
Abstract
Species distribution models are widely used by ecologists to estimate the relationship between environmental predictors and species presence and abundance records. In this paper, we use compiled faunal assemblage records from archaeological sites located across southwest Asia and southeast Europe to estimate and to compare the biogeography of ancient wild and early domestic cattle (Bos primigenius and Bos taurus). We estimate the contribution of multiple environmental parameters on the explanation of variation in abundance of cattle remains from archaeological sites, and find that annual precipitation and maximum annual temperature are significant predictors of abundance. We then formulate, test, and confirm a hypothesis that states the process of cattle domestication involves a change in the types of environmental ranges in which cattle exploitation occurred by applying a species distribution model to presence-only data of wild and domestic cattle. Our results show that there is an expansion of cattle rearing in more temperate environments, which is a defining characteristic of the European early Neolithic.
Keywords
Introduction
What role does the environment play in explaining regional variability in subsistence behaviour? It is understood that environmental differences at continental scales structure the type and distribution of resources and thus impact settlement and mobility strategies (e.g. Binford, 2001), but there have been few attempts to predict more precisely how environmental variability may explain resource use strategies at smaller scales. This paper considers this question from the perspective of spatial and temporal variability in the use of wild and domestic cattle in southwest Asia and southeast Europe. More specifically, we use statistical and ecological models to test a hypothesis that predicts that regional variance in cattle use patterns can be accounted for by differences in rainfall, temperature and hydrology, and that between the earlier and later Neolithic, cattle use expands into new niches as humans play a greater role in cattle management. Through this, we provide additional insight into the biogeography of early cattle-rearing, and how and to what extent the structure of the environment influences resource choices. It is important to highlight our expectation that a significant amount of variability will not be accounted for by environmental factors and consequently that resource use patterning is likely being driven by a range of cultural decisions that are historically and locally contingent.
The development of cattle pastoralism during the 11th and 10th millennia BP represents a major shift in the development of the ‘Neolithic package’. In addition to their meat, cattle provide a source of social capital and mobile wealth alongside a range of new resources that become important during the Neolithic and into the Bronze Age, including hide, blood, milk, dung and traction. Yet, whilst the spread of domestic cattle appears to have been rapid and widespread throughout the Levant and eastern Mediterranean, their incorporation into the pre-existing socio-economic system was neither inevitable, nor straightforward (Arbuckle and Makarewicz, 2009). Wild cattle, or aurochs, occur in a range of symbolic representations and ritual deposits throughout the early part of the Neolithic (i.e. Pre-Pottery Neolithic A (PPNA) and Pre-Pottery Neolithic B (PPNB) – see Table 1 for periodization and calibrated date ranges), for example, at Çatalhöyük (Hodder, 2006; Mellaart, 1967), Kfar Hahoresh (Horwitz and Goring-Morris, 2004) and Göbekli Tepe (Peters and Schmidt, 2004) (see Figure 1 for key site locations). Remains of early domestic cattle, meanwhile, are first found in the northern Levant by at least the middle PPNB (MPPNB) and increase in occurrence across the Levant through the late PPNB (LPPNB). However, domesticates did not entirely replace wild cattle in either their social or economic role. At the LPPNB site of Basta, in the southern Levant, for example, Horwitz et al. (1999) have identified both wild and domestic cattle, indicating that well-established hunting strategies persisted despite the introduction of domestic resources (cf. Conolly et al., 2011). The aim of this paper is to develop an understanding of the factors influencing the observed regional variation in the use of cattle, and to build a better appreciation of the ecological factors that may be influencing the development and spread of cattle pastoralism.
Archaeological chronology and approximate calibrated kilo-years before present (kyr cal. BP).
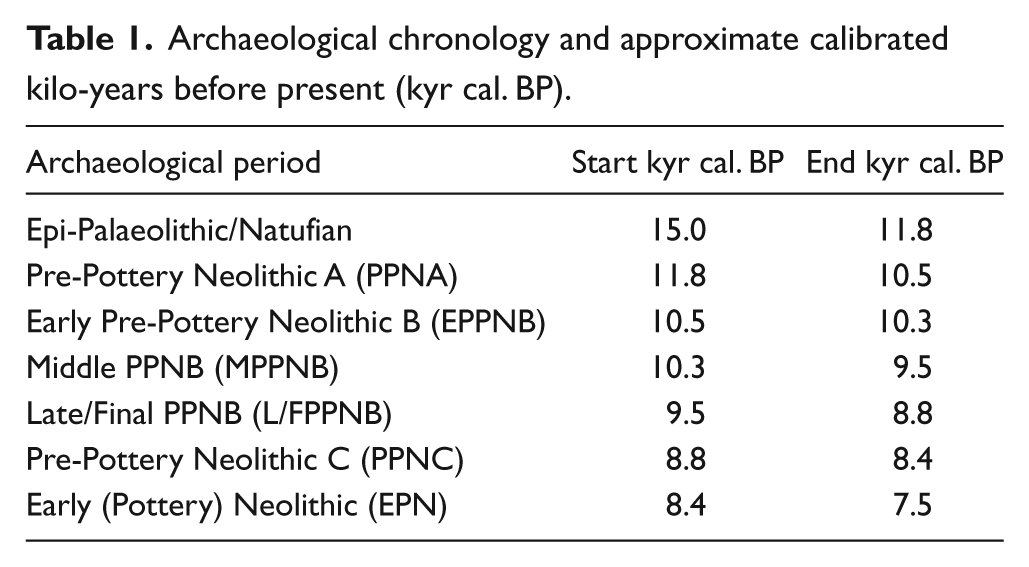

Key sites referred to in the text. 1: Argissa Magoula; 2: Kazanluk; 3: Ovčarovo Gorata; 4: Mentese; 5: Hacilar; 6: Höyücek; 7: Çatalhöyük; 8: Ortos & Ais Yorkis; 9: Tenta; 10: Ras Shamra; 11: Mezraa-Teleilat; 12: Halula; 13: Nahr el Homr; 14: Mureybet; 15: Dja’de; 16: Hayaz Tepe & Gritille Höyük; 17: Göbekli Tepe; 18: Gürcütepe; 19: Çayönü Tepesi; 20: Körtik Tepe; 21: Tell es Sin; 22: Bouqras; 23: Qermez Dere; 24: M’lefaat; 25: Ali Kosh; 26: Sarab; 27: Hagoshrim; 28: Kfar Hahoresh & Munhatta; 29: Tsur Nathan; 30: Abu Gosh & Motza; 31: Ashqelon; 32: Zahrat edh-Dhra; 33: Azraq; 34: Jebel Naja.
Cattle ecology
Cattle and sheep tend to derive the majority of their forage energy from grazing on seasonally available herbaceous grasses as opposed to more perennial woody species, which are the preferred forage of goats (Coughenour et al., 1985: 620–621; Dahl and Hjort, 1976: 244; Russell 1988: 58). However, unlike sheep and goats cattle are generally unable to meet their water requirements from plants and require permanent standing water (Behnke, 1980: 26: Dahl and Hjort, 1976: 239; Russell, 1988: 57). In addition, most modern domestic cattle maintain poor thermal stability when under heat stress (Bianca, 1968: 110; MacFarlane, 1968: 172; Schmidt-Nielsen, 1979: 77), and thus tend to be regarded as high maintenance animals (Russell, 1988: 55–58). There are, however, a number of cattle breeds that demonstrate a greater tolerance for dry adapted plant species and increased efficiency of water use. Corriente cattle, for example, are widespread throughout much of southwest America and Mexico, where they have adapted to the arid environment through aggressive browsing (Russell et al., 2000: 2314). As with all ruminants, climate, nutrition, and level of management will also effect the reproductive efficiency of cattle (Vanderplassche, 1982). Thus base ecological parameters clearly play a central role in the maintenance and breeding success of modern domestic cattle, but it is an open question as to what types and ranges of ecological parameters wild and early domestic cattle were adapted to in the late Pleistocene and early Holocene.
The distribution of southwest (SW) Asian aurochs (B. primigenius) from Late-Pleistocene and early-Holocene sites in the 12th and 11th millennia BP stretches along much of the Levantine coast and its hinterland and along the foothills of the Zagros, indicating adaptation to moist through semi-arid woodland and park woodland conditions that existed in the coastal, littoral and foothill regions of the Mediterranean in the early Holocene (Roberts et al., 2011; Uerpmann, 1987: 71–72). This suggests that SW Asian aurochs exploited a different, or at least broader, range of niches than observed for modern breeds of taurine cattle (B. taurus taurus). Correspondingly, recent attempts at reconstructing the ecology of European aurochs suggest that unlike domestic cattle, they were, in fact, a more wetland-adapted species, favouring marshes and areas of open marshland in forests (van Vuure, 2002, 2005). These observations require us to consider more critically our predictions for the range of environmental tolerance in the herds of wild and domestic taurine cattle in SW Asia, which is essential for developing a better understanding of the ecological context of animal domestication. The remainder of this paper takes a step towards that goal and examines the distribution of early cattle remains from archaeological sites in SW Asia, the eastern Mediterranean and southeast Europe in order to assess the relative importance of ecological constraints on the spread and adoption rate of domestic cattle.
Cattle pastoralism: Archaeological and genetic evidence
Recent work on the origins of domestic livestock distinguishes between the archaeozoological identification of fully domesticated animals and the ethno-archaeological, or more specifically ethno-zoological, identification of pre-domestic animal husbandry (Vigne et al., 2003; Zeder, 2008). This issue has come under increasing scrutiny as advances in analytical techniques provide greater insight into the stages preceding full-scale domestication. In particular, increasing chronological resolution and improved methods of demographic profiling have demonstrated that early animal management does not immediately lead to observable gross morphological change in bones and teeth, the traditional marker usually associated with animal domestication.
At Tell Mureybet in the middle Euphrates Ducos (1978a) has argued that an increase in adult Bos remains during mid-11th millennium (EPPNB) indicates a shift towards pre-domestic animal husbandry, or ‘proto-élevage’ (Ducos, 1993). This term has caused some consternation among researchers – Peters et al. (1999: 30), for example, refer to the term as misleading, advocating instead the term ‘selective hunting’ to describe herd management or selective culling preceding gross morphological change (Helmer, 1994; Vigne et al., 2000; Zeder, 2008). Recent work on Cyprus lends considerable support to the role of human in cattle management prior to domestication. At Shillourokambos, cattle – which are not an endemic species to Cyprus – are present in the earliest levels, accounting for ~10% of the mammalian remains (Vigne and Buitenhuis, 1999). The Shillourokambos cattle are equivalent in size to the late-12th/early-11th millennium cal. BP aurochs from the Middle Euphrates, the latter also demonstrating considerable metric range (Vigne et al., 2000). The presence of vertebrae and foot bones also suggest close-to-site or on-site butchery, negating the importation of butchered aurochs body parts from mainland SW Asia. Hence, cattle appear to have been imported as livestock into Cyprus during the mid-11th millennium, and whilst it is possible that they may have been introduced as hunting stock, it has been argued that they are more likely that they represent the remains of individuals from a managed rather than wild herd (Vigne and Buitenhuis, 1999).
To date, the earliest published evidence for morphologically confirmed domestic cattle on the basis of size diminution comes from the northern Levant, at Tell Halula and Dja’de (Helmer et al. 2005; Peters et al., 1999), dating to the late 11th millennium cal. BP (MPPNB). From the mid 10th millennium (LPPNB), domestic cattle, as demonstrated by a distinct size diminution and shift in the age and sex ratios, are found throughout the northern Levant at sites such as Gürcütepe and Hayaz Tepe (Peters et al., 1999: 40), to the south at Bouqras and Tell es Sinn (Clason, 1979/1980) and to the west at the coastal site of Ras Shamra (Helmer, 1989, 1994).
This wide and varied pattern of contexts in which early domestic cattle are found somewhat tempers any introduction hypotheses (e.g. Becker, 2002; Horwitz et al., 1999; Peters et al., 1999), and raises the question: Did cattle undergo a single domestication event in the northern Levant, or were distinct populations of wild aurochs independently domesticated in more than one SW Asian location? Genetic data clearly demonstrate an expansion of domestic cattle out of the Near East between 9000 and 11,000 BP (Achilli et al., 2008; Beja-Pereira et al., 2006; Cymbron et al., 2005; Troy et al., 2001). However, on the basis of this evidence, the question of whether domestication occurred as a result of a single event within SW Asia is as yet unresolved. Five different domestic mtDNA haplotypes within haplogroup T have been identified for taurine cattle, with at least three and possibly all five originating in the Near East (Achilli et al., 2009; Troy et al., 2001). Beja-Pereira et al. (2006) support a demographic expansion of cattle from a small Near Eastern population. Likewise, Achilli et al. (2008) propose that the sequence divergence of the T haplogroup indicates a narrow bottleneck, with the mtDNA falling into subclades that support a single founder sequence for taurine cattle in the Fertile Crescent. However, the authors do note that several founder sequences with little variation could result in the same pattern (Achilli et al., 2008: 158). Furthermore, recent work on Y chromosome haplotypes indicates a greater contribution of male aurochs DNA to the genetic pool, suggesting at least some introgression of domestic stock with wild populations. In Europe, there is some evidence for backcrossing and hybridization between domestic female stock and wild males (Achilli et al., 2009; Götherström et al., 2005), but this hypothesis is not supported in other analyses (Bollongino et al., 2008). As it is likely for early pastoralists to have maintained a degree of residential mobility, especially in more marginal areas to protect vegetation from overgrazing, it is not unlikely that domestic female stock would have come into contact with extant aurochs populations.
Despite the obvious nutritional advantages that large Bovidae offer to hunter-gatherers and early farmers, our comparative analysis shows that the frequency of all cattle (Bos sp.) recorded in archaeological assemblages stays relatively low and constant in representation over the duration of the earlier Neolithic (Figure 2). Cattle constitute on average between 1% and 5% total Number of Identified Specimens (NISP) in faunal assemblages dating to the 11th and 10th millennia cal. BP, but in the 9th millennium rise dramatically in number (note the distribution of these are positively skewed so Figure 2 uses a arcsine transformation to normalize frequencies in order to facilitate comparisons). The increase, corresponding to the latter part of the Aceramic Neolithic through the Pottery Neolithic, is largely being driven by the increase in the use of domestic cattle in Anatolia and SE Europe, at sites such as Ovčarova Gorata in Bulgaria (Todorova and Vajsov, 1993) and Fikirtepe in Turkey (Boessneck and von den Driesch, 1979), which have recorded well over 30% Bos remains by total NISP. At these sites cattle may be over-represented because faunal samples were hand-collected rather than sieved. However, this is true for many assemblages in the Levantine region, but even so they do not show such dramatic increases. For example, between the earlier and later phases of Halula in Syria (Saña-Segui, 1999) Bos increases from about 15% to about 20% of total NISP, and at Sarab in Iraq (Bokonyi, 1977) Bos is only 1.2%, and more arid sites such as Jebal Naja in Jordan (Martin, 1994) have no recorded cattle. Figure 3 shows the percentage of total NISP (hereafter %NISP) of wild and domestic cattle, as defined by the original analyst, plotted by approximate date. We accept that this latter graph is also problematic given that there is constant revision of how wild from domestic cattle should be distinguished, but the general pattern is likely to hold, namely that from about 11,800 to 9000 cal. BP, there is a relatively slow increase in the use of domestic cattle, although wild cattle remains are always common. After 9000 cal. BP, wild and domestic species are used in roughly equal proportions, and after 8500 cal. BP (corresponding to the earliest introduction and development of the Neolithic in SE Europe), domestic cattle increase exponentially. This demonstrates that following the appearance of domestic cattle it takes as long as 2000 years before they begin to fix within early agricultural communities as the dominant domestic animal species.
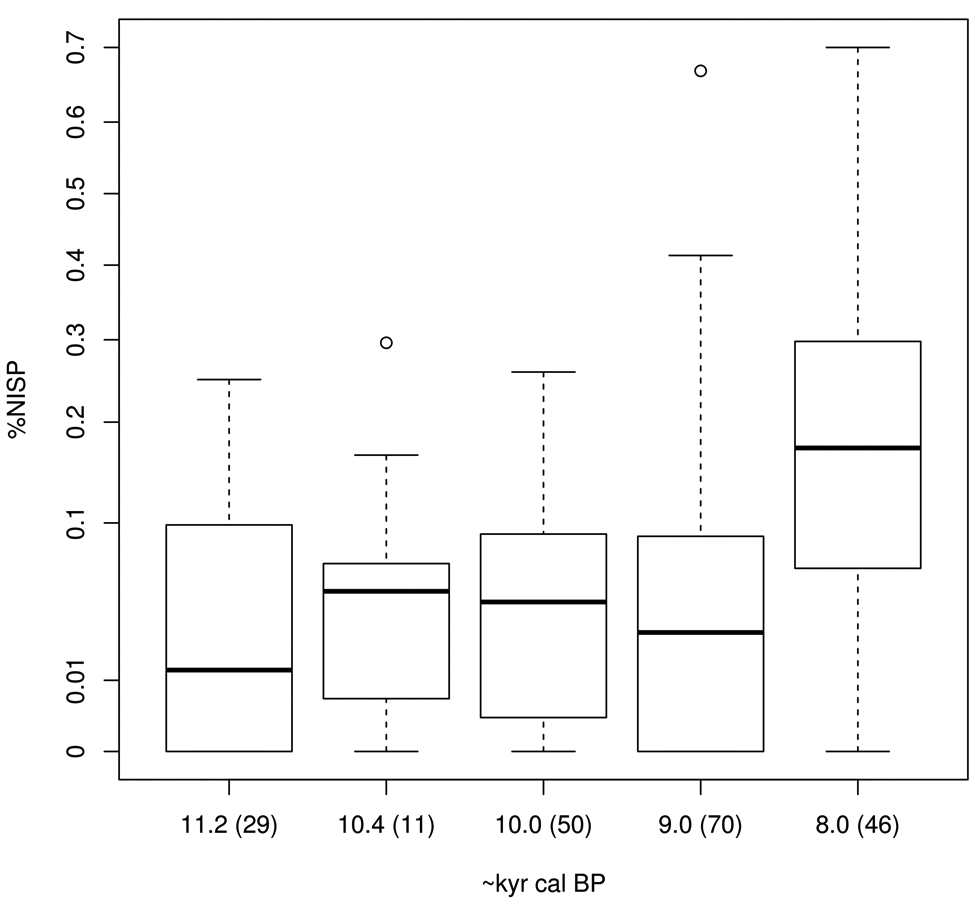
Boxplot of cattle frequencies from phases between the 12th and 8th millennium BP (numbers of phases given in parentheses following approximate date).

Mean %NISP for ‘wild’ and ‘domestic’ cattle between the 12th and 8th millennium BP (see Figure 2 for phase counts). Note that this distinction is problematic, and mostly reflects differing proportions of large (~wild) and small (~domestic) cattle.
It is also important to note that there is considerable regional variation in these patterns reflecting important differences in faunal exploitation behaviour, as documented in Conolly et al. (2011). In the remainder of this paper, we examine the factors influencing the observed differences in the frequency of cattle use by considering the environmental contexts of the sites that have recorded and published zooarchaeological assemblages.
Hypotheses
Our earlier work established that there is significant regional patterning in the different amounts and proportions of animal species recorded at Neolithic archaeological sites in SW Asia (Conolly et al., 2011). In general there is a clear separation between sites in the Mediterranean and southern Levantine region and sites in the northern Levant and Anatolia – with the former dominated by greater proportions of wild taxa throughout the PPN and the latter by an early reliance on domestic animals. We hypothesized that this was likely being driven to some degree by the significant biogeographical differences in the study region (i.e. annual precipitation ranging from <10 mm to greater than 1500 mm; elevation varying from c. 400 m below to nearly 5000 m above sea level, and with major hydrological systems cutting through highly variable landforms). We proposed that the variation in proportion of species represented at sites is thus a reflection of the density of those species available in the ecological niches available for exploitation by the inhabitants of sites, as has been similarly proposed for the Palaeolithic (cf. Morin, 2008). This hypothesis received tentative support from initial biogeographic models (Conolly et al., 2010), suggesting that upwards of 40% of the identified variability in cattle Number of Identified Specimens (NISP) recorded at over 100 sites in SW Asia could be accounted for by the interaction of three critical environmental variables: elevation, rainfall and temperature. The purpose of this section of the paper is to revisit this proposition in a more rigorous manner to assess: (i) the degree to which environmental variables are able to account for spatial variation in the exploitation pattern of cattle at Neolithic sites in SW Asia; and (ii) if the strength of the correlation between exploitation patterns and environmental variables changes between the early and later part of the Aceramic Neolithic; and (iii) how the environments differ between sites using wild versus domestic cattle. As noted in the introduction, we expect that the majority of observed variability in cattle use will not be accounted for by environmental factors and other factors are likely more important. However, we should not begin testing for causation against cultural or other determinants without also establishing how and to what extent environments are driving these observed patterns.
These general questions lead us to develop two hypotheses that can be formally tested. The first hypothesis proposes that regional differences in base environmental conditions (e.g. rainfall, temperature and their seasonal fluctuations) will explain a significant proportion of the documented variability in the proportion of cattle bones identified from archaeological sites across SW Asia. The second hypothesis is that these variables will differ between the earlier Neolithic (PPNA to LPPNB: c. 11.8 to 9.0 kyr cal. BP) and later Neolithic (PPNC to early PN: c. 9.0 kyr cal. BP to 7.5 kyr cal. BP). The prediction is that because there is an increasing propensity for humans to be more actively involved in the management of cattle in the later Neolithic, there should be a regional pattern that shows that cattle are being exploited in different types of environments over time. More specifically, we should see that some environnments become increasingly associated with domestic cattle, reflecting an increase in cattle use in those niches most suitable for cattle rearing.
Data set compilation
Zooarchaeological data
The source of our data for testing these hypotheses comes from the Origins and Spread of Stock Keeping (OSSK) project data base. The data base contains the records of absolute number of identified specimens (NISP) by taxa and chronological phase from over 300 sites in SW Asia and SE Europe. The archaeological data set for this study consists of presence or numbers of identified specimens (NISP) of vertebrate taxa from 135 Neolithic sites (256 chronologically separate phases) from SW Asia (Jordan, Syria, Israel, Palestinian Territories, Sinai, Iran, Iraq and Turkey), the eastern Mediterranean (Cyprus and Crete) and SE Europe (Greece and Bulgaria), listed in Table 2. The geographic distribution of these sites is shown in Figure 4. For 96 of these sites (185 phases) the full suite of potentially exploitable fauna published at each site was recorded, including wild and domestic mammals, birds, fish, molluscs and crustaceans. These taxa were entered into our data base at the level of identification published by the original analysts, whether to species or genera, or at the more general level of family (or ‘type’), or according to body size (e.g. large/small mammal). Age and sex trends, biometric information, pathological signatures, and body part distribution were also recorded where available.
Sites used in the analysis and whether abundance (A) or presence-only (P) species data available.

Site locations used in this analysis. For site names please refer to Table 2. A compiled list of references of faunal assemblages used in this and previous studies is available for download as supplementary data in Conolly et al. (2011) or by contacting the corresponding author.
Of the 256 archaeological phases used in this analysis 33 record no Bos remains, and 71 record Bos only as present, with no quantification of abundance. Of the remaining phases, 63 possess between 0.1 and 9.9% cattle by total mammal NISP, 66 between 10.0% and 49.9%, and there are ten phases (from six sites: Atlit Yam, Fikirtepe, Hacilar, Koprivec, Mentese and Ovčarova Gorata), where more than 50% of the total NISP is reported to be cattle. Note that Atlit Yam and Hacilar have very low sample sizes (<500 total NISP) and their species abundance measures are thus problematic. As we explain below, sites with small sample sizes are not included in the statistical analysis.
For the purposes of this paper we have reduced the data in two important ways. First, for the linear models we sidestepped the fundamental but complicated issue of what constitutes domestic status cattle versus managed or wild cattle by simply summing species-level data (i.e. B. primigenius or B. taurus) to the genus level. We excluded bones recorded only to family (Bovidae) as this may describe non-cattle remains, and excluded all records that recorded data to species presence only. We then calculated the proportion of Bos as a percentage of the total number of identified mammal specimens (hereafter %MNISP) to the level of family or higher, and used this measure as an index for cattle abundance at a particular location. A site or site phase with taxa recorded as presence-only was excluded from the analysis as we could not accurately calculate MNISP values.
Second, the data set was divided into two subsamples – the first containing phases from 11.8 to approximately 9.0 ka cal. BP (calibrated radiocarbon kiloyears BP, hereafter for simplicity ka), which includes the PPNA through the late and final PPNB. The second subsample combines phases from 9.0 ka to about 7.5 ka or the PPNC through to the early pottery Neolithic and the first phases of farming communities in eastern Europe. The basis for this split was that the early set of phases are primarily characterized by the use of wild cattle, whereas the second set of phases often have a mix of wild and domestic cattle. Dividing the data this way allows us to examine whether there are changes in the environmental contexts associated with chronology. To reduce the inflationary effects of double-counting, multiple phases from a single site were amalgamated into a single site record by summing NISP counts. Finally, all records with fewer than 500 total mammal NISP were excluded, resulting 34 locations with abundance records for the earlier period, and 38 for the later (Table 3).
Bos abundance and environmental co-variates

Second, for the species distribution models we used both abundance and presence records, retained the original analysts’ assessment of the domestic status of cattle (i.e. wild or domestic), and assigned a presence score as explained further below. When the domestic status of cattle in an entire phase or archaeological site was uncertain (e.g. where cattle were recorded as wild or domestic, or wild/domestic, or status not recorded, as Bos sp.), the entire site or phase was excluded from further analysis. We acknowledge that the operational definition of what constitutes ‘wild’ versus ‘domestic’ varies by analyst, is subject to revision, and is not necessarily binary. However, at the large scales in which we are working, we anticipate that variation in definitional and/or observational practices will not undermine the general patterns we aim to identify. In our sample there are 118 sites or phases where the presence of wild cattle is recorded, and 101 where domestic cattle are recorded. A total of 45 phases record the presence of both species in use at the same period. Where there were two or more phases at a single site location recording the same species of cattle they were reduced to a single observation point. The final data consists of 62 unique locations with wild cattle, and 49 unique locations with domestic cattle.
Environmental data
We used modern (averaged records from 1950 to 2000) rather than reconstructed environmental data in this analysis. There are three reasons for this. First, the spatial scale required to distinguish the potential environmental correlates of differences in cattle representation at sites (for example) either side of the Jordan Valley means that we need relatively high-resolution data that exceed that of any available reconstructed environmental model for SW Asia. Second, modern environmental data provide us with a range of baseline variables to use, including rainfall and rainfall periodicity and seasonal variations in temperature that are known to impact vegetation and thus faunal biogeography. Third, because we are not interested in reconstructing the absolute climatic predictors of cattle abundance but simply which variables strongly co-vary with abundance, and how these vary relatively over time and space, the absolute changes between modern and mid-Holocene environments do not concern us. We are assuming with caution that the relative differences between regions have remained stable, and thus our measures of differences between regions also reflect past relative differences, despite the evidence for overall increasing regional aridity between the early and mid Holocene (Roberts et al., 2011).
Table 4 lists the independent environmental variables we used in this study together with their source. Second order variables that describe the coefficient of variation (seasonality) of first order variables are also listed. All data sets are publicly available, and can also be obtained by contacting the lead author.
Independent environmental variables (source: SRTMv2, WorldClim, and Natural Earth).

Method: Species distribution modelling
Species distribution models (SDMs) are widely used by ecologists to estimate the relationship between environmental predictors and species presence/absence or abundance records (Franklin, 2009). There are several different approaches to modelling appropriate for different objectives and types of data (for a recent review and comparison, see Tognelli et al., 2009). For this paper we have used two complementary approaches: Maximum Entropy modelling of presence-only data (MaxEnt version 3.3.3e, Dudik et al., 2010) and linear modelling on abundance proxy data. MaxEnt has been demonstrated to perform well for obtaining estimates for the spatial distribution of species based on environmental covariates using presence-only data (Elith et al., 2011; Phillips et al., 2006; Tognelli et al., 2009), and linear models are simple but commonly used statistical methods useful for providing estimates of the relationship between species abundance records and environmental covariates (see Austin, 2002; Guisan et al., 1999 for considered reviews and pitfalls of the approach). We are aware that linear modelling of proportional data is problematic as it erroneously treats the dependent variable as continuous, and multinomial logistic models are preferable and often more robust (Zhao et al., 2001). However, as we do not wish to use the results in a formal predictive way but instead are only testing a hypothesis of covariance, our approach is reasonable.
It is important to identify two important assumptions underlying the reconstruction of species distribution patterns using archeological data: (1) that the record of animal bones (or more specifically for this paper, cattle bones) assigned to an archaeological site (or phase of a site) provides unbiased information on the presence or abundance of cattle in the local catchment of the site’s location; (2) that sites represent a random selection of locations sampled from across the full range of possible environments that the species may be found in the study region. Both of these assumptions are unlikely to be wholly true and our approach consequently requires careful consideration of sample biases. First, zooarchaeological data obtained from archaeological site records are not simple reflections of ‘natural’ species abundance. Archaeological subsistence records are formed by human agents who select, process and discard animal resources using complex rules that often, but not always, involve non-optimal decisions based on imperfect information, meaning that faunal records may not necessarily be accurate representations of even relative local species abundance. In additional, species may be hunted or herded at some distance from a site, distorting the relationship between the environment in which an animal lived versus where it is recovered archaeologically. Further, taphonomic effects (preservation) and sampling and recording error introduce additional noise into records that also reduce our ability to use zooarchaeological records to estimate the ecological determinants of species presence/absence or abundance. Second, archaeological site locations are non-randomly distributed in space with respect to environmental covariates, reflecting human choices concerning suitable locations in which to live. The type and local expression of covariates define the types of environments suitable for human settlement and subsistence; choices which are multifaceted and balance a range of competing needs (e.g. among others factors, access to suitable building materials and other raw materials, access to water, access to different types of subsistence, as well as the distribution of existing other settlements for ensuring suitable social distances). Although the distribution of suitable site locations will overlap to some extent with the distribution of species exploited by a community, without independent measure of species presence or abundance, it is difficult to determine this spatial relationship.
Despite these sources of bias, we are able to adjust our expectations of the models to accommodate these discrepancies between the ideal and the reality of using archaeological data in this way. First, the MaxEnt model only uses the presence of species (as described below) to estimate environmental covariates, which removes the effect of noise around species abundance scores. Second, we alter our definition of the response variable from reflecting the niches of wild and domestic cattle to the niches in which humans are exploiting wild and domestic cattle, rather than the cattle themselves. Third, for the linear modelling approach, we use a proxy for species abundance data, %MNISP, to explore how cattle use differs quantitatively in different environments. However, we restrict our hypotheses to a level supported by the quality of the data: i.e. we only test whether specific environmental covariates predict the variance in cattle abundance. Ecologists using modern species records go well beyond this and estimate more precisely how specific covariates determine species abundance. Archaeological data is too coarse to attempt this and the results would likely be spurious, so we do not extend the analysis to this level of explanation.
Model 1: Linear modelling of cattle abundance records
Multiple linear regression analysis was used to test whether there is a significant relationship between the covariates (or subsets of the covariates) in Table 4 and the abundance of Bos sp. at archaeological sites (or discrete phases of multiperiod sites). Abundance is expressed as the proportion of Bos to the total mammal NISP for that phase or site. Neither the dependent variable nor independent covariates are normally distributed, so they required transformation prior to analysis. Frequency values for cattle were arcsine transformed, which is a common method for positively skewed frequency data (Legendre and Legendre, 1998; see Crawley, 2005: 247–248). A cube root transformation was applied to all environmental variables, which normalized them sufficiently for the purposes of the analysis. Significant (R2>0.7) correlation was noted between three pairs of variables: prcov and prdry, prwet and prtot, and tprng and tpcov. To reduce confounding errors, prdry, prwet and tprng were removed from the analysis.
The data used for this analysis are presented in Table 3. Our first hypothesis predicts that the interaction of a set of independent environmental variables is able to explain some of the observed variation in cattle %MNISP values. To test this we first ran an exhaustive step-wise model selection routine (‘regsubsets’ in R). This uses the Mallows’ Cp statistic (Mallows, 1973) to assess the strength of models on the basis of their explanatory power, taking into account the problem of overfitting (the tendency for models to increase their fit as new terms are added, but with progressively less explanatory power attached to each additional term – i.e. a model with fewer strong variables is better than a model with many weaker predictive variables, even though the overall R2 value may be larger in the latter than the former). Model selection was thus oriented to identifying the smallest subset of variables that could explain a significant proportion of the variance in the dependent variable.
Model 2: MaxEnt of wild and domestic cattle presence records
To test our hypothesis that proposes there is a change in the range of environments associated with wild versus domestic cattle use, we develop a Maximum Entropy (MaxEnt) species distribution model. MaxEnt is designed to work with presence-only records, which has advantages for archaeological data where absence data are often missing and/or there are likely significant sampling biases associated with absences. The modelling procedure (more completely explained in Elith et al., 2011) first calculates the local environmental covariate values (Table 4) at the sites of observed species presence records (from sites listed in Table 2), and establishes how these values differ from a random sample (or multiple random samples) of the covariates in the study region. This provides estimates of multivariate distribution of suitable habit conditions (Franklin, 2009: 196–200). The conditional probability of species occurrence is calculated based on the ratio between the density vector of covariates at locations where species are present, against an estimate of covariate density across the study area. This ratio is treated as a logit score so that it can be used to predict the odds probability of species presence. The contribution of each covariate to predicting species presence is examined (individually and as a part of a set of potentially correlated variables) by varying its value but holding other covariates constant, which provides an estimate of its significance to the model. In addition, the species records can be partitioned into training and test samples so that the significance of the predictions can be estimated. In our implementations, we randomly partitioned the data into equal-sized training and test samples and used the MaxEnt toolkit’s built-in jackknife routine to establish each variable’s contribution to the model. The output is a spatial model that depicts the conditional probability of occurrence of a species across the study region alongside information on the significance of each of the covariates to explaining species presence in geographical and environmental space. As there is a stochastic component to deriving estimates of the background covariate density (by default a set of 10,000 random locations is sampled), we ran multiple models on multiple partitioned sets for cross-validation of the estimates of the species probability and the significance of the covariates.
Results
Linear models
As described, two models were constructed: one for Bos abundance for sites assigned to between 11.8 and 9.0 ka (n=34), and one for Bos abundance from sites with phase dating to between approximately 9.0 and 7.5 ka (n=38). In the early group there is very poor goodness of fit between environmental data and Bos abundance scores. The interaction of two variables, total annual precipitation seasonality (prtot) and maximum annual temperature (tpmax) can at best only account for about 20% of the variance of Bos %MNISP across the 34 locations (raw R2=0.21, adjusted R2=0.16). These two variables are collectively interpreted as expressing relative aridity, such that low annual precipitation and high maximum temperatures are more arid then regions with lower temperatures and/or more precipitation (cf. Pahari and Murai, 1999). Analysis of the Q-Q plots (not shown) to assess the goodness-of-fit of the distribution show that two sites are significant outliers: Ais Yorkis (Cyprus: Croft, 1998; Simmons, 2003) and Ortos (Cyprus: Croft, 2003). These sites report considerably fewer (1% or less) cattle than their environmental settling predicts, but as cattle are not expected in great numbers in early Neolithic Cyprus (Vigne, 2008) this is not surprising. After the removal of these two outliers the goodness-of-fit improves and annual precipitation explains about 25% of the variance in cattle %MNISP for the remaining 32 locations (Figure 5; Table 5a). However, although the correlation is statistically significant, the considerable noise around the signal doesn’t provide much additional insight. For the early phases, we conclude that abundance data can only partially be accounted for by regional variability in temperature and precipitation and that other factors, such as the presence of competing high-rank prey species as well as cultural traditions that influence prey choice, may be better predictors of cattle use. These others factors will, of course, require testing.

Relationship between environmental variables and cattle abundance (Bos %MNISP). The y-axis scales are arcsin transformed. (a) Early Phases Bos by Precipitation – R2=0.26, p=0.003; (b) Late Phases Bos by Precipitation – R2=0.37, p<0.001; (c) Late Phases Bos by Maximum Temperature – R2=0.32, p<0.001.
Early phases linear model results.
Standardized Coefficients
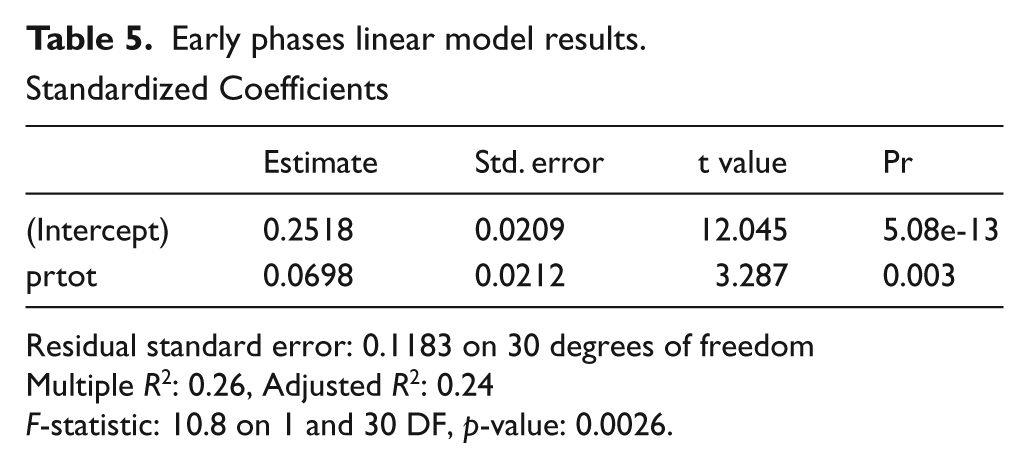
Residual standard error: 0.1183 on 30 degrees of freedom
Multiple R2: 0.26, Adjusted R2: 0.24
F-statistic: 10.8 on 1 and 30 DF, p-value: 0.0026.
For the 38 later Neolithic locations, the strength of the association between the dependent and independent variables significantly increases. Slightly under 40% of the variance of cattle %MNISP is explained by regional differences in aridity as expressed by the interaction of two variables: annual precipitation and maximum temperature. This pattern was confirmed by permutation. Examination of goodness-of-fit plots shows that there are three significant outliers: Ovčarova Gorata (Bulgaria: Nobis, 1986), Halula (Syria: Saña-Segui, 1999, 2000), and Tenta (Cyprus: Croft, 1982, 2005). The first two have significantly more cattle than predicted given their location: Ovčarova Gorata has over 66% cattle, which is high and likely misrepresents the contribution of smaller taxa. At Halula a more realistic 20% cattle by %MNISP is reported, which is high only within the general regional context, but can be accounted for by its position on the Euphrates where there is both ready access to water and a strong local tradition of cattle use extending back into the earlier Neolithic. At Tenta cattle are not recorded, despite the availability of cattle on Cyprus in this period, but the island context may lead to local reductions in availability. Removal of these outliers from the model improves the fit, and regional precipitation and temperature variability explains almost 50% of the variance in cattle NISP across the study region (Table 6, Figure 5b, c).
Late phases linear model results.
Standardized Coefficients
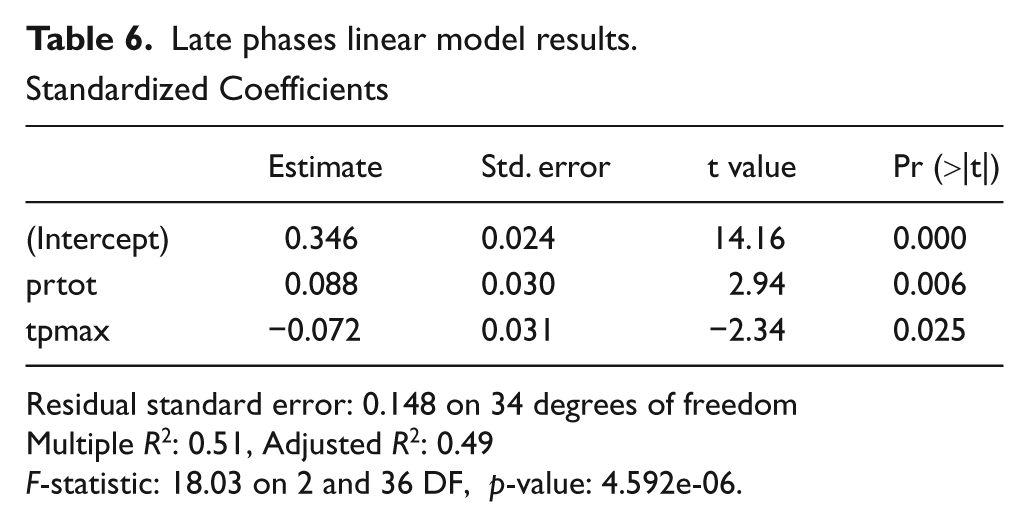
Residual standard error: 0.148 on 34 degrees of freedom
Multiple R2: 0.51, Adjusted R2: 0.49
F-statistic: 18.03 on 2 and 36 DF, p-value: 4.592e-06.
We should point out that these results are not spatially stationary processes – the goodness-of-fit of the model almost certainly varies across space, and thus we can assume that environmental factors explain the variation in cattle use better in some regions than others. Our preliminary tests of this using geographically weighted regression do in fact show a tendency for the fit to improve from east to west, although the details and interpretation of this pattern is beyond the scope of this paper.
Our first hypothesis is thus demonstrably true: regional environmental variation does explain some of the variation in cattle %MNISP – from about 25% to 50% in the earlier and later Neolithic phases, respectively. Importantly, the one variable identified as having the strongest relationship – total precipitation – is the same between the two periods, although the strength of this relationship between precipitation and cattle use becomes stronger after 9.0 ka. An equally significant observation is that temperature becomes a significant predictor in the later period. We interpret this as arising from a process in which, initially, communities exploiting cattle were located in warmer and semi-arid regions of the study region (i.e. the Levantine and upper Euphrates regions), although within these regions of initial use precipitation is only a weak determinant of use patterns. After about 9.0 ka, cattle had become more prevalent in subsistence economies and the range of use expanded into temperate regions of central and western Anatolia and Greece and Bulgaria. These are areas with higher amounts of annual precipitation and lower temperatures, which would have reduced the risks and enabled greater investment in cattle, further strengthening the observed statistical relationship between cooler climates and increased cattle use after 9.0 ka.
We feel it is worth emphasizing that despite the majority of the variation in NISP values not being accounted for by these models, the fact that we are able to explain nearly 50% of the variation in observed cattle abundance at later Neolithic sites using modern environmental covariates is remarkable, especially given that abundance values are subject to a range of sample errors as articulated earlier.
MaxEnt
Our second hypothesis proposed that over time there should be changes in the types of environments in which wild and domestic cattle are exploited, driven by the expansion of domestic cattle use in the later part of the early Neolithic. We tested this using a MaxEnt based species distribution model for presence-only records of wild and domestic cattle, for sites with wild and/or domestic cattle listed in Table 2. As we have already explained, the distinction between wild and domestic in early-Holocene cattle is not straightforward and taking original analysts’ distinctions at face-value is problematic, especially for assemblages published several decades ago. In our data set, the difference between ‘wild’ and ‘domestic’ may be mostly expressing size differences, which may reflect differences in domestic status – but not necessarily so. We thus accept that these models have error terms that are difficult to estimate; but we remain confident that the overall trends identified are likely to hold true.
The map of predicted probabilities for wild cattle based on the mean output of the cross-validated MaxEnt models is shown in the upper part of Figure 6. Of the 11 covariates, distance to river, precipitation seasonality, and maximum annual temperature are the top three predictors of wild cattle presence. Regions where wild cattle are predicted as being more likely on sites on the basis of environmental suitability are the coastal areas of the southern Levant, the arc running north then east along the Taurus and Zagros Mountains, coastal parts of Cyprus (on environmental suitability – not necessarily availability), and western Anatolia and, at slightly lower probabilities, the Aegean side of the Greece. Examination of the (modern) environmental covariates indicates that these regions are characterized by low to moderate seasonal rainfall, with an interquartile range of total annual precipitation of 300 to 500 mm, 60 to 100 mm of which falls in the wettest month (December or January), and low seasonal temperature fluctuations with an annual maximum median of 36°C. These can be generalized as typically semi-arid Mediterranean environments, with the more arid areas lying farther inland, and south of the more seasonally variable mountainous areas of central and eastern Turkey and the Zagros. These regions correspond to the dense forest and woodlands of the littoral regions and the arc of park-woodland and steppe-woodland vegetation communities running from the southern Levant along the Zagros, as reconstructed for post 9000 BP environments (Moore et al., 2000: 78–80).

MaxEnt derived predicted probability distribution for ‘wild’ (a) and ‘domestic’ (b) cattle. Note caution expressed in Figure 3.
The modelled probability distribution of sites using domestic cattle is shown in the lower part of Figure 6. The top four predictive variables are average temperature, total annual precipitation, maximum temperature, and precipitation in the wettest month. The region where sites using domestic cattle are predicted to be located overlaps areas with wild cattle exploitation, but extends into central and eastern Anatolia, and southeastern Europe east of the Dinaric Alps. Within these areas, the coastal area of the southern Levant, the Mediterranean coastal regions of southeastern Turkey, parts of Cyprus, and Aegean coastal regions, including Crete, are subregions with relatively high probabilities for sites that exploit domestic cattle. These regions are characterized by higher annual rainfall with an interquartile range of 450 to 600 mm/yr, but up to 800 mm/yr in parts of western Anatolia and northern Greece, and lower median maximum temperatures of 32°C. Comparison of the two maps shows that the spatial distribution of the higher (p Bos>0.6) likelihood regions shifts westward, and slightly expands in total area: The proportion of the study area at p Bos>0.7 is 3.7% for wild cattle and 5.9% for domestic cattle. This supports our hypothesis that the process of domestication involved an expansion of the types of environments in which cattle were exploited, with the expansion focused in cooler and better watered areas of NW Anatolia and northern Greece and Bulgaria.
Discussion
We predicted at the onset that a significant proportion of the identified regional variation in cattle NISP could in part be explained by independent environmental variables. This was confirmed and, although not an altogether surprising conclusion, it is an important observation that can be built upon, especially for developing explanations for the regional variations in the uptake of domesticates. However, it is the outliers to this model of Bos abundance that are equally interesting: For example, the positive outliers in the early phases (PPNA–LPPNB) include Göbekli Tepe (Turkey: Peters et al., 2005; von den Driesch and Peters, 1999, 2001), Halula (Syria: Saña-Segui, 1999), Mezraa-Teleilat (Turkey) at 19.9% NISP (Ilgezdi, 2008), Abu Gosh (Israel: Davis, 1978; Ducos, 1978b; Ducos and Horwitz, 2003; Horwitz, 2003), and Munhatta (Israel: Ducos, 1968) (Figure 1). These sites were predicted to have no more than 10% cattle as a proportion of mammal taxa, but instead have up to 17%. The importance of cattle in the Euphrates region has been commented on by others (e.g. Peters et al., 1999), but our data show that cattle are similarly important at some sites in the Mediterranean Levant. At the other end of the scale, in addition to the two Cypriot sites considered earlier there are three further significant negative outliers where cattle are predicted to be up to 10% of the assemblage but do not exceed 4%. These are Ali Kosh (Iran: Hesse, 1978), Körtik Tepe (Turkey: Arbuckle and Özkaya, 2006), and Motza (Israel: Sapir, 2005). Ali Kosh is at the extreme of our distribution in the eastern Zagros, and although it may have been a centre for early goat domestication (Zeder, 2005), cattle evidently did not play an important role in the economy. The latter two sites are relatively early, where there is more pronounced variability in exploitation patterns; fewer cattle than expected seems likely to be a stochastic effect.
In the later phases of the Neolithic there is a clear increase in the frequency of cattle use, although the highest proportions of cattle are observed in sites in Greece and Bulgaria. The environmental variables used to predict cattle use also highlight the greater potential for cattle in SE Europe compared with SW Asia, providing support for our second hypothesis that the later phases of the Neolithic include an expansion into temperate environments that supported increased use of cattle. The residuals for the later phases are informative: In addition to Ovčarovo Gorata and Halula referred to previously, high positive deviations are also found at Hagoshrim (Israel: Haber, 2001; Haber and Dayan, 2004), Fikirtepe (Turkey: Boessneck and von den Driesch, 1979), Ovčarovo Gorata (Bulgaria: Nobis, 1986), Koprivec (Bulgaria: Manhart, 1997), and Sofia-Slatina (Bulgaria: Todorova and Vajsov, 1993) (Figure 1). The NW Anatolian and Bulgarian sites represent later phases of the early Neolithic, where use of managed species is more important, and it is entirely possible that they had developed more specialized exploitation patterns with an emphasis on domestic cattle. This is in keeping with the general character of the faunal exploitation patterns of the SE European early Neolithic use a less diverse range of taxa than in PPNB SW Asia. It is less clear why Hagoshrim has relatively high representation of cattle (33% MNISP versus 13% predicted for its location). The faunal sample was collected with careful control (Haber and Dayan, 2004: 1589) so it is not biased towards larger species, but it does represent a locally high concentration of cattle exploitation that requires further consideration.
The significant negative outliers in the later phases include Kazanluk in Bulgaria (Dennell, 1978), where domestic cattle contribute less than 10% to the assemblage, whereas it is predicted to comprise over 20%; and Erbaba (Turkey: Arbuckle, 2008), where cattle accounts for 5.5% of the assemblage, but 20% is predicted based on location. It is worthwhile noting that these outliers, although individually interesting and potentially significant for locally contingent reasons, are nevertheless not so extreme as to undermine the modelling exercise. They are minor exceptions to a general pattern that predicts a significant increase in the use of cattle in western Anatolia and SE Europe on the basis of the important differences in environment compared to the more arid areas of SW Asia. Moreover, this exercise is important as it identifies specific assemblages that deviate from the expected patterns, which provides encouragement for continued assessment of their characteristics.
The MaxEnt model provides additional support for the modelled difference between the western and eastern parts of our study area, with the probability of domestic cattle being much higher for south-central Anatolia and SE Europe. Given the significant (25%+) contribution that cattle make to several faunal assemblages in the western part of the study area, it is also reasonable to suggest that the growth of Neolithic communities in more temperate environments provided opportunities for the expansion of cattle use, which are less drought-tolerant than ovicaprines. Thus, after approximately 8500 cal. BP when Neolithic economies have begun their expansion into central Europe, the foundation had already been set for cattle-rearing to emerge as a defining characteristic of early European farming (Bogaard, 2005; Manning et al., 2012).
Footnotes
Funding
The research was funded by grant AH/D503434/1 from the UK Arts and Humanities Research Council.