
Abstract
In Northern Patagonia, the long-term vegetation–climate relationships that gave shape to southern South American forests have been the subject of extensive investigations in the Andean regions of Argentina and Chile for about a century. The Río Manso Superior originates in one of the tongues of the Cerro Tronador Glacier, near the international border, discharging into the Pacific Ocean after feeding present Lago Mascardi and flowing east to west on a mountain landscape. Ongoing pollen studies in this watershed provide us with a modern analogue to better understand the vegetation history of the region. A pollen record from a 15 m long sediment core retrieved from Lago Mascardi (41°08°S, 71°34°W) contains continuous evidence of vegetation changes in the Río Manso watershed extending back to the last deglaciation. High Andean steppe vegetation with forest patches, and extended waterlogged areas gave place to a forest, probably deciduous, during the Lateglacial–Holocene transition. A forest diversification took place during the climate amelioration that encompassed the deglaciation whilst the vegetation became more open before the onset of the Huelmo-Mascardi cold reversal. A mixed Nothofagus-Austrocedrus forest expanded during the middle Holocene. This forest became denser under the higher climate variability registered in the region during the late Holocene as shown by independent published data. Statistical analyses of modern pollen samples along an altitudinal transect from low Nothofagus forest and shrubland to high Andean semi-desert support this interpretation. Pollen results are discussed in the context of paleoenvironmental reconstructions at a regional scale.
Introduction
In Northern Patagonia, precipitation and topography determine the arrangement of vegetation in a west to east gradient across the continent. Additionally, sharp climate and vegetation altitudinal gradients occur in the Andes: from west to east and low to high altitudes, Valdivian, North Patagonian and Sub-Antarctic rain forest communities thrive at the western Andean flank. High Andean vegetation prevails near the summits. Deciduous, evergreen Patagonian forest and steppe are replaced from high to low elevations at the eastern flank.
The long-term vegetation–climate relationships in the region have been the subject of exhaustive research that started in 1928–1929 (Auer, 1958) and continues to the present. Studies of fossil pollen, developed in the Chilean Lake District (Heusser, 1966, 2003; Villagrán, 1985) and in the area of Nahuel Huapi National Park (NHNP) (Markgraf, 1983: 84), providing evidences of the major vegetational and paleoclimate changes since the last glaciation.
Westerly winds from the South Pacific Ocean are the major source of moisture, while temperature and precipitation are the main driving factors of vegetation at both flanks of the Andean range, precipitation playing a significant role at the eastern side.
Both vegetation and glacial histories have been taken as evidence of the timing, magnitude, and frequency of latitudinal changes in the westerly winds during and since the Lateglacial maximum. These processes are still poorly understood, and constitute an active area of research (e.g. Abarzúa et al., 2004).
Conflicting results and divergent interpretations of the paleoecological records have led to disagreements about the timing, magnitude, and direction of climatic events during the Lateglacial–Holocene Transition (LG-H) in the region (Moreno, 2000). At the western Andean slopes, three different models of climatic change based on biological proxies (beetles and pollen) have been proposed for this complex period. A ‘single step warming’ (Hoganson and Asworth, 1992), a ‘two step model’, with a cooling episode (Heusser et al., 1996), and a ‘multiple step model’ with several cooling events (Heusser et al., 1996; Moreno, 2000).
At the eastern Andean flank, evidences of full glacial environments at the northeastern coast of Lago Nahuel Huapi by 17 000 14C BP (Markgraf and Bianchi, 1999) were preceded by the rearrangement of the regional hydrological system during the LG-H starting c. 14 000 14C BP (Bianchi et al., 1999). The single step warming model (Hoganson and Asworth, 1992) prevailed in the region until a glacial re-advance as well as a series of climate fluctuations between 15 000 and 8000 14C BP, were shown for the area of Lago Mascardi (Ariztegui et al., 1997; Hajdas et al., 2003).
These findings were supported by a synchronous cold reversal at Huelmo, a palynological locality in the Lake District, Chile (Moreno, 1998; Moreno et al., 2001). Therefore, this climatic event was named Huelmo-Mascardi cold reversal (Hajdas et al., 2003).
Interestingly, the first 14C dated pollen record encompassing the Lateglacial and Holocene periods at the eastern side of the Andes was obtained at Mallín Book, a sedge bog located at the shore of Lago Mascardi (Markgraf, 1983). Although notable vegetation changes are recorded in Mallín Book, they occur with a different timing and no evidences of cold episodes have been proposed in this site for the LG-H (Markgraf, 1983).
To the east of Lago Mascardi no conclusive evidence of a cool reversal during LG-H has been proposed (Jackson, 1996; Markgraf, 1984; Markgraf and Bianchi, 1999; Whitlock et al., 2006). Therefore, the area of Lago Mascardi’s response to deglaciation seems to be in phase with deglaciation episodes at the western side of the Andes, about 300 km to the west, but different to the continuous warming trend suggested for pollen localities located about 30 km towards the northeast (Markgraf, 1984; Markgraf and Bianchi, 1999; Whitlock et al., 2006).
The early Holocene was marked by the amelioration of climatic conditions that favoured forest expansion in the entire region (Bianchi, 1999, 2007; Jackson, 1996; Markgraf, 1983, 1984; Whitlock et al., 2006).
The Río Manso Superior (Upper Manso River) valley is located between the summit of Cerro Tronador, the highest peak at the Chilean–Argentinean border in Patagonia, and the western arm of Lago Mascardi. This mountain landscape is characterized by steep climate and vegetation altitudinal gradients. Dendrochronological studies indicate the sensitivity of this area to climatic fluctuations during the last 1000 years (Masiokas et al., 2009b; Villalba et al., 1990). Because of the mentioned characteristics the area was chosen as an appropriate location to reconstruct vegetation history at millennial and submillennial timescales in the deep Andean glacial valleys on the Argentine side.
Although the time period between 13 000 and 9000 14C yr BP has been studied in detail in previous papers, the entire Holocene pollen sequence of Lago Mascardi has only been briefly discussed (Bianchi, 1999, 2007).
We present here a detailed continuous pollen record of a sediment core retrieved at Lago Mascardi, encompassing the Lateglacial and the Holocene periods, calibrated by an altitudinal transect of modern pollen samples from the Río Manso Superior valley. The comparison of this information with other records from localities at the east and west of the Andean range improved our understanding of vegetational changes through a broader range of climatic periods regionally defined using different proxies (Ariztegui et al., 2007; Masiokas et al., 2009b).
Environmental setting
The Río Manso hydrological system is located within the boundaries of the Nahuel Huapi National Park (NHNP) connecting several water bodies at the front of the Andean range and having a Pacific Ocean derivation (Figure 1A, B).
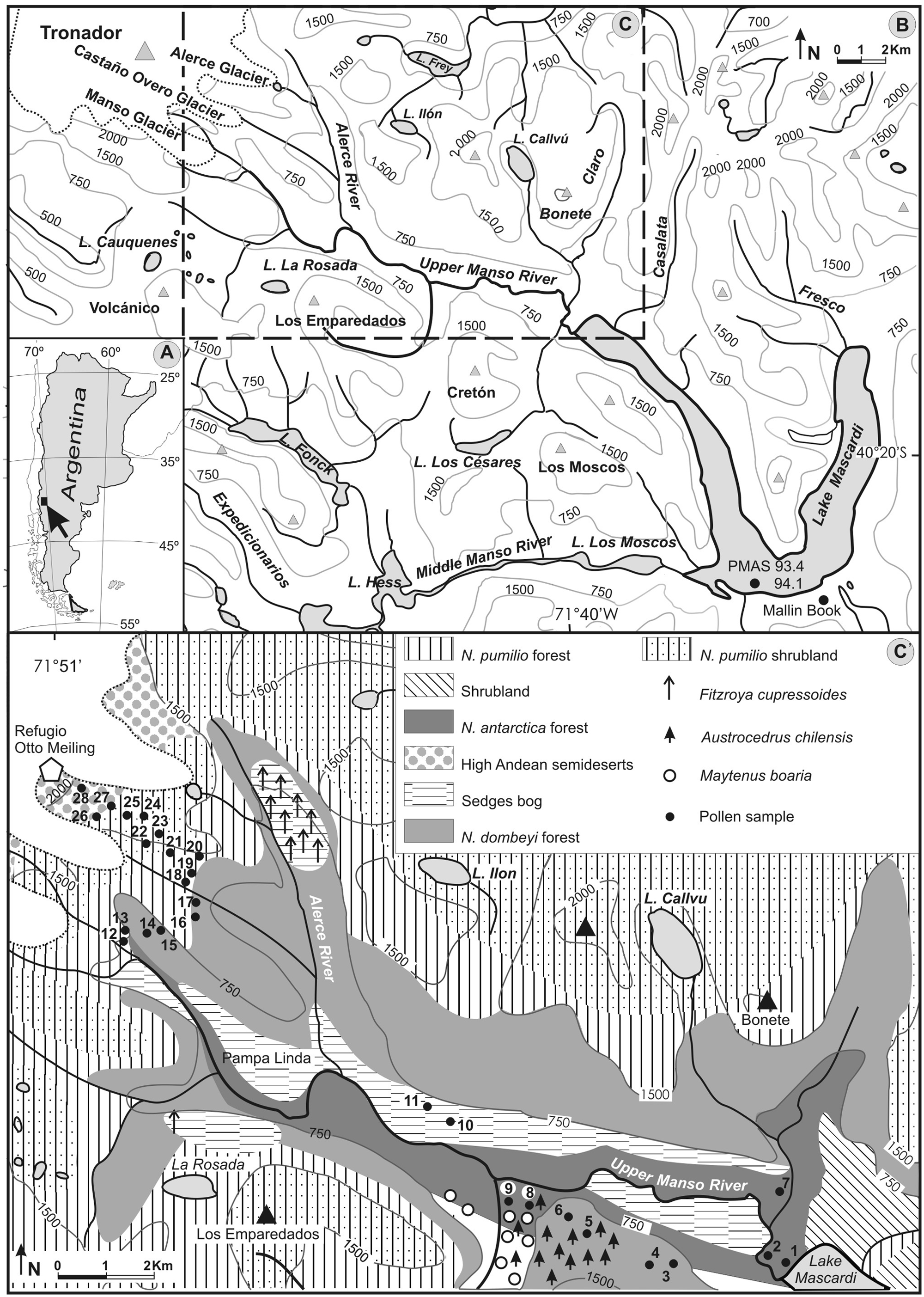
(A) Location map. (B) Area of study indicating palynological sites discussed in the text (solid circle). (C) The Río Manso Superior basin. (C’) Vegetation units along the upper Manso River basin and transect of surface soil and moss pollen samples (solid circles)
The Río Manso Superior is fed by melt waters from three Cerro Tronador’s glaciers which have undergone important variations during recent and Neoglacial times (Masiokas et al., 2009a). It flows to the northwest end of Lago Mascardi (41°31'S, 73°W, 793 m). This oligotrophic lake (39.2 km2, 218 m depth) has the Río Manso Medio as its only outlet.
The drainage basin of the valley of Río Manso Superior (247 km2) has a NW–SE orientation showing an abrupt topography, corresponding to an ancient glacier valley (Figure 1C’).
The climate in the valley is cold-temperate, and the mean annual precipitation (3000 mm), falls mostly in winter (Gallopín, 1978). A thermal gradient is observed along the valley with annual temperatures declining from 7.6°C at Lago Mascardi, to 4.3°C at the 1411 m elevation (http://www.cricyt.edu.ar).
The studied area lies within a high diversity section of the Sub-Antarctic forest (Figure 1C’). The vegetation is arranged in altitudinal strata along the valley (Correa Luna and Dimitri, 1969; Martín and Mermoz, 2005). The modern distribution and floristic composition of these communities provide analogues to facilitate the interpretation of the paleovegetation records.
Methodology
Modern pollen studies
An altitudinal transect of pollen samples along the Río Manso Superior valley was carried out in 2004 to study the modern pollen–vegetation relationships and for a better interpretation of the changes in the pollen record of Lago Mascardi.
A total of 28 soil surface/moss pollen samples were collected along this altitudinal transect from Lago Mascardi (793 m a.s.l.) to Refugio Mailing (2000 m a.s.l.) (Figure 1C’). Standard procedures were applied (Bennett and Willis, 2001).
Lycopodium spores were added as markers. Pollen Atlas and reference collections were used for identification (D’Antoni and Markgraf, 1978; Heusser, 1971). A total of 91 pollen taxa were grouped in ecological units to facilitate interpretation (Table 1). Data were transformed to percents and pollen concentration values. Percent values were analyzed applying CONISS constrained cluster analysis, using Cavalli–Sforza’s chord distance and Edward dissimilarity coefficient. The output dendrogram was used for the zonation of the pollen record. Statistical analyses were performed with TILIA 2.0 and TGVIEW 2.02. (Grimm, 1993, 2004).
Pollen types identified in modern and core PMAS 94-1 samples, arranged according to vegetation units

Only found in core samples.
Palynological richness (E (Tn)) was estimated by rarefaction analysis (Birks and Line, 1992) and carried out using PSIMPOLL 4.10 (Bennett, 2003).
Core PMAS 94-1
Sediment cores were retrieved from Lago Mascardi during summer 1993/1994 within the framework of an Argentinean–Swiss Scientific collaboration programme (e.g. Ariztegui et al., 1997; Bianchi, 1999). Twin piston cores, PMAS93.4 and PMAS94.1, were collected 1 m apart from a water depth of 30 m at the southern coast of the lake containing a continuous record of environmental changes extending back to 15 000 14C yr BP (Figure 1B).
Both cores display almost identical stratigraphies as shown in high-resolution seismic profiles (Ariztegui et al., 2001, 2008). They can be easily correlated using magnetic susceptibility profiles which reflect conspicuous lithological changes and the presence of tephras associated with the significant volcanic activity of the region (Figure 2). Two main lithological units have been distinguished. The oldest sediments comprise a laminated sequence with variable development in quality and thickness of the lamination as well as amount and nature of the organic fraction (Ariztegui et al., 2001; Hajdas et al., 2003). The uppermost Holocene unit is mostly composed of fine siliciclastic sediments interbedded with numerous tephra layers and a variable content of extremely well-preserved remains of the subaquatic moss Rhynchostegium (Ariztegui et al., 1997).
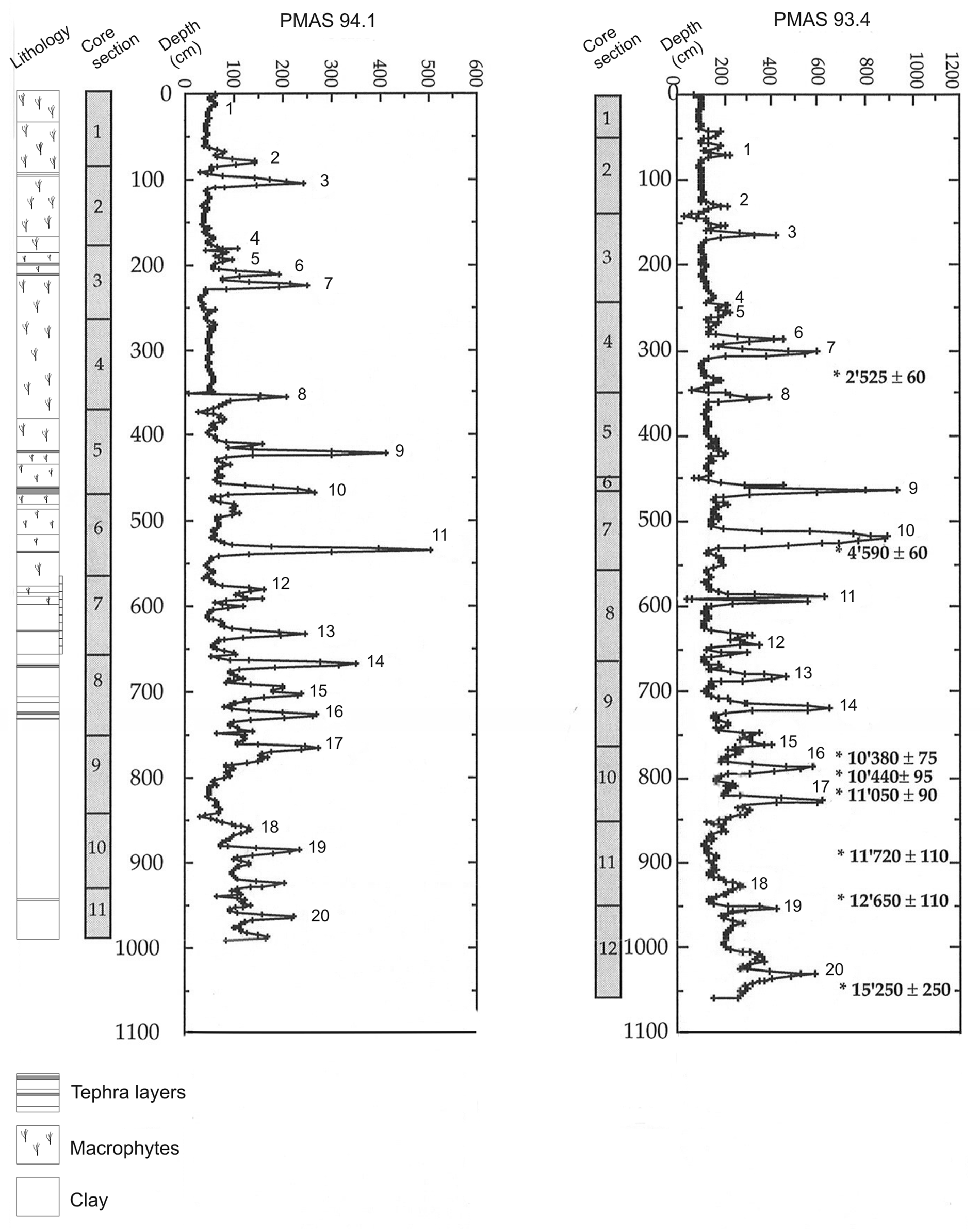
Lithology of core PMAS 94.1. Magnetic susceptibility profiles of sedimentary cores PMAS 93-4 and 94-1. Numbers indicate the position of the tie points used for core correlation
A high-resolution pollen record has been produced in core PMAS94.1 that has been dated interpolating a robust age model developed for core PMAS93.4 (see below).
Chronology
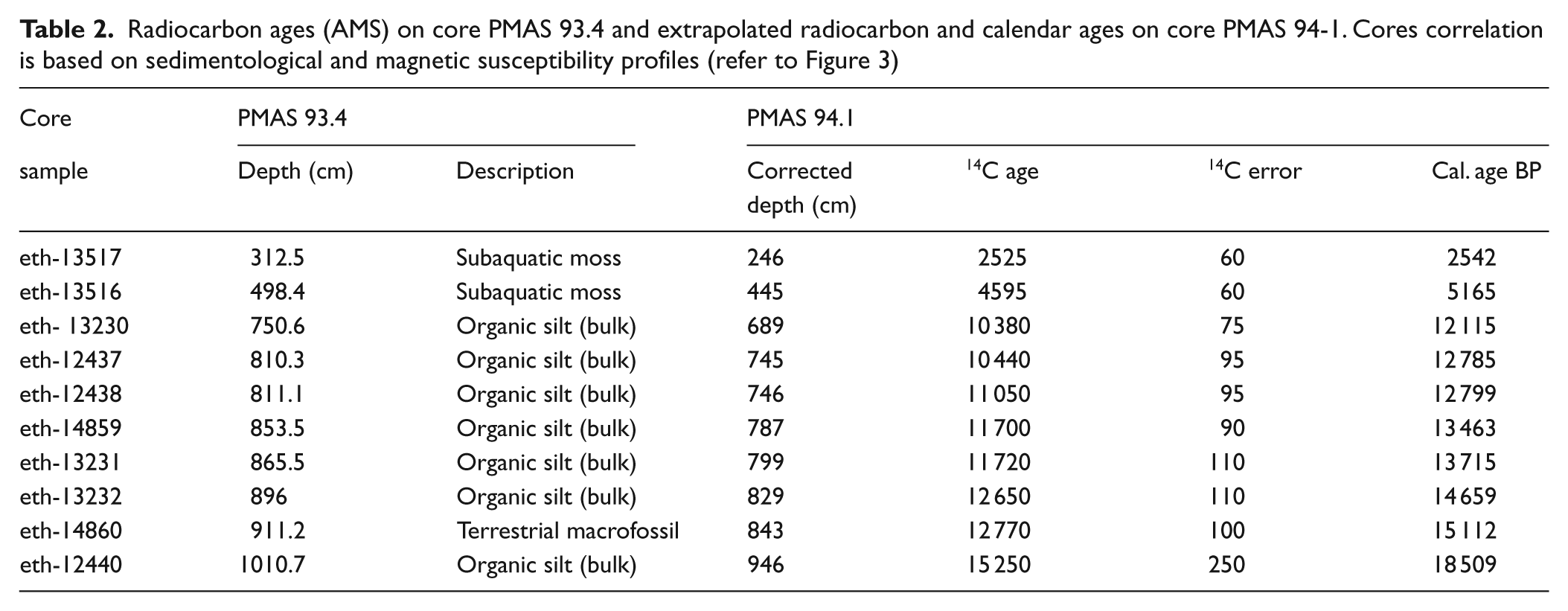
Lago Mascardi chronology was produced by interpolation of ten AMS radiocarbon ages obtained from terrestrial macrofossils, subaquatic mosses and bulk sediment samples on core PMAS93.4 (Ariztegui et al., 1997; Hajdas et al., 2003). Radiocarbon ages were calibrated using the program CALIB 6.0.1 (Stuiver et al., 1993, 1986–2010). Calibration was performed with the Southern Hemisphere curve (SHCal04) (McCormac et al., 2004) for samples < 10 000 14C yr BP (2525±60 14C yr BP; 4595 ±60 14C yr BP). Not being reliable or possible to use this curve for older samples, the remaining ages were calibrated with the Northern Hemisphere curve (IntCal04) (Reimer et al., 2004).
Table 2 contains a detailed list of the radiocarbon data. These ages were assigned to the appropriated depths of the pollen profile by comparing magnetic susceptibility and lithological profiles which allowed defining tie points, corresponding to ash layers, between cores. A constant sedimentation rate between those points was assumed for the purpose of developing the age model. The age–depth model was obtained by MCAge software using a smoothing spline (0.2 stiffness coefficient) and a bootstrap approach (Monte Carlo sampling) (Higuera, 2008; Higuera et al., 2009). The confidence intervals for the age–depth models, reflecting the combined uncertainty of all age estimates, were derived from 1000 bootstrapped chronologies. For each bootstrapped chronology, each age used to develop the chronology was selected randomly based on the probable distribution of calibrated 14C dates. The final chronology represents the median age at each depth from all the runs (Figure 3).
Radiocarbon ages (AMS) on core PMAS 93.4 and extrapolated radiocarbon and calendar ages on core PMAS 94-1. Cores correlation is based on sedimentological and magnetic susceptibility profiles (refer to Figure 3)
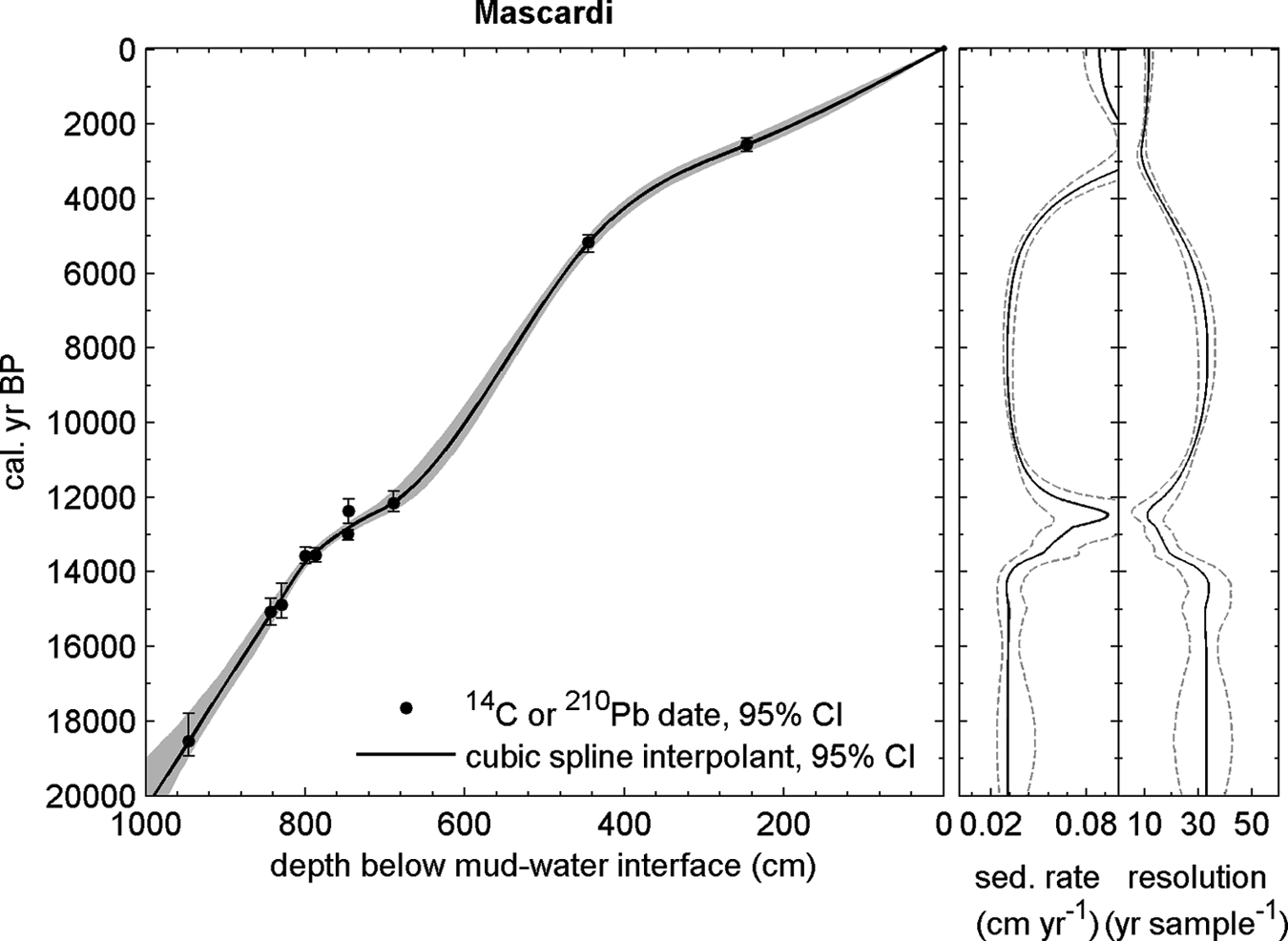
Age versus depth model of Lake Mascardi PMAS 94-1 core record. Sedimentation rates were assumed to be constant between tie points
Core PMAS 94-1 pollen studies
A total of 99 sediment samples were collected along the core and prepared for pollen analyses using standard procedures (Bennett and Willis, 2001). Raw data were transformed to percents excluding aquatic types. TILIA 2.20 was used for analyses C2 was used for graphics (Juggins, 2003). Pollen accumulation rates (PAR grains cm2/yr) were calculated by dividing the pollen concentration values by the temporal resolution provided by the age model. Palynological richness (E (T n )) was estimated by rarefaction analysis (Birks and Line, 1992) and carried out using PSIMPOLL 4.10 (Bennett, 2003). Percent values were analyzed applying CONISS constrained cluster analysis, using Cavalli–Sforza’s chord distance and Edward dissimilarity coefficient. The output dendrogram was used for the zonation of the pollen record (Grimm, 1993, 2004) (see Figure 5).
Results
Upper Manso River transect
Several vegetation units can be broadly depicted using variations in both percentage and concentration of pollen samples along the altitudinal transect (Figure 1C’; Figure 4). They are as follows.
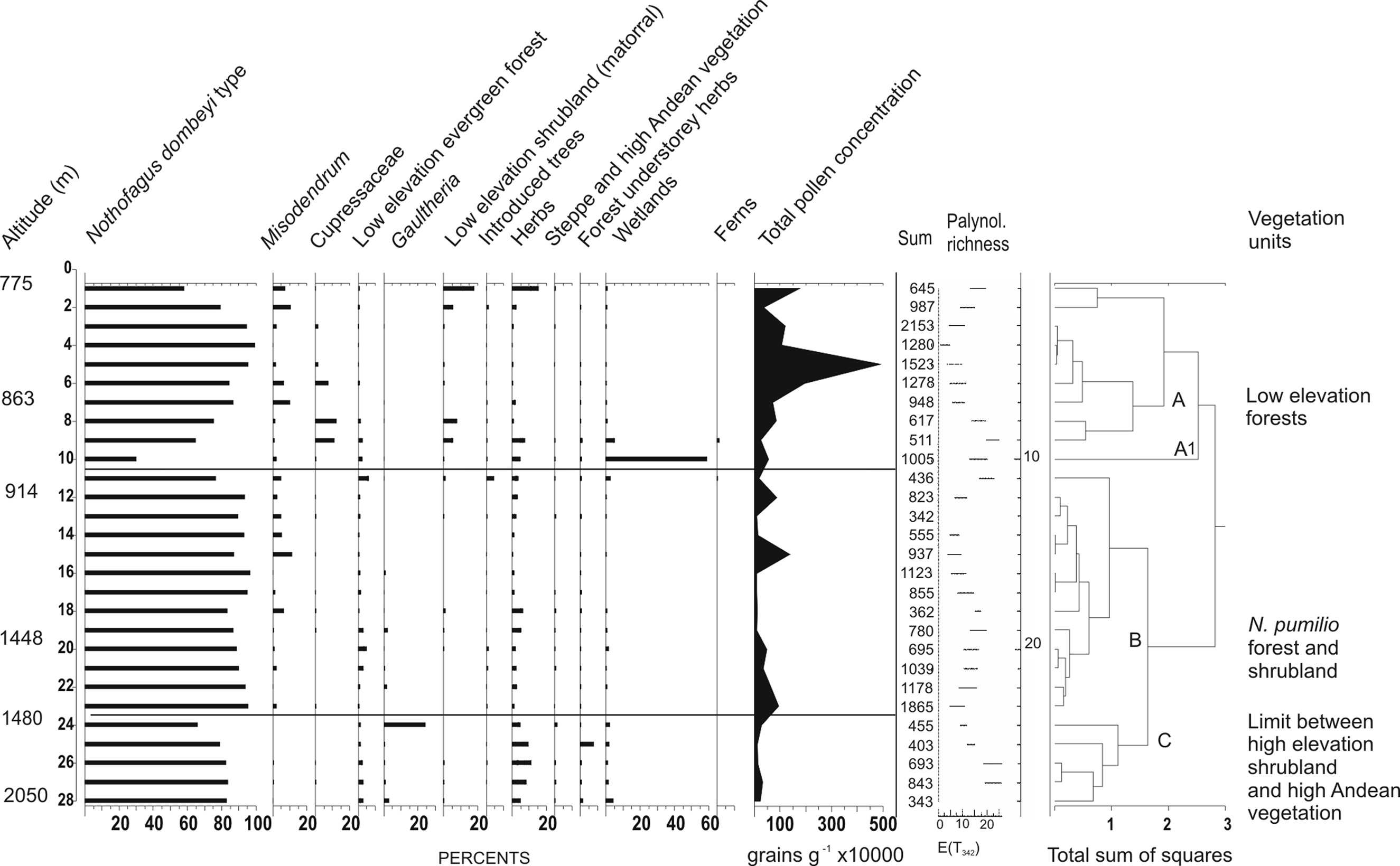
Percentage, total pollen concentration values (grains/g), and palynological richness of modern pollen samples along a transect in the Upper Manso River basin. Zonation based on results of constrained cluster analysis
Low elevation Nothofagus antarctica forest and xeric shrubland ‘matorral’ (modern pollen samples 1, 775 m; 2, 776 m and 7, 863 m elevation)
This unit develops at valley bottoms on poor soils in relation to disturbance processes. It is the most affected by human activities. Frequent elements co-occurring with Nothofagus antarctica are Diostea juncea, Discaria chacaye, and Lomatia hirsuta.
Values of Nothofagus dombeyi-type fluctuated from 65 to 93% of the pollen sum. Schinus (11%), Verbenaceae (6.4%), and Poaceae (11%) attained their maxima, while Misodendrum reached 10%.
Austrocedrus chilensis forest (modern samples 5, 824 m; 6, 825 m; 8, 876 m; 9, 875 m)
This unit includes open and closed canopy forests that extend on slopes below 1100 m a.s.l. This xerophytic conifer co-occurs with N. antarctica and Maytenus boaria at the easternmost locations of the valley while to the west it forms a mixed forest with N. dombeyi.The highest values of Cupressaceae (12–11%) were found at this unit and at mixed N. dombeyi-Austrocedrus chilensis forest (7%). Lomatia (4%) and Maytenus (2%) are best represented at the eastern most section of the A. chilensis forest where the forest is intermixed with shrubland.
Nothofagus dombeyi forest (modern samples 3, 793 m; 4, 792 m; 12, 914 m; 13, 920 m; 14, 930 m; 15, 931 m)
Located at low elevation slopes (1000–1100 m a.s.l.), the highest arboreal stratum is dominated by individuals of Nothofagus dombeyi. In wet spots evergreen trees such as Fitzroya cupressoides form patches (see Figure 1C’), while Saxegothaea conspicua and Podocarpus nubigenus grow as isolated individuals. Tall canes of Chusquea culeou form dense thickets. Hydrangea intergerrima and Gaultheria mucronata are frequent species in the understorey. Misodendrum reached 11% at the highest elevation of this unit, from there decreasing with altitude.
Nothofagus pumilio forest (modern samples 16, 1000 m; 17, 1001 m; 18, 1250; 19, 1300; 20, 1448 m)
This unit is characterized by a closed canopy stratum. It extends in mountain slopes at elevations between 1000 and 1500 m a.s.l. Shrubs such as Drimys winteri, Berberis sp. and Maytenus dysticha can be found at the understorey. At its lowest elevations the evergreen N. dombeyi co-dominates, forming a mixed forest.
Nothofagus pumilio shrubland (modern samples 21, 1449 m; 22, 1477 m; 23, 1478 m; 24, 1480 m; 25, 1670 m)
It is a narrow layer at 1400–1700 m, between the N. pumilio forest and the high Andean vegetation patches. It is formed by a dense stratum of shrubs of Escallonia alpina and Empetrum rubrum is common in the understorey. Values of Podocarpus (3%) and Drimys winteri (2%) were the highest in the forest while maximum values of Gaultheria (24%) Valeriana (8%) Apiaceae (5%) Papilionatae (3%) were found at the limit between forest and shrubland.
High Andean vegetation (modern samples 26, 1676 m; 27, 1800 m; 28, 2050 m)
It is a discontinuous unit developing at the summits above 1700 m a.s.l. It is formed by a stratum of xerophytic grasses and cushion plants such as Festuca monticola and Nassauvia sp., with a low vegetation cover. Asteraceae (3%) attained its maximum; Cyperaceae reached 4% and Gaultheria 3%.
Sedge bogs ‘mallines’ (modern samples 10, 912 m; 11, 913 m)
Waterlogged soils are frequent in flat areas at different altitudes showing diverse vegetation in which species of Cypereace, Poaceae and Juncaginaceae are dominant. Escallonia virgata and Gunnera magellanica are also frequent. Escallonia (5%) is almost restricted to this unit. Cyperaceae was not excluded from the pollen sum and attained its maximum (59%). Pinaceae reached 4.6%, indicating the proximity of planted trees. Values of Nothofagus ranged from 30% at the sedge bog to 99% at N. dombeyi forest, and reached 83% at the high Andean patches, suggesting that this taxon is over-represented above the treeline. Pollen of Cupressaceae was restricted to vegetation units where the tree is present; indicating its dispersion along the valley is poor. The same is valid for the low elevation shrubland dominant elements.
Values of Gaultheria and ‘forest understorey herbs’ were higher at the N. pumilio forest and shrubland. The patchy distribution of shrubland and stronger winds at higher altitudes favour pollen dispersion.
Palynological richness E (T 342) was the highest at the limit between N. pumilio shrubland and high Andean vegetation. Values were also high at the sedge bog and at the limit between the low elevation mixed forest and shrubland. The lowest values of this index were registered at low and high elevation Nothofagus dombeyi forests.
Total pollen concentrations were the highest at low elevation forests, particularly at mixed Nothofagus-Austrocedrus, related to a high pollen production (Bianchi and Olabuenaga, 2006). The lowest values were recorded at the limit between low and high elevation forests where dense foliage restricts dispersal. Pollen concentrations were also low at the high Andean vegetation unit.
Vegetation units were further grouped by constrained cluster analysis. All types of low elevation forests and woodland, from altitude 775 to 876 m were gathered in cluster A. The sedge bog sample (912 m) corresponds to A1. Samples from mixed and pure N. pumilio forests and the lower limit between N pumilio forest and shrubland from altitude 913 to 1478 m were reunited in Cluster B. Samples of high elevation N. pumilio shurbland and high Andean vegetation (1480–2050 m) were gathered in Cluster C.
Pollen diagrams of core PMAS 94.1
Seven pollen zones were identified through the core (Figure 5).
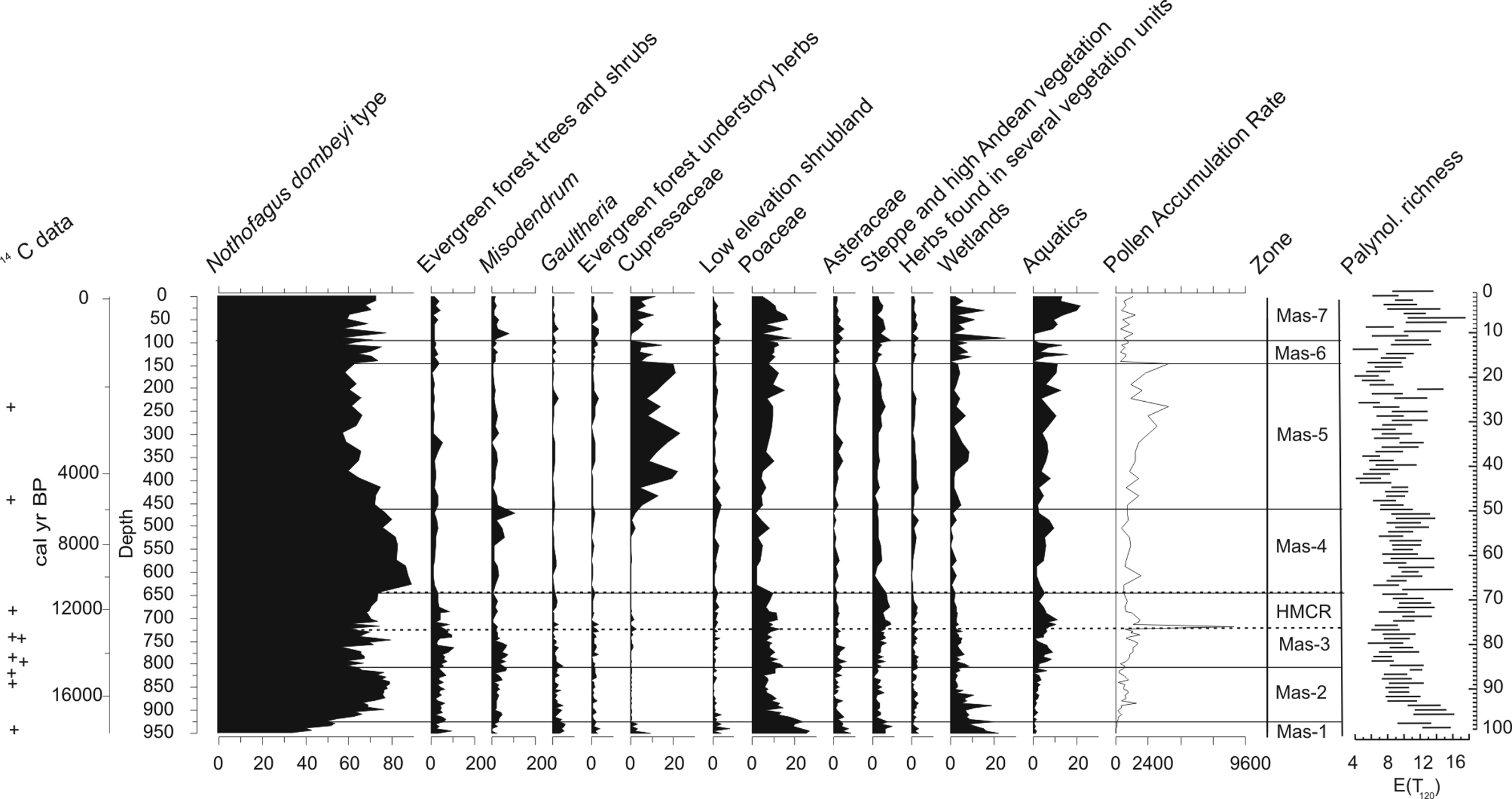
Percentage, total pollen accumulation rates (PAR), and palynological richness values of core PMAS 94-1 samples. Zonation is based on results of constrained cluster analysis. The dotted line indicates the Huelmo–Mascardi cold reversal. + indicates position of 14C dates
Zone Mas-1 (950.2–927.7 cm depth; 18 643–17 889 cal. BP)
At Zone Mas-1 (950.2–927.7 cm depth; 18 643–17 889 cal. BP) the percentage values of all categories of herbaceous types, including wetland taxa (mean 15%) reach high values. Poaceae (mean 23%), ‘low elevation shrubland’ types (mean 3%) and Gaultheria (mean 5%) attain their highest values at the bottom of this zone, showing a decreasing trend. Nothofagus exhibits the lowest values of the whole section (35–51%), while ‘low elevation evergreen trees and shrubs’ taxa show significant values 10%). Cupressaceae, probably Fitzroya cupressoides, present at the bottom (mean 4%) decreases towards the top.
Palynological richness values, E (T 120) are high reaching 16 taxa at the bottom. PAR values are very low (mean < 100 grains cm2/yr). Palynological richness and pollen concentration values are similar to those recorded at the limit between N. pumilio shrubland and high Andean semi-desert vegetation units.
Zone Mas-2 (927.7–800.8 cm depth; c. 17 889–13 754 cal. BP)
A sharp increase in Nothofagus (maximum 79%) is observed in this zone with a declination at the top (60%). Misodendrum (mean 3%) remains low. A decline is observed in Poaceae (mean value 11%), Asteraceae and ‘steppe and high Andean herbs’ taxa. E (T 120) values are low to medium. PAR rises from <100 to 546 grains/cm2 per yr.
Zone Mas-3 (800.8–635.6 cm depth; c. 13 754–11 007 cal. BP)
At Zone Mas-3 Nothofagus pollen fluctuates and declines to 57% while Gaultheria and Misodendrum show a decreasing trend. ‘Steppe and high Andean herbs’ as well as ‘evergreen forest trees and shrubs’ increase to reach their highest values. Poaceae continues to diminish, showing a mean value of 9%. Total pollen accumulation rate PAR (<400 to <8000 grains/cm2 per yr) shows a sharp increase at the middle of this section. E (T 120) values are medium to high.
Zone Mas-4 (635.6–462 cm depth; c. 11 007–5609 cal. BP)
This zone features the decline of Poaceae, falling to its lowest values (mean 3.5 %) plus a prominent increase in Nothofagus that reaches a maximum of 90% at the bottom, with a decreasing trend towards the top of the section. ‘Low elevation evergreen forest’ taxa and Cupressaceae show low values. ‘Steppe and high Andean vegetation’ taxa are low at the bottom, showing an increasing trend. Palynological richness values, E (T 120) oscillates between 6 and 9.
Zone Mas-5 (462–143 cm depth; c. 5609–1538 cal. BP)
This zone displays a rapid increase of Cupressaceae to attain its highest values (mean 14%). Nothofagus and Misodendrum decline abruptly at its expense. Poaceae increases. PAR reaches a mean of 2000 grains/cm2 per yr.
Zone Mas-6 (143–93.6 cm depth; c. 1538–1010 cal. BP)
At Zone Mas-6 Nothofagus recovers while Cupressaceae fluctuates. ‘Evergreen forest trees and shrubs’ as well as ‘forest understorey herbs’ increase slightly. ‘Steppe and high Andean semi-desert’ taxa increase slightly. Palynological richness values are low to medium and PAR values are low (<650 grains/cm2 per yr).
Zone Mas-7 (93.5–0.1 cm depth; 1010 cal. BP–recent)
At Zone Mas-7 the values of Nothofagus and Cupressaceae fluctuate. All herb taxa increase. An abrupt growth of the aquatic fern Isoetes (21%) at the bottom accompanies a sharp rise in wetland taxa. Aquatics (mean 11%) attain their maxima in this zone. Palynological richness values are medium to high. PAR shows low to medium values (between <300 and <1400 grains/cm2 per yr). Exotic pollen taxa, Pinus and Betula appear in the record and Asteraceae fenestrate increases. An expansion in wetland herbs and aquatics is observed.
Discussion
The comparison with modern pollen samples (this paper; Markgraf et al., 2002; Páez et al., 2001) has allowed the reconstruction of the long-term vegetation history in the watershed of Lago Mascardi and some aspects of climate trends and variability were inferred.
Lateglacial–Holocene transition
The Mas-1 pollen assemblage shows the decline of open vegetation with steppe and high Andean vegetation elements, as well as extended bogs with patches of forest, probably deciduous. At a regional scale, before c. 18 000 cal. BP, this kind of vegetation mosaic dominated the landscape. All vegetation units recorded along the altitudinal transect in Río Manso Superior valley were represented in this zone, suggesting that the entire vegetation gradient from evergreen forest to steppe was more constrained than in recent times. This prediction was formerly proposed for pollen localities in Chile at the same latitude (37°–42°S) for the Glacial period (Villagrán and Armesto, 1993).
The decline of Cupressaceae that occurred after c. 18 600 cal. BP may have followed a local expansion of Fitzroya cupressoides from neighbouring valleys favoured by regionally extended open landscapes and waterlogged soils during deglaciation. To the west, along the Chilean coast, Lateglacial pollen sequences indicate the dominance of Nothofagus dombeyi type and substantial amounts of non-arboreal pollen (Poaceae, Asteraceae and Cypereceae) during this period (Villagrán, 1985, 1990).
At Mallín Book, the expansion of steppe into Andean valleys is inferred (Markgraf, 1983). However, based on our findings we propose an alternative interpretation for the valley of Río Manso Superior. Before starting to decline at c. 18 000 cal. BP, the high Andean vegetation, sharing several taxa with steppe, still occupied extended areas in the Andean glacial valleys subsequently to its expansion during the Glacial period at the western Andean flanks.
Steppe, extended wetlands and patches of humid forest prevailed at pollen sites located to the east (Jackson 1996; Markgraf, 1984; Whitlock et al., 2006).
The fact that there was only open vegetation in the region suggests a drier and probably colder climate than today.
During c. 17 900–13 700 cal. BP increasing temperatures and wet conditions resulted from rapid deglaciation, promoted a fast expansion of the forest from protected valleys in the cordillera. Mas-2 pollen assemblage suggests a fast expansion of a dense-canopy forest by this time. During the Glacial period, deciduous species of Nothofagus were dominant along the altitudinal gradient at the western Andean flank, supported by pronounced seasonality (Villagrán and Armesto, 1993). Similarly, at the foothills of the eastern Andean flank, these species may have been the dominant tree elements (Markgraf and Bianchi, 1999; Whitlock et al., 2006). With the increasing temperature during LG-H, N. pumilio may have expanded uphill from lowland localities to reach the altitudinal position that it occupies today. In the case of N. antarctica, its tolerance to poor soils and its adaptation to fire could have allowed its eastward expansion where it coexists today intermingled with shrubland taxa.
Between c. 13 700 and 11 000 cal. BP forest development in the Río Manso valley was interrupted by a brief period of forest decrease and expansion of steppe and high Andean herbs, including Poaceae, as a consequence of climatic changes. Trends in biogeochemical indicators in sediments were taken as clear evidence of a re-advance of El Manso Glacier between 13 000 and 11 500 cal. yr BP (11 400–10 200 14C yr BP) under increasing moisture and decreasing temperature at the waterheads (Ariztegui et al., 1997). Mas-3 suggests a period in which the forest became more open. This fast and sharp decline in the forest dominant species started approximately 700 years before the onset of the Huelmo-Mascardi cold reversal, at c. 13 000 cal. BP and continued until its termination at c. 11 500 cal. BP. This change in vegetation was definitively related to climate. Hence, a conclusive explanation of the asynchronous response of the sedimentary and pollen records to cooling is not yet available. However, similar asynchronous responses have been recorded elsewhere during Holocene cold episodes. At Laguna Las Vizcachas (50°42'S, 71°59'W) pollen and diatom records preceded the beginning of the ‘Little Ice Age’ as recorded by glacier fluctuations in Southern Patagonian Icefield (Flavia Quintana, personal communication, 2010).
In the Chilean Lake district 41°S, the North Patagonian Rain Forest (NPRF) dominated by Nothofagus, P. nubigena and conifers, developed between about 15 200 and 14 300 cal. BP. Palynological studies on the Huelmo site showed the appearance and subsequent increase of pollen of Podocarpus nubigena at c. 14 400 cal. BP. This trend was interpreted as a downvalley shift of mixed NPRF communities, suggesting a lowering of vegetation belts in response to cooler conditions (Moreno et al., 2001).
The opening of the forest after its expansion during the LG-H transition is more evident in Lake Mascardi than in other localities to the east. In Lago Escondido, a forest opening of smaller dimensions in magnitude was observed by this time (Jackson, 1996). At Lago El Trébol the vegetation change observed between 12 000 and 9000 cal. BP was taken as evidence of the eastward forest expansion, at expense of xerophytic shrubland and steppe (Whitlock et al., 2006). In relative terms, the similarity between the pollen values of this time period with the values recorded in soil samples at the limit between N. pumilio forest and shrubland may indicate that deciduous species of Nothofagus were occupying lower altitudes from where they expanded uphill (N. pumilio) and eastward (N. antarctica).
The increase in steppe and high Andean taxa during the HMCR reinforce the idea that some species have moved down along the altitudinal gradient during cold periods. A lower location of vegetation units during the Glacial Maximum and the HMCR has been previously proposed for the Isla de Chiloé and the Lake District in Chile, respectively (Hajdas et al., 2003; Moreno et al., 2001; Villagrán, 1990).
Unfortunately, at the eastern Andean flank, three main factors prevent testing the hypothesis of the lowering of vegetation units during cold periods by means of pollen analysis: (1) Pollen grains from the dominant species of evergreen N. dombeyi, and deciduous forest N. pumilio and N. antarctica cannot be distinguished from each other under optical microscope. (2) Pollen grains from higrophylous (Fitzroya/Pilgerodendron) and xerophytic trees (Austrocedrus chilensis) in the Cupressaceae cannot be distinguished from each other using an optical microscope for counting (Jackson, 1996; Markgraf, 1984; Whitlock et al., 2006). (3) Most of the pollen taxa that characterize the high Andean vegetation unit are also found at low elevations in the steppe. Because of these limitations, inferences concerning the dominance of deciduous species over evergreen species of Nothofagus and hygrophilous versus xerophytic species in the Cupressaceae family remain as predictions. The increase of evergreen forest trees and shrubs during the Lateglacial–Holocene transition could be also interpreted as the contribution of long distance pollen sources. Recent findings showed that pollen transport over the Andes due to the westerlies is significant and that pollen trajectories follow glacial valleys as transport corridors from west to east (Pérez et al., 2009).
Early and middle Holocene
The features observed at Mas-4 may be indicating that the forest became very dense. The comparison of trends in palynological richness and number of taxa with those of soil samples along the altitudinal transect led us to think of the expansion of an evergreen forest dominated by N. dombeyi. During the early and middle Holocene (c. 11 007–5600 cal. BP) the evergreen species N. dombeyi may have probably contributed to the expansion of Nothofagus. Evergreen species of Nothofagus spent glacial periods in lowland elevation sites in the Chilean coast (Villagrán and Armesto, 2003) where these taxa were a major component of low and high elevation forests. However, the fast and synchronous increase of this taxon recorded at both sides of the Andes during LG-H allow us to support the hypothesis that protected valleys in the main cordillera may have been suitable forest refuges from where some species expanded eastward with the amelioration of climatic conditions (Bianchi, 1999; Markgraf et al. 1995; Pastorino and Gallo, 2002). In the Lake District, forest diversification after c. 11 000 cal. BP included the expansion of thermophilous rainforest species and the disappearance of Podocarpus. The character of the vegetation change at this time suggests a warming that led to interglacial climate conditions. Fire and vegetation disturbance were also prominent features between c. 15 000 and 11 100 cal. BP in some localities of this area (Moreno et al., 2001).
The pollen assemblage of Mas-5 indicates a major vegetation change. The Nothofagus forest is replaced by a closed canopy mixed forest with the co-dominance of Austrocedrus.
During the middle Holocene, a substantial change in forest composition at Lago Mascardi occurred. With the expansion of Austrocedrus, at c. 5400 cal. BP, the old Nothofagus forest was replaced by a mixed Nothofagus-Austrocedrus forest with a closed-canopy.
To the west of LM, in Isla Grande de Chiloé, 43°S, a mosaic of Valdivian/North Patagonian rainforest species started at c. 6000 cal. yr BP, along with a moderate increase in fire activity (Abarzúa and Moreno, 2008).
To the east of LM, about c. 6000–3500 cal. BP Austrocedrus chilensis expanded abruptly. At wet localities, such as Lago Mascardi, Mallín Book, and Lago El Trébol this expansion occurred to the detriment of Nothofagus (Bianchi, 1999; Markgraf, 1983; Whitlock et al., 2006). To the southeast, A. chilensis extended on land formerly occupied by shrubs about 2000 years later, and continued up to the present into the steppe. An increase in climate variability favouring fire occurrence has been inferred from independent data in both palynological sites (Whitlock et al., 2006). Unfortunately, a fire history record of the Lago Mascardi area is not yet available.
Late Holocene
Zone MAS-6 indicates that a more dense forest with evergreen species was developed in the Lago Mascardi watershed at c.1500 cal. BP, and paleoclimatic conditions similar to modern ones were established. Changes in the last two millennia including a decrease in Austrocedrus and an increase in Nothofagus in both wet and dry localities on the NHNP, suggest wetter conditions than before (Jackson, 1996; Markgraf, 1983; Markgraf and Bianchi, 1999).
Glaciological and sedimentological studies in this watershed and adjacent valleys provide additional information on climate fluctuations and their forcing mechanisms.
Tree ring-based glaciological research in The Río Manso Superior valley has shown that the Río Manso Glacier has reacted sensitively to climate change during the ‘Little Ice Age’ (LIA), with well-identified major glacial advances between
Investigations in annually laminated sediments of proglacial Lago Frías, located at a neighboring valley, reflect variations in the transport of glacially derived clay and silt to the basin that can be directly linked to changes in climate. Sedimentological evidence combined with a chronological model indicate variations in varve thicknesses showing two frequencies centered at 16.4 and 10.5 years and a main frequency located between 2.5 and 3.0 years pointing towards a dominant El Niño/Southern Oscillation (ENSO) signal (Ariztegui et al., 2007).
Regionally the establishment of more closed forest is related to increased precipitation that allowed the forest to expand eastward into the land previously occupied by woodland and steppe (Whitlock et al., 2006). A possible explanation for this regional increase in precipitation, especially during summer months, may be related to the onset of ENSO variability (Markgraf and Bianchi, 1999).
Intensified fire activity in Isla Grande de Chiloé, 43°S, at 3000 cal. BP has also been associated with the increase in climate variability generated by ENSO (Abarzúa and Moreno, 2008; Abarzúa et al., 2004).
Recent times
In terms of vegetation dynamics, MAS-7 (1010 cal. BP–present) may imply a period in which the forest becomes more open. The effect of human impact on vegetation becomes noticeable at the top of this zone. Exotic pollen taxa, Pinus and Betula appear in the record and Asteraceae fenestrate increases, suggesting the introduction of weeds of the Asteraceae family. The expansion in wetland herbs and aquatics could be the combined effect of mild climatic conditions and the anthropogenic effect on waterlogged soils and bogs (mallines) due to the introduction of cattle. Regionally land use increased first by Native Americans and subsequently by European settlers (Markgraf, 1983; Veblen and Markgraf, 1988). Part of the observed fluctuations, however, can be partially explained by climate variability at a decadal scale. Recent short-term climatic events could not be precisely tracked in the pollen records analyzed in this paper because of the sampling resolution. Tree ring-based glaciological studies in the region of Cerro Tronador, however, have contributed detailed evidence of high frequency climate variability. Several short periods of glacier readvance or stasis has been identified at the Río Manso Superior valley for the periods since mid-1870s. These pulses of glacier advance have been linked to multiyear periods of overall cooler and wetter conditions in the region. Tropospheric warming in conjunction with a significant decrease in precipitation, has probably contributed to the generalized glacial recession observed in most glaciers of Cerro Tronador during the last three decades (Masiokas et al., 2009b).
Conclusions
All pollen records from mid latitudes on the eastern Andean flank, including Lago Mascardi show the expansion of a humid Nothofagus forest during LG-H transition. This forest was best developed between 13 000 and 5000 cal. BP.
The amelioration of the climatic conditions during LG-H, the altitudinal and longitudinal shifts of vegetation units, and plant migration from glacial refugia that occurred after deglaciation may have been dominant factors controlling vegetation change in the Río Manso Superior valley and Lago Mascardi watershed. However, because of the methodological limitations mentioned above, these assumptions remain as predictions.
An increase in steppe and high Andean semi-desert herbs, as well as Poaceae, at expenses of Nothofagus after 13 700 cal. BP was initiated earlier than the major changes in the sedimentological record attributed to the HMCR. The position and intensity of westerly winds have been most probably responsible for the observed paleoenvironmental and vegetational changes in the Río Manso Superior basin. These changes also have been identified at pollen sites to the west in south central Chile. With the exception of the HMCR, major vegetation changes found in Lago Mascardi also have been tracked at the easternmost localities of the NHNP, today occupied by the mixed forest and the forest-steppe ecotone (Bianchi, 1999; Jackson, 1996; Markgraf, 1984; Whitlock et al., 2006).
Open forests, probably deciduous, dominated the landscape during LG-H while dense evergreen forest has become dominant since the early Holocene.
The dramatic change in forest structure generated by the expansion of Austrocedrus chilensis at c. 5400 cal. BP in Lago Mascardi has been registered at easternmost localities between 6000 and 2700 cal. BP (Whitlock et al., 2006).
The presently available palynological information is not enough for modeling the vegetation response to rapid climatic events that occurred in the deep Andean glacial valleys on the Argentine flank during the late Holocene.
New localities in neighbouring basins are being studied at present. A larger data network will allow us to answer questions on processes controlling vegetation dynamics at a regional scale (Virginia Iglesias, personal communication, 2010).
Because of the methodological limitations mentioned above, our findings cannot support the existence of a regionally lower treeline in the Andean valleys during the Glacial period and this assertation remains thus a prediction. We place our expectations in population genetics studies, now in progress in the region.
Ongoing sedimentology, geochemistry, dendro- and sclerochronology studies in northern Patagonian lakes are providing independent high resolution proxies which will contribute to explain processes influencing vegetation dynamics, to define climatic trends more precisely and to explain their forcing mechanisms more accurately.
Footnotes
Acknowledgements
Funding was provided by the Lagos Comahue project (CONICET 1992) and Swiss National Science Foundation (NF 21-37689.93). The senior author was supported by a Swedish Institute Fellowship to continue this research at the Geology Department, Uppsala University in 2004.Gonzalo Sottile is thanked for assisting with running software and Mauro Passalía for helping with graphics. We are also grateful to the anonymous reviewers for their valuable comments that helped to improve the manuscript.