
Abstract
Laminated sediments are unique archives of palaeoenvironmental and palaeoceanographic conditions, recording changes on seasonal and interannual timescales. Diatom-rich laminated marine sediments are examined from Dumont d’Urville Trough, East Antarctic Margin, to determine changes in environmental conditions on the continental shelf from 1136 to 3122 cal. yr BP. Scanning electron microscope backscattered electron imagery (BSEI) and secondary electron imagery are used to analyse diatom assemblages from laminations and to determine interlamina relationships. Diatom observations are quantified with conventional assemblage counts. Laminae are primarily classified according to visually dominant species identified in BSEI and, secondarily, by terrigenous content. Nine lamina types are identified and are characterized by: Hyalochaete Chaetoceros spp. resting spores (CRS); CRS and Fragilariopsis spp.; Fragilariopsis spp.; Corethron pennatum and Rhizosolenia spp.; C. pennatum; Rhizosolenia spp.; mixed diatom assemblage; Stellarima microtrias resting spores (RS), Porosira glacialis RS and Coscinodiscus bouvet; and P. glacialis RS. Formation of each lamina type is controlled by seasonal changes in sea ice cover, nutrient levels and water column stability. Quantitative diatom assemblage analysis revealed that each lamina type is dominated by CRS and Fragilariopsis sea ice taxa, indicating that sea ice cover was extensive and persistent in the late Holocene. However the lamina types indicate that the sea ice regime was not consistent throughout this period, notably that a relatively warmer period, ~3100 to 2500 cal. yr BP, was followed by cooling which resulted in an increase in year round sea ice by ~1100 cal. yr BP.
Introduction
Phytoplankton productivity in the Southern Ocean is dominated by diatoms. Their siliceous frustules are well-preserved in Antarctic sediments and their abundance and diversity make them a powerful tool for reconstructing sea-surface conditions and climate in the Holocene. Laminated diatom-rich sediments occur with a circum-Antarctic distribution in inner shelf basins and troughs and provide a unique opportunity for constructing ultra-high resolution records of environmental change (Armand and Leventer, 2010; Leventer et al., 2002, 2006, 2010; Pike and Stickley, 2007).
In 2001, a remarkably thick Holocene laminated diatom ooze sediment sequence, with laminations extending to the sediment surface, was discovered in the Dumont d’Urville Trough (DUT) (Figure 1a) off the Adélie Land coast, East Antarctica (Leventer et al., 2001). These sediments accumulate here in a sediment drift because the trough morphology creates a natural sediment trap. DUT dissects the continental shelf in a SE–NW direction just offshore of the Astrolabe and Zélée Glaciers (Figure 1a), two of the largest of several outlet glaciers that currently drain into the trough (Domack et al., 1991).
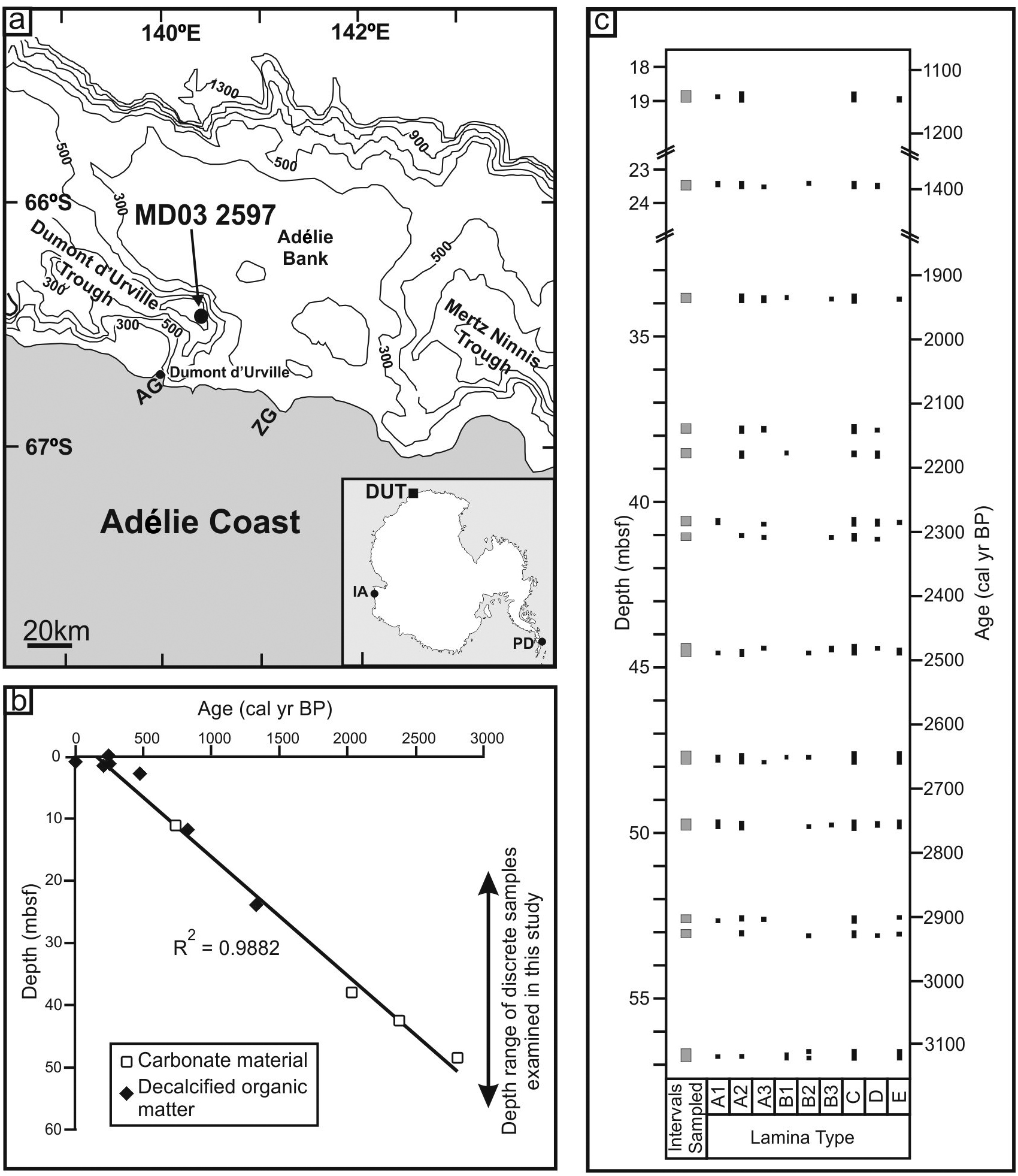
(a) Maps showing the location of Dumont d’Urville Trough (DUT) with coarse scale bathymetry in metres (adapted after Domack et al., 1989). Core site MD03-2597 is indicated. AG: Astrolabe Glacier; ZG: Zélée Glacier. Inset is the location of DUT on the East Antarctic Margin. IA: Iceberg Alley; PD: Palmer Deep. (b) Calibrated age–depth model for MD03-2597. (c) Distribution of discrete intervals sampled from MD03-2597 (grey blocks). Distribution of identified lamina types are indicated with black blocks (mbsf: metres below sea floor).
This area is influenced by the westward-flowing surface Antarctic Coastal Current and upwelling Modified Circumpolar Deep Water. Brine rejection as a result of sea ice formation contributes to the production of dense High Salinity Shelf Water which flows northwards as part of Adélie Land Bottom Water, a significant source of Antarctic Bottom Water (Marsland et al., 2004; Rintoul, 1998; Williams et al., 2008). As a result of strong katabatic winds (Periard and Pettre, 1993), a polynya currently forms in the vicinity of the DUT in the winter (66°11′S, 139°31′E) (Adolphs and Wendler, 1995; Arrigo and Van Dijken, 2003). Fast ice formation typically commences each April and grows westwards by pack ice interception but is relatively unstable over DUT because of the lack of grounded icebergs (Massom et al., 2009). During the summer most of the fast ice melts back towards the coast, however the exact timing of this melt back is subject to significant interannual variability (Massom et al., 2009).
During the last glacial maximum the East Antarctic Ice Sheet (EAIS) was grounded on the outer continental shelf (Anderson et al., 1980; Mackintosh et al., 2011) with elevations of up to 1000 m (Goodwin and Zweck, 2000). At this time the ice streams of the Zélée and Astrolabe Glaciers occupied DUT, bypassing the outer shelf banks (Eittreim et al., 1995). Deglaciation of the Adélie Land continental shelf occurred around 11 cal. kyr BP, with terminal glacier recession at the inner part of the shelf around 8.5 cal. kyr BP (Denis et al., 2009a).
Previously, diatom-rich laminated Antarctic sediments from the East Antarctic Margin have been successfully used to reconstruct early-Holocene climate and environments (e.g. Leventer et al., 2006; Maddison et al., 2006; Stickley et al., 2005). In these studies, the laminated sediment sequences did not extend to the sea floor so in order to improve understanding of the late-Holocene Neoglacial climatic phase around the East Antarctic Margin we examine laminated diatom ooze sediments from DUT. Our high-resolution approach allows us to investigate seasonal changes in sea ice, nutrients and surface water column stability conditions in DUT during this time period.
Materials and methods
Material and core chronology
Calypso core MD03-2597 (66°24.74′S, 140°25.26′E; 1025 m water depth) was collected from DUT (Figure 1a) during the MD130-Images X cruise (CADO-Coring Adélie Diatom Oozes) on board RV Marion Dufresne II in 2003 (Michel et al., 2003). Throughout much of its length the 57.34 m long core consists of black to dark olive grey diatom ooze laminations (Figure 2). As only four carbonate samples were AMS radiocarbon dated from MD03-2597, the core chronology was developed using additional total organic carbon dates from two proximal DUT cores (jumbo piston core NBP01-01 JPC17B (66°24.852′S, 140°25.164′E 1048 m water depth) and kasten core NBP01-01 KC17B (66°24.865′S 140°25.168′E 1048 m water depth) (Table 1). The carbonate radiocarbon ages were calibrated assuming a local reservoir age of 1200 yr which is only slightly higher than a recent general Southern Ocean Holocene reservoir age estimate of 1144±120 yr (Hall et al., 2010). The ages of the total organic carbon samples are older than the carbonate ages owing to the reworking of old organic carbon, therefore, a local reservoir age of 1600 yr was used for the total organic carbon decalcified samples. A best fit line was constructed through the 11 calibrated dates to construct an age model (Figure 1b). As a result of high sedimentation rates at this site, 13 discrete sediment intervals up to 25 cm long were sampled from MD03-2597 between 18.75 and 56.83 mbsf (Figure 1c) to provide an insight into the peak Neoglacial conditions. Age estimates for this interval are 1136 to 3122 cal. yr BP and the base of the core is dated at 3149 cal. yr BP.

Core photograph (56.78 to 56.63 m below sea floor (mbsf)) and backscattered electron imagery (BSEI) photograph of an example of annually laminated sediment deposition. Lamina types are labelled. CRS: Hyalochaete Chaetoceros spp. resting spores; MDA: mixed diatom assemblage; P.glacialis RS: Porosira glacialis resting spores.
Radiocarbon ages for core MD03-2597, NBP0101 JPC17B and NBP0101 KC17B.
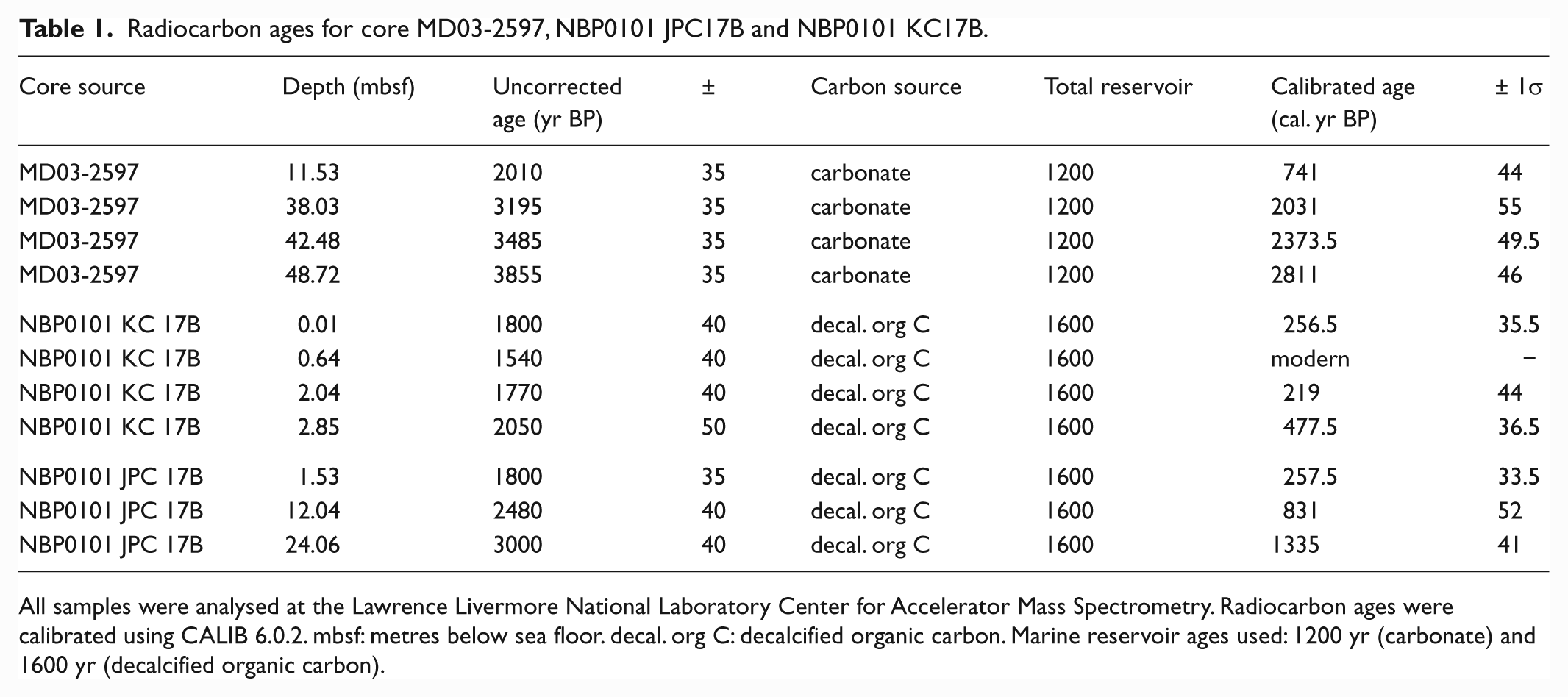
All samples were analysed at the Lawrence Livermore National Laboratory Center for Accelerator Mass Spectrometry. Radiocarbon ages were calibrated using CALIB 6.0.2. mbsf: metres below sea floor. decal. org C: decalcified organic carbon. Marine reservoir ages used: 1200 yr (carbonate) and 1600 yr (decalcified organic carbon).
Scanning electron microscope analysis
A fluid displacive resin embedding technique (Pearce et al., 1998; Pike and Kemp, 1996) was used to prepare polished thin sections from the sediment slabs. These were examined on a Cambridge Instruments (LEO) S360 scanning electron microscope (SEM) using backscattered electron imagery (BSEI). Photomosaics were produced from low-magnification images (×20) of each polished thin section (Figure 2) and then, using the mosaic as a map, laminae were examined using higher magnification (×100–1000) to investigate the sediment fabric and perform qualitative analysis of the main diatom species present (Figure 3). The laminae were primarily classified according to visually conspicuous diatom species and, secondarily, by the terrigenous grain content versus biogenic content. The distinction between terrigenous and biogenic laminae is qualitative and follows methods used in previous studies (e.g. Kemp, 1990; Maddison et al., 2005; Pike and Kemp, 1997). In BSEI biogenic laminae are dark as a result of the almost pure diatom ooze composition (low atomic number, carbon-based resin infilling the high porosity of the diatom frustules) and terrigenous laminae are bright because of the greater proportion of clay and silt grains (high average atomic number of the more closely packed terrigenous grains). These two types of laminae are visually distinct in BSEI (Figure 2). To aid diatom species identification from BSEI photographs, blocks of sediment (~0.5 cm3) were cut from the sediment slabs and fractured parallel to the laminations. The blocks were mounted onto SEM stubs, allowed to dry for 24 h and then coated in Au-Pd (90:10). Topographic secondary electron imagery (SEI) analysis of the fractured surface was conducted using a Veeco FEI (Philips) XL30 Environmental SEM with FEG (Field Emission Gun).

Backscattered electron imagery (BSEI) and secondary electron imagery (SEI) photographs of lamina types. (a) BSEI photograph of biogenic laminae characterized by Hyalochaete Chaetoceros spp. resting spores (CRS); (b) SEI photograph of biogenic laminae characterized by CRS; (c) BSEI photograph of biogenic laminae characterized by CRS and Fragilariopsis spp.; (d) SEI photograph of biogenic laminae characterized by CRS and Fragilariopsis spp.; (e) BSEI photograph of biogenic laminae characterized by Fragilariopsis spp.; (f) SEI photograph of biogenic laminae characterized by Fragilariopsis spp.; (g) BSEI photograph of biogenic laminae characterized by Corethron pennatum and Rhizosolenia spp.; (h) SEI photograph of laminae characterized by C. pennatum and Rhizosolenia spp.; (i) BSEI photograph of biogenic laminae characterized by C. pennatum; (j) SEI photograph of biogenic laminae characterized by C. pennatum; (k) BSEI photograph of biogenic laminae characterized by Rhizosolenia spp.; (l) SEI photograph of biogenic laminae characterized by Rhizosolenia spp.; (m) BSEI photograph of mixed diatom assemblage terrigenous laminae; (n) SEI photograph of mixed diatom assemblage terrigenous laminae; (o) BSEI photograph of terrigenous laminae characterized by Stellarima microtrias resting spores, Porosira glacialis resting spores and Coscinodiscus bouvet; (p) SEI photograph of terrigenous laminae characterized by S. microtrias resting spores, P. glacialis resting spores and C. bouvet; (q) BSEI photograph of terrigenous laminae characterized by P. glacialis resting spores; (r) SEI photograph of terrigenous laminae characterized by P. glacialis resting spores. All scale bars: 50 microns. CRS: Hyalochaete Chaetoceros spp. resting spores. PhCh: Phaeoceros Chaetoceros spp. Frag: Fragilariopsis spp. F.kerg: Fragilariopsis kerguelensis. F.obliq: Fragilariopsis obliquecostata. C.pen: Corethron pennatum. CPG: Corethon pennatum girdle bands. Rhizo: Rhizosolenia spp. RGB: Rhizosolenia spp. girdle bands. R.ant: Rhizosolenia antennata f. semispina. S.micro: Stellarima microtrias resting spores. P.gl: Porosira glacialis resting spores. C.bou: Coscinodiscus bouvet.
Quantitative diatom assemblage analysis
Quantitative diatom slides were prepared using the settling technique described by Scherer (1994) and fixed with Norland Optical Adhesive (Refractive Index 1.56) in order to quantify diatom assemblages and abundance in selected laminae. An Olympus BX40 microscope (with phase contrast, at ×1000 magnification) was used to count a minimum of 400 diatom valves per slide. Two separate counts per slide were made: (1) a total species count; and (2) an Hyalochaete Chaetoceros spp. resting spore (CRS)-free count. The CRS-free counts allow trends of less common, but ecologically important, species to be revealed (Allen et al., 2005; Leventer and Dunbar, 1996). Relative abundances were calculated as percentages of either the total diatom assemblage or CRS-free assemblage (Table 2a and b). Average absolute diatom abundances were calculated as millions of valves per gram of dry sediment (mv/gds) (Table 2a) as detailed in Scherer (1994). Diatom taxonomic identification follows Hasle and Syvertsen (1997), Armand and Zielinski (2001), Johansen and Fryxell (1985) and Jordan et al. (1991).
(a) Relative abundance of diatoms counted, all species. (b) Relative abundance of diatoms counted, Hyalochaete Chaetoceros spp. resting spore (CRS)-free.
Table 2(a)


mv/gds: millions of valves per gram of dry sediment; FOV: Field of View; mbsf: metres below sea floor. Slides were prepared in a beaker with a diameter of 10 cm and the area of the microscope field of view used during counting was 0.000356 cm
Results
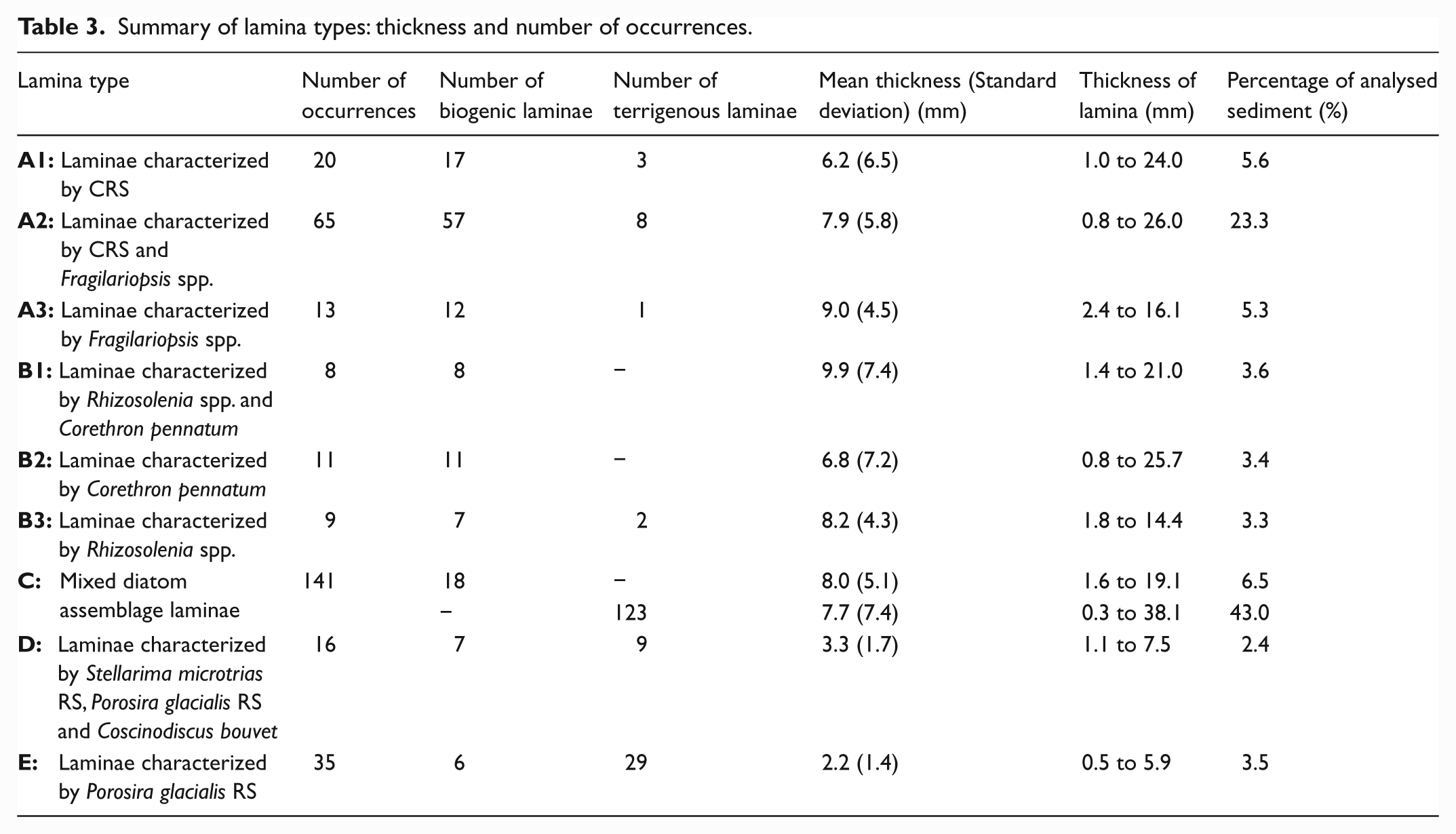
The late-Holocene diatom-rich laminated sediments from DUT consist of biogenic and terrigenous laminae. A total of 318 laminae are present in the 13 analysed laminated intervals (Figure 1c, Table 3). The terrigenous laminae contain silt- and clay-sized lithogenic grains, the abundance of which varies between laminae. Terrigenous laminae range from 0.3 to 38.1 mm and biogenic (i.e. pure diatom ooze) laminae range from 0.8 to 26.0 mm in thickness. No correlation between lamina thickness variations and core depth was observed in these sediments. Using BSEI and SEI nine types of lamina were characterized (and named) by visually conspicuous diatom species. In addition, the lamina types were classified according to their biogenic versus terrigenous content. The biogenic and terrigenous forms of a particular lamina type (LT) are reported together. Diatom assemblages in the BSEI/SEI-defined laminae were subsequently quantified; at least three samples of each lamina type were used to calculate average relative and absolute diatom abundances (Table 2a and b).
Summary of lamina types: thickness and number of occurrences.
Lamina type A1: Laminae characterized by CRS
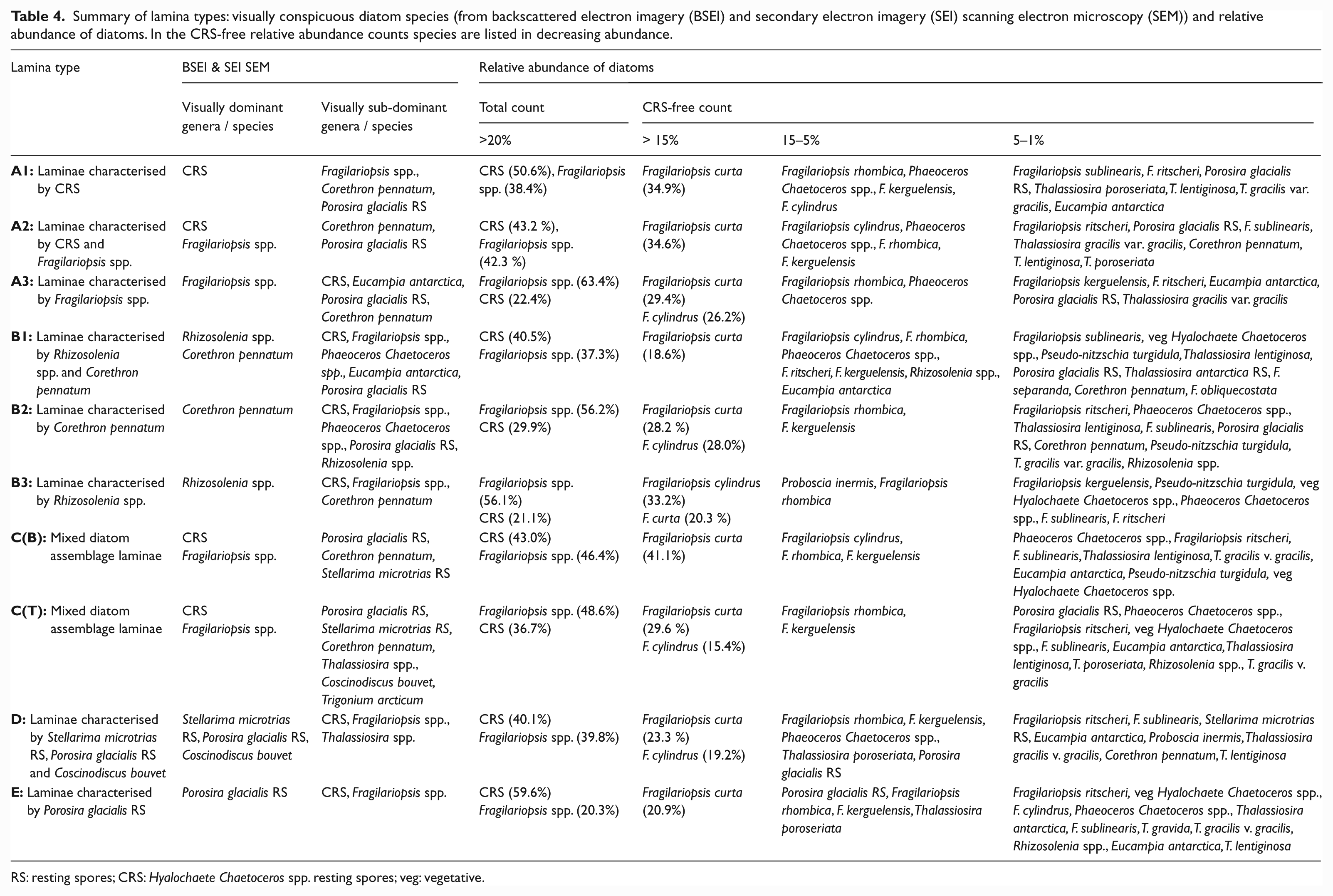
Inspection of LT A1 laminae using BSEI and SEI revealed that the diatom assemblage is visually dominated by CRS (Figure 3a and b). Visually sub-dominant species were Fragilariopsis spp., Corethron pennatum and Porosira glacialis resting spores (RS) (Table 4). Quantitative diatom assemblage analyses (Tables 2a and 4) confirmed the dominance of CRS and sub-dominance of Fragilariopsis spp., however, C. pennatum and P. glacialis RS are numerically rare (< 2%). The CRS-free counts (Tables 2b and 4) are dominated by Fragilariopsis curta and sub-dominated by F. rhombica, Phaeoceros Chaetoceros spp., F. kerguelensis and F. cylindrus (each between 5 and 15%). LT A1 occurs 20 times in the studied intervals, 17 of which are biogenic (Table 3). These laminae range in thickness from 1.0 to 24.0 mm, making up ~6% of the analysed sediments.
Summary of lamina types: visually conspicuous diatom species (from backscattered electron imagery (BSEI) and secondary electron imagery (SEI) scanning electron microscopy (SEM)) and relative abundance of diatoms. In the CRS-free relative abundance counts species are listed in decreasing abundance.
RS: resting spores; CRS: Hyalochaete Chaetoceros spp. resting spores; veg: vegetative.
Lamina type A2: Laminae characterized by CRS and Fragilariopsis spp.
CRS and Fragilariopsis spp. are the most visually conspicuous taxa in BSEI and SEI analysis of LT A2 laminae (Figure 3c and d). The total diatom assemblage is co-dominated by CRS and Fragilariopsis spp. (Tables 2a and 4). In CRS-free counts (Tables 2b and 4) the assemblage is dominated by F. curta and sub-dominated by F. cylindrus, Phaeoceros Chaetoceros spp., F. rhombica and F. kerguelensis (each between 5 and 15%). The visually sub-dominant species (BSEI and SEI), Corethron pennatum and Porosira glacialis RS, account for 3.6% of the assemblage (CRS-free). There are 65 occurrences of LT A2 in the analysed intervals, ~90% of which are biogenic (Table 3). This LT makes up almost a quarter of the analysed sediments and individual laminae range in thickness from 0.8 to 26.0 mm.
Lamina type A3: Laminae characterized by Fragilariopsis spp.
Fragilariopsis spp. are visually conspicuous in BSEI and SEI analyses (Figure 3e and f) and also dominate the quantitative diatom assemblage (F. curta and F. cylindrus are the most abundant species; Tables 2a and 4). CRS sub-dominate the total species count and F. rhombica and Phaeoceros Chaetoceros spp. sub-dominate the CRS-free counts (Table 4). LT A3 occurs 13 times, 12 of which are biogenic. The laminae range in thickness from 2.4 to 16.1 mm and comprise 5.3% of the studied sediment intervals (Table 3).
Lamina type B1: Laminae characterized by Rhizosolenia spp. and Corethron pennatum
BSEI and SEI analyses reveal that LT B1 are visually dominated by Rhizosolenia spp. and Corethron pennatum (Figure 3g and h). However these two taxa do not dominate the total quantitative diatom assemblage (Tables 2a and 4) because of their relatively large elongate cells and their propensity to break up (usually fragments are not counted during quantitative diatom assemblage analysis). The assemblage is in fact co-dominated by CRS and Fragilariopsis spp. In CRS-free counts the diatom assemblage is dominated by Fragilariopsis curta and sub-dominated by four other Fragilariopsis spp., Phaeoceros Chaetoceros spp. Rhizosolenia spp. and Eucampia antarctica (Table 4). C. pennatum constitutes 1.2% of the CRS-free count. LT B1 are the least common type in the analysed intervals and all eight occurrences are biogenic (Table 3). The laminae range from 1.4 to 21.0 mm thick and constitute 3.6% of the analysed sediment.
Lamina type B2: Laminae characterized by Corethron pennatum
Corethron pennatum visually dominates BSEI and SEI analyses (Figure 3i and j) of LT B2, but this species only constitutes 1.4% of the CRS-free quantitative diatom assemblage (Tables 2b and 4). Fragilariopsis spp. and CRS dominate, accounting for a total of 86% of the total species count. In CRS-free counts (Tables 2b and 4) the assemblage is co-dominated by Fragilariopsis curta and F. cylindrus and is sub-dominated by F. rhombica and F. kerguelensis. All 11 LT B2 are biogenic and range in thickness from 0.8 to 25.7 mm making up approximately 3% of the analysed sediment (Table 3).
Lamina type B3: Laminae characterized by Rhizosolenia spp.
Rhizosolenia spp. are visually conspicuous in BSEI and SEI analyses (Figure 3k and l), however, this genus does not dominate the quantitative diatom assemblage (<3%; Table 2a). Rhizosolenia antennata f. semispina is the most common species identified during counting. Fragilariopsis spp. and CRS are the dominant genera in this LT, accounting for 77% of the total species count. In CRS-free counts (Tables 2b and 4) F. cylindrus and F. curta are dominant, and Proboscia inermis and F. rhombica are sub-dominant. There are nine occurrences of LT B3 laminae, seven of which are biogenic and two are terrigenous (Table 3). The laminae range in thickness from 1.8 to 14.4 mm, making up 3.3% of total sediment thickness.
Lamina type C biogenic (B) and terrigenous (T): Mixed diatom assemblage laminae
LT C has been split into two forms, because of minor differences observed in diatom assemblages during SEM analyses which relate to the biogenic and terrigenous content of the laminae. However, these forms were insufficiently distinct to allow two separate lamina types to be defined.
LT C(B) are visually dominated by CRS and Fragilariopsis spp. and sub-dominated by Porosira glacialis RS, Corethron pennatum and Stellarima microtrias RS in BSEI and SEI analyses. The quantitative diatom assemblage is also dominated by CRS and Fragilariopsis spp. but not sub-dominated by Porosira glacialis RS, Corethron pennatum and Stellarima microtrias RS as these three species together only account for <2% of the assemblage (Tables 2a and 4). Fragilariopsis curta, F. cylindrus, F. rhombica and F. kerguelensis are the most common Fragilariopsis species present in this lamina type (Tables 2b and 4). The laminae range in thickness from 1.6 to 19.1 mm and make up 6.5% of analysed sediments.
In BSEI and SEI analyses LT C(T) are also visually dominated by Fragilariopsis spp. and CRS (Figure 3m and n) but the sub-dominant species are more diverse than in LT C(B) (Table 4). The quantitative diatom assemblage is dominated by CRS and Fragilariopsis spp. (Tables 2a and 4). The visually sub-dominant species Thalassiosira spp. and P. glacialis RS together account for ~11% of the CRS-free assemblage, whereas C. pennatum and S. microtrias RS are much less common (both <1% of the assemblage) (Tables 2b and 4). Coscinodiscus bouvet and Trigonium arcticum are also observed in BSEI and SEI analyses but are not observed during counting, this is possibly due to their large frustule sizes making them susceptible to fragmentation during slide processing. The CRS-free counts (Tables 2b and 4) are dominated by F. curta and F. cylindrus, and are sub-dominated by F. rhombica and F. kerguelensis. A diverse range of taxa each contribute between 1 and 5% to the assemblage of this LT (Table 4). LT C(T) are the most common lamina type in the analysed intervals and range in thickness from 0.3 to 38.1 mm, making up 43% of the analysed sediment (Table 3). LT C(B and T) laminae occur 141 times in the analysed intervals (18 LT C(B) and 123 LT C(T)).
Lamina type D: Laminae characterized by Stellarima microtrias RS, Porosira glacialis RS and Coscinodiscus bouvet
Stellarima microtrias RS, Porosira glacialis RS and Coscinodiscus bouvet are visually conspicuous in BSEI and SEI (Figure 3o and p) analysis of LT D. Both S. microtrias RS and P. glacialis RS are present in the CRS-free assemblage (1.6% and 5.0%, respectively) however C. bouvet is not observed, probably because of fragmentation of their large frustules. The quantitative diatom assemblage (Table 2a) is dominated by CRS and Fragilariopsis spp. The CRS-free assemblage is dominated by F. curta and F. cylindrus and sub-dominated by F. rhombica, F. kerguelensis, Phaeoceros Chaetoceros spp., Thalassiosira poroseriata and P. glacialis RS (Tables 2b and 4). LT D laminae are present 16 times throughout the analysed intervals; seven biogenic and nine terrigenous. These laminae range in thickness from 1.1 to 7.5 mm and constitute ~3% of total sediment thickness (Table 3).
Lamina type E: Laminae characterized by Porosira glacialis RS
Porosira glacialis RS are visually conspicuous in BSEI and SEI analyses (Figure 3q and r) and constitute 14.9% of the CRS-free assemblage (Table 2b). The quantitative diatom assemblage (Tables 2a and 4) is dominated by CRS and Fragilariopsis spp. CRS-free counts (Tables 2b and 4) are dominated by F. curta and Thalassiosira spp. and sub-dominated by P. glacialis RS, F. rhombica and F. kerguelensis. The most common Thalassiosira species present is T. poroseriata. LT E occurs 35 times in the analysed sediment intervals, six of which are biogenic and 29 are terrigenous (Table 3). This lamina type ranges in thickness from 0.5 to 5.9 mm. It comprises 3.5% of the analysed sediment, but is the third most commonly occurring of all lamina types.
Lamina relationships
Boundaries between biogenic and terrigenous laminae are gradational and are very occasionally bioturbated. Faecal pellets occur throughout the analysed intervals, but are not common place (<20 occurrences observed). The diatom taxa CRS and Fragilariopsis spp. dominate diatom assemblages in all of the lamina types (Tables 2a and 4). LT E have the greatest absolute abundance of diatoms (1164.9 mv/gds) and LT C(T) laminae have the lowest abundance (556.1 mv/gds) (Table 2a). On average, the absolute abundance of diatoms in LT A1-3 (851 mv/gds) is higher than in LT B1-3 (659 mv/gds). LT A1-3 and LT C are present throughout the core (Figure 1c). In the youngest interval examined (1136 to 1149 cal. yr BP) only LT A1, A2, C and E occur. LT B1-3 and LT E occur more commonly before 2475 cal. yr BP, and occurrences of LT D are more common after 2475 cal. yr BP (Figure 4b).
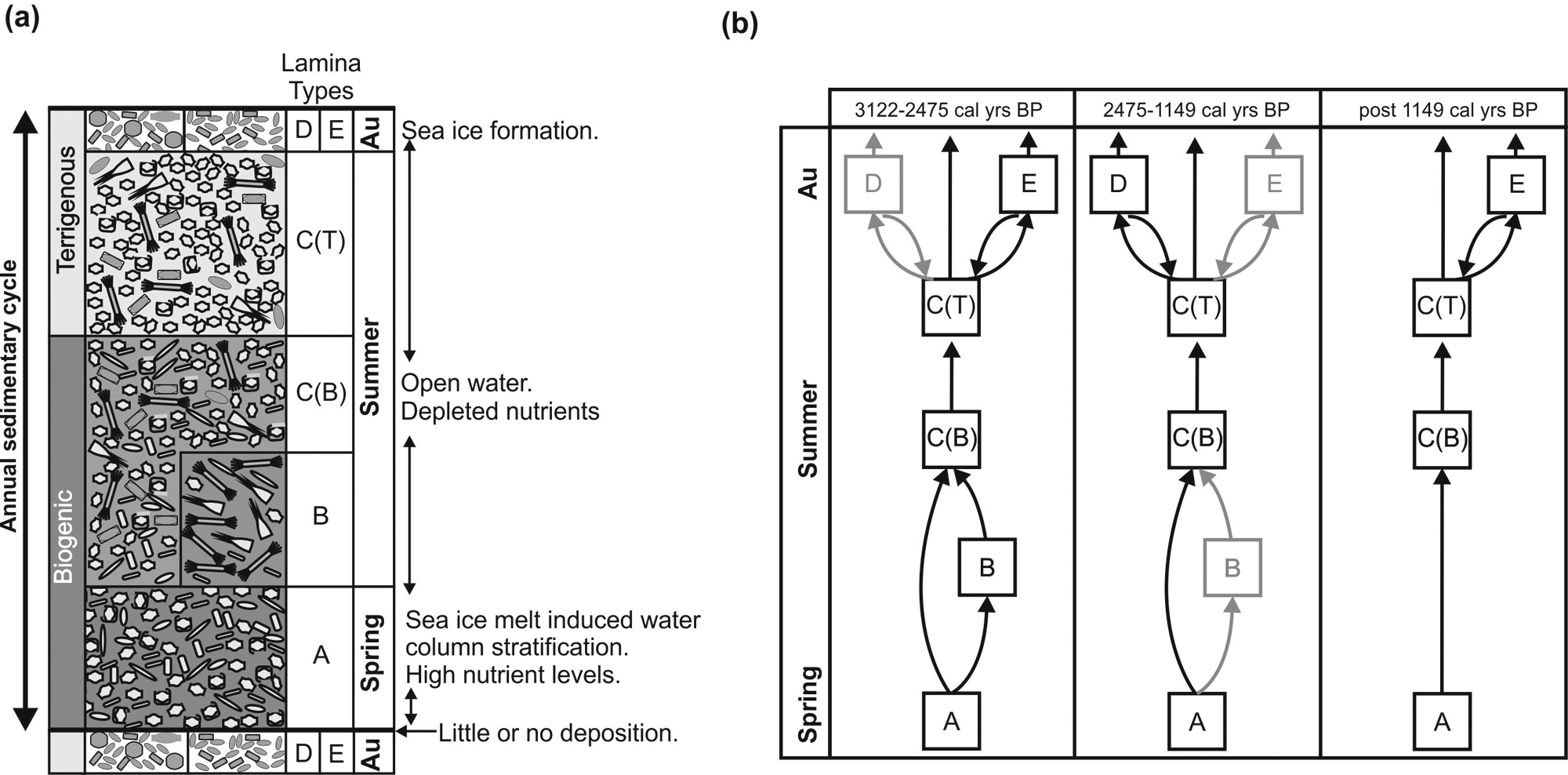
Schematic diagrams of typical sequences of annual lamina deposition in laminated sediments from Dumont d’Urville Trough (a) general; (b) for three discrete time periods. Au: Autumn. A: Lamina types characterized by Hyalochaete Chaetoceros spp. resting spores and/or Fragilariopsis spp. (LT A1–3); B: Lamina types characterized by Corethron pennatum and/or Rhizosolenia spp. (LT B1–3); C(B): Mixed diatom assemblage biogenic laminae; C(T): Mixed diatom assemblage terrigenous laminae; D: laminae characterized by Stellarima microtrias resting spores (RS), Porosira glacialis RS and Coscinodiscus bouvet; E: laminae characterized by Porosira glacialis RS.
Several permutations in the order of lamina occurrences have been observed in the analysed core intervals, however, a typical sequence of laminations has been identified and is represented schematically in Figure 4a. Laminae in the lower part of the lamina sequences are biogenic and characterized by LT A1-3. These laminae are usually succeeded by LT C(B); however, occasionally, LT B1-3 may occur between LT A1-3 and LT C(B) laminae. In the upper part of the lamina sequences, laminae have a higher terrigenous content with a more mixed diatom assemblage (LT C(T)). The lamina sequence ends with terrigenous LT D or LT E (terrigenous or biogenic). Not every lamina type occurs in every sequence (e.g. Figures 2 and 4b).
Discussion
Seasonal signal
The lamina types are discussed in groups determined by the diatom species that are visually conspicuous in BSEI and SEI analyses. While there are a range of processes that can produce biogenic or terrigenous laminae types, such as dilution, sediment transport and changes in surface water productivity, we propose the most appropriate mechanisms according to the glaciological/oceanographic conditions prevailing in the season. We suggest that each lamination was created as a result of a seasonal productivity event and, therefore, have determined that a total of 120 discrete years of sediment were deposited within the 13 sediment sections examined. This value is in agreement with the number of years calculated using the calibrated age–depth model (117 years, Figure 1b). Approximately two-fifths of all the identified annual sediment increments are composed of more than two laminae. The repetition of laminae is irregular and is similar to that found in Holocene postglacial sediments from the Mertz Ninnis Trough, George V Coast (Maddison et al., 2006) and unlike rhythmically paired laminations which are found in postglacial sediments from Palmer Deep, Antarctic Peninsula (Leventer et al., 2002; Maddison et al., 2005; Pike et al., 2002) and Iceberg Alley, Mac.Robertson Shelf (Stickley et al., 2005) (Figure 1a). The gradational nature of lamina boundaries implies that there was gradual, not abrupt, change in productivity and sedimentation through the year. This is likely to be a result of the open trough geometry (Leventer et al., 2006). The presence of laminae throughout the core indicates that there was minimal or no bioturbation, probably as a combination of low oxygen conditions in the trough and ultra-high-sedimentation rates (2 cm/yr in the interval investigated). In the following discussion, water column (e.g. water column stability, nutrient levels) and sea ice conditions which prevailed at the time of deposition are determined from diatom assemblages present in the different lamina types.
Spring: Laminae characterized by CRS (LT A1), by CRS and Fragilariopsis spp. (LT A2) and by Fragilariopsis spp. (LT A3)
The proposed spring lamina types have similar diatom assemblages dominated by varying amounts of CRS and Fragilariopsis spp. (F. curta, F. cylindrus and F. rhombica). Hyalochaete Chaetoceros is one of the most abundant genera in the modern ocean (Armand et al., 2005), favouring near coastal environments (Zielinski and Gersonde, 1997) and is indicative of high primary productivity (Donegan and Schrader, 1982). High concentrations of CRS in Antarctic sediments are believed to be a result of well-stratified waters, created by austral spring sea ice melt (Crosta et al., 1997; Leventer, 1991). Leventer (1991, 1992) has suggested that CRS formation may be triggered by reduced salinity or depleted nitrogen near the sea ice edge. F. curta and F. cylindrus are both associated with pack and fast ice, with F. curta being the most common species found in this environment (Garrison, 1991). F. curta and F. cylindrus have also been observed in high abundances in the water column near melting sea ice edges (Dierssen et al., 2002; Fryxell, 1989; Kang and Fryxell, 1992, 1993; Leventer and Dunbar, 1996). F. rhombica is a sub-dominant species in these three lamina types and the relative abundance is comparable with modern abundances in the region of the core site (Leventer, 1992). F. rhombica is also associated with pack or fast ice (Garrison, 1991; Garrison and Buck, 1989; Garrison et al., 1982, 1983; Gersonde, 1986; Horner, 1985; Krebs et al., 1987). Low, but persistent abundances of Porosira glacialis RS are present in these lamina types. The exact conditions that promote resting spore formation are not known, but it has been suggested that P. glacialis share similar growth requirements to Thalassiosira antarctica and therefore form as a result of environmental stress induced by sea ice formation and decreasing light levels in the Autumn (Maddison et al., 2006; Pike et al., 2009; Stickley et al., 2005). The vegetative form of P. glacialis is associated with waters adjacent to the coast or sea ice (Hasle, 1973) but has not been observed to live within sea ice (Garrison, 1991; Horner, 1985; Watanabe, 1988).
High abundances of CRS, F. curta and F. cylindrus in these lamina types are ascribed to spring melt-induced productivity at the sea ice edge. The change in dominance of CRS and Fragilariopsis spp. between lamina types is the result of subtle environmental adjustments e.g. rapidity of sea ice edge break up, proximity of sea ice edge, and nutrient levels in the water column. The persistent presence of P. glacialis RS in the spring laminae is interpreted as the release of diatoms that were trapped in sea ice at the end of a previous growing season. The rare terrigenous forms of these lamina types are most probably a result of early season hemipelagic suspension or bottom transport of terrigenous material from the coast (Denis et al., 2009a; Dunbar et al., 1985; Presti et al., 2003). These mechanisms cannot be distinguished from examining the sedimentary fabric in BSEI.
Late spring/early summer: Laminae characterized by Corethron pennatum and Rhizosolenia spp. (LT B1), by Corethron pennatum (LT B2) and by Rhizosolenia spp. (LT B3)
Late spring/early summer lamina types have comparable diatom assemblages that are visually dominated by Corethron pennatum and/or Rhizosolenia spp. in BSEI and SEI analyses because of their large size making them conspicuous. C. pennatum is a cosmopolitan, widespread species (e.g. Fenner et al., 1976; Fryxell and Hasle, 1971; Leventer et al., 1993) with a preference for ice-free open waters (Fryxell and Hasle, 1971; Makarov, 1984). C. pennatum cells are capable of positive buoyancy (Crawford, 1995) which may allow them to exploit a well-stratified water column (Leventer et al., 2002). Mass sinking of C. pennatum blooms is thought to occur when the water column destabilises (Kemp et al., 2000). Rhizosolenia spp. also bloom in open water conditions with minimal sea ice influence (Alldredge and Silver, 1982; Harbison et al., 1977; Kemp et al., 1999) and migrate up and down within the water column exploiting different environments (Villareal et al., 1993, 1996). R. antennata f. semispina, the most common Rhizosolenia species in the late spring/early summer laminae, is cosmopolitan and can be found between the sea ice zone and the Subantarctic Zone (Fenner et al., 1976). Pseudo-nitzschia turgidula, also present in these lamina types, favours relatively warm and ice-free waters and has not been observed living in sea ice unlike other Antarctic Pseudo-nitzschia spp. (Almandoz et al., 2008; Hesgeth and von Quillfeldt, 2002). Proboscia inermis is only common in laminae characterized by Rhizosolenia spp. Apart from the taxonomy of P. inermis (Jordan et al., 1991), not much is known about its ecological preferences. This co-occurrence with R. antennata f. semispina has also been observed at other East Antarctic Margin sites (Maddison et al., 2006; Stickley et al., 2005) and has been used to infer that P. inermis prefers open waters with little sea ice influence (Stickley et al., 2005). Leventer et al. (2002) suggests that Rhizosolenia spp. and Proboscia spp. are both able to migrate below the thermocline to take advantage of nutrients in the water column at depth.
In terms of relative diatom abundance, CRS, F. curta and F. cylindrus dominate these late spring/early summer laminae similar to the three spring lamina types (A1–3), however, there are less F. curta and more F. cylindrus in the late spring/early summer. Laminae characterized by C. pennatum and Rhizosolenia spp. have previously been observed in several circum-Antarctic locations (Bahk et al., 2003; Leventer and Dunbar, 1996; Leventer et al., 1993; Maddison et al., 2006; Pudsey, 1990; Stickley et al., 2005; Taylor and Sjunneskog, 2002). These studies have attributed these laminae to early season warmth creating a meltwater-stabilised water column with little sea ice influence. Leventer et al. (1993, 1996) and Maddison et al. (2006) conducted quantitative diatom abundance analyses on these lamina types found in Granite Harbour, Palmer Deep and Mertz Ninnis Trough. In these studies the relative abundance of C. pennatum and Rhizosolenia spp. were much higher than in similar DUT laminae, suggesting that the water column was more weakly stratified above DUT than in those other locations. Laminae with high relative abundance of Rhizosolenia spp. in DUT would have formed beneath comparatively more oligotrophic surface waters (Kemp et al., 2000; Stickley et al., 2005).
The diatom assemblage within these three lamina types suggests that the biogenic forms were deposited in late spring/early summer. There are only two occurrences of the terrigenous form of laminae characterized by Rhizosolenia spp. and those occur in the uppermost part of the lamina succession, most likely a result of continued water column stratification in the late summer when terrigenous input to the sediments increases as a result of summer glacial melting (Denis et al., 2009a).
Summer/late summer: Mixed diatom assemblage (MDA) laminae (LT C(B and T))
CRS and Fragilariopsis spp. dominate the quantitative diatom assemblage of biogenic and terrigenous MDA laminae in a similar manner to spring and late spring/early summer lamination types, however, BSEI analyses have provided us with an opportunity to assess more subtle changes in diatom assemblages. Stellarima microtrias and Porosira glacialis are reported to prefer open water conditions in the summer, when there is <30% sea ice concentration (Armand et al., 2005). Rhizosolenia spp. (particularly R. antennata f. semispina), Corethron pennatum and Thalassiosira spp. also indicate that open water conditions prevailed prior to the deposition of this lamina type (Alldredge and Silver, 1982; Crosta et al., 2005a; Fenner et al., 1976; Fryxell and Hasle, 1971; Harbison et al., 1977; Makarov, 1984). Very little has been reported on the distribution or ecological preferences of Coscinodiscus bouvet or Trigonium arcticum. C. bouvet is considered to be a neritic species (Priddle and Thomas, 1989) and it has been suggested that T. arcticum lives epiphytically on macro-algae at water depths of 200–300 m (Hendey, 1937; Tomas, 1966). T. arcticum has also been observed in postglacial laminated sediments in the nearby Mertz Ninnis Trough (Figure 1a) and is suggested to have been swept off surrounding shallow banks into the trough by current action (Maddison et al., 2006).
In the seasonal succession of laminae, the biogenic form of MDA laminae usually occurs preceding the terrigenous form. This, combined with the ecological information above, suggests that biogenic MDA laminae were deposited in summer after the sea ice edge had retreated. The position of the terrigenous form in the uppermost part of the lamina succession, along with inferred open water conditions suggests that deposition occurred in summer/late summer. Diatom abundance in the terrigenous form is lower than in the biogenic form because of dilution by clays and silts from seasonal glacier melting, or sediment re-suspension events. Sedimentary fabric examination does not provide sufficient evidence to determine which source supplied the terrigenous material to the laminae.
Autumn: Laminae characterized by Stellarima microtrias RS, Porosira glacialis RS and Coscinodiscus bouvet (LT D)
Stellarima microtrias is associated with shelfal ice and surrounding waters (Hasle et al., 1988). In autumn, in the Weddell Sea, S. microtrias RS have been observed in high abundances under sea ice (Fryxell, 1989). Porosira glacialis is ecologically similar to T. antarctica (Pike et al., 2009) and forms resting spores as a result of autumnal sea ice formation, diminishing light levels and increasing water salinity (Maddison et al., 2006; Pike et al., 2009; Stickley et al., 2005). Coscinodiscus bouvet has been observed in coastal waters in the summer (Garibotti et al., 2005) and reports indicate that it can be transported considerable distances by surface currents (Priddle and Thomas, 1989). Thalassiosira spp. associated with summer open water conditions are also common in this lamina type. The most common Thalassiosira spp. in these laminae is T. poroseriata, however, very little is known about this species other than it is cosmopolitan (Hasle and Syvertsen, 1997) and is found in all sectors of the Southern Ocean (Johansen and Fryxell, 1985).
Environmental conditions inferred from the diatom assemblage and the position of this lamina type at the end of the seasonal succession suggests that these laminae were produced as a result of autumnal flux to the sediment. The biogenic form is most likely due to high diatom flux diluting the terrigenous input from hemipelagic re-suspension events (Denis et al., 2009a; Dunbar et al., 1985; Presti et al., 2003).
Autumn: Laminae characterized by Porosira glacialis RS (LT E)
Porosira glacialis RS are produced as a response to sea ice formation in autumn. The high relative abundance of the resting spores and the typical thinness of this lamina type suggest that relatively short, intense episodes of resting spore formation and deposition took place. Multiple occurrences in a year may suggest repeated sea ice edge advance and retreat over DUT. The abundance of P. glacialis has been observed to notably decrease when winter sea ice concentration is >85% (Armand et al., 2005; Pike et al., 2009). Years where laminae characterized by P. glacialis RS are not present could indicate that the sea ice margin re-advanced later in the year after the growing season, the winter sea ice concentration was exceptionally high or vegetative cell abundance in the summer water column was exceptionally low (Pike et al., 2009). The fewer biogenic forms of this lamina type most likely result from high diatom flux diluting the terrigenous grains.
The annual cycle
Cyclical changes between the seasonally constrained lamina types have allowed us to determine a typical annual sequence of flux in these sediments (Figure 4a). Not every lamina type is present in every annual deposit reflecting subtle differences in environmental conditions between years (Figures 1c and 4b).
As solar radiation increases in spring, sea ice starts to fragment and melt and the environment adjacent to the sea ice edge provides suitable conditions for blooms of Hyalochaete Chaetoceros spp. and sea ice-related Fragilariopsis species. Which taxa becomes dominant in the surface waters is controlled by subtle shifts in the stability of the water column, salinity levels and the degree and nature of sea ice fragmentation. Therefore, the sedimentary record comprises either laminae characterized by: (i) LT A1: CRS, a consequence of relatively early seasonal sea ice melt, creating a well-stratified water column with reduced salinity (Crosta et al., 1997; Leventer, 1991); (ii) LT A2: Fragilariopsis spp., a result of a pack or fast ice-dominated environment with some ice edge melting (e.g. Garrison, 1991; Leventer and Dunbar, 1996); or (iii) LT A3: CRS and Fragilariopsis spp., a product of a well-stratified water column influenced by sea ice.
In late spring/early summer, surface water conditions dictate whether one of two diatom assemblages thrives. First, early seasonal warmth, combined with stable open water conditions in front of the sea ice edge, provides an environment suitable for blooms of Corethron pennatum and Rhizosolenia spp. The dominance of either C. pennatum or Rhizosolenia spp. is dependent on nutrient levels in the water column (Kemp et al., 2000; Leventer et al., 2002; Stickley et al., 2005), with Rhizolsolenia spp. appearing to prefer slightly more oligotrophic conditions. These circumstances produce laminae characterized by: (i) B1: C. pennatum, (ii) B2: Rhizosolenia spp. or (iii) B3: Rhizosolenia spp. and C. pennatum. Second, and more commonly, surface waters are re-supplied with nutrients by mixing of the water column. This promotes a diverse diatom assemblage which results in LT C(B): MDA biogenic laminae. Later in the summer the MDA terrigenous laminae (LT C(T)) are produced either as a result of glacial melting increasing the supply of terrigenous material, decreasing diatom productivity or a sediment re-suspension event, increasing the relative proportion of terrigenous material in the sediment.
In autumn, as light levels fall, temperatures cool and sea ice forms, vegetative forms of Stellarima microtrias and Porosira glacialis produce resting spores and blooms of Coscinodiscus bouvet are advected into the region by surface currents. This results in a lamina characterized by S. microtrias RS, P. glacialis RS and C. bouvet (LT D) or a laminae characterized by P. glacialis RS (LT E). A transition from late summer/autumn laminae to spring/summer laminae indicates that there was very little, or no, deposition during the winter when the core site is covered by sea ice.
Holocene environments
The quantitative co-dominance of CRS and Fragilariopsis spp. (particularly F. curta and F. cylindrus) in all lamina types indicates the persistent influence of sea ice throughout much of the growing season in Dumont d’Urville Trough (DUT) between 3122 and 1136 cal. yr BP, signifying that the laminated sediments were deposited during the cool climatic phase known as the Neoglacial. Previous East Antarctic margin palaeoenvironmental studies have suggested that the Neoglacial commenced anywhere between ~4000 and ~1000 cal. yr BP (e.g. Cremer et al., 2003; Kulbe et al., 2001; Taylor and McMinn, 2002), however, our findings corroborate DUT studies that indicate the Neoglacial was established prior to 3000 cal. yr BP (e.g. Crosta, 2009; Crosta et al., 2005b, 2008). The laminated sediments from MD03-2597 contain higher average absolute abundance of diatoms than contemporaneous sediments from slightly further offshore DUT (MD03-2601, Crosta et al., 2005b; Denis et al., 2006). This higher absolute abundance is likely to be related to the more coastal nature of MD03-2597 (~30 km offshore compared with ~60 km for MD03-2601) combined with the fact that site MD03-2597 has a longer ice-free productivity season than MD03-2601 (Pike et al., 2009).
In a study based on the ratio between the relative abundance of Porosira glacialis RS and Thalssiosira antarctica RS in Holocene sediments, increases in winter sea ice concentration, cooler spring seasons with increased sea ice and slightly warmer autumn seasons with less sea ice were identified in the DUT region at approximately 2200 cal. yr BP (Pike et al., 2009). This change in environment coincides with the end of a long period of glacial advance in the region (Denis et al., 2009a) and a decrease in occurrences of laminae characterized by Corethron pennatum and/or Rhizosolenia spp. (warm, open water diatoms) and laminae characterized by P. glacialis RS after 2475 cal. yr BP (Figure 4b). However, the highest relative abundance of P. glacialis RS occurs at 1730 cal. yr BP (a period characterized by a lack of P. glacialis RS laminae) which corresponds with a maximum in the P. glacialis RS:T. antarctica RS ratio observed in Pike et al. (2009). This apparent dichotomy, decrease in occurrence of laminae characterized by P. glacialis RS despite an increase in overall abundance of individual frustules, reflects the change to slightly warmer autumn conditions with later onset of sea ice production. Cooler spring conditions promote higher abundances of the vegetative cell standing stock of P. glacialis, thus providing the potential for greater resting spore production, hence flux to the sediment (Pike et al., 2009). The longer, warmer autumn would inhibit the usual abrupt resting spore formation event, leading instead to an extended period of resting spore formation and lack of lamina formation in the sediment. This scenario is supported by other diatom records from the DUT that identify decreasing abundance of summer diatoms (Denis et al., 2009b) and increasing surface water stratification (Crosta et al., 2008) after 2000 cal. yr BP. After 2475 cal. yr BP laminae characterized by S. microtrias RS, P. glacialis RS and C. bouvet are more common which further indicates that less sea ice was present in the autumn (Figure 4b). By 1149 cal. yr BP only laminae characterized by CRS, CRS and Fragilariopsis spp., mixed diatom assemblage and P. glacialis RS were present (Figures 1c and 4b), which suggests further cooling and more sea ice influence throughout the year. Two further glacier advances were noted between 2000 and 1000 cal. yr BP and modelling data that shows ice-free season duration also decreased from approximately 2000 yr BP (Denis et al., 2009a).
Diatom and sedimentary lamina analyses, supported by additional environmental data from the DUT, suggest that sea ice conditions were not constant in this region during the late-Holocene Neoglacial climate period. Early seasonal warmth and associated early sea ice melt occurred from 3122 to 2475 cal. yr BP. Subsequently sea ice melt occurred later in spring and autumn became warmer. By 1149 cal. yr BP sea ice influence had increased year round. This undoubtedly is a result of climatic fluctuations that are recorded in deuterium isotope records from several East Antarctic ice cores (Masson et al., 2000) as well as local glacier dynamics (Denis et al., 2009a).
Conclusions
The late-Holocene laminated sediments from Dumont d’Urville Trough provide an important high-resolution archive of climate, ocean and sea ice conditions between 3122 and 1136 cal. yr BP. A total of 318 laminae were identified which comprised 120 years of deposition. Nine lamina types were interpreted as seasonal productivity events. The quantitative diatom assemblage of each lamina type was dominated by CRS and Fragilariopsis spp. indicating that this region, in the late Holocene (Neoglacial), was influenced by sea ice all year round. However, environmental conditions were not constant at this site during the Neoglacial and the diatoms and laminations record the deterioration from a relatively warm period, 3122–2475 cal. yr BP, with early seasonal warmth and melting of the sea ice, though a cooling period with increasingly icy spring conditions and slightly warmer autumns (post-2475 cal. yr BP), to year-round increased sea ice influence by 1136 cal. yr BP. This, and other studies of seasonal-scale sea ice records from the Dumont d’Urville Trough (e.g. Denis et al., 2006) will aid in realising the potential of recent (February 2010) IODP drilling through the Holocene sediments of the Adélie Basin (Expedition-318-Scientists, 2010).
Footnotes
Acknowledgements
EJM thanks Lawrence Badham for preparing the polished thin sections and Peter Fisher for instruction on the use of SEM facilities (Cardiff University). We thank Amy Leventer and Xavier Crosta for constructive discussions and Claire Allen and one anonymous reviewer for valuable suggestions which have improved the manuscript. We also thank the staff and scientific party aboard the R/V Marion Dufresne during the MD130 Images X cruise (CADO) and the staff of the Antarctic Research Facility (Florida State University) for their help with sampling and core photography.
Funding
This research was funded by Natural Environment Research Council (NERC) postgraduate research studentship NER/S/A/2002/10350 (EJM at Cardiff University), NERC research grant NER/A/S/2001/01106 (JP), and National Science Foundation (NSF) research grant 9909837 (RD).