
Abstract
Recent interpretations of the archaeology of domesticated rice evolution and the development of agriculture in the Lower Yangzi Valley are being informed by new genetic, palaeoenvironmental, archaeological and archaeobotanical data. This paper explores current views on early rice production in the region and attempts to expand the discourse to provoke exploration of new and modified questions. Rice-specific botanical through large-scale ecological issues and causality are examined. This paper argues that understanding rice domestication and production, although important, should not be the exclusive goal of research on early agriculture in the Yangzi Valley. Despite the preliminary nature of the investigations at Kuahuqiao (c. 8000–7000 cal. BP), evidence of resource management, and potential domesticated resources or their ancestors that includes pig (Sus scrofa), peach (Prunus persica), apricot (P. armeniaca), Japanese apricot (P. mume), foxnut (Euryale ferox) and water chestnut (Trapa spp.) underscore the need to broaden the scope of inquiry on early food/resource production. The complexities of the archaeological and environmental record in the Yangzi valley indicate that nuanced, complex explanations for rice domestication and agricultural origins rather than deterministic, single cause explanations are necessary in order to move forward.
Keywords
Introduction
In the nearly 15 years since Shen and I assessed the then-current status of our understanding of East Asian rice domestication and the spread of rice production in Northeast Asia (Crawford and Shen, 1998) considerable archaeological and genetic data have been amassed. The rice genome has been sequenced (Yu et al., 2002) and the archaeological record for early rice and its cultural and environmental context is rapidly developing (e.g. Crawford, 2006; Fuller and Qin, 2009; Fuller et al., 2010b; Jiang and Liu, 2006; Pan 2011; Zhao, 2010a; Zong et al., 2007, 2011 (this issue); ZPICRAZM, 2004). The literature on rice genetics and its relationship to crop improvement is specifically addressing domestication in order to understand how to improve the modern crop. Thirteen years ago a few lines of evidence (phytoliths, charred and uncharred rice remains) indicated that early Neolithic populations in China appear to have been using rice, but how early its use began, the nature of the rice populations at the time, and under what circumstances rice was coming into use were issues that needed careful scrutiny. We urged considered rather than hasty interpretations and that more AMS radiocarbon dates were required before a proper assessment could be made. We were also concerned about the casual use of criteria such as grain size and shape for identifying rice as either wild or domesticated (Crawford and Shen, 1998: 864). Few, if any, directly dated rice remains had been reported and we were responding in part to what was, in our view, inappropriate attempts to date the oldest domesticated rice to roughly 11 500 BP (Normile, 1997). Many of these issues are being evaluated today and some are actively and constructively debated (e.g. Fuller, 2007; Fuller et al., 2007; Jiang and Liu, 2006; Liu et al., 2007). The geological, climatic, anthropogenic, and cultural context in which rice was one of many resources supporting the burgeoning human presence on the evolving landscape of what is now central-eastern China are being documented and are, in part, the subject of this special issue of The Holocene. This paper does not provide a comprehensive overview of the archaeological record because others have already done so (see above). However, the discourse is somewhat more limited than it could be despite better data being available today and criteria for assessing domestication and agriculture being established. In an attempt to expand the discussion I raise interpretive complications and questions for further research. The implications of alternative taxonomies of rice, the wide range of domestication-related traits (DRTs) in rice rather than a narrow suite of two or three traits, new archaeobotanical techniques to recognize those traits, the lack of an Upper Palaeolithic record combined with the negligible number of early Holocene sites in the Lower Yangzi valley, and the intellectual focus that emphasizes rice are considered. These issues are also considered in the context of broader ecological and anthropological questions. This paper is a personal reflection on issues that I hope will stimulate further discussion beyond narrowly framed explanations. As such, I try to raise questions for further research rather than provide answers to the many perplexing questions surrounding rice domestication and early agriculture in the Yangzi valley.
Taxonomy and characteristics of domesticated rice
Rice (Oryza sativa) is a grass (Poaceae) whose relationship to other members of the genus has for a long time been ‘a matter of opinion’ (Vaughan et al., 2008a). In fact, the relationship among most domesticated plants and their close relatives has been problematic because of the varying ability of crops and their wild relatives to cross and produce fertile hybrids. Harlan and de Wet (1971) recognized that cultivated plant classification was intuited through idiosyncratic characteristics and that these plants tend to be overclassified (Harlan and de Wet, 1971: 20). Interpretation of archaeological rice remains must be cognizant of the classification schemes. Overclassifying modern rice may lead to overclassifying archaeological material and vice versa. The primary gene pool consists of what we normally conceptualize to be a species, that is, members of the primary gene pool can regularly cross and produce reasonably fertile offspring (Harlan and de Wet, 1971; Harlan et al., 1973). The domesticated and wild forms are, in this conceptualization, subspecies. In this respect East Asian rice is overclassified.
In the 1990s when Shen and I published our synopsis we summarized key issues related to the wild taxa most closely related to domesticated rice and indicated that cultigen rice and its wild ancestor(s) are considered to be the same species by some taxonomists (Crawford and Shen, 1998: 863). There are either two species ancestral to domesticated Asian rice, O. rufipogon and O. nivara, or one species, O. rufipogon. The Flora of China lists three native species: O. rufipogon, O. officinalis and O. meyeriana (Liu and Phillips, 2006). The latter two species have different genomes than O. rufipogon/nivara/sativa and their distributions are tropical although their historical distribution is not clear. O. nivara is annual and O. rufipogon is perennial; they are allotropic because of habitat adaptations but have no genetic barriers to crossing (Morishima, 2001: 60). The annual habit is a response to seasonally fluctuating water while the perennial habit is a response to deeper waters and fewer disturbances (Morishima, 2001: 68). Perennial rice, because it spreads vegetatively, has very low seed production and is outcrossing while annual rice produces abundant seeds and is mainly self-pollinating (Vaughan et al., 2008b). The distinct distributions of the two putative ancestral rice species reinforce the view that they are separate species. Morishima (2001) not only accepts that O. nivara and O. rufipogon are the same species (O. rufipogon) but that O. sativa and O. rufipogon are also the same species. The variation between annual and perennial types is continuous (Joshi et al., 2000: 1318; Vaughan et al., 2008b). Perennial rice increases seed production when habitats dry out. Conversely, the cultigen is still essentially a perennial and can survive several years if it is not harvested (Lu, Baorong, personal communication, 2011). Ratooning is also an alternative to double cropping in some regions (International Rice Research Institute (IRRI), 1988).
Further complicating matters is a form of rice, O sativa f. spontanea (weedy or red rice) that accompanies domesticated rice wherever it grows and nowhere else (Xia et al., 2011). Phenotypic characteristics of weedy rice resemble those of domesticated rice; flowering time, tillering, awn structure and hull characteristics are particularly variable in weedy rice (Gealy et al., 2009: 627). Research on weedy rice is directed to its eradication and is quite active because this plant can reduce yields of rice today by as much as 80% (Olsen et al., 2007: 812). Unlike wild rice, it has a tendency to self-pollinate (Xia et al., 2011) as do all annual rices. Weedy rice is also genetically closer to cultivated rice than wild rice (Cao et al., 2006) and it is generally agreed that weedy rice is a diverse taxon resulting from hybridization of wild and domesticated rice (Gealy et al., 2009; Olsen et al., 2007; Xia et al., 2011). The weedy rice in North American rice fields has been hypothesized to have arisen from incidental selection of feral crop volunteers but most evidence points to the ancestors of North American weedy rice being weedy hybrids introduced from Asia (Gealy et al., 2009; Olsen et al., 2007: 815). Because wild rice cross-pollinates easily, hybridization primarily (but not exclusively) results from introgression of genes from cultivated rice into wild rice populations (Xia et al., 2011: e16189). Distinguishing domesticated rice from wild and spontaneous types is not necessarily straightforward in the archaeological record. When and under what circumstances the first weedy rice appears is an archaeological issue well worth examining in relation to the evolution of domesticated rice. Finally, a weedy grass, Echinochloa crus-galli var. oryzicola has coevolved with rice as a mimic and is widely distributed with cultivated rice (Barrett, 1983). The seed is distinct from seeds of other varieties of E. crus-galli and examining the archaeological record for its evolution will also be useful in understanding the evolution and spread of rice (e.g. D’Andrea, 1995).
A variety of DRTs comprise the domestication syndrome in rice; most are related to the predictability and yield of grain production (Table 1). These traits are, for the most part, quantitatively expressed (Sweeney and McCouch, 2007) and tend to be retained once established because domesticated rice has a significantly higher rate of self-pollination than does wild rice or weedy rice (O. sativa f. spontanea). Table 1 lists most of the commonly reported traits in temperate Japonica (with a few from Indica) in order to illustrate the complexity of the evolution of this cultigen. Several traits form domestication blocks; most are on five chromosomes (Xiong et al., 1999: 250). For example, the genes for red pericarp, shattering and seed dormancy are associated on chromosome 7 while increased panicle branching and reduced tillering are on chromosome 8 (Ji et al., 2006; Miura et al., 2010). These linkages could help identify proxies for traits that might not be detectable in the archaeological record yet.
Comparison of key traits of wild and domesticated rice (with emphasis on Japonica in China)

Domestication-related traits or DRTs.
Shattering insures seed dispersal in the wild whereas reduced shattering in the crop reduces the number of seeds that are lost, maximizing human seed harvest potential. This is the classic trait used to discriminate wild from domesticated grasses (Fuller and Allaby, 2009; Fuller et al., 2010a: 14). Yet shattering in rice is not simply turned on or off. It results from the growth of one or two layers of cells (the abscission layer) that separates the grain from its pedicel. The layer is complete (continuous) in wild rice but still present although incomplete (discontinuous) in most domesticated rice (Li et al., 2006). Longitudinal sections do not show the abscission layer cutting the vascular bundle in either wild or domesticated rice (Li et al., 2006). A small-celled abscission layer still develops in Japonica rice, but in completely non-shattering varieties it is not functional (Ji et al., 2006). The abscission layer develops in degrees and in a variety of ways in all rice and abscission layer morphology is variable in modern rice varieties and is related to the degree of shattering (Ji et al., 2006: 1002). Some varieties are completely non-shattering while other varieties have moderate to easy shattering (Ji et al., 2006: 1002). The abscission layer is less continuous in varieties with low or no shattering spikelets and, conversely, more continuous in rice that has moderate to high shattering spikelets. Furthermore, shattering may be expressed in varying degrees depending on harvesting methods (Ji et al., 2006). Mechanical threshing is required to remove the seeds from the plant but more force may be required depending on abscission layer development. Moderate-shattering as opposed to hard-shattering is preferred for hand harvesting (Ji et al., 2006). The environment may also play a role in abscission layer development but little is known about this (Ji et al., 2006: 1002). Selection of non-shattering in rice may well have involved balancing ease of threshing against non-shattering and the magnitude of the benefit of a non-shattering mutation (Zhang et al., 2009: 718). These studies indicate that non-shattering is a complex trait and potentially has concomitant morphological variation.
Reduced seed dormancy leads to uniform germination (Sugimoto et al., 2010; Vaughan et al., 2008a). Dormancy is an advantage in the wild because seeds can survive for long periods until conditions are appropriate to sprout. In crops, selection by people favours reduced dormancy because a complete loss of dormancy means that seeds will sprout too early and be difficult to store. Limiting dormancy permits relatively uniform germination. Seed dormancy, prevalent in some plants such as Chenopodium, is not solely based on thinning seed-coats. Seed dormancy has not been specifically addressed in the archaeological rice record. Seed coat-imposed dormancy does not appear to be the case in rice. The endosperm and coleorhiza surrounding the radicle are likely influencing dormancy in rice (Sugimoto et al., 2010: 5795).
Synchronicity of flowering (Table 1) is correlated with the timing of fruit ripening, another trait advantageous to humans who can harvest at a predictable time rather than cope with the irregular seed maturation of wild rice. An increase in the number of grains per plant is also important to productivity. The number of grains is higher in the crop because the panicle has more branches that are also densely packed. Related to this trait is a reduction of tillering and the development of an erect growth habit favoring denser planting (Jin et al., 2008). During the growth phase preceding flowering, wild rice is prostrate with many, short tillers (Pang et al., 1996). This plant architecture keeps the density of plants low and shades out competitors. People may have selected against prostrate growth if they were trying to increase grain production (Jin et al., 2008). The number of tillers is inversely correlated with the number of panicle branches; the traits appear to be controlled by the same gene (Miura et al., 2010: 548) so that selecting for upright growth at the vegetative stage would have permitted more plants per unit area and larger numbers of seeds per plant to be produced. Specific human behaviours may be linked to rice plant architecture (Fuller et al., 2010a).
Wild rice seeds are more uniform in shape than seeds of domesticated rice yet still have considerable morphological variation among and between subpopulations (Table 1; Takano-Kai et al., 2009). Seed widths tend to be larger in Japonica thereby increasing the mass of the seeds (Shomura et al., 2008: 1024). Grain length and thickness are less important contributors to increased grain weight (Song et al., 2007). At least four genes contribute to seed size: GW2 (grain width 2), SW5 (seed width 5), GIF1 (grain incomplete filling 1), and GS3 (grain size 3 in Indica) (Shomura et al., 2008; Song et al., 2007; Takano-Kai et al., 2009). The deletion of the allele qSW5 appears to be associated with increased width and may have been selected for by people and that qSW5 deletion appears earlier than the defective qSH1 that is involved with loss of shattering (Shomura et al., 2008: 1026). Rice is also able to increase seed mass despite tough hulls that limit grain size by rapid grain-filling so that more mass can accumulate quickly. How this is accomplished is an active area of research in rice botany (Kobata et al., 2006; Wang et al., 2008).
The annual habit is critical because of the higher seed productivity over perennial rice. Habitats with regular, annual disruption would favour annual rice such as annual flooding reinforced by monsoons (Sato, 1996). Rice plants with less prostrate growth and fewer tillers could potentially be selected for management; however, the archaeological record is so far silent on these and other plant architecture traits such as more inflorescence branching that correlates with higher numbers of seeds. Isolation of annual, managed rice from perennial wild rice may not have been difficult owing to the preponderance for annual wild and domesticated rice to self-pollinate and the different habitat requirements for annual and perennial rice. Introgression from domesticated rice to wild rice also means that some wild rice populations would coevolve as domestication proceeds, probably clouding the already murky distinctions among rice remains at archaeological sites. Some features can be observed in the archaeological record: grain shape and size, presence and characteristics of awns, and shattering but these represent only a few traits among which others were potentially undergoing early selection by people. Finally, the wide range of DRTs and crossings of wild, weedy, and domesticated rice raise questions about how and when to recognize domesticated rice in the archaeological record. Multiple lines of evidence including quantity of grains, the presence and quantity of weedy rice, morphology of not only grains and spikelets but of culms and other plant tissue that may be preserved in wet deposits, weed mimics, and archaeological context are all potential data sources. Weed flora, currently being explored by Dorian Fuller and his team, will also be a productive source of information.
Archaeological rice
The archaeological record for rice presents many challenges, particularly because the remains of rice from archaeological deposits do not present some of the traits that could be relevant to understanding the initial selection process such as annual habit and architecture. Among the many DRTs (Table 1) non-shattering and grain size/shape changes are understandably privileged in archaeological discussions because charred seeds are common in the archaeological record and wet sites have good preservation of spikelet bases that have clues to the development of non-shattering rice (Ahn, 1993; Crawford and Shen, 1998; Fuller and Qin, 2010; Fuller et al., 2007, 2010; Liu et al., 2007; Zheng, 2007). Variation of the physical indications of shattering versus non-shattering has also made significant contributions to the discussion of rice domestication in ways that have not been applied to other grains (Fuller et al., 2009).The presence and characteristics of awns is also contributing to the discussion (Liu et al., 2007; Tang et al., 1993). Seed size studies rely primarily on length and width, although two recent studies include other measurements (Fuller et al., 2010b; Zhao and Gu, 2010). Width has been emphasized in some studies (e.g. Fuller et al., 2010b) because of the understanding that it is under separate genetic control. Seed size variability (see Table 1) is not attributed much significance although the data indicate that archaeological rice seed size variation is considerable; only one study includes an ANOVA (Liu et al., 2007a, b). Thickness is not normally taken into consideration, nor is shape, except for calculating ratios of length and width. Grain shape and the extent to which it evidences the domestication process may be complex, although it should be assessed case by case (see Rovner and Gyulia, 2007 for a detailed discussion). The role of heating and charring of grains needs to be carefully investigated. The duration and temperature of firing and in some cases the heating rate affect the size of charred seeds of wheat, pea and sunflower differentially (Braadbaart, 2007; Braadbaart and Bergen, 2004; Braadbaart and Wright, 2007) so we might expect the same for rice.
To date, all comparisons of wild rice and domesticated grain length and width measurements document a significant overlap; that is, the small grains being recovered from archaeological sites cannot distinguish wild and domesticated rice (Ahn, 1993; Zhao and Gu, 2010); however, a tendency to increased grain width over time has been suggested and grain size increase is thought to precede a general increase in non-shattering spikelet frequencies (Fuller et al., 2010b; Purugganan and Fuller, 2009). This is also predicted by genetic research (Shomura et al., 2008: 1026). However, the trend illustrated in Fuller et al. (2010b: figure 5b) is not yet statistically verified and may be significantly influenced by the 3300
Considering this dilemma, Zhao and Gu (2010) undertook a multivariate analysis of rice grains, comparing modern comparative specimens of wild and domesticated reference samples with each other. They relied on grain length, width and thickness, but added embryo length and width to the discussion. They also measured spikelet bases but found these measurements to have no diagnostic utility. From the five variables they determined discriminant function coefficients and developed formulae to determine whether rice grains are wild or not. They conducted blind tests that showed a high degree of accuracy with known samples (Zhao and Gu, 2010: 263). Charring seems to have little impact on the results, possibly because the embryo size has a significant impact on the results. Their paper is in Chinese and difficult to obtain in the west so the formulae are cited below:
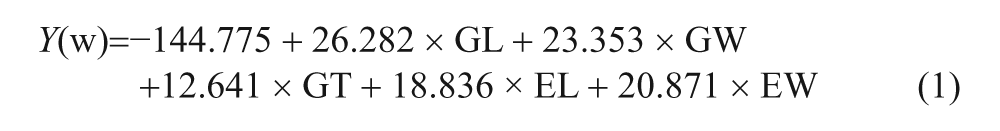
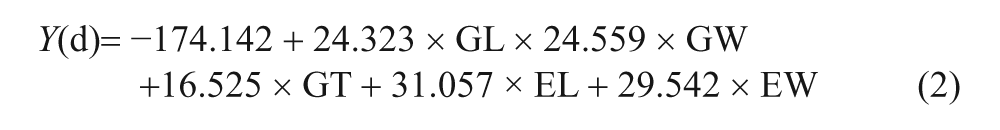
Grain measurements are entered into both formulae and the results are compared. If Y(w) is larger than Y(d) then the grains are from wild rice and conversely the grains are from domesticated rice. They applied this formula to four archaeological samples. The proportion of domesticated morphology grains is as follows in chronological order from 8000 to 5000 BP: Bashidang is 47%, Dingjiagang is 67%, Chengtoushan is 79%, and Yejiamiao is 92%. The results show an increase in proportion of domesticated morphology rice over time (Zhao and Gu, 2010: 264). This analysis is not without problems, however. Several of the variables have significant linear correlations so subsequent renditions of this study should only use independent variables. The distinction between the seeds of wild and domesticated populations may not be quite so clear. Furthermore, seed size studies should also consider weedy rice (O. sativa f. spontanea)
Fuller et al. (2009) emphasize that immature grain harvesting needs to be taken into account when assessing the seeds and spikelets. This is a novel and refreshing perspective that until recently has rarely been considered when assessing the archaeological evidence for domestication and is important because it is critical to the discussion of whether domesticated rice was present at sites and in what proportions. The hypothesis is that wild rice seed would have been harvested before the spikelets shattered and were lost. This perspective is not universally accepted (Liu et al., 2007a; Pan, 2008). The premise assumes that hunter-gatherers need to harvest immature grasses to maximize the harvest otherwise shattering would lead to significant losses. However, the ethnographic evidence documents hunter-gatherer exploitation of mature, wild grasses for the most part. Furthermore, mature seed collecting was important before food production in Southwest Asia and collecting grains after they fell to the ground was likely an important strategy there; most of the wild barley (Hordeum vulgare ssp. spontaneum recovered at Netiv Hagdud is mature (Kislev et al., 2004). The extensive collection of grass seeds at the Palaeolithic Ohalo II site are all ‘fully mature’ (Weiss et al., 2004: 9552). Not only is shattering an issue, but wild grass seeds do not mature synchronously so collection strategies need to consider this. Harris (1984) cites one example in Australia where hunter-gatherers harvested grass seeds just before they began to shatter (as does Fuller, 2007) but Harris also cites the Owens Valley for whom the most detailed data on grass harvesting are available. In this case, the Paiute harvested mature grass seeds. Furthermore, people harvest mature wild rice today using simple techniques such as twisting stalks together (Vaughan et al., 2008a), much as First Nations (Native Americans) do with Zizania spp. and has been documented for African wild rice harvested near Lake Chad (Harlan, 1985). Harlan (1985) describes the extensive exploitation of a wide range of wild grasses in Africa and describes the many strategies people used to harvest ripe, wild grass seeds. Anderson (2005) also describes the wide range of mature grass seeds collected using seed beaters in western North America.
To me, it is not so obvious that people would have had strategies to harvest immature rice. Nevertheless, signs of immature rice seed harvesting are evidenced in the archaeological record. Small grains and empty husks are thought to be indications of immature rice (Fuller et al., 2007). Other reasonable explanations for empty spikelets and smaller grains have been suggested (Liu et al., 2007b; Pan, 2008). Furthermore, uncharred rice spikelets are common at wet sites but no uncharred rice is reported from wet sites in East Asia. We have been testing this issue and, while spikelets contain significant silica content that aids in spikelet preservation, the starchy rice grains do not appear to preserve in wet contexts. Subsequently, another set of criteria have been reported that appear to distinguish immature and mature rice spikelet bases as well as shattering and non-shattering spikelets (Fuller et al., 2009). A recent PhD dissertation (Pan, 2011) finds variation among mature and immature spikelet bases (using different mechanical separation methods) of ten rice types (wild, cultigen and crosses). She notes that protruding rachis bases are not all necessarily from immature plants. Investigators might do well to expand their investigation of spikelet base morphological variation given the complexities of abscission layer development and variation discussed earlier. Different balances of shattering through semi-shattering to non-shattering may be related to harvesting techniques and degree of introgression among the continuum from wild, weedy through domesticated rice (whatever the later is at particular times). The views that immature rice was significant to early rice harvesters is a critical research area that is provocative yet I think still not conclusive.
Large-scale issues
Understanding rice domestication is entangled in contrasting views of the meaning of the concepts of agriculture and hunting-gathering that embodies an explicit dichotomy. In recent years this dichotomy has been critiqued in general (Smith, 1998a, 2001) as well as in specific cases (Crawford, 2008; Denham, 2008, 2011). One solution has been to identify a range of relationships between plants and animals on the one hand and people on the other and to refer to them as ‘in between’ or low-level food producers; they are neither transitional to agriculture nor transitional from hunting-gathering (Smith, 2001). Domestication has normally been considered the most important feature of agriculture so, legitimately, investigators have focused on the extent to which domesticates are represented in the archaeological record. This is not so much the case today because of the understanding that the physical evidence of domestication is often not clear and we should not presume to know which organisms in the archaeological record were not undergoing a form of management or selection (e.g. Crawford, 2011; Zeder et al., 2006). I do not like, nor use, the term ‘pre-domestication cultivation’ for these reasons as well as the directional nature of the concept. Furthermore, the archaeological record is difficult to interpret, especially if we rely on common sense definitions and easily identified characteristics of important crops such as rice. Agreement on how to classify food-producing regimes is not easily attained (Harris, 2007). Such conceptualizations may not be empirically based definitions but may in fact be influenced by privileged, usually Eurocentric perspectives (Deur and Turner, 2005). For me, determining if a group is agricultural is a classification exercise that distracts from the more interesting questions of how and under what circumstances people selected certain traits in plants and animals, and what the range of plant and animal use patterns mean and how they developed, stabilized or changed. Mutualism is a more general issue that includes rice management and selection so we should investigate a wide range of plant (and I would add animal) management strategies (Zeder et al., 2006).
Research on domestication, and I might add agricultural origins, focuses on either the specifics of time, place, and organisms or issues related to explaining domestication and the rise of agriculture by identifying forcing mechanisms (Smith, 2007; Zeder, 2006). The recent discussions of rice domestication and the development of agriculture in the Lower Yangzi Valley are no exception. The debates concerning whether specific remains are domesticated or not, the timing of rice domestication and whether it is a lengthy process, and where and when certain characteristics first appear fall into the first category (Fuller et al., 2007, 2009; Liu et al., 2007a, b; Zheng, 2007). Explanations of why and how domestication and agriculture developed in the region are few but range from possible climatic triggers such as the Younger Dryas (YD) (Boaretto et al., 2009; Cohen, 1998) to a multivariate analysis of climate and other factors (Chen, 2004).
Climatic determinist models have been critiqued, not only in their substance but in terms of the precise correlation of cultural and climatic episodes (Maher et al., 2011). The YD hypothesis in China relies on no high quality data so the record for the earliest rice management being later than the YD is no surprise. In any case, the YD and other climate changes do not fit the archaeological data, particularly in the southern Levant where this climatic trigger has been thought to have had some merit; in fact, culture changes related to sedentism and agriculture appear before the climate shifts in question in SW Asia (Maher et al., 2011). In the specific case of Abu Hureyra, the main study to hypothesize a specific role for the YD, the sudden climate change may have pushed people to make novel choices so that they could maintain their community year-round (Moore et al., 2000). This meant cultivating plants such as grasses that had previously been harvested from wild stands that were becoming scarce. Whether this hypothesis is correct is not crucial to the East Asian case but the Abu Hureyra study has value because it examines multiple lines of evidence including vegetation changes, seasonality, habitats exploited, and biotic signals of cultivation. The model proposes that plant and animal management were initially developed to maintain the community and its familiar resources under changing circumstances, not to change to some new, agriculture-based life.
In the Yangzi case, we could ask the same question: what strategies were people undertaking and under what modified conditions when plant and animal management were seen to be viable choices in order to maintain an economy that relied on exploiting resources from their ‘wild’ habitats? Chen (2004), in fact, hypothesizes that in both North and South China agriculture developed in ecotonal zones. He notes that small seed plant harvesting and even cultivation could have been common during the Pleistocene but that realistic food production was only feasible under certain conditions such as reduced patchiness, increased rainfall, and expanded floodplains (Chen, 2004). Furthermore, cultivation, domestication and agriculture need not be responses to a reduction of specific wild resources. Food production appears to begin in areas with relatively rich resources (Smith, 1998b) and in eastern North America the coalescence of a crop complex was an enhancement of hunting and gathering economies in resource-rich river valleys and was not a response to packing or carrying capacity challenges (Smith and Yarnell, 2009).
Another study evaluates the shift away from acorn procurement to rice production and explores whether a climatically induced reduction in acorn availability may have triggered rice cultivation (Fuller and Qin, 2010). The study is predicated on the position that a popular trigger for agricultural beginnings is climate change and that we should find out how climate changes ‘adversely affected’ vegetation (Fuller and Qin, 2010: 141). Implicit in this discussion is that we should be looking for a resource stress that pushed people to take up food production under adverse conditions but that we haven’t found them yet. This approach does not have much power elsewhere so it probably is not the best approach in China either. In fact, because acorns were abundant when rice cultivation was initiated, this hypothesis fails (Fuller and Qin, 2010: 155) and supports the view that agriculture developed in contexts of resource abundance.
Other plants, mutualism and niche construction
If people were targeting rice for management, then why assume that it was the only organism being managed? The Kuahuqiao site is instructive (Zong et al., 2007; ZPICRAZM, 2004). The occupation represents a well-developed adaptation at the mouth of the Qiangtang River with low hills to the northeast and southwest and at the time it was occupied it was near the coast. The most recent occupation layer is about 1 m below sea level. The habitation lasted for about a millennium from 8000 to 7000 cal. BP. The plant and animal remains include marine (e.g. dolphin, Delphinidae) and freshwater aquatic and terrestrial resources, dogs (Canus familiaris), and pig (Sus scrofa) management (if not domestication) (ZPICRAZM, 2004). Woody plants that include acorn (Quercus spp.), peach (Prunus persica/P. simonii), apricot (P. armeniaca), and Japanese apricot (P. mume) as well as foxnut/gorgon (Euryale ferox) and Trapa (T. bicornis and T. quadrispinosa) that are classified (Chen et al., 2007) as subspecies of T. natans or water chestnut), and are usually considered ‘wild’ and having been harvested from ‘natural’ habitats. Another woody plant seed is illustrated in the first Kuahuqiao site report (Camellia sinensis, tea; ZPICRAZM, 2004) but the species identification has been retracted so it is not discussed here. Categories such as ‘natural’ are culturally loaded terms. For example, geometric fields are usually thought of as not natural in the west but in Japan rice paddies are usually considered part of nature (Ohnuki-Tierney, 1993). Conceptualizing pre-agricultural peoples in the Hangzhou Bay region as nut-gatherers who eventually switched to agriculture is a far too constrained, reductionist hypothesis. Just as I have argued for the Jomon that we should expand our theoretical repertoire beyond simplified dichotomies and not be limited by strict application of hunting-gathering theory, we should consider a broader range of perspectives in the Yangzi valley discussion (e.g. Pan, 2011).
Peach and apricot are known mainly as cultivated plants whose origins are obscure but they are considered to be native to China (USDA, 2011a, 2011b). Eurayle and Trapa, too, are crops in China today. Nevertheless, at the very least these taxa raise questions about the early history of these plants that we know today to be important crops. Zong et al. (2007) also suggests that the extensive Typha growth in the wetlands at Kuahuqiao was a managed resource. Should we not be testing whether these are in the initial or even more developed stages of arboriculture and/or cultivation and management? Furthermore, some species of oak (Quercus sp.) are fire tolerant and burning is evident at Kuahuqiao (Zong et al., 2007). Acorn may have been collected from a large area requiring remote camps as Fuller and Qin (2010) hypothesize for Tianluoshan. This is a good foundation from which to examine deviations from the model so what deviations may be considered? In California, Native Americans managed acorn production by burning (Anderson, 2005; Lewis, 1972); oaks were also encouraged by Native American burning in Illinois (McClain et al., 2010) as were tree masts and fruit in general in North America (Abrams and Nowacki, 2008). Evidence that the Jomon in the Tohoku region managed nut production is quite strong (Kitagawa and Yasuda, 2004). Fuller and Qin (2009: 94) raise the possibility for Zhejiang Province but do not develop the concept in later work. It is, however, being examined by Pan (2011). I am not arguing here that, in fact, the landscape or a wide range of plants were managed or cultivated at Kuahuqiao but limiting the discussion to rice and passive acorn harvesting in the Lower Yangzi region is masking other important questions about anthropogenesis and niche construction well before the Hemudu culture.
The apparent decline in oak in the pollen record from the region might argue against oak management but does it? Although Fuller and Qin (2010: 151) interpret the decline, peaks and dips in oak pollen at Kuahuqiao to mean that oak was changing in local abundance due to climate, another interpretation may be considered. The pollen diagram in Zong et al. (2007) is ambiguous and the investigators are reluctant to attribute the changes in the pollen profile to climate because of anthropogenic effects (Innes et al., 2009: 2290). The pollen percentage for oak is calculated on the basis of all terrestrial pollen (only five arboreal taxa and four terrestrial herbs, two of which are small and large-pollen grasses). The most common pollen is from only two groups of plants, oaks and small grass pollen. In a more detailed treatment, pine is a minor component throughout the sequence (Innes et al., 2009). Because oak and grass are percentages of the same total pollen, the percentages covary. Both oaks and grasses could be abundant in the local area because of management practices so the percentages do not necessarily signal an oak decline. An increase in grass in the area would make oak appear to be declining while it actually was not. Microcharcoal is most abundant during the period of high grass pollen percentages indicating that anthropogenic fire is a significant factor in the local ecology. A detailed examination of the microcharcoal could assist in identifying its source. Acorns are abundant at Kuahuqiao and if they are abundant throughout the sequence, then they are clearly maintained as an important resource. Radiocarbon dating acorns from a variety of contexts would be one piece of evidence among other tools that could help assess whether arboriculture took place.
Finally, Brian Hayden (2011) suggests that early rice management may have involved swamp cultivation that involved clearing and ploughing/spading seasonally dry wetlands and planting rice in them. Although rice may have been a component of the flora of these habitats, rice might have been transplanted or sown in these locations to increase its production and encourage annual rice, as opposed to perennial rice, to flourish (Hayden, 2011: 80). These locations may also have begun to isolate these preferred varieties of rice in contexts where gene flow among wild, weedy and managed rice was relatively free compared with modern agricutural fields (Jones and Liu, 2009). In this scenario, humans pushed rice to produce more seed by encouraging the annual forms to grow in drier soil. Hayden goes on to suggest that the increased risk of this endeavor may have enhanced the prestige of rice, particularly if it was being used in feasts (Hayden, 2011: 81).
Settlement and geoarchaeological issues
The geoarchaeology focus of this issue of The Holocene is an opportunity to bring novel data sets to the agricultural origins discussion. The Kuahuqiao people integrated themselves into a complex wetland and terrestrial environment (Innes et al., 2009; Zong et al., 2007). The summer monsoons strengthened in the area from the beginning of the Holocene until 9000 cal. BP, then weakened until the beginning of the Hypsithermal when the monsoons once again strengthened (c. 7600 cal. BP in the region) (Innes et al., 2009: 2289) providing enabling conditions for annual rice to flourish. They also propose that receding flood management was a tool that permitted increased productivity of aquatic resources. The people in this region appear to have been adept at risk management, with a wide variety of techniques and resources at their disposal in a seasonally varying wetland setting. In other words, evolutionary potential was high.
Since 1998 a number of sites with occupations that immediately precede Kuahuqiao and Xiasun such as Xiaohuangshan, and Shangshan have been reported and provide new insight into the period during which cave/rockshelter sites were only known previously (Diaotonghuan, Xienrendong, Yuchanyan, Zengpiyan). Diaotonhuan and Xienrendong are still the only sites in this list with good evidence for rice from the Late Pleistocene to early Holocene boundary. Extensive flotation sampling at Zengpiyan has no convincing evidence of early agriculture (Zhao, 2010b). New excavations took place in the mid-2000s at Yuchanyan in order to examine the dating of the site, the late Upper Palaeolithic adaptations in the region, and whether rice was part of the subsistence of the occupants. Resource intensification is often cited as being associated with the earliest shift to agriculture but the evidence for it at Yuchanyan is equivocal (Prendergast et al., 2009). Only a few rice remains have been reported from Yuchanyan (Yan, 1997; Yasuda, 2002) and given the miniscule sample size and lack of AMS dates on the rice from Yuchanyan I doubt that Yuchanyan has much to contribute to our understanding of early rice and human interaction.
The recently reported open-air sites (Xiaohuangshan and Shangshan) are likely only a small sample of the occupations that were in the area at the time. Not only have water levels and the coastline varied over the millennia but the Hangzhou Bay and Shanghai region are highly developed with a population of at least one-third of Canada’s. Unfortunately much of the archaeological record in the area is gone, although considerable land area can still potentially be surveyed. The impact of receding and advancing sea levels on the lower Yangzi has been well documented in this region that, on average, is little more than 5 m above sea level today (Zong et al., 2007). The nature of the coastlines before 10 000 cal. BP have not been considered to any great extent yet. The lack of evidence for Upper Palaeolithic sites and the paucity of occupations immediately preceding Kuahuqiao may not only have been erased by modern development but by rising sea levels after the LGM. Coastal archaeology is beginning to demonstrate that an archaeological record exists offshore in some parts of the world (Flemming, 2004). Hangzhou Bay is relatively shallow with an average depth of about 30 m. Land area comprising most of the continental shelf extending up to 600 km from the current coast would have been exposed between the LGM when sea levels were about 120 m lower than today (Jiao, 2006; Lu et al., 2002; Zong et al., n.d.). Rising sea levels at the end of the Pleistocene have been proposed as a cause of population pressure related to triggering a shift to agriculture (Binford, 1968) but this is not what I am suggesting here. These now-inundated regions may well have been the locales for Upper Palaeolithic sites and the initial coastal-brackish water and freshwater wetland-terrestrial adaptations that appear so well developed at Kuahuqiao, Xiasun, and possibly Shangshan and Xiaohuanghan (although the research at these sites is still ongoing). A core from near the palaeo-estuary of the Yangzi River retrieved about 2800 phytoliths among which small numbers of rice phytoliths are found between c. 13 900 and 13 000 cal. BP during a mild period before the YD (Lu et al., 2002) suggesting that rice was growing in the region relatively early; the suggestion that this rice is an early domesticated form (p. 382) does not make much sense however. Rice would have been even more feasible to include in the resource suite as the climate warmed at the beginning of the Holocene and monsoonal systems eventually reached this latitude.
Conclusions
Research on the origins of agriculture and the evolution of domesticated rice in the Yangzi valley is developing rapidly in the context of an engaged Chinese archaeological community, interdisciplinary and international collaborations, and genetic research that focuses on crop improvement. This paper reflects on these important developments by assessing interpretive complexities and indicating where research may be directed in the future. DRTs in rice are extensive and are, for the most part, quantitave traits. The taxon’s environmental responsiveness, hybridization/crossing ability, and the association of weedy rice almost universally with the crop have potential consequences to the interpretation of archaeological rice. Ancient DNA research could be productive given the growing knowledge base of the genetic underpinnings of certain DRTs. Research on rice, although crucial, should not cast a long shadow on other important human–plant–animal interactions. Broader conceptualizations of human–plant–animal interactions beyond traditional perspectives related to agriculture are essential in order to move forward. One recent integrative model was published just as I completed this paper (Hayden, 2011). We still need to investigate the history of use and domestication of other important resources in the archaeological record, not just rice. Research on acorn use has begun to broaden the scope, but the view that plants that eventually become economically important in China are all wild in the archaeological record assumes that we know the meaning of ‘wild’ with respect to these plants. In fact, we do not. Archaeologists need to question the extent to which many of these plants, and potentially others, were components of a managed landscape. Although not examined in any depth in this paper, anthropogenesis, niche construction, and the potentials of behavioural ecology may help to understand the complexities of human adaptations in the Lower Yangzi Valley in the Late Pleistocene and early Holocene. The limitations of the archaeological record in the lower Yangzi valley also need to be understood given the large-scale landscape transformations, climate change, and modern development in the region. The interdisciplinary nature of this special issue of The Holocene is another step in the right direction.
Footnotes
Acknowledgements
I would like to thank Yongqiang Zong for the opportunity to contribute to this special issue. Yongqiang Zong, Gyoung-Ah Lee and Pan Yan read earlier drafts of this paper and provided insightful comments.The reviewers of this paper were generous with their time, advice and insight and have helped me come to a better understanding of the complex issues in this paper. Discussions with Baorong Lu of Fudan University have also been edifying. Chen Shen helped me with some of the literature. I am grateful to the staff of the Kuahuqiao Project for their assistance and generosity with information.
My research and travels in China have been sponsored by the Social Sciences Research Council of Canada (SSHRC, grant 72022345), the University of Toronto Mississauga, Canada and Shandong University, Jinan, China.