeatedly make a large number of fictitious choices).
To address the trade-off between experimental control and ecological validity, a number of researchers have recently combined laboratory tasks and neuroimaging methods with assessments of real-life behavior using smartphone-based momentary ecological assessments (i.e. experience sampling). Although this brain-as-predictor approach (Berkman & Falk, 2013) is correlational in nature, the few recent studies that have used it yielded promising results, indicating, for instance, that individual differences in real-life self-control failures are associated with activity in brain areas involved in cognitive control and response inhibition (Berkman, Falk, & Lieberman, 2011; Krönke et al., 2018; Lopez, Hofmann, Wagner, Kelley, & Heatherton, 2014).
Here, we employed the brain-as-predictor approach to test a theoretical model, according to which self-control rests critically on the degree to which neural value representations are modulated by anticipated future outcomes of decisions (Berkman, Hutcherson, Livingston, Kahn, & Inzlicht, 2017; Goschke, 2014; Hare, Camerer, & Rangel, 2009). Thus, in contrast to the predictions of dual-systems theories, which postulate that a “reflective” system encoding long-term goals suppresses an “impulsive” system encoding short-term value (Berkman et al., 2017; Heatherton & Wagner, 2011; Hofmann, Friese, & Strack, 2009), our hypothesis was that choices between conflicting options result from a dynamic integration process whereby the subjective value of each alternative is calculated by integrating various short- and long-term gains (e.g., the immediate rewarding effects of alcohol consumption) and costs (e.g., hangover on the next day and negative health outcomes). The resulting integrated value signal reflects the individual’s preferences at the moment of decision and will determine the enactment of the most valued option. We thus assumed that individual differences in self-control depend on the relative weight with which long-term consequences enter this value-integration process, rather than on the suppression of an impulsive system encoding short-term value. Although our hypothesis did not exclude the possibility that self-control may also be supported by reducing the relative weight of short-term outcomes in the value-integration process, we assumed that especially highly tempting immediate rewards would exert a modulating influence on the value-integration process in most individuals but that self-controlled individuals would more strongly anticipate long-term consequences during decision conflicts, thereby enhancing the weight of those consequences on the integrated value signal (Kruschwitz et al., 2018; Peters & Büchel, 2010a).
Consistent with this hypothesis, neuroimaging evidence indicates that subjective value across a wide range of stimulus domains correlates with activation in the ventromedial prefrontal cortex (vmPFC; Bartra, McGuire, & Kable, 2013; Chib, Rangel, Shimojo, & O’Doherty, 2009; Kable & Glimcher, 2007; Levy & Glimcher, 2012; Lin, Horner, Bisby, & Burgess, 2015; Peters & Büchel, 2010b; Plassmann, O’Doherty, & Rangel, 2010; Rangel & Hare, 2010) and that the vmPFC is involved in the processing of both real and imagined rewards (Bray, Shimojo, & O’Doherty, 2010; Lin et al., 2015). Moreover, self-controlled choices in laboratory tasks depend on how strongly this value signal is modulated by long-term consequences (e.g., the healthiness of food items) but not by short-term consequences (Hare et al., 2009).
Thus, we derived two predictions concerning the pattern of neural activation and its relation to real-life self-control. First, looking beyond previous evidence for a role of the vmPFC in encoding the subjective value of primary, monetary, or social reinforcers (Bartra et al., 2013), we predicted that brain activity in the vmPFC should be parametrically associated with the subjective value of daily behaviors from a wide range of thematic and motivational domains. Second, and critically, we predicted that individuals in whom value signals in the vmPFC are more strongly modulated by anticipated long-term consequences of behavioral options should show a lower propensity to commit self-control failures in a laboratory decision task as well as in real-life choice conflicts. To test these predictions, we combined functional MRI (fMRI) during a value-based imaginative decision-making task with smartphone-based experience sampling (Fig. 1).
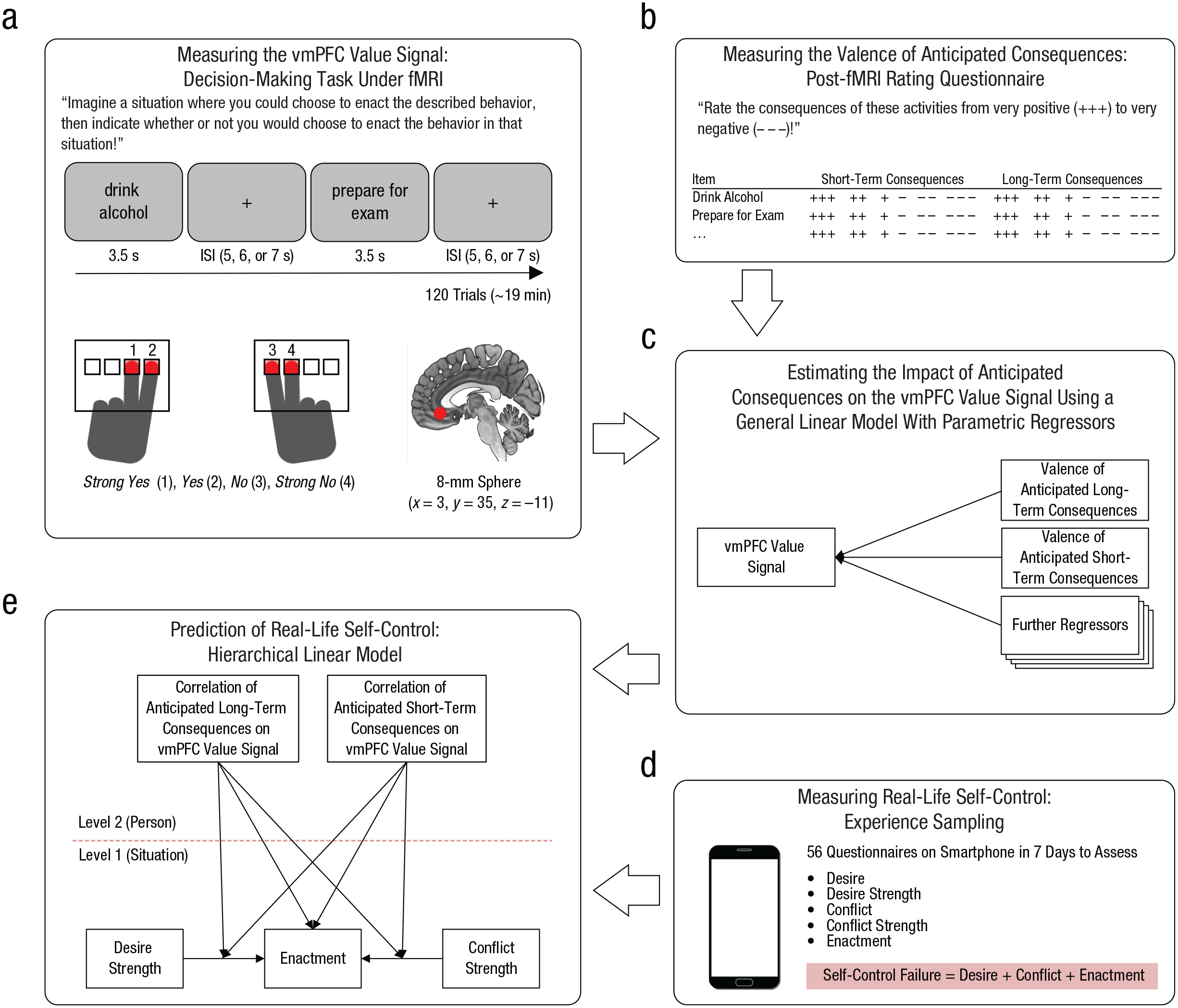
Overview of the data-collection and analysis strategy. In the value-based imaginative decision-making task (a), participants were instructed during functional MRI (fMRI) to imagine a realistic situation in which they would have the option to enact a specific behavior and then to indicate on a 4-point scale the strength of their preference to enact the behavior in the imagined situation. Neuroimaging analyses were restricted to a ventromedial prefrontal cortex (vmPFC) region of interest that was defined a priori (see the text). To measure the valence of anticipated consequences (b), we asked participants to rate the anticipated short- and long-term consequences of the previously seen items on 6-point scales ranging from very positive (+++) to very negative (−−−). To estimate the impact of anticipated consequences on the vmPFC value signal by parametric analysis (c), we defined a general linear model that included parametric regressors for the valence of anticipated long- and short-term consequences. Individual regression weights were used as person-level predictors in the subsequent hierarchical linear model. Smartphone-based experience sampling (d) was used to acquire data on real-life self-control failures. Real-life self-control was predicted (e) by hierarchical linear modeling with conflict situations (Level 1) nested within participants (Level 2). Arrows pointing to rectangles represent main effects, and arrows pointing to other arrows represent interactions. ISI = interstimulus interval.
Method
Participants
We recruited 225 young adults from a representative community sample in Dresden, Germany, for a longitudinal investigation on the onset and course of addictive disorders (for previous studies from this project, see Krönke et al., 2018; Wolff et al., 2016). This sample size is much larger than ones used in previous studies that employed a brain-as-predictor approach. Participants were paid €40 for completing the scanning session and smartphone-based experience sampling. All participants provided written informed consent. The study was approved by the local research ethics committee of the Technische Universität Dresden and conducted in accordance with the Declaration of Helsinki. Following a structured interview, we excluded participants if they had (a) neurological conditions that might affect cognition or motor performance; (b) magnetic-resonance contraindications; (c) lifetime schizophrenia or psychotic symptoms; (d) bipolar, somatoform, anxiety, obsessive-compulsive, or eating disorders; or (e) major depression in the past 4 weeks. Seventeen participants were excluded because of incomplete acquisition of fMRI or experience-sampling data. Fourteen participants were excluded because of floor and ceiling effects—no conflicts reported during experience sampling, too few conflicts (0%–10%) during the fMRI session, or no self-controlled decisions during the fMRI task. Thus, 194 participants (age range = 20–27 years, 118 female) were included in the analyses reported here.
Materials, design, and procedure
Value-based imaginative decision-making task
Neural correlates of subjective value were measured using blood-oxygen-level-dependent (BOLD) fMRI in a value-based imaginative decision-making task in which participants indicated their subjective preference for a broad range of daily behaviors (Fig. 1a). Participants were shown 40 short descriptions that had been carefully selected to evoke conflict between the anticipated short- or long-term consequences of the actions described (e.g., “drink alcohol,” “prepare for exam,” “play video games,” “clean kitchen”). For each item, participants were instructed to imagine a realistic choice situation. They then had to decide whether they would perform the action and to indicate the strength of their preference on a 4-point scale (strong yes, yes, no, strong no). To facilitate interpretation of the subsequent parametric analysis of decision value, we reversed the scale so higher numbers indicated higher decision value (strong no = 1, strong yes = 4). Each trial consisted of an item presented on screen (3.5 s) followed by a fixation cross (jittered interstimulus interval of 5 s, 6 s, or 7 s; average interstimulus interval = 6 s), yielding an average trial length of 9.5 s. After each participant responded, the item remained visible on screen until the end of the 3.5-s stimulation period. Forty different items were randomly presented and repeated three times, yielding a total number of 120 trials and a total duration of 19 min.
After fMRI scanning, participants rated the value of the anticipated short- and long-term consequences of each of the 40 items on a 6-point scale from very positive to very negative (Fig. 1b; for more details about the rating procedure, see the Supplemental Material available online). To facilitate interpretation of the subsequent parametric analysis with regard to the impact of anticipated short- and long-term consequences, we reversed the scale so that higher numbers indicated more positive ratings (very negative = 1, very positive = 6). Ratings were performed after the fMRI task so the decision process during fMRI would not be influenced by the rating process. Ratings were z-scored before they were used as parametric regressors in subsequent analyses.
For each participant, items were individually classified as conflict items when he or she assigned a positive rating to anticipated short-term consequences and a negative rating to anticipated long-term consequences, or vice versa. Decisions on conflict trials were classified as self-controlled when the participant expressed a preference to resist a temptation associated with negative long-term outcomes or expressed a preference to endure an aversive situation associated with positive long-term outcomes.
fMRI data acquisition and analysis
Functional images were acquired using a T2*-weighted gradient-echo echo-planar-imaging sequence (echo time = 25 ms, repetition time = 2 s, flip angle = 78°, slice thickness = 3.2 mm, matrix = 64 × 64, field of view = 19.2 cm, in-plane resolution = 3 mm × 3 mm) on a Siemens MAGNETOM Trio A Tim System 3T scanner with a 32-channel head coil. Thirty-four axial slices, oriented parallel to the anterior commissure-posterior commissure line covering the whole brain, were acquired. In addition, high-resolution anatomical images were acquired (echo time = 2.26 ms, repetition time = 1,900 ms, flip angle = 9°, matrix = 256 × 256, field of view = 25.6 cm, 591 sagittal slices, slice thickness = 1 mm) and coregistered with the functional images. Statistical Parametric Mapping software (Version 8; Wellcome Department of Cognitive Neurology, London, United Kingdom; http://www.fil.ion.ucl.ac.uk/spm/) was used for preprocessing and statistical analyses of fMRI data. After realignment and slice-time correction (to the middle slice), data were normalized to standard Montreal Neurological Institute (MNI) space using the unified-segmentation approach, which is based on the separation of gray matter, white matter, and cerebrospinal fluid (voxel size = 3 mm). Data were spatially smoothed using an 8-mm full-width half-maximum Gaussian filter.
We computed two general linear models (GLMs). To test our first prediction—that brain activity in the vmPFC is parametrically associated with subjective value—we computed GLM 1, which modeled BOLD activity and included two regressors of interest: (a) decision trials and (b) decision trials modulated by decision value (parametric regressor with four levels). Note that decision value is the net value of a specific decision option that integrates subjective and individually weighted (short- and long-term) benefits and costs (Peters & Büchel, 2010b). For each item, decision value was operationalized as the preference strength that participants expressed in the value-based imaginative decision-making task: strong no, no, yes, strong yes (note that the original scale was reverse scored to facilitate interpretation). One first-level single-participant contrast was computed to assess neural correlates reflecting decision value (parametric regressor decision value vs. baseline; for whole-brain peak activations, see Table S1 in the Supplemental Material).
Multilevel Logistic Regression Results: Predicting Self-Control Failures From Desire Strength, Conflict Strength, and Ventromedial Prefrontal Cortex (vmPFC) Modulation by Anticipated Long- and Short-Term Consequences
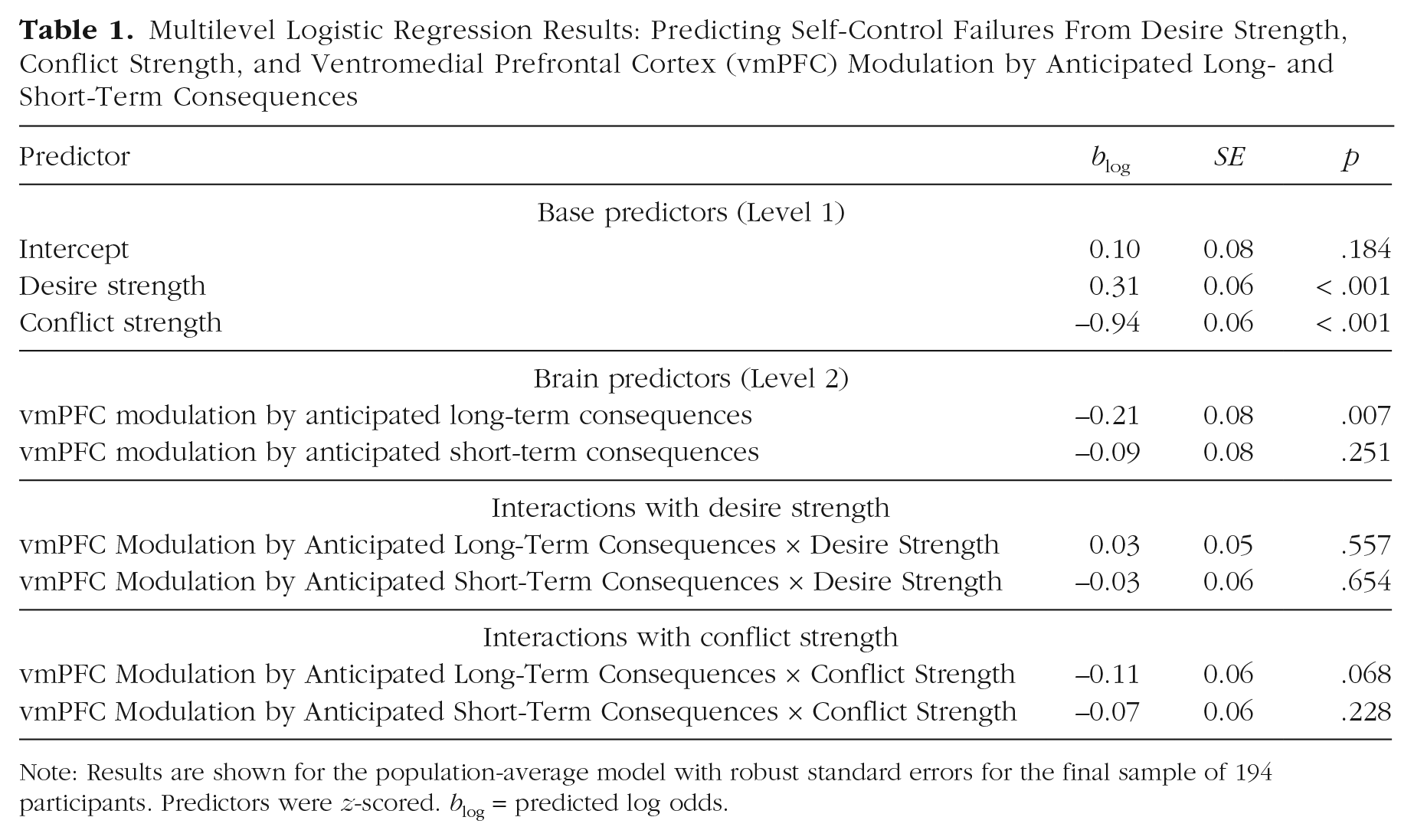
Note: Results are shown for the population-average model with robust standard errors for the final sample of 194 participants. Predictors were z-scored. blog = predicted log odds.
To test our second and critical prediction, we first determined to what extent each participant’s anticipated long-and short-term consequences were reflected in vmPFC activity (Fig. 1c). To this end, a second GLM (GLM 2) was estimated that included three regressors of interest—(a) decision trials, (b) decision trials modulated by anticipated long-term consequences (parametric regressor with six levels), and (c) decisions modulated by anticipated short-term consequences (parametric regressor with six levels)—as well as a fourth regressor: (d) interaction of anticipated short- and long-term consequences, capturing additional variability in the data. Values for the parametric regressors of anticipated short- and long-term consequences were derived from participants’ post-fMRI ratings. Two first-level single-participant contrasts were computed to assess vmPFC modulation by anticipated long-term consequences (parametric regressor anticipated long-term consequences vs. baseline) and to assess vmPFC modulation by anticipated short-term consequences (parametric regressor short-term consequences vs. baseline; for whole-brain peak activations, see Table S1).
For both GLMs, regressors were modeled as boxcar functions with a duration equal to the participant’s reaction time for that trial. The six motion parameters estimated during realignment, session constants, and missed trials were included as regressors of no interest. Further, for all GLMs, the regressors of interest and the missed trial regressor were convolved with a canonical form of the hemodynamic response. Data were high-pass filtered with a cutoff period of 100 s to account for the relatively fast pacing of our experiment. Note that two separate GLMs were set up because net decision value (the independent variable in GLM 1) was assumed to result from the integration of short- and long-term consequences (independent variables in GLM 2).
Extraction of parameter estimates from the region of interest (ROI)
The MNI coordinates reported by Levy and Glimcher (2012) in their meta-analysis (x = 3, y = 35, z = −11) were chosen as the center of the vmPFC ROI. These coordinates were also used in previous research (e.g., Hare et al., 2009), and the corresponding brain region is known to be involved in the representation of subjective value during decision-making. We extracted β values from this region for each of the four levels of the parametric regressor decision value (strong yes, yes, no, strong no) using the rfxplot toolbox (Gläscher, 2009). A search volume (sphere of 8-mm radius) was centered around the vmPFC ROI, and βs were extracted from individual spheres (5-mm radius) around single-participant peaks within this ROI to allow for variability between subjects (Hare et al., 2009). Finally, percentage signal change was calculated in the individual peak voxel for each level of the parametric regressor decision value using the MarsBaR approach (Brett, Anton, Valabregue, & Poline, 2002):

where β(task) refers to the parameter estimate of the effect of interest, max(HRF) is the maximum of a single event of the current duration convolved with the current basis function, and β(constant) is the parameter estimate if the current session is constant. Across participants, averaged percentage signal change and standard errors of the mean were plotted for each level of the parametric regressor. To test whether the parametric modulation was statistically significant, we conducted a repeated measures analysis of variance including one within-participants factor (decision value) with four levels (strong no, no, yes, strong yes).
For GLM 2, percentage signal change associated with anticipated long- and short-term consequences was extracted in the same vmPFC ROI from the same individual participant peaks as identified with GLM 1.
Assessment of real-life self-control failures
Real-life self-control failures were assessed using an experience-sampling procedure adapted from the work of Hofmann, Baumeister, et al. (2012; Fig. 1d). Participants were provided with smartphones equipped for experience sampling (using the customizable application movisensXS; movisens, 2014), which they carried with them continuously for 7 days. Eight alarms per day were issued randomly within a 14-hr window, starting at either 8 a.m., 9 a.m., or 10 a.m., depending on participants’ habitual waking hours. When accepting an alarm, participants were prompted to complete a short questionnaire to examine the occurrence of self-control failures in the hour preceding the alarm. First, they were asked whether they had experienced a desire within the past hour. If they reported a desire, they were asked (a) to indicate the strength of the desire on a scale from 1 (very weak) to 6 (very strong), (b) to select the respective type of desire from a list of 19 categories (e.g., eating, sleeping, drinking, and smoking; for an overview of desire domains by incidence, see Fig. S1 in the Supplemental Material), and (c) to indicate whether the desire was in conflict with a superordinate goal. If they reported a conflict, participants were asked to rate the conflict strength on a scale from 1 (very weak) to 6 (very strong) and to indicate whether they had attempted to resist the desire and whether they had enacted the desired behavior. Each participant completed up to 56 questionnaires, depending on response rates (see Table S2 in the Supplemental Material). Self-control failures were operationalized as occasions on which participants enacted conflict-laden desires.
Multilevel Logistic Regression Results: Predicting Self-Control Failures From Desire Strength, Conflict Strength, Ventromedial Prefrontal Cortex (vmPFC) Modulation by Anticipated Short- and Long-Term Consequences, and Trait Self-Control
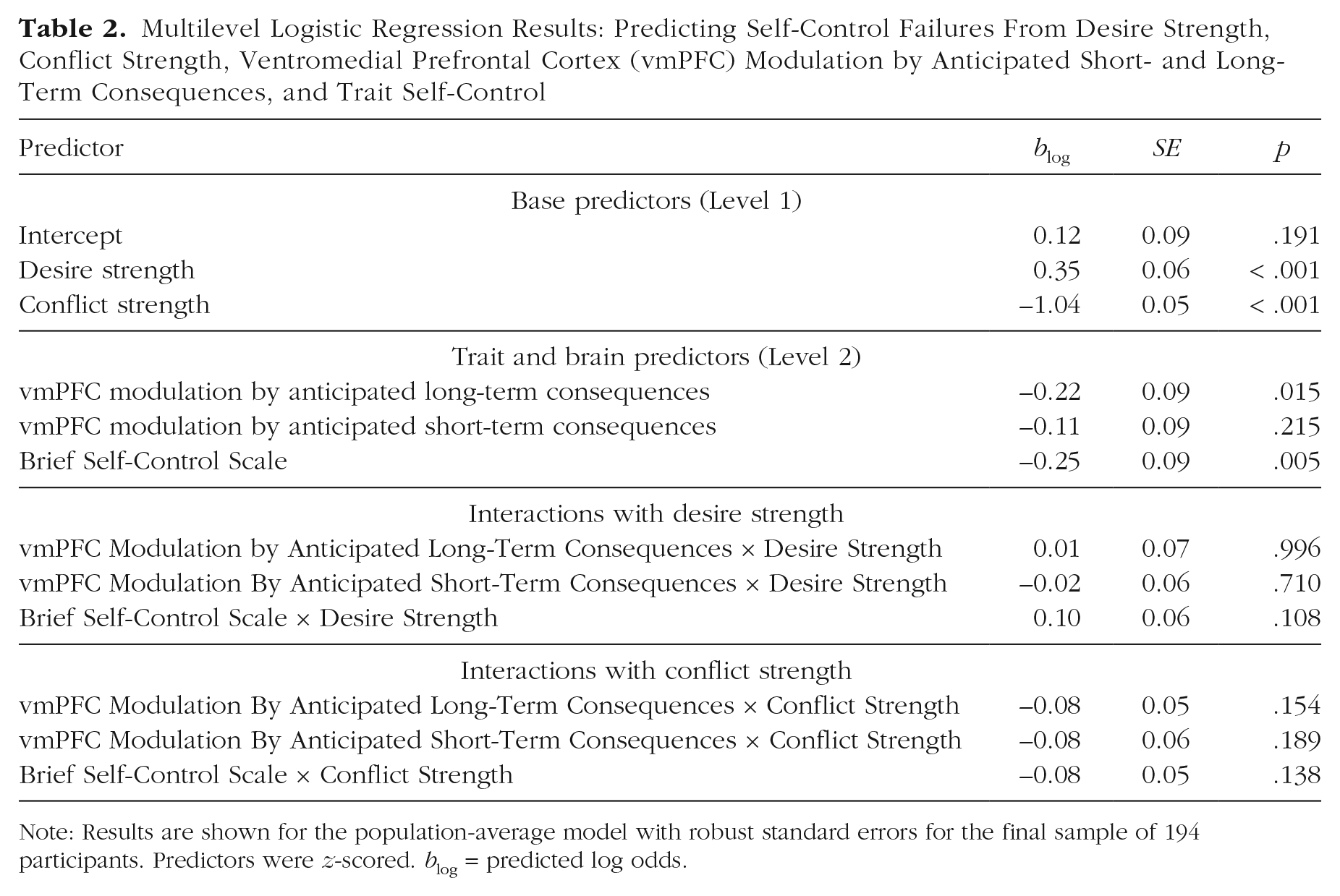
Note: Results are shown for the population-average model with robust standard errors for the final sample of 194 participants. Predictors were z-scored. blog = predicted log odds.
Self-reports of trait self-control
Participants’ self-evaluations of trait self-control were assessed with a German version of the Brief Self-Control Scale (BSCS; Bertrams & Dickhäuser, 2009; Tangney, Baumeister, & Boone, 2004). The BSCS comprises 13 items (e.g., “I am good at resisting temptation”; “People would say that I have iron self-discipline”; “Sometimes I can’t stop myself from doing something, even if I know it is wrong”) and is one of the most widely used questionnaires in self-control research. High BSCS scores indicate high levels of trait self-control.
Prediction of real-life self-control failures from fMRI data
To test whether the modulation of vmPFC activity by anticipated long-term consequences predicted self-control failures, while taking into account the nested structure of the experience-sampling data, we subjected fMRI and experience-sampling data to logistic multilevel regression analyses (Raudenbush, Bryk, Cheong, Congdon, & du Toit, 2011; Fig. 1e). Because enactment of a desire indicates a self-control failure only when the desire is in conflict with long-term goals, only those situations in which such conflicts were reported were analyzed. Two hierarchical linear models (HLMs) were built, with conflict situations (Level 1) nested within participants (Level 2). Level 1 predictors for both HLMs were conflict strength and desire strength. Individual percentage signal changes associated with vmPFC modulation by anticipated long- and short-term consequences were incorporated as z-scored Level 2 predictors in HLM 1. In line with the brain-as-predictor approach, a second HLM was computed (HLM 2), extending the original model by including z-scored BSCS scores as an additional Level 2 predictor.
Results
Imaginative self-control during the value-based decision-making task
On average, items involved conflicts between the value of short- and long-term outcomes in 62.85 trials (SD = 20.83, range = 15–108) out of 120 total trials, that is, in 52.3% of trials (SD = 17.35). In 28.28 of those trials (SD = 16.29, range = 1–80), that is, in 43.2% (SD = 18.85) of conflict trials, participants indicated that they would make a self-controlled decision in the imagined situation (enacting a behavior that was rated to have negative short-term consequences or not enacting a behavior that was rated to have positive short-term consequences). As predicted, in-task self-control was reliably correlated with vmPFC modulation by long-term consequences (r = −.233, p < .001) but not with vmPFC modulation by short-term consequences (r = −.029, p = .693; for scatterplots, see Fig. S2 in the Supplemental Material). Note that the correlation between in-task self-control and vmPFC modulation by long-term consequences was significantly larger than the correlation between in-task self-control and vmPFC modulation by short-term consequences, t(191) = −1.998, p < .05. In 34.56 trials (SD = 14.33, range = 8–79), participants indicated that they would likely fail to use self-control. The average percentage of missed responses was low (M = 2.4%, SD = 2.98).
Consistent with the idea that optimal decision-making requires the integration of several relevant aspects, within-participants correlations of all 194 participants showed that decision value was determined by both long-term consequences (mean r = .471, 95% confidence interval, or CI = [.436, .505]) and short-term consequences (mean r = .552, 95% CI = [.518, .587]). Note that these correlations indicate that across all individuals (i.e., irrespective of their degree of self-control), both short- and long-term consequences influence the integrated decision value. This is, of course, perfectly compatible with our primary hypothesis that individual differences in the weight of long-term outcomes determine the probability of making self-controlled decisions.
Real-life self-control during experience sampling
On average, participants responded to 43.65 (SD = 9.30) of the 56 issued alarms (77.9%) and reported 31.55 desires (SD = 9.73), 11.58 (SD = 7.33) of which were conflict laden (36.7%). Of the conflict-laden desires, 6.13 (SD = 4.91) were enacted and resulted in self-control failures (52.9%; for descriptive statistics of experience-sampling data, see Table S2).
vmPFC activity correlates with decision value
Before vmPFC activity was related to real-life self-control, we determined for each participant individually the vmPFC region in which activity increased parametrically with decision value (GLM 1). Betas reflecting decision value were extracted from 5-mm spheres around individual peaks to allow for variability between subjects. A repeated measures analysis of variance with the within-participants factor decision value (strong no, no, yes, strong yes) showed a strong effect for a parametric increase of vmPFC activity with decision value (Fig. 2a), F(3, 570) = 27.03, p < .001. Although the statistical significance of this effect is not very surprising given that the analysis was restricted to spheres around individual peak voxels that were significantly correlated with decision value, we have included this result for illustration. That a vmPFC region reflecting decision value was found is consistent with previous results (e.g., Hare et al., 2009) in showing that the vmPFC encodes a common value signal for imagined daily behaviors.
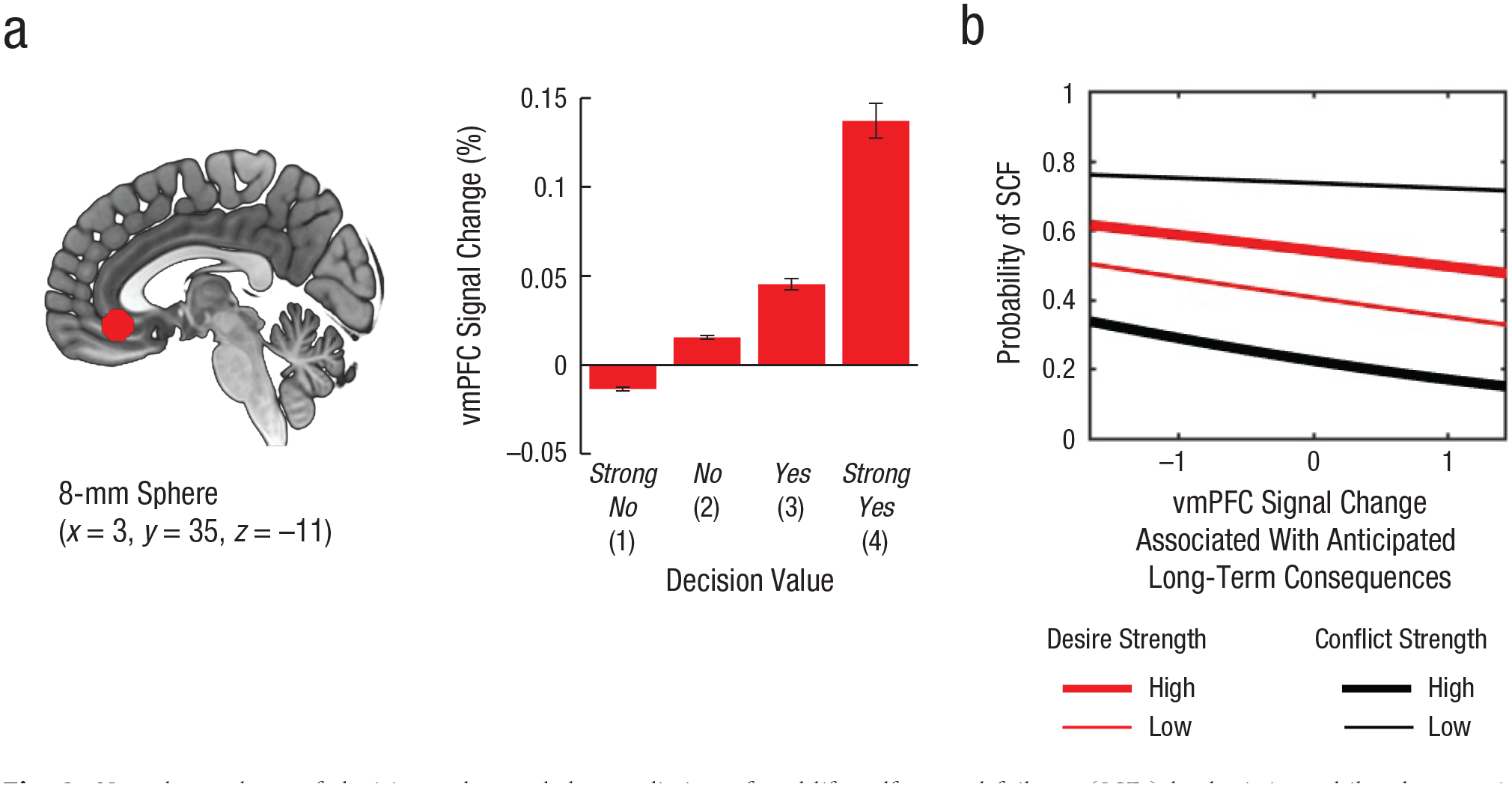
Neural correlates of decision value and the prediction of real-life self-control failures (SCFs) by logistic multilevel regression analysis. Percentage signal change was extracted from individual 5-mm spheres in an 8-mm ventromedial prefrontal cortex (vmPFC) region of interest (a; highlighted in red in the brain image). The graph shows percentage signal change for each of the four decision values. Error bars depict standard errors of the mean. The probability of real-life SCFs (b) is shown as a function of the vmPFC percentage signal change associated with anticipated long-term consequences, separately for high (1 SD above the mean) and low (1 SD below the mean) levels of desire and conflict strength. Predicted log-odds for SCFs have been transformed to probabilities. Values on the x-axis range from the 5th to the 95th percentiles.
Prediction of real-life self-control
To predict real-life self-control, we built two HLMs with conflict situations (Level 1) nested within participants (Level 2). Level 1 predictors for both HLMs were conflict strength and desire strength, and Level 2 predictors were (a) vmPFC modulation by anticipated long-term consequences and (b) vmPFC modulation by anticipated short-term consequences (HLM 1). Further, HLM 2 included BSCS scores as an additional Level 2 predictor.
Situational variables
Not surprisingly, and in line with previous findings (Hofmann, Baumeister, et al., 2012), the frequency of self-control failures increased with desire strength (blog = 0.30, SE = 0.06, p < .001, odds ratio, or OR = 1.36, 95% CI = [1.21, 1.52]) and decreased with conflict strength (blog = −0.95, SE = 0.06, p < .001, OR = 0.39, 95% CI = [0.34, 0.44]), indicating that self-control failures were most likely in situations in which desire strength was high, conflict strength was low, or both (Table 1 and Fig. 2b).
vmPFC modulation by anticipated long- and short-term consequences
Most importantly, percentage signal change associated with vmPFC modulation by anticipated long-term outcomes (GLM 2) was reliably inversely correlated with self-control failures (blog = −0.21, SE = 0.08, p = .007, OR = 0.81, 95% CI = [0.69, 0.94]). Individuals whose vmPFC signal was modulated more strongly by the value of anticipated long-term consequences were less likely to commit self-control failures in real-life conflict situations (Fig. 2b and Table 1). The interaction between the vmPFC modulation by anticipated long-term consequences and conflict strength just missed significance (blog = −0.11, SE = 0.06, p = .068, OR = 0.90, 95% CI = [0.80, 1.01]; Table 1). There were no statistically significant interactions between vmPFC modulation by anticipated long-term consequences and desire strength or conflict strength (all ps > .05; Table 1). Consistent with our prediction that self-control failures primarily reflect an insufficient modulation of neural value signals by anticipated long-term consequences, percentage signal change associated with the vmPFC modulation by anticipated short-term outcomes was not reliably associated with self-control failures (blog = −0.09, SE = 0.08, p = .251, OR = 0.91, 95% CI = [0.77, 1.07]). Note, however, that based on the Bayesian 95% credible interval ([–0.19, –0.05]), inferences regarding a zero effect of short-term consequences should be treated with caution. 1 There were no significant interactions of percentage signal change associated with the vmPFC modulation by anticipated short-term consequences with desire strength or conflict strength (all ps > .05; Table 1). See Table S3 in the Supplemental Material for the prediction of real-life self-control by behavioral ratings and Table S4 in the Supplemental Material for the prediction of real-life self-control by behavioral ratings and vmPFC modulations.
Trait self-control
HLM 2 extended the original model by including BSCS scores as an additional Level 2 predictor. Importantly, the main effect of vmPFC modulation by anticipated long-term consequences remained significant when BSCS score was included as an additional Level 2 predictor (blog = −0.22, SE = 0.09, p = .015, OR = 0.80, 95% CI = [0.67, 0.96]; Table 2). This shows that vmPFC modulation by anticipated long-term consequences explained variability in self-control failures that was not explained by trait self-control. Self-control failures were nonetheless also predicted by BSCS score; as expected, participants with low trait self-control were significantly more likely than participants with high trait self-control to commit self-control failures (blog = −0.25, SE = 0.09, p = .005, OR = 0.78, 95% CI = [0.65, 0.93]). There were no interactions of trait self-control with desire strength or conflict strength.
Discussion
In contrast to an increasing number of reports of zero relationships between laboratory tasks measuring executive functions and measures of real-life self-control (Eisenberg et al., 2019; Enkavi et al., 2019; Saunders et al., 2018), the present study showed that, in a large sample of 194 participants, investigating the role of anticipations of future consequences in valuation processes is a promising direction for elucidating the cognitive and neural mechanisms mediating real-life self-control. Our findings contribute both to our understanding of the role of value-modulation processes in the vmPFC for self-control and to the debate concerning the ecological validity of behavioral and neuroimaging laboratory tasks for the prediction of individual differences in real-life self-control. Expanding previous evidence for the involvement of the vmPFC in the computation of subjective value (Lin et al., 2015), our study showed that activity in the vmPFC reflected the subjective value of (imagined) daily behaviors related to a wide range of different goals and desires. Most importantly, our study provided the first evidence that the degree to which this neural value signal is modulated by anticipated long-term consequences is reliably associated with individual differences in the propensity to commit self-control failures in real-life conflict situations. Thus, in contrast to dual-systems theories, which conceptualize self-control as the suppression of an impulsive system by a reflective system (Heatherton & Wagner, 2011; Hofmann et al., 2009), our results provide consistent evidence for a process model according to which self-control (both in a laboratory decision task and during real-life conflicts) depends on the modulation of a common value signal by anticipated future outcomes (Berkman et al., 2017). In contrast, the modulation of the vmPFC value signal by short-term consequences was neither correlated with in-task self-control (Fig. S2) nor a significant predictor of real-life self-control (Fig. 2). Although this pattern of results suggests that individual differences in self-control primarily or exclusively depend on the degree to which anticipations for future consequences modulate valuation processes, we can of course not exclude the possibility that (despite our large sample size) we might have missed a small effect of short-term valuations. The possible relevance of the ability to attenuate the impact of short-term outcomes for self-control thus remains an open question for further research.
Our findings go in important ways beyond previous studies, which showed that self-control in laboratory food-related choices depends on the relative weight with which the vmPFC value signal is modulated by short-term taste and long-term health information (Hare et al., 2009; Hare, Malmaud, & Rangel, 2011). Our results provide direct evidence that not only relatively inconsequential choices in laboratory tasks but also self-control in real-life choice conflicts reflect individual differences in how anticipated long-term consequences are weighted into integrated neural representations of subjective value. This conclusion is consistent with recent evidence suggesting that the vmPFC is involved in the assignment of affective value to imagined future episodes (Benoit, Szpunar, & Schacter, 2014; Lin et al., 2015) and thereby supports farsighted choices (Benoit, Gilbert, & Burgess, 2011; Peters & Büchel, 2010a). Moreover, our results suggest that the well-documented role of cognitive control networks (including the dorsolateral prefrontal cortex) in self-control (Berkman et al., 2011; Berkman et al., 2017; Hare et al., 2011; Krönke et al., 2018; Lopez et al., 2014; Wolff et al., 2016) may at least partly consist of the top-down adjustment of the relative weight with which long-term outcomes and their anticipated affective consequences are integrated into a common value representation in the vmPFC (Rudorf & Hare, 2014). From an applied and clinical perspective, this assumption together with our present results suggests that self-control in real-life choice conflicts may be facilitated by interventions or environmental cues that help individuals to direct attention to long-term consequences and promote future-directed thought during conflict-laden decisions (Baumeister, Vohs, & Oettingen, 2016; Hare et al., 2011).
In light of the often low retest reliabilities of behavioral tasks measuring self-control (Enkavi et al., 2019) and the lack of correlations between self-reports of self-control and tasks measuring self-control (Eisenberg et al., 2019; Saunders et al., 2018), our results suggest that it is important to include different measures of self-control as predictors for real-world outcomes. Of note, we found that, in addition to behavioral and neural predictors of real-life self-control, a self-report measure of trait self-control (BSCS) was a valid predictor of real-life self-control, but the two sets of predictors explained unique portions of the variability in real-life self-control. This finding is in line with recent evidence suggesting that the relation between trait self-control and real-life behavior is mediated primarily by the degree to which individuals have acquired beneficial habits that help them avoid self-control conflicts (Galla & Duckworth, 2015). By contrast, cognitive control processes supporting future thinking and the modulation of valuation processes by anticipated long-term consequences may be particularly relevant when individuals are confronted with acute temptations (as we assessed with our experience-sampling protocol) and have not yet acquired habitual response patterns supporting self-controlled behavior.
In conclusion, our study provides evidence that the modulation of value signals in the vmPFC by anticipated long-term consequences improves self-control not only in the laboratory but also in real-life choice conflicts. This demonstrates the usefulness of a brain-as-predictor approach as a means to overcome the gap between laboratory research and real-life behavior.
Supplemental Material
Kronke_Supplemental_Material – Supplemental material for Predicting Real-Life Self-Control From Brain Activity Encoding the Value of Anticipated Future Outcomes
Supplemental material, Kronke_Supplemental_Material for Predicting Real-Life Self-Control From Brain Activity Encoding the Value of Anticipated Future Outcomes by Klaus-Martin Krönke, Max Wolff, Holger Mohr, Anja Kräplin, Michael N. Smolka, Gerhard Bühringer and Thomas Goschke in Psychological Science
Footnotes
Transparency
Action Editor: Michael Inzlicht
Editor: D. Stephen Lindsay
Author Contributions
T. Goschke conceptualized the research hypothesis and study design. K.-M. Krönke, A. Kräplin, and M. Wolff recruited and screened participants, K.-M. Krönke acquired the functional MRI data, and M. Wolff implemented the experience-sampling procedures. K.-M. Krönke and M. Wolff analyzed the data, with contributions from H. Mohr, A. Kräplin, and T. Goschke. K.-M. Krönke, M. Wolff, and T. Goschke wrote the manuscript, with contributions from A. Kräplin, M. N. Smolka, and G. Bühringer.
Notes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.