
Abstract
Central-European raised bogs, developed on mountain summits, are specific ecosystems of high conservation importance, but their history remains largely unknown. Pollen, macrofossils, testate amoebae and peat characteristics were analysed in a peat sequence of the Vozka bog (Hrubý Jeseník Mountains, Eastern Sudetes, Czech Republic). Past water chemistry and water-table depths were reconstructed by transfer functions calibrated from recent testate amoeba data and long-term environmental averages. Peat initiation started in the middle Holocene (approximately 4200
Introduction
Ombrotrophic bog habitats accumulate a large amount of organic material consisting of the micro- and macroremains of organisms (Mauquoy and van Geel, 2007; Tolonen and Turunen, 1996). This provides an opportunity to study ecosystem development directly and thus to re-evaluate succession theories previously based on time–space substitution studies. Autogenic succession from lakes or groundwater-fed fens to rainwater-fed bogs is the most common successional pathway documented by stratigraphical analyses (Kuhry et al., 1993; Magyari et al., 2001; Michaelis, 2002; Walker, 1970). Paludification is another important process leading to the development of rainwater-fed bogs in the absence of groundwater discharge (Charman, 2002). Climate favourable for paludification particularly occurs in boreal-oceanic regions and high mountain regions (Steiner, 1992), both of which are characterised by high precipitation and low evapotranspiration. Boreal and high-mountain bogs are rather sensitive to allogenic impacts, such as hydrological changes, grazing or air pollution (Bragazza et al., 2004; Jiroušek et al., 2011; Pellerin et al., 2008; Sjögren et al., 2007), which may lead to the changes in both the species composition and the peat properties, including the level of decomposition and compression (Franzén, 2006; Sjögren et al., 2007). These changes may alter or even reverse the normal successional development (Hájková et al., 2011a; Hughes and Dumayne-Peaty, 2002).
Central-European raised bogs that developed on impermeable bedrock at mountain summits are of high conservation importance. They are usually situated at a flat terrain of a mountain ridge and are thus saturated either exclusively by rainwater or by both rainwater and surface water coming from the adjacent slopes. Historically, they have been affected by husbandry from the early medieval period or even earlier (Gobet et al., 2005; Novák et al., 2010). More recently, many have disappeared because of drainage and afforestation, and the remaining ones are subjected to increasing nutrient, mineral and toxic-compound input through aerial liming and atmospheric deposition (Hájková et al., 2011a; Jiroušek et al., 2011). In order to evaluate the recent ecosystem changes in the longer temporal context, detailed knowledge about mires and landscape development during the Holocene is needed. Unfortunately, these bogs have been poorly studied in terms of the local vegetation development inferred from macrofossil and/or testacean analyses (but see Speranza et al., 2000). The lack of palaeoecological information particularly holds for the Hrubý Jeseník Mountains, the second highest mountain range in the Czech Republic, located in the easternmost part of the European Hercynian mountain system. In that region, the most hitherto studied peat sequences are young, they started to form in the late Holocene (Opravil, 1959; Rybníček and Rybníčková, 2004; Treml et al., 2006, 2008). Although all of these studies have concentrated on landscape development, they did not generate unequivocal reconstructions of regional vegetation and, moreover, have been challenged by the results of soil-charcoal analyses (Novák et al., 2010).
Modern palaeoecological studies often use the multiproxy approach, which allows the detailed reconstruction of mire succession, landscape cover and human impact (De Vleeschouwer et al., 2009; Lamentowicz et al., 2010). The principal objective of this study is to apply the multiproxy approach, including peat stratigraphy, pollen, macrofossil, testate amoebae, microcharcoal and soil analysis, to the peat sequence of the representative summit bog in the Hrubý Jeseník Mountains, considered to be the oldest bog in the study region (Opravil, 1959). The main aims are: (1) to study its local vegetation history, including reconstruction based on transfer-function models and (2) to increase the information about Holocene landscape development at the high altitudes of the Eastern Sudetes and its possible effect on bog development.
Study site and methods
Site
Jeseníky Mts (Gesenke in German) are the easternmost part of the Bohemian Massif (Figure 1). The highest area, called Hrubý Jeseník Mountains (Hohes Gesenke, Altvatergebirge), reaches the alpine belt and consists of gneiss and schist bedrock. According to the occurrence of light-demanding stenoendemits and patterned soils, its highest peaks are thought to have been treeless during the entire Holocene.
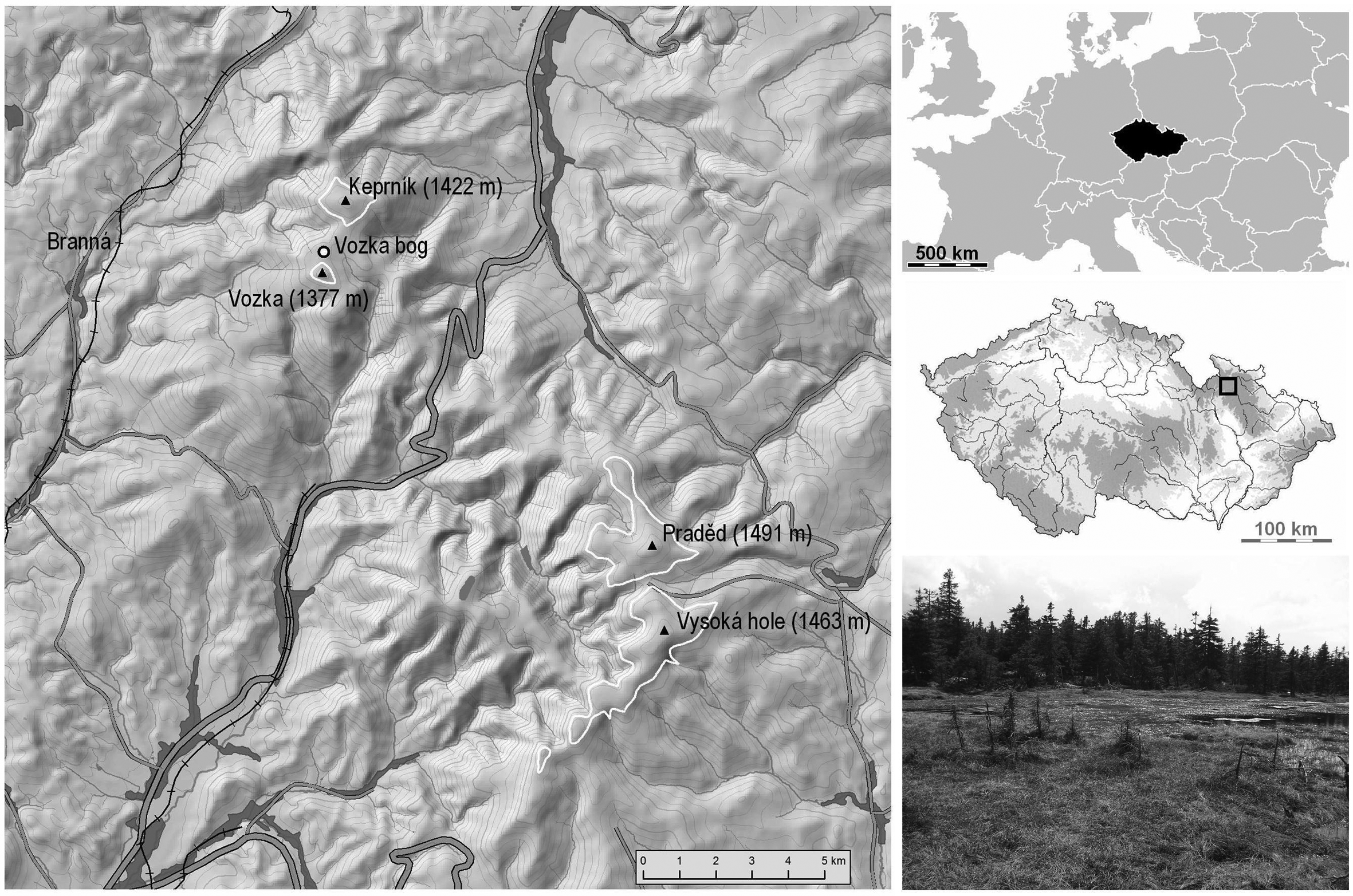
Site map of the Vozka bog, situated in the summit area of the Hrubý Jeseník Mountains, Czech Republic. Approximate position of the natural treeline is depicted by a white line. Open-face excavation was carried out at the place shown on the photo.
In this region, the existence of high-mountain ombrotrophic bogs is conditioned by the coinciding high annual mean precipitation (1900 mm), low mean temperature (2.2°C), and favourable topography. According to geological maps (available at http://www.geology.cz/ accessed 4 August 2011), 42 small peat deposits occur above 1100 m a.s.l., situated either on the summit plateau, in saddles or on gentle slopes. The studied Vozka bog (1320–1350 m a.s.l.) is situated in the northwestern part of the Hrubý Jeseník Mountains, close to Keprník Mountain, a summit with an isolated island of alpine vegetation (Figure 1). It lies in the National Nature Reserve Šerák-Keprník, slightly below the recent treeline. According to hydromorphological classification (Charman, 2002), it could be classified as a raised bog. It is surrounded and locally covered by spruce woodland. The bog surface is formed of hollows (Drepanoclado fluitantis-Caricetum limosae), flat lawns and small hummocks with the vegetation classified as subcontinental montane bog vegetation (Andromedo polifoliae-Sphagnetum magellanici, for details see Chytrý, 2011).
Sediment sampling and analyses
The peat section was taken from the deepest point (50°9′31.22″N, 17°6′24.7″E, 1330 m a.s.l.), located close to the largest bog pool. The section of 165 cm deep organic sediment, lying directly on partially weathered gneiss was taken from an open face excavation and stored in metal boxes (10 cm × 10 cm × 50 cm). Colour was determined according to the Munsel soil colour charts (Munsell Color Company, 1954).
The section was sampled and analysed as follows: samples from each 5 cm section were analysed for dry bulk density, ash content and pollen spectra; samples from each 10 cm section (except for the uppermost sample) were analysed for testate amoebae; and monoliths with length of 5 cm were cut subsequently to obtain a sample of 100 cm3 volume for macrofossil analysis.
Samples of 6 cm3 in fresh state were cut in order to obtain dry bulk density and ash content. Peat samples were dried for 7 days at room temperature and weighed. The same samples were then dried at 105°C for 1 h to remove the air moisture, weighed, burnt for 3 h at 550°C in a kiln and then weighed again.
Samples for pollen analysis (1–2 cm3) were treated with acetolysis, and the two deepest samples were additionally treated with HF, according to Faegri and Iversen (1989). Tablets with Lycopodium spores were added in order to calculate the pollen concentration and pollen influx (Stockmarr, 1971). Pollen were identified using the reference literature (Beug, 2004; Punt, 1976; Punt and Blackmore, 1991; Punt and Clarke, 1980, 1981, 1984; Punt et al., 1988, 1995, 2003; Moore et al., 1991; Reille, 1992) and the reference collection at the Institute of Botany ASCR. The determination of spores and non-pollen palynomorphs follow Moore et al. (1991) and van Geel et al. (1980–1981, 2003).
Peat samples for testate amoebae determination were macerated for 24 h in the distilled water and 3 ml of solution were analysed after sedimentation. All tests in this volume were determined and counted (e.g. Laminger et al., 1981). In order to calibrate fossil testate amoeba assemblages, modern assemblages and measured environmental data were used. A net of 30 permanent plots established in eight natural mires in the same study region were utilised. The water level below the surface was measured and bog water samples were taken periodically over 15 years, with 21 chemical variables analysed (for details see Hájková et al., 2011b; Rybníček, 1997). From each permanent plot, water was squeezed from bryophyte tufts and concentrated by sedimentation for 24 h. As the recent testate amoebae were better preserved than the fossil ones, the total number of individuals was counted in 0.1 ml of sample. Species were determined with help of Bartoš (1954), Charman et al. (2000) and Ogden and Hedley (1980).
For macrofossil analysis, monoliths of peat were rinsed with running water on 1 mm, 630 μm and 200 μm mesh sieves. Seeds were determined according to Beijerinck (1974), and wood fragments according to Schweingruber (1978) and Gregus (1959). Leaf and tissue determination follows Mauquoy and van Geel (2007). Material was also compared with the macrofossil reference collection at the Institute of Botany ASCR.
Additionally, chemical analyses of peat were carried out at various depths of the profile. The peat was leached by Goehler’s agent. Nitrogen was detected by distillation with water vapour and sodium hydroxide addition, with released ammonia absorbed to boric acid and then titrated by sulphuric acid. Phosphorus was detected spectrophotometrically after colouration by a molybdenum agent. Potassium was detected by flame atomic emission spectrometry. Calcium was determined by atomic absorption spectrophotometry.
The age of four samples was initially determined using conventional radiocarbon dating of bulk samples. Later, macrofossils from three samples were analysed by Acceleration Mass Spectrometry (for details see Table 1).
Radiocarbon dates of the Vozka peat samples.

UGAMS: Centre of Applied Isotope Studies, The University of Georgia, USA; GrN: Centre for Isotope Research, Groningen, the Netherlands; Conv.: conventional dating; E. vaginatum: Eriophorum vaginatum; BP: before present;
Data processing
Radiocarbon ages were calibrated by OxCal (Bronk Ramsey, 2009) using the calibration curve Bomb04NH1 (Hua and Barbetti, 2004) for sample UGAMS8799 and IntCal09 (Reimer et al., 2009) for other samples. Inversion between GrN-31530 and UGAMS-10457 may be caused by different dating methods, however the overlap between the 95.4% ranges of the two dates suggests that both dates may be correct. An age–depth model was created with OxCal (Poisson deposition, k=0.8, Figure 2). In the main text the age of each sample refers to a centre of interpolated calibrated age with 68.2% probability.

Sediment characteristics of the Vozka bog section (from left to right): peat stratigraphy and colour (names follow Munsell Color Company, 1954); components of peat; peat characteristics: dry bulk density (dark grey line) and ash content (i.e. loss-on-ignition, light grey line); zonation of the sediment; age–depth relationship based on seven radiocarbon dates (sampling year is highlighted by arrow for better visibility). Abbreviations: dark r. b.: dark reddish brown.
The results of pollen, macrofossil and testate amoeba analyses are presented in diagrams created by PolPal (Nalepka and Walanus, 2003). Pollen and charcoal concentrations and influx were calculated according to Faegri and Iversen (1989) and Hicks and Hyvärinen (1999). For a better delimitation of assemblage zones in pollen diagram, agglomerative cluster analysis (ConsLink) implemented in PolPal was used.
Principal component analyses (PCA) of pollen, macrofossil and testate amoeba data were carried out using Canoco 4.5 software (ter Braak and Šmilauer, 2002) in order to compare the main gradients in species data variation. The PCA analysis was used because of the short gradient in species data (below 3 s.d. units for all proxies). For pollen and testate amoebae, log-transformed percentage abundance values were used. For macrofossils, where different scales were used for seeds, woods and bryophytes, the presence-absence data was used with the pseudospecies generated in the case of Sphagnum magellanicum, S. russowii and/or Eriophorum vaginatum dominance (i.e. > 50% of sample volume).
To carry out the palaeoenvironmental reconstruction, a calibration data set of 30 modern samples of testate amoebae was applied. Species data in percentages per sample and 15 yr environmental averages were used. Only five environmental variables (water level, water conductivity, pH, and calcium and magnesium concentrations) significantly explained the variation in recent testate amoeba data in the canonical correspondence analysis (Monte Carlo test, P <0.01; results not shown). Fifteen-year averages were used because of the higher portion of explained variability in recent species data as compared with single measurements or mean values from shorter periods. As water conductivity, calcium and magnesium concentrations provided the same results, only the reconstruction of calcium concentration is presented. The root mean square error of prediction (RMSEP) was calculated by bootstrap cross-validation (Crowley, 1992) and the model with the lowest RMSEP (weighted averaging) was used. Analyses were carried out using C2 software.
Results
Sediment stratigraphy, age and soil analyses
According to sediment composition, dry bulk density and ash content there were three main zones distinguished in the section (Figure 2). Dry bulk density and ash content curves mostly gave similar trends. Both variables reached higher values in the lowermost layer (VOs-1; 165–130 cm), then slowly decreased (VOs-2; 130–110 cm) to minimal values that were stable towards the surface (VOs-3; 110–0 cm). The only exception to this trend was recorded around 50 cm, where an abrupt peak in both soil characteristics is evident.
There are two main components of peat in the section: Eriophorum vaginatum, which is dominant in zone VOs-1, is gradually altered in the transitional zone (VOs-2) by Sphagnum species, which became dominant in the zone VOs-3. The mineral admixture and leaves of non-sphagnous mosses are present in the zone VOs-1. Wood of dwarf shrubs occurs in a small amount (up to 5%) whereas vascular plant tissues reach values of 5–50% of the sample volume throughout the entire section.
Both the calibrated and uncalibrated ages of seven samples are presented in Table 1. Organic matter started to accumulate around 4180 yr
Pollen analysis
According to the pollen record, and with the help of the Conslink method, the profile is divided into five zones, reflecting the main changes in pollen spectra (Figure 3).
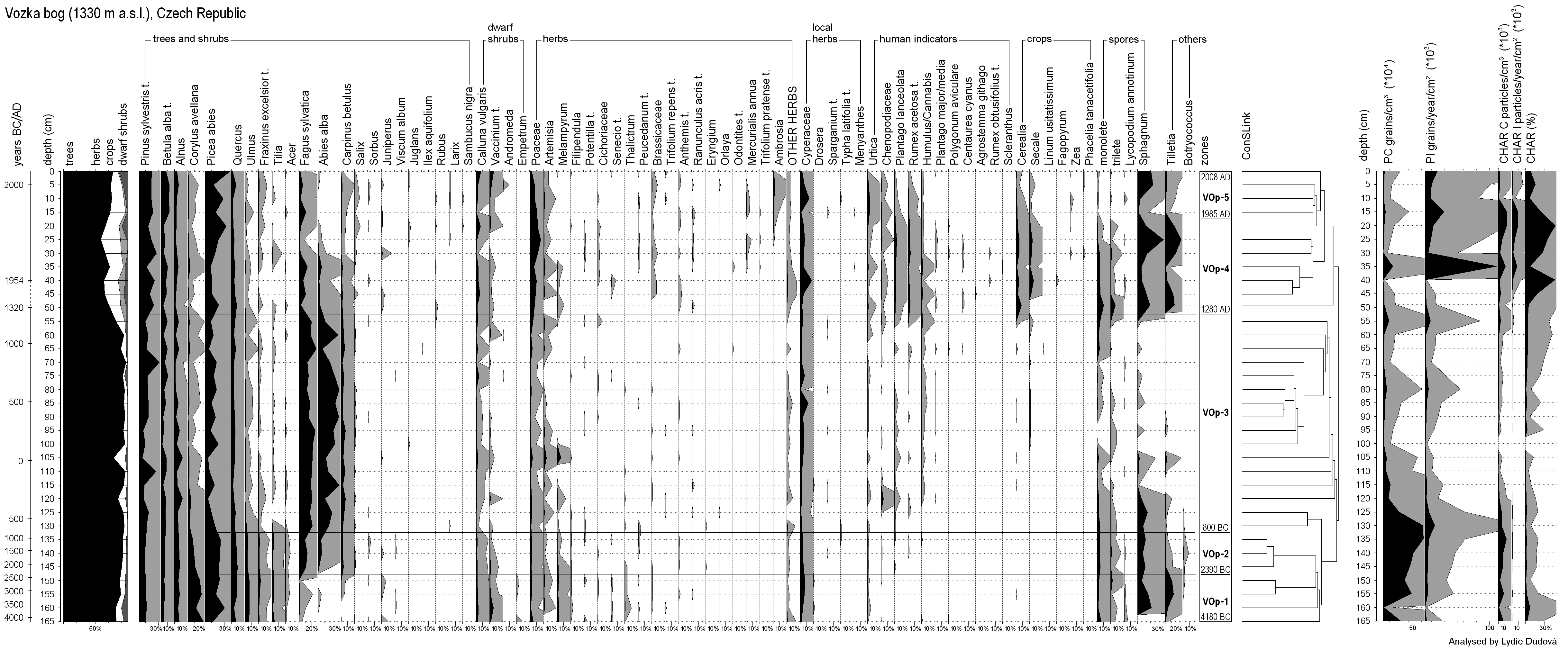
Pollen diagram of the Vozka bog based on total pollen sum, excluding local herbs (from left to right): approximate time scale in calibrated years, depth scale, cumulative tree/herb pollen diagram (without local herbs), percentage diagram of selected taxa, zonation with approximate age of boundaries, ConsLink dendrogram, pollen concentration (PC), pollen influx (PI), charcoal concentration (CHAR C), charcoal influx (CHAR I) and charcoal percentages (based on total pollen sum, CHAR %).
Zone VOp-1 (4180–2390
The main features of the VOp-2 zone (2390–800
Samples of zone VOp-3 (800
Zone VOp-4 (
In the uppermost zone, VOp-5 (
Macrofossil analysis
The profile was divided into three zones (Figure 4). The first zone (VOm-1; 165–110 cm) is characterised by a dominance of Eriophorum vaginatum tissues. Seeds of Potentilla erecta were present only in the deepest layers. Leaves of bryophytes Polytrichum sp., Warnstorfia fluitans, Sphagnum magellanicum and S. russowii were found in this zone. Twigs of dwarf shrubs (Calluna vulgaris, Vaccinium myrtillus and Empetrum nigrum) and the wood, peridermis and needles of Picea abies also occurred.
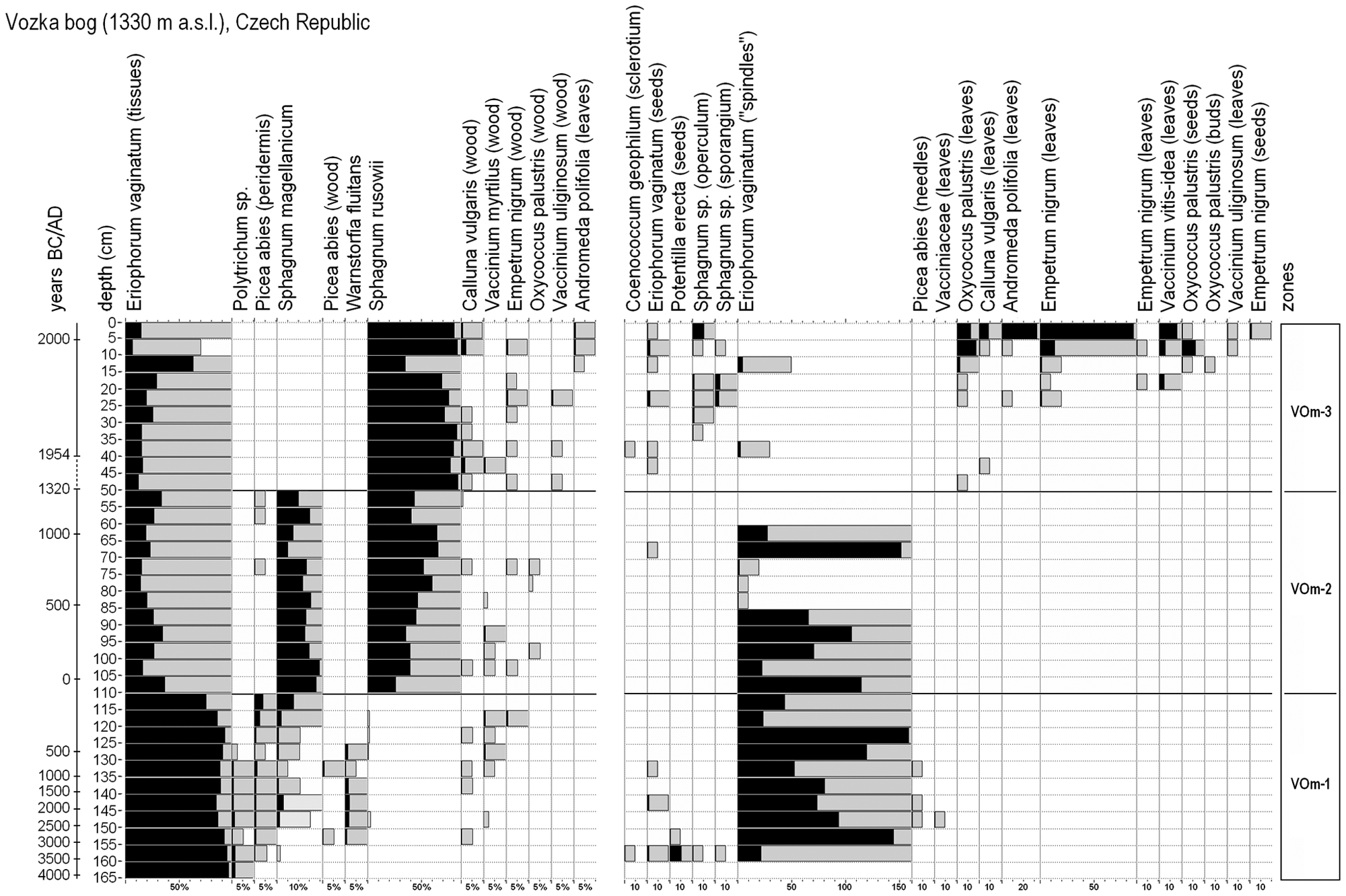
Macrofossil diagram and zonation of the Vozka bog. Macrofossils are given either in volume percentages (plant tissues, bryophytes, wood; on the left) or in absolute numbers (seeds, leaves, spindles; on the right).
In the following zone (VOm-2; 110–50 cm) stems and leaves of both Sphagnum species were more abundant than before, especially Sphagnum russowii. Eriophorum vaginatum strongly decreased its abundance contrary to the former zone. From this event, vegetation did not differ substantially from the recent layer consisting of Sphagnum magellanicum, S. russowii, Vaccinium species, Empetrum nigrum, Oxycoccus palustris, Andromeda polifolia and other bog species.
In the upper zone (VOm-3; 50–0 cm) Sphagnum rusowii became the dominant species in the bryophyte layer, whereas Sphagnum magellanicum disappeared. Representation of Eriophorum vaginatum tissues in peat was still low. The dwarf shrubs Calluna vulgaris, Oxycoccus palustris agg. and Empetrum nigrum increased their abundance and Andromeda polifolia and Vaccinium uliginosum also appeared.
Testate amoebae analysis
Cyclopyxis eurystoma is the only species present in the 155 cm sample, while in the deepest sample no evidence was found (Figure 5). The first developmental phase (VOt-1, 165–115 cm) is characterised by the presence of Cyclopyxis eurystoma, Archerella flavum, Arcella catinus, Assulina seminulum, Hyalosphenia papilio and Trigonopyxis arcula. Amphitrema wrightianum was found with high abundance in the sample 125 cm and then it entirely disappeared.
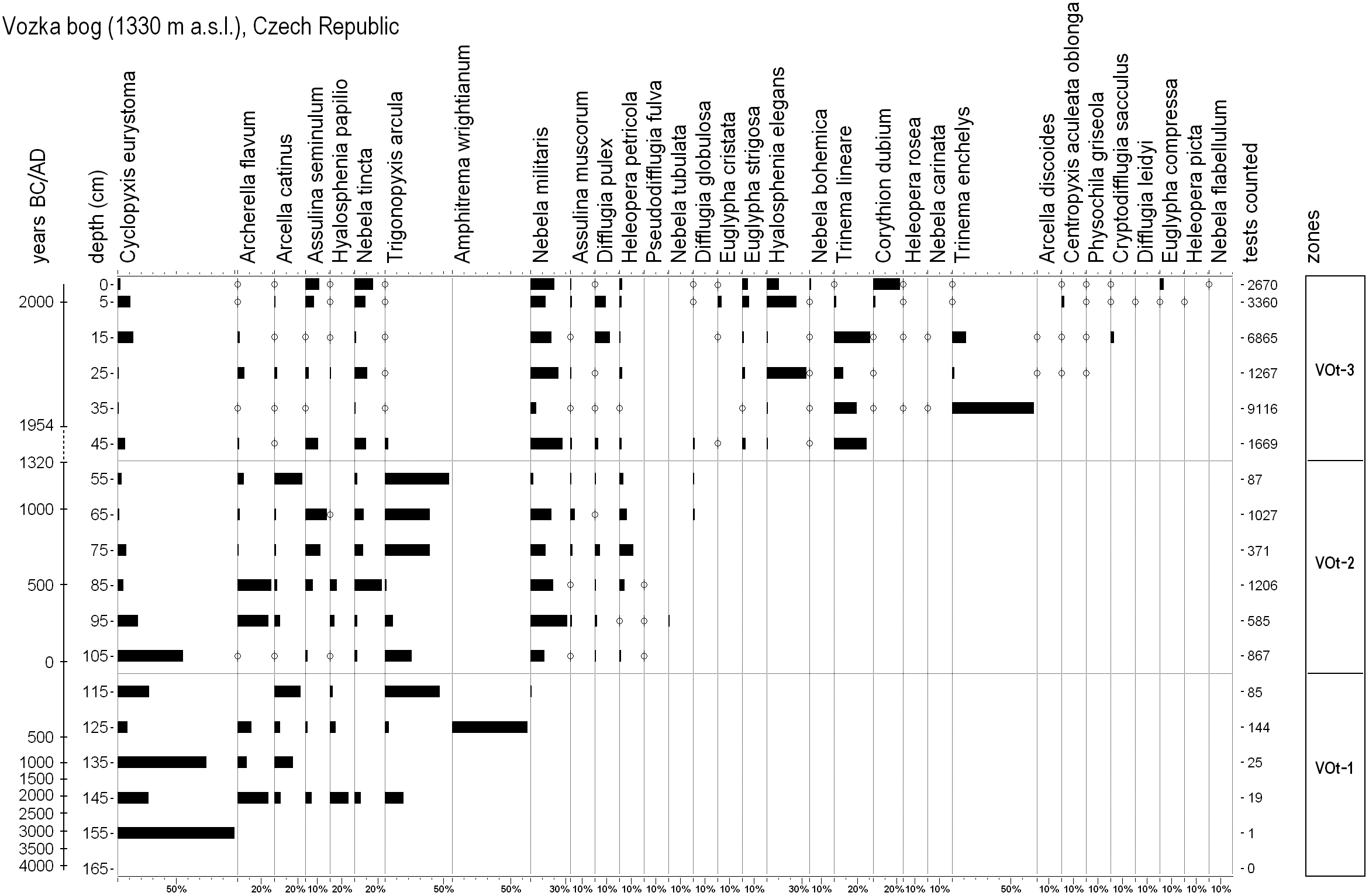
Percentage diagram of testate amoebae of the Vozka bog based on total sum of testate amoebae. Total sum of tests counted in each sample is provided at the end of the diagram. Histograms of values lower than 0.9% are highlighted by circles.
In the following zone (VOt-2, 105–55 cm) Assulina muscorum, Difflugia pulex, Heleopera petricola and Pseudodifflugia fulva appeared and Nebela militaris reached high abundance. Archerella flavum, Cyclopyxis eurystoma and Hyalosphenia papilio were more abundant in the lower part of this zone (105–85 cm), whereas Trigonopyxis arcula, Assulina seminulum and Nebela tincta in the upper (85–65 cm). At the end of this zone, Difflugia globulosa appeared and Pseudodifflugia fulva entirely disappeared from the profile.
Species richness of the upper part of the profile (zone VOt-3, 45–0 cm) was higher in comparison with the former zones, probably because of young peat which enabled good test preservation. Trinema lineare appeared in higher abundances between 45 and 15 cm and Trinema enchelys between 35 and 15 cm. Euglypha strigosa, Hyalosphenia elegans and Nebela bohemica occurred constantly through the entire zone. Some species, such as Centropyxis aculeata oblonga, Corythion dubium, Cryptodifflugia sacculus, Euglypha compressa, E. cristata and Physochila griseola, were characteristic of the youngest peat layers (0–15 cm).
Palaeoenvironmental reconstruction
Sample scores on the first PCA axis of macrofossils and testate amoebae show similar patterns, whereas that of regional pollen differs (Figure 6). Nevertheless, all three proxies reflect abrupt environmental changes at 50 cm. The result of the PCA analysis of local pollen (dwarf shrubs and local herbs, see Figure 3) is not shown because of its low representativeness as a result of low number of species and a certain stochasticity in pollen record (e.g. Empetrum pollen were not present in some depths despite the occurrence of its macrofossils).

From left to right: (1) sample scores of regional pollen, macrofossils and testate amoebae on the first axis of Principal Component Analysis (PCA); eigenvalues: 0.405 (regional pollen), 0.379 (macrofossils) and 0.326 (testate amoebae); cumulative percentage variance of species data: 40.5 (regional pollen), 37.9 (macrofossils) and 32.6 (testate amoebae); (2) reconstructions of water-table depth (WTD), water pH and water calcium concentration inferred from testate amoebae with sample-specific bootstrap errors of reconstruction; (3) directly measured concentrations of total nitrogen (total N), phosphorus (P), potassium (K) and calcium (Ca) in peat.
The relationships between the observed and predicted values of WTD, pH and calcium (Figure 7), the values of RMSEP and maximum bias results show the good quality of the transfer function models and their usefulness for the reconstructions. The species composition of testacean communities in the beginning of bog development indicates wetter site conditions contrary to the recent water regime. The water-table probably declined approximately 3–13 cm below the bog surface (Figure 6). The water level was highest around 470
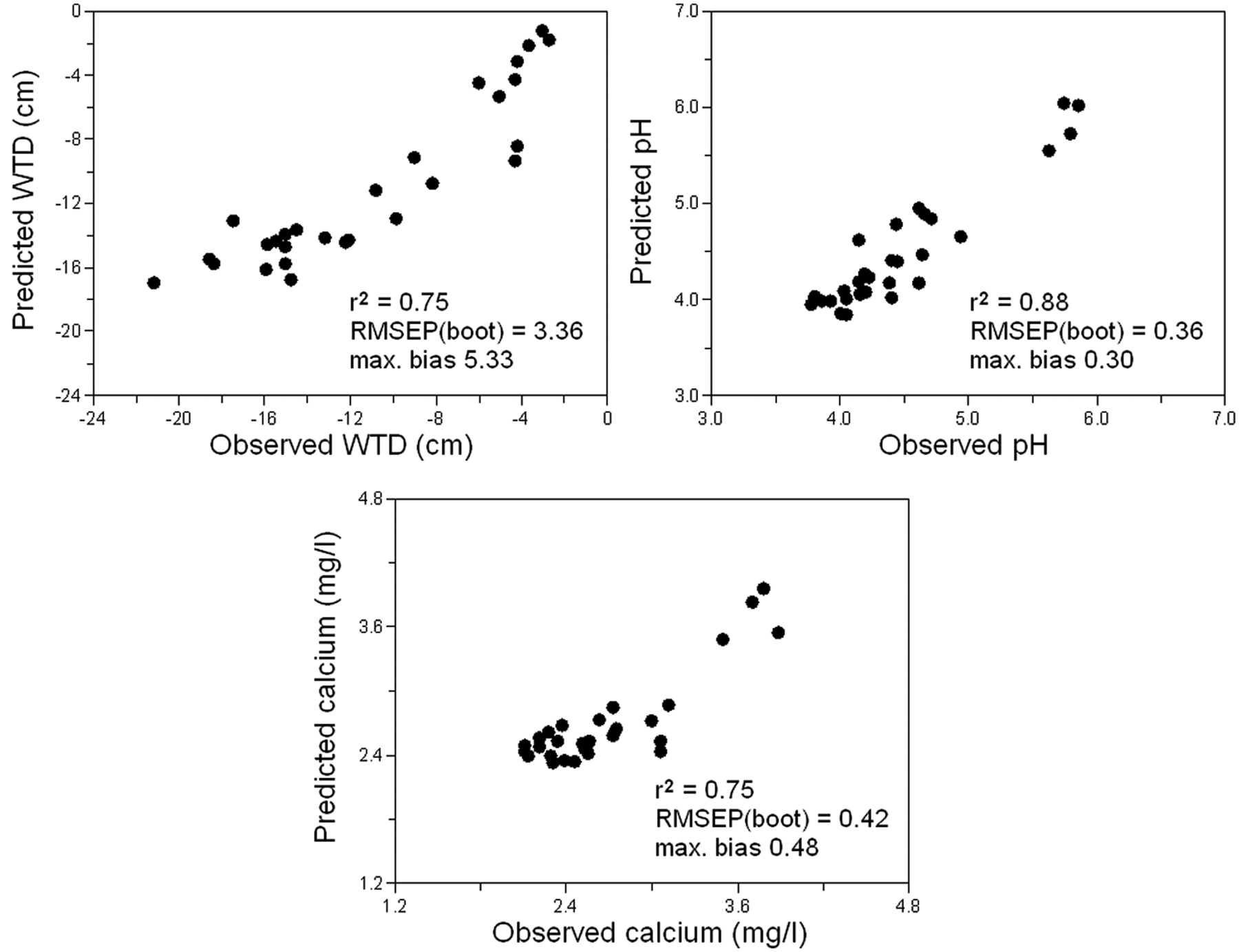
Comparison of observed and predicted values of water-table depth (WTD), pH and calcium concentrations. The model with the lowest RMSEP (weighted averaging) was used.
The predicted water pH varies only slightly, being somewhat higher in the beginning of the bog development in the vegetation dominated by Eriophorum vaginatum (4.0–4.7) than in the second zone with Sphagnum magellanicum (3.9–4.4). In the last developmental phase (the last 60 years), the predicted water pH again increases (4.2–5.1). The predicted calcium concentration fluctuates only slightly, reaching the highest values in the last developmental phase with peaks at 35 cm (3.27 mg/l) and 15 cm (3.04 mg/l) below the bog surface.
The directly measured concentrations of phosphorus and calcium were lowest in the deepest parts of the peat profile and increased upwards (Figure 6).
Discussion
Origin and development of an ombrotrophic bog ecosystem
The Vozka summit bog originated more than 6000 cal. yr ago, in the second half of the Holocene climatic optimum. In contrast, the formation of most other summit bogs in the region started much later, mostly between 850
Peat started to accumulate on very gentle slopes, most likely in a small terrain concavity where rainwater had temporarily accumulated. A presumption that summit locations exclude groundwater discharge is supported by macrofossil and local-pollen records that do not suggest the influence of spring groundwater. At the beginning of bog development, macrofossils and pollen spectra suggest the occurrence of Eriophorum vaginatum tussocks with Calluna vulgaris, Vacciniaceae and Polytrichum species, with shallow open water amongst particular tussocks, indicated by Warnstorfia fluitans leaves, Amphitrema wrightianum tests and Botryococcus cells. Such vegetation could resemble those occurring recently in bog laggs (e.g. Eriophoro vaginati-Sphagnetum recurvi) or initial poor fens (Sphagno-Caricion canescentis, for details see Chytrý, 2011). Acidicole communities of plants and testate amoebae correspond well with mineral-poor crystalline bedrock, indicated by low mineral concentrations in the two deepest peat samples (Figure 6).
As the representation of Sphagnum species was low in the first developmental stage, Eriophorum vaginatum probably played an important role in both the peat accumulation and the fen–bog transition. The analogous situation was documented by Hughes and Dumayne-Peaty (2002) in South Wales. They consider Eriophorum vaginatum to be an equally important ‘ecosystem engineer’ similar to Sphagnum species, which are generally believed to play a crucial role in autogenic bog development because of their ability to acidify the environment and isolate the mire surface from the effect of groundwater (Clymo, 1963; Walker, 1970; but see Soudzilovskaia et al., 2010). However, it cannot be excluded that space among Eriophorum tussocks was overgrown by Sphagnum magellanicum or another Sphagnum species.
In the sample from 45 to 50 cm, Sphagnum russowii increased its representation at the expense of S. magellanicum. Although Sphagnum magellanicum disappeared only at the sampling site, its significant retreat was noticed in the same region during the last decades (Hájková et al., 2011a), which may reflect either increasing nitrogen deposition or aerial liming, i.e. the processes reconstructed also by transfer-function models and enhanced mineral values in the topsoil peat samples (Figure 6).
Peat stratigraphy and reconstructed environmental conditions show a relatively stable environment (pH 4.0–5.0, calcium 2–3 mg/l, WTD 5–20 cm) supporting rather slow vegetation changes. The long-term stability of bog vegetation at the millennial scale was previously stressed by Rydin and Barber (2001), however many peat sequences from bogs reveal either development from lakes and calcium-rich fens (e.g. Kuhry et al.,1993; Ponel et al., 2011; Walker, 1970) or alternating forested and open phases (e.g. Dudová et al., 2010). The origin and development of the Vozka bog, conditioned by the climate and topography without a groundwater influence, resembles the development of blanket bogs in oceanic parts of Europe (Mauquoy and Barber, 1999; Solem, 1986) and North America (Davis, 1984; Rydin and Jeglum, 2006), and is only rarely documented from inland Europe (e.g. from extremely humid mountain regions of Scandinavia; Luoto and Seppälä, 2000). In Central Europe, similar developments from a topogenous poor fen to a Sphagnum-dominated bog have been recorded rarely, e.g. in the Jura Mountains (Sjögren and Lamentowicz, 2008), in the Vosges (Janssen et al., 1975) and in the Giant Mountains (Speranza et al., 2000). Shifts in the dominance from Cyperaceae to Sphagnum, triggered by hydrological changes, were further documented in the Swiss Alps (van der Knaap et al., 2011). Unfortunately, most of the recent studies lack a complete macrofossil record, which makes a more detailed comparison impossible.
Variation in peat decomposition and its possible causes
Variations in peat properties throughout the profile reflect different peat-forming plant communities and different conditions under which the peat accumulated. Dry bulk density, correlated with the Sphagnum abundance (Figure 2), reflects, at least partially, low decomposition rates of Sphagnum litter (Bragazza and Freeman, 2007). The bottom part of the peat section was formed of more decomposed Eriophorum vaginatum-Polytrichum peat. Besides good decomposability of vascular plant litter, a high degree of peat decomposition may be explained by fluctuating water regime. Polytrichum usually indicates a fluctuating water-table (Laitinen et al., 2008), which might have occurred in the initial phase of the Vozka bog development because of the exclusive dependence of local vegetation on rain and surface water. In addition, Eriophorum-peat exhibits lower water retention abilities compared with Sphagnum-peat. Testate amoebae indicate rather high water-tables in the initial stage, however they might not reflect short-term water-table declines because of their quick recolonisation (Laggoun-Défarge et al., 2008).
An abrupt change in the ash content and dry bulk density, indicating an increased rate of peat decomposition, was found at a depth of about 50 cm (Figure 2). Moreover, there is a large difference in the age of samples from 50 and 40 cm (c. 600 cal. yr). There are two possible explanations of this difference: either peat accumulated extremely slowly under unfavourable conditions (e.g. low water levels) or there is a hiatus in peat sequence caused by either the decomposition of previously accumulated peat or erosion. Recent studies show that the occurrence of a decomposed peat layer close to the mire surface is a general phenomenon in the Alps (Sjögren et al., 2007) and in southern Sweden (Franzén, 2006). In the Alps, pastoralism connected with trampling and fertilization was identified as the most probable reason for this stratigraphical pattern (Sjögren et al., 2007) thanks to pollen indicators and spores of coprophilous fungi found in the particular layers. By analogy, fertilization is assumed to be the main reason for the secondary decomposition of subsurface peat layers in southern Sweden (Franzén, 2006). In the vicinity of the Vozka bog several pastures and meadows were recorded
Regional vegetation history and its relationship to settlement
The results presented here suggest that the bog development was largely independent of regional vegetation history (Figure 6). Changes in the pollen spectra of the Vozka bog, providing a record of high mountain vegetation changes since the middle Holocene, put some older results into a new perspective. Large amounts of Picea, Corylus, Ulmus and other broadleaved tree pollen, found in the oldest peat layers, are also reported from the eastern part of the region (Rybníček and Rybníčková, 2004) as well as from the Western Sudetes (Jankovská, 2007a; Speranza et al., 2000; Svobodová, 2004) for the same period. Mixed spruce woodlands were reconstructed with an admixture of elm, lime and sycamore in the vicinity of the Vozka site during the rather wet and warm middle Holocene (zone VOp-1). The dominant role of spruce is supported by macrofossils found in the peat (Figure 4). The high amount of Corylus pollen probably reflects the occurrence of hazel in the surrounding supramontane forests. Rybníček and Rybníčková (2004) presented the hypothesis of shrubby hazel stands occurring above the treeline and hence vicarising Pinus mugo which is naturally absent from the Hrubý Jeseník Mountains. However, the relative representation of hazel pollen in peat sequences from the Krkonoše Mountains, where Pinus mugo naturally occurs, is similar to that found in the Vozka bog (Jankovská, 2007a; Speranza et al., 2000; Svobodová, 2004). Therefore, it can be presumed that Corylus was an admixture of mixed spruce woodlands below the treeline. Alternatively, hazel could play the role of pioneer species at disturbed patches and natural openings. The regular presence of hazel in zonal mountain woodlands is further supported by pollen assemblage for consequent time periods, when a fundamental transformation of forest vegetation took place: some woodland species (Ulmus, Acer) were replaced by Fagus and Abies, while others (Picea) reduced their altitudinal range. According to the results, this transformation was relatively slow and gradual, spanning approximately 1600 years (VOp-2, 2390–800
The mixed spruce woodlands of the middle Holocene probably had a rather open tree canopy enabling the regular presence of Corylus avellana. This hypothesis is supported by the occurrence of Juniperus, which can grow directly in supramontane woodlands close to the treeline. Alternatively, juniper might only grow in treeless habitats and its rather high presence in the pollen spectrum may merely reflect good pollen dispersal throughout open woodlands.
Since 800
Although a distinctive human impact on the high mountain vegetation has been traditionally expected since the 13th century
Around
As the peat section is reliably dated to approximately
Occurrence of Larix
One of the most important pollen records is that of Larix decidua. While in the Western Carpathians larch occurred frequently during the last glacial and persisted there until the present day (Jankovská, 2007b; Jankovská and Pokorný, 2008), in the neighbouring Bohemian Massif it probably disappeared during the Lateglacial/Holocene transition (Jankovská, 2007b). Nevertheless, according to written sources, larch wood from the Jeseníky Mountains was used in the 16th century (Mathiolli, in Nožička, 1957). The Jeseníky Mountains are therefore considered as the only region of the Bohemian Massif where Larix survived throughout the entire Holocene, but direct evidence is still missing. The pollen grain of Larix found at approximately 580
The Vozka bog in a broader context
The origin of the Vozka bog is dated back to 4180

Summarized development of local and regional vegetation of the Vozka bog based on all biotic proxies (macrofossils, testate amoebae and regional pollen).
Changes in the surrounding vegetation generally coincide with main climate variations during the Holocene. The gradual substitution of forest dominants lasting from 2390
Although changes in the local and regional vegetation composition at the Vozka bog are influenced mostly by climate (until Medieval time), they are not synchronous. It is suggested that this is a consequence of different sensitivities to climatic events: local vegetation may change under several extreme years, whereas the transformation of zonal vegetation, dominated by long-living trees, is possible only under long-term climatic conditions.
Footnotes
Acknowledgements
We are grateful to Martin Kočí, Radim Hédl, Dušan Lekeš, Ivan Tůma and Petr Holub for help with taking the profile, to Zuzana Formánková for laboratory work, and to the staff of the Administration of the Protected Landscape Area Jeseníky for their support and permission to research. We thank Jakub Halama, Mariusz Lamentowicz and Pim van der Knaap for useful comments. Special thanks to Jacqueline van Leeuwen for help with pollen identification. We are grateful to Michal Hájek, Per Sjögren and an anonymous reviewer for critical revision of the manuscript.
Funding
The research was supported by the grant project GA206/08/0389, institutional support of Masaryk University and long-term research development project no. RVO 67985939.