
Abstract
Based on published pollen data from 44 sites in the Loess Plateau and surrounding areas, we recalculate values of Picea abundance digitized from the original diagrams and construct six isopoll maps to reconstruct the distribution of spruce forest at 2000 yr intervals during the Holocene. Spruce forest gradually expanded from about 10 ka BP ago and reached its peak distribution around 8 ka BP. It retained its maximum extent between 8 ka BP and 6 ka BP, broadly covering the subalpine area and stretching to the valleys of the western Loess Plateau with high densities and even reaching the edges of the adjacent desert regions. After 6 ka BP, spruce forest began to retreat to higher elevations. This trend was accelerated after 4 ka BP. After 2 ka BP, it disappeared completely from the Loess Plateau and most of its surrounding areas. Precipitation strongly dominated by the Asian summer monsoon was the primary controlling factor in the distribution of spruce before 2 ka BP. After about 2 ka BP, the increasing intensity of human activity became the primary factor in the disappearance of spruce in the Loess Plateau, resulting in an enormous impact on the present landscape.
Introduction
Spruce (Picea sp.), which is distributed widely in Eurasia and North America, is the characteristic tree of taiga and mountain forest in middle- and high-latitude areas of the Northern Hemisphere. The history of spruce forests can be a valuable tool for understanding environmental changes, ecological restoration and human impacts on the environment because of its sensitivity to climatic variability, paleodrought, and human-induced deforestation.
As early as the 1980s, the migration history of spruce during the postglacial period of Eurasia and North America was reconstructed in continental pollen maps based on published pollen and macrofossil data (Davis, 1983; Huntley and Birks, 1983; Webb et al., 1983). Spruce is also considered an important component in the reconstruction of Holocene vegetation in North China based on pollen data (Ren and Beug, 2002; Ren and Zhang, 1998). These previous studies provide broad-scale evidence on the history of spruce. However, higher-resolution studies focused on particular regions that are sensitive to climate will help us to reconstruct the variation in spruce distributions in detail over time and better understand the dynamic component.
The Loess Plateau and its surrounding area, where spruce is the dominant species of the conifer forest and needle-broadleaved mixed forest, is an important area for the study of paleoenvironment change as well as geomorphological study. Understanding the natural habits of spruce in the area is significant for researching landscape development, ecological restoration, and human impacts on the environment. In recent decades, new pollen records with high-resolution 14C dates have been published, providing a data source to elucidate the mechanisms of paleoenvironmental changes during the Holocene. Here, we collate 44 high quality pollen records with age data in the Loess Plateau and surrounding areas and construct isopoll maps of Picea pollen at 2000 yr intervals during the Holocene, following the methods suggested by Webb et al. (1983) and Ren and Beug (2002). These maps reveal the temporo-spatial patterns of spruce distribution during different periods of the Holocene.
Study area
The Loess Plateau and its surrounding area (33–40°N; 99–110°E) is located in North and Northwest China. It exhibits complex terrain, including the Qinling Mountains, the Northeastern Tibetan Plateau, the Qilian Mountains, the Liupan Mountains, the Muus Desert and the southern margin of the Badain Jaran and Tengger Deserts (Figures 1 and 2). The mean elevation of the Tibetan Plateau is 4000 m above sea level (a.s.l.), and the valleys of the Loess Plateau and adjacent deserts are located at elevations of about 1200 m a.s.l.
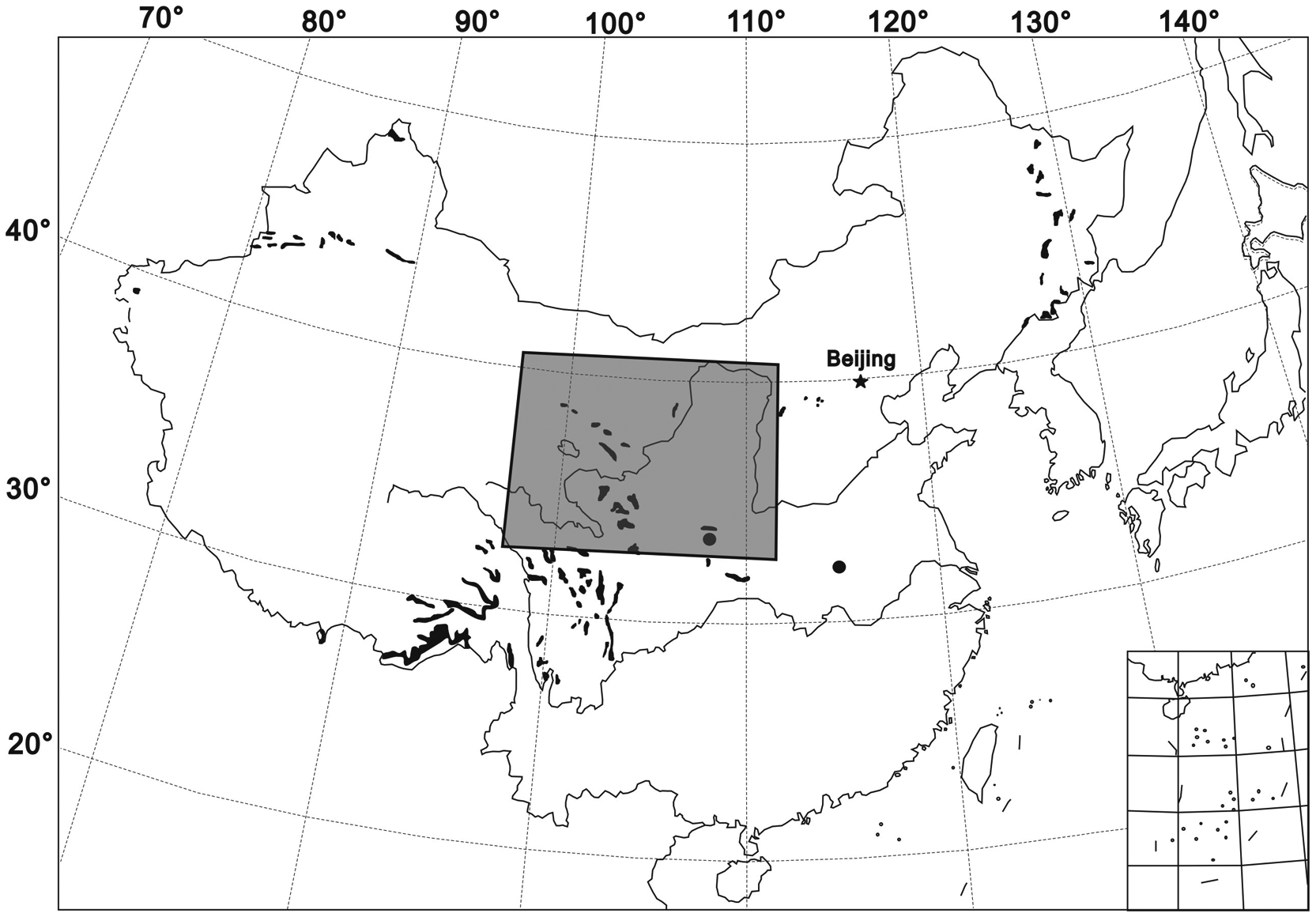
The modern distribution of spruce in China (black shading) and within study area (gray box)
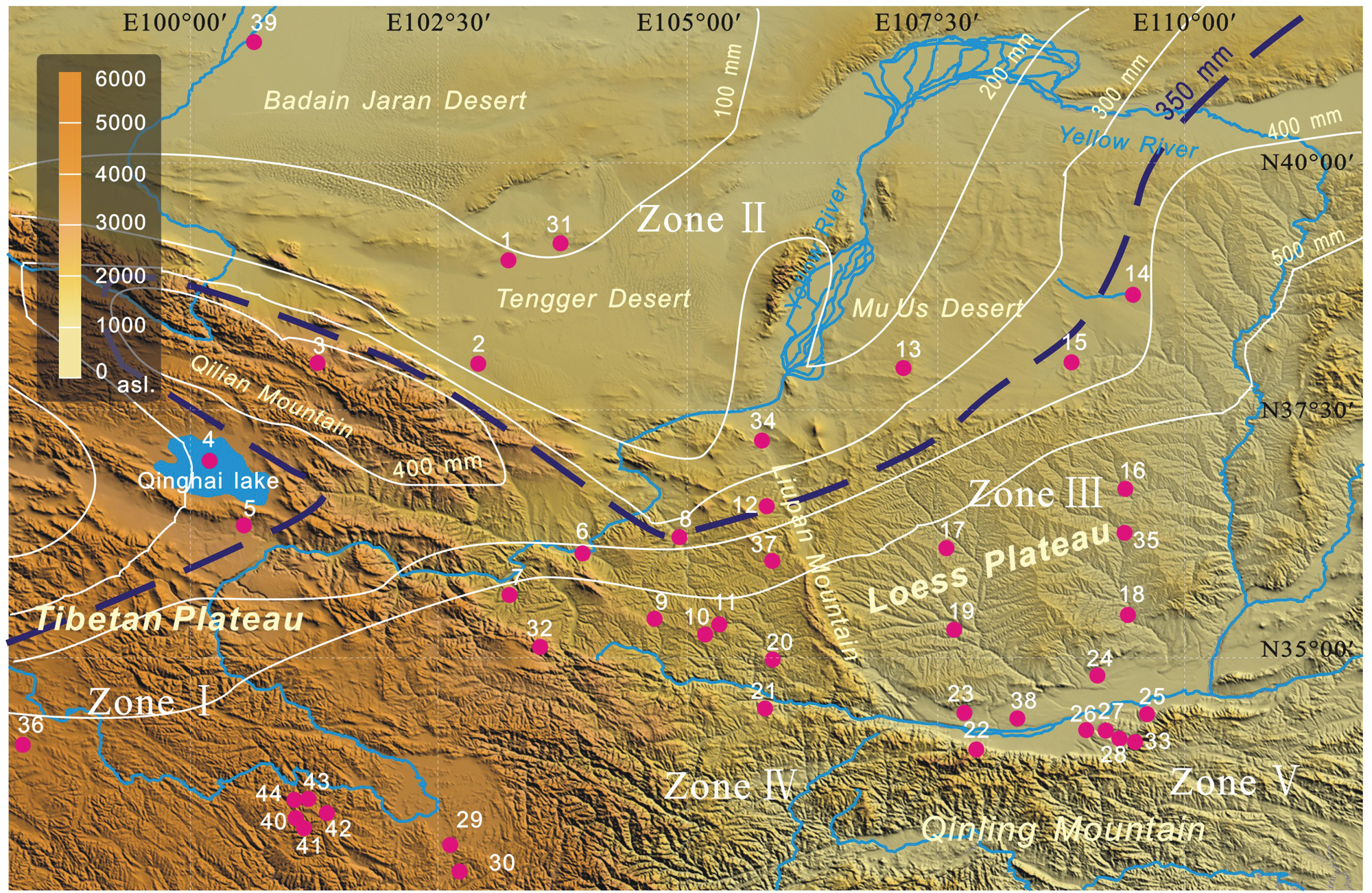
Major landforms, climate zones, isohyets in the study area and locations of pollen data sites used in the study (site numbers as in Table 2). Zone I, alpine subfrigid zone; Zone II, the extreme arid zone; Zone III, semi-arid temperate zone; Zone IV, alpine temperate zone; Zone V, semi-humid warm temperate zone
The study area has climatic patterns controlled by the Asian monsoon, which is a dynamic component of the modern climate system of China. Precipitation controlled by the Asian monsoon decreases gradually from southeast to northwest and plays an important role in the natural landscape of the study region. The boundary between the monsoon zone and the arid climate zone runs approximately along the northwestern margin of the Loess Plateau and corresponds to the 350 mm isohyet of mean annual precipitation (Institute of Geography of CAS, 1999). In addition, landform also plays an important role in regional climate. In the mountainous regions of Qilian and the eastern Tibetan Plateau, mean annual rainfall can reach more than 450 mm/yr because of the orographic upward wind. And in the nearby Badain Jaran and Tengger Deserts, mean annual precipitation is less than 200 mm. Under the influence of these factors, climatic conditions of the study area encompass five zones: the extreme arid zone, semi-arid temperate zone, alpine subfrigid zone, alpine temperate zone, and semi-humid warm temperate zone.
The study of modern vegetation in the Loess Plateau and surrounding areas shows that spruce (including the five Picea species native to the region) is the dominant tree of forests in the mountainous region (Table 1). In the northeastern Tibetan Plateau Spruce (Picea sp.) is primarily distributed between the elevations of 2000 and 4200 m a.s.l. (Hou, 1983; Li, 1999; Lu et al., 2008; Tibetan Investigation Group, 1985). In the Qinling Mountains, Liupan Mountains, and Qilian Mountains (Hou, 1983; Liu et al., 2002), the Picea forest zone is usually restricted to elevations between 1500 and 2700 m (Table 1). Regions in Northwest China below 1400 m a.s.l. are generally too dry to support boreal forest (Institute of Geography of CAS, 1999; Yan et al., 2004).
Distributions and ecological tolerances of Picea species in the study area (Lu et al., 2002, 2008)
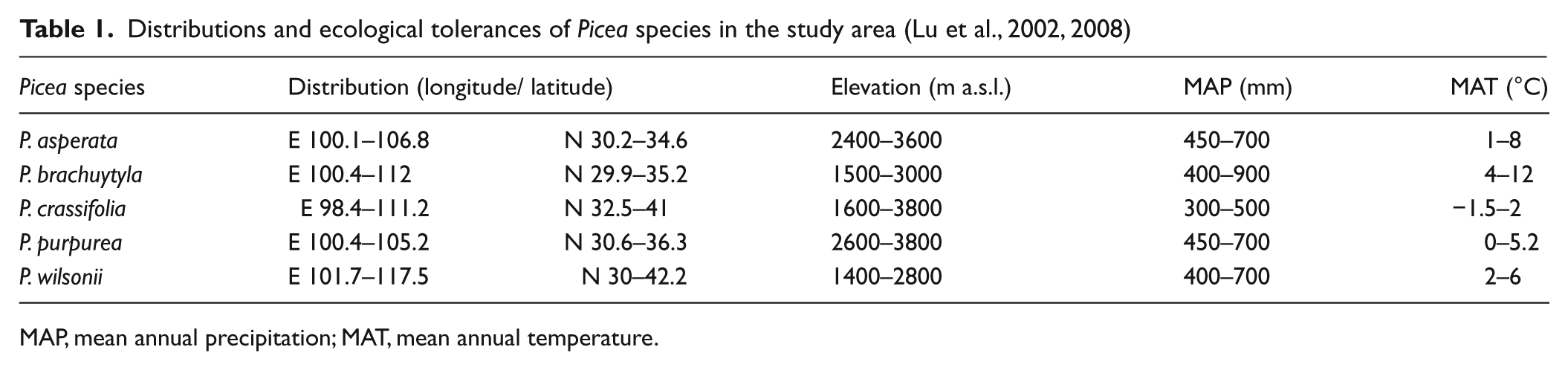
MAP, mean annual precipitation; MAT, mean annual temperature.
Pollen–vegetation relationships of Picea in the study area
Modern pollen spectra are crucial for interpreting vegetation history from fossil pollen data (Bradshaw and Webb, 1985; Davis, 1983; Minckley and Whitlock, 2000). In recent decades, many studies of modern pollen rain have been reported, and vegetation types can be represented effectively by modern pollen spectra in China (Liu et al., 1999; Lu et al., 2008; Shang et al., 2009; Shen and Tang, 1992; Song et al., 1997; Sun et al., 1996a, b; Yan et al., 2004; Zheng et al., 2007). Some studies of modern pollen rain have focused on vegetation types in the Tibetan Plateau (Yu et al., 2001) and helped to generate a semi-quantitative reconstruction of the vertical zonation of vegetation types and mean annual temperatures across the last 50 ka in the Qilian Mountains (Herzschuh et al., 2005). Studies of a modern data set based on 857 samples from the Tibetan Plateau and Xinjiang, China indicate that the spatial pattern of Abies and Picea pollen is highly correlated with the modern distribution of these taxa in the alpine environment and is affected by variation in vegetation, climate and elevation (Lu et al., 2004, 2008).
It was generally considered that saccate pollen, such as that of Picea and Pinus, can be transported long distances through the air. But, the average weight of Picea pollen is 72.8×10−9 g/grain, much heavier than Pinus (18.4×10−9 g/grain) and Betula (6.1×10−9 g/grain) pollen, whose potential for long-distance transport is well known (Wang and Wang, 1983). The smaller saccus of Picea compared with that of Pinus pollen also limits the aerial transport of Picea pollen (Sugita, 1993; Tallantire, 1977). Therefore, large amounts of Picea pollen are not usually transported very far from source areas. In spruce forests, the modern pollen assemblage is dominated by Picea pollen, but the percentage of Picea decreases sharply away from the forest edge and is usually lower than 2% in most surface samples from shrublands, meadows and grasslands (Li, 1999; Shang et al., 2009). If the percentage of Picea pollen is greater than 15%, spruce forest is usually inferred to exist nearby (Li, 1998). However, the percentage of Picea pollen can be as high as 15% in desert and tundra regions with little to no vegetative cover, even if the closest spruce forest is 10 km away (Cheng et al., 2004; Yang et al., 2004). Pollen production and concentration are very low in regions with little vegetation. Thus, Picea pollen can reach a relatively high proportion even though the absolute number of Picea pollen grains transported over such a long distance is small.
Data and methods
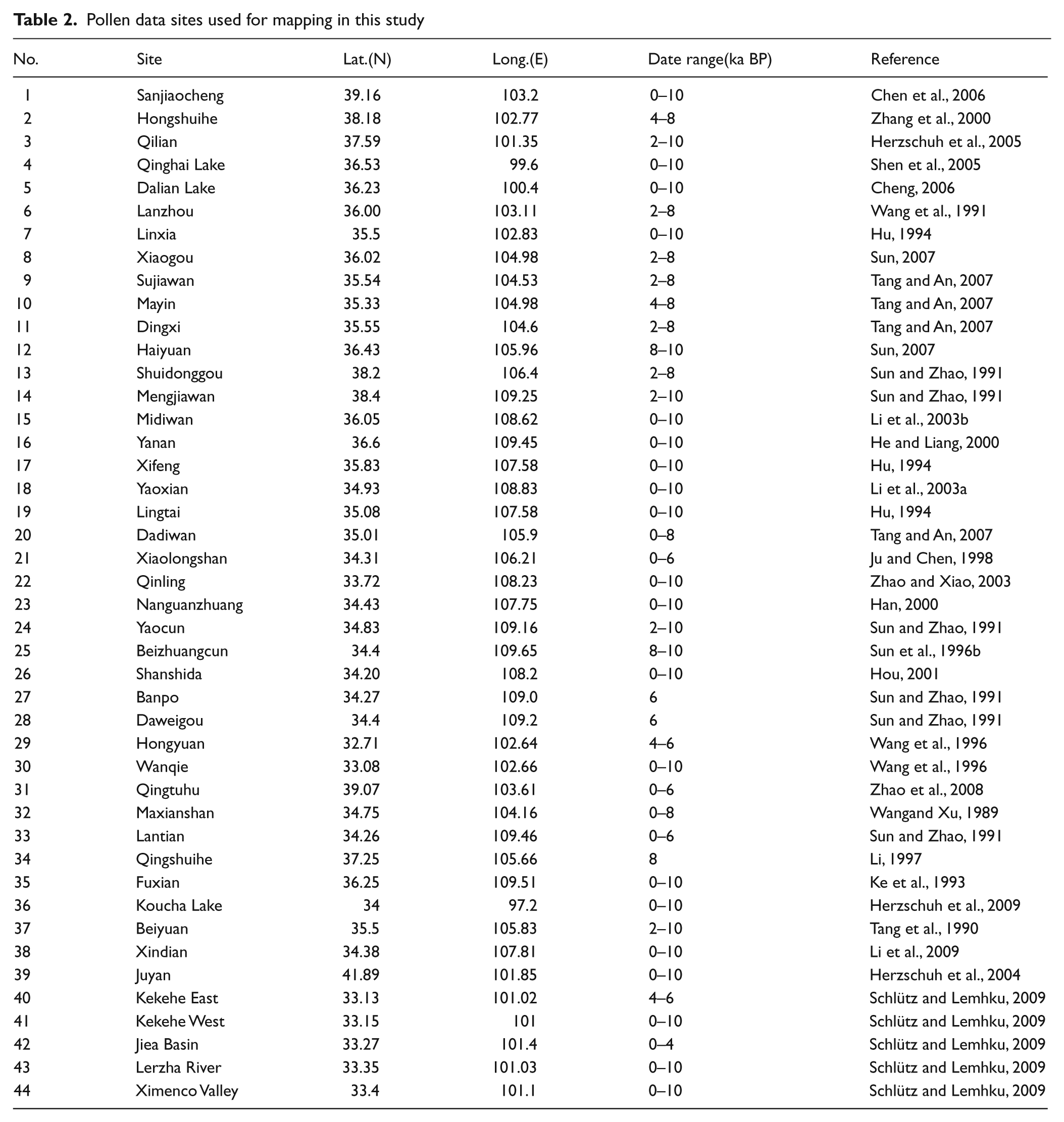
All data used here were taken from the Holocene pollen data base, recent publications, and Master’s or Doctoral dissertations (Figure 2, Table 2). Pollen sites were selected on the basis of sediment type, elevation, sampling interval or time resolution, and radiocarbon dating. The 44 pollen sites obtained from lake sediments, peat and loess samples represent geographic units including loess tableland, river valley, mountainside, desert area, basin and plateau area etc. (Figure 2, Table 2). Absolute pollen count of most selected data is over 300. Some pollen data from the loess sections, which the minimum pollen count is 200 grains, were also selected because of the lower pollen concentration in loess sediments.
Pollen data sites used for mapping in this study
In some published reports that we selected for mapping, including those for Lantian, Fuxian and Sujiawan (Ke et al., 1993; Sun and Zhao, 1991; Tang and An, 2007), Picea and Abies data were not given separately in reports originally published because these trees exhibit similar ecological characteristics in the study area. Values for Picea pollen digitized from the original diagrams were recalculated based on descriptions in the reports and on the average proportions of Picea and Abies pollen at nearby sites for which published data on the two species had not been combined, and where the ratio of Picea to Abies is usually greater than 3:1 (Sun and Zhao, 1991; Tang and An, 2007). Generally, the error due to recalculation can be regarded as small because the total percentage of Picea and Abies at these sites was very low and would affect the resulting isopoll map only slightly.
Most ages of the pollen data were determined by 14C dating. Dates used for mapping are given in calibrated 14C years. Some age data from loess sections were dated by luminescence dating, which gives results consistent with calibrated 14C years. The ages of a few pollen data sets from loess sections that had not been dated were estimated based on the stratigraphy of loess sediments, for which the age framework has been well established in studies of the Loess Plateau (Kukla et al., 1988; Liu, 1988).
We chose a 2000 yr interval for isopoll mapping. Six maps were generated representing time slices at 10 ka, 8 ka, 6 ka, 4 ka, 2 ka and 0 ka BP. The pollen maps were generated automatically using the software Surfer Version 8.0 for Windows, which was improved for application to vegetation and ecological reconstruction by Ren and Beug (2002). The isoline intervals were defined as 10%, 5%, 3%, and 1%. The contours were drawn using medium smoothing, and the grid size was 74 rows by 100 columns (130 square km per grid).
Results
In the early Holocene (around 10 ka BP), higher values of Picea pollen from the Qilian Mountains, northeastern Tibetan Plateau, Liupan Mountains, and Qinling Mountains indicated that spruce was widely distributed among these mountain areas, which are still the core areas of modern spruce distribution (Figure 3a). The percentages of Picea pollen in the northeastern Tibetan Plateau and Qinling Mountains remained greater than 40% throughout the Holocene (Zhao and Xiao, 2003), indicating that the spruce forest in this area was relatively stable with high population densities. The abundance of spruce pollen was greater than 10% in the Qilian Mountains, including relatively low-elevation areas at the foot of the mountains which had spruce forest (Herzschuh et al., 2005; Shen et al., 2005). Sparse spruce forest was present in the Liupan Mountains. The abundance of Picea pollen in the valleys, such as at Xining, Lanzhou, Tianshui and Guanzhong in the western and southern Loess Plateau, was generally less than 5%. Spruce forest with high population density was thus mostly distributed in mountainous areas with elevations greater than 2000 m a.s.l.

Isopoll maps for Picea at 10, 8, 6, 4, 2 and 0 ka BP in China
In the early Holocene in the northwestern part of the study area, Picea abundances were greater than 15%, on average in pollen records from lacustrine sediments in deserts, such as at Zhuye Lake in the Tengger Desert (Chen et al., 2006). In the Muus Desert and in Juyan Lake in the Badain Jaran Desert, Picea abundances were generally greater than 5% during this period (Herzschuh et al., 2004; Li et al., 2003b). Around 8 ka BP, the proportions of Picea pollen increased throughout the study region, reaching their greatest extent and largest population densities (Figure 3b). Picea expanded in the Liupan Mountains and the valleys of the Loess Plateau and reached its peak value in pollen spectra during the last 10 ka (Sun, 2007). Similar increasing trends also occurred in Lake Qinghai and Lake Dalian in the northeastern Tibetan Plateau (Cheng, 2006; Shen et al., 2005). The abundance of Picea pollen remained greater than 40% in the Qinling Mountains (Zhao and Xiao, 2003). In the Tengger and Muus deserts, the abundance of Picea pollen continued to rise (to greater than 5%) (Chen et al., 2006; Li et al., 2003b), indicating expansion of the spruce forest and increasing spruce population density.
Around 6 ka BP, the spruce forest extent began to shrink in the Liupan Mountains, the valleys of the western Loess Plateau, and the northeastern Tibetan Plateau, but spruce still covered a wide region with a relatively high density (Figure 3c). Abundant fossil spruce charcoal has been found in Neolithic culture layers from around 6 ka BP near Lake Qinghai (Kaiser et al., 2007), showing that mountainous spruce forest was distributed close to Lake Qinghai. Spruce forest remained at a relatively high density in the southern Qilian Mountains, Tianshui Basin of the western Loess Plateau, southern Liupan Mountains, and northern Qinling Mountains at an average elevation of 1200 m a.s.l. Spruce forest with high population densities existed during the late Yangshao and Majiayao culture periods (6–5 ka BP) (Li et al., 2007; Zhao et al., 1990). Desert spruce forest islands declined to levels similar to those found at 10 ka BP. However, the distribution of spruce forest did not obviously change between 6 ka and 8 ka BP around the Qinling Mountains.
At about 4 ka BP, the distribution and density of spruce forest declined remarkably in the northeastern Tibetan Plateau, western Loess Plateau, Liupan Mountains, and northwestern deserts (Figure 3d). The percentage of Picea pollen in Lake Qinghai and Lake Dalian declined substantially (to less than 1%) after 4.5 ka BP. Pollen records from the western Loess Plateau, such as those from Sujiawan, Dadiwan, and Xishanping, indicate major vegetation changes. Forests, especially spruce forest, declined considerably around 4 ka BP (Cheng, 2006; Li et al., 2007; Tang and An, 2007).
Around 2 ka BP, the pollen data show spruce forest retreating toward the mountains (Figure 3e). Pollen records from the valleys of the Loess Plateau show that spruce forest declined sharply; with the abundance of Picea pollen generally below 1%. Picea pollen disappeared from the desert areas where spruce forest islands had existed before 4 ka BP. The extent of spruce forest declined in the Qilian and Qinling Mountains, but increased in the north of Liupan Mountains (Herzschuh et al., 2005; Sun, 2007; Zhao and Xiao, 2003).
Discussion and conclusion
The distribution and population density of spruce forests is strongly related to climatic conditions in the Northern Hemisphere (Bartlein et al., 1986; Overpeck et al., 1992). Temperature has been considered to be the controlling factor in the distribution of spruce forest for Arctic Eurasia, especially in northeastern Europe and Siberia, minimum winter temperature has a strong controlling effect on the distribution of spruce (Huntley and Prentice, 1988). In the study areas, based on the relationship between temperature and modern spruce distribution in China, Wang (1990) has proposed that the average temperatures of May and September are more important than winter temperature in controlling the growth and development of spruce forest. Generally, in the northern high latitudes or upper part of mountainous forest areas, temperature is the most important limiting factor for spruce growing.
In addition to temperature, water is considered another important controlling factor for spruce forest distribution especially in the semi-arid and arid areas. Most of the spruce forests distribute in regions where the average annual precipitation is between 400 and 850 mm (Lu et al., 2004, 2008). In the Tianshan Mountains in Xinjiang, Schrenk spruce (Picea schrenkiana) forest is distributed on southern slopes to elevations of 2300 m, but extends downward on northern slopes to 1400 m because of the heavier precipitation brought by westerly winds in northern slopes (Hou, 2001). In the same way, aridity has limited the distribution and development of Koyama spruce (P. koraiensis) and Yeso spruce (P. jezoensis) in the Daxinganling Mountains in Northeast China: these species are distributed only on riverbanks, overcast slopes, and moist areas on low-elevation slopes (Zheng, 1983).
As mentioned above, both temperature and precipitation are important factors controlling the growth of spruce. Just as Tranquillini (1976) proposed in his study of mountainous forests, the upper limit of mountainous forests is normally controlled by temperature, while the lower limit is primarily controlled by precipitation. This is the representative pattern of spruce distribution in the study areas.
Holocene climatic variability has been reported from ice cores, lacustrine sediments, speleothems and loess-paleosol sequences (Bianchi and McCave, 1999; Enzel et al., 1999; Feng et al., 2006; O’Brien et al., 1995; Wang et al., 2005; Zhao et al., 2009, 2010). The Holocene optimum was a warm and moist period that occurred between 10 and 6 ka BP in the Loess Plateau and surrounding areas (An et al., 2000, 2003, 2005; Feng et al., 2006; Shi and Kong, 1992). The climate became less moist with the weakening of the Asian monsoon after about 5 ka BP and then changed to a cool-dry climate in the Loess Plateau and northeastern Tibetan Plateau (An et al., 2000; Shi and Kong, 1992). On the Tibet-Qinghai Plateau, climate changes as a result of changes in the intensity of the East Asian Summer Monsoon in the mid Holocene, probably was the key factor for the widespread forest decline (Herzschuh et al., 2010).
The Picea pollen contour maps for the Loess Plateau and surrounding areas show that the distribution and density of spruce increased from about 10 ka BP and expanded from the Qilian Mountains, Qinling Mountains, and Liupan Mountains to relatively low-elevation areas in the western Loess Plateau and some northwestern desert areas. Spruce forest reached its largest extent and peak density at around 8 ka BP, and remained as such to 6 ka BP. However, spruce forest declined after 6 ka BP and nearly disappeared from the valleys of the western Loess Plateau, retreating to the Liupan and Qinling Mountains, after 4 ka BP. Spruce forest also disappeared from the Lake Qinghai Basin and retreated to the Qilian Mountains after 4 ka BP. Spruce forest disappeared completely from the Loess Plateau and most of the surrounding areas and was found only north of the Qilian and Qinling Mountains after 2 ka BP (Figure 4).

Patterns of Spruce distribution change in the study area
Compared with the Picea isopoll maps with these Holocene climate changes, we find that the widest distribution and highest population density of spruce forest corresponds closely to the Holocene optimum, when greater precipitation levels were controlled by the Asian monsoon. Although, it is believed that the distribution of spruce forest is controlled by mean annual temperature during the glacial and interglacial cycles (Xu et al., 1980), the temperature is relatively stable through the Holocene compared with that in glacial and interglacial cycles. Moreover, the lower limit, rather than the upper limit, of the spruce forest in the study areas, which is primarily controlled by precipitation, changes remarkably through the Holocene (Figure 4). Abundant precipitation probably enabled spruce to grow in low-elevation valleys in the Loess Plateau and to extend to the edges of desert regions. With the weakening of the Asian monsoon after 5 ka BP, spruce forests continued to decline in the Loess Plateau and surrounding areas because of lessening precipitation. Therefore, we conclude that temperature is not the main controlling factor in spruce distribution in the Loess Plateau and surrounding areas, but variations in spruce distribution and density are primarily controlled by precipitation, which is strongly dominated by the Asian monsoon.
The relatively high percentages of Picea pollen (greater than 5%) in desert areas in the humid period, early and mid Holocene, probably indicate the presence of developed spruce forest islands near to the sample sites. However, some researchers have attributed the presence of high concentrations of Picea pollen in the lake sediment in desert areas of the early and middle Holocene to long-distance transport of Picea pollen by rivers from the Qiliang mountains hundreds of kilometers away (Cheng et al., 2004; Zhu et al., 2001). Spruce charcoal fossils dated to the early Holocene have been found in the Hunshandake Desert of Inner Mongolia, where average annual rainfall is about 200 mm and the survival of the psammophytic spruce (Picea mongolica) forest still exists on sandy soils there today (Cui and Liu, 1997; Xu et al., 1993). The spruce forest treeline should have moved downward in the humid period, even reaching close to the margins of these sandy areas as a result of the increase in regional rainfall and the improvement in soil moisture.
Although the distribution of spruce is primarily controlled by precipitation, it is difficult to explain why spruce forest disappeared completely from the Loess Plateau and most of its surrounding areas after 2 ka BP. The period from 2 ka BP to the present is not the driest episode during the Holocene (Wang et al., 2005); the ‘Medieval Warm Epoch’, a warm and moist period, occurred between
Chinese agricultural history began as early as 10 000 years ago (Lu et al., 2009). The Loess Plateau is famous as the home of the Dadiwan, Yangshao, and Majiayao cultures in the Neolithic period, and the Qijia and pre-Zhou cultures in the Bronze Age. Agriculture became the dominant economic activity during the Holocene, as recorded by more than 1000 archaeological sites (Li et al., 2009). Farmers need more arable land for crops as populations expand. Therefore, human activities undoubtedly influenced vegetation cover and reduced native vegetation on the most arable land areas (e.g. Björkman, 1997; Li et al., 2007; Nunez and Vuorela, 1978). Furthermore, conifer wood is useful for construction. The decline of spruce forest might not be related to climate change but to selective harvesting of conifers (Figure 5; Li et al., 2007). Therefore, we conclude that ancient farmers reduced spruce forest areas during the Neolithic and Bronze Ages (Cui and Liu, 1997; Li et al., 2007).

Pollen spectrum of Xishanping Neolithic site in Gansu (Li et al., 2007)
The decline of spruce between 6 and 2 ka BP was a gradual process in the study region rather than a sudden change. Considering the relatively sparse human population, the land use type and the use of stone tools, we conclude that the shrinking and retreat of spruce forest was primarily caused by climatic drying between 6 and 2 ka BP. However, human activities strongly affected the vegetation cover during this time. With the development of societies and widespread use of iron tools after 2 ka BP, agriculture and human populations increased rapidly during the Han (202 BCE–220 CE) and Tang Dynasties (618–907 CE) (Figure 6; Ge, 2002; Li et al., 2009). The deforestation process was much stronger during that period than during the Neolithic and Bronze Ages. In the last 600 years, the human population density of the study region increased dramatically during the Ming and Qing Dynasties and reached almost 250 million in the industrial era. Farming expanded to semi-arid regions and around oases in arid regions. The increasing intensity of human activities became the primary factor in the disappearance of spruce forest from the Loess Plateau, which has had an enormous impact on the present landscape.
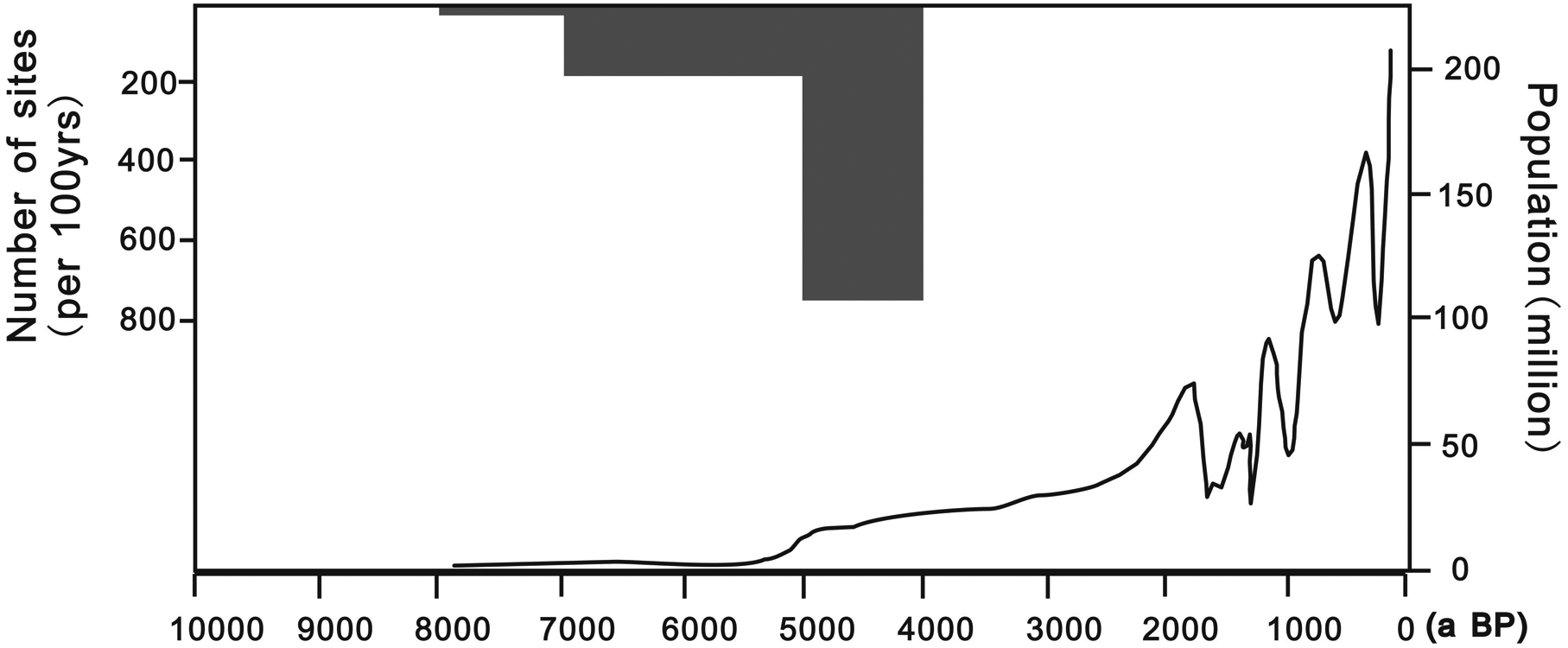
Increase in population of China of recent 10 kyr. The population after 4000 yr BP is calculated from previous Chinese government records and historical documents, and that before 4000 yr BP was calculated by the number and the size of archeological sites (Li et al., 2008)
Footnotes
Acknowledgements
We are grateful to the National Key Basic Research Fund (2010CB950200), the Innovation Program of the Chinese Academy of Sciences (KZCX2-YW-Q1-03), the Most Special Fund for SKLL (LQ0701), and the Grants NSFC (41001062) for supporting the project.