
Abstract
Ancient Loulan, an important city on the Silk Road, disappeared about 1500 years ago. The environmental conditions associated with the vicissitude of ancient Loulan have been debated since the city was rediscovered in ad 1900. However, little paleobotanical evidence concerning vegetation and environment in this area has so far been available. In this study, phytoliths and diatoms extracted from 16 samples including two fossil camel coprolites from sites of Loulan and Milan indicate that the landscape of ancient Loulan was a typical oasis, where reeds, grasses of Paniceae and Pooideae probably grew along with some shrubs. Also, some typical brackish diatoms might live in some water bodies in the catchment of an ancient lake, Lop Nor. Our results also suggest that common millet as staple crop, and foxtail millet and possibly naked barley as non-staple crops were the main food source for ancient Loulan and Milan residents between ad 50 and 770.
Introduction
The ancient city of Loulan is located on the western bank of a now dried-up lake, Lop Nor, in Xinjiang, China (Figure 1). It was once the political, economic, and cultural center of West China from c. 77 bc to ad 550 (Ban, 92; Xia et al., 2007). The ancient city, a staging post for traders and migrants, was also an important junction between central and western China on the middle route of the Silk Road (Figure 1). However, this once prosperous city became a desert and depopulated zone filled with wind-eroded gravels after the Buddhist monk Xuanzang passed through the area in about ad 645 during the Chinese Tang Dynasty (Xuanzang, 645).
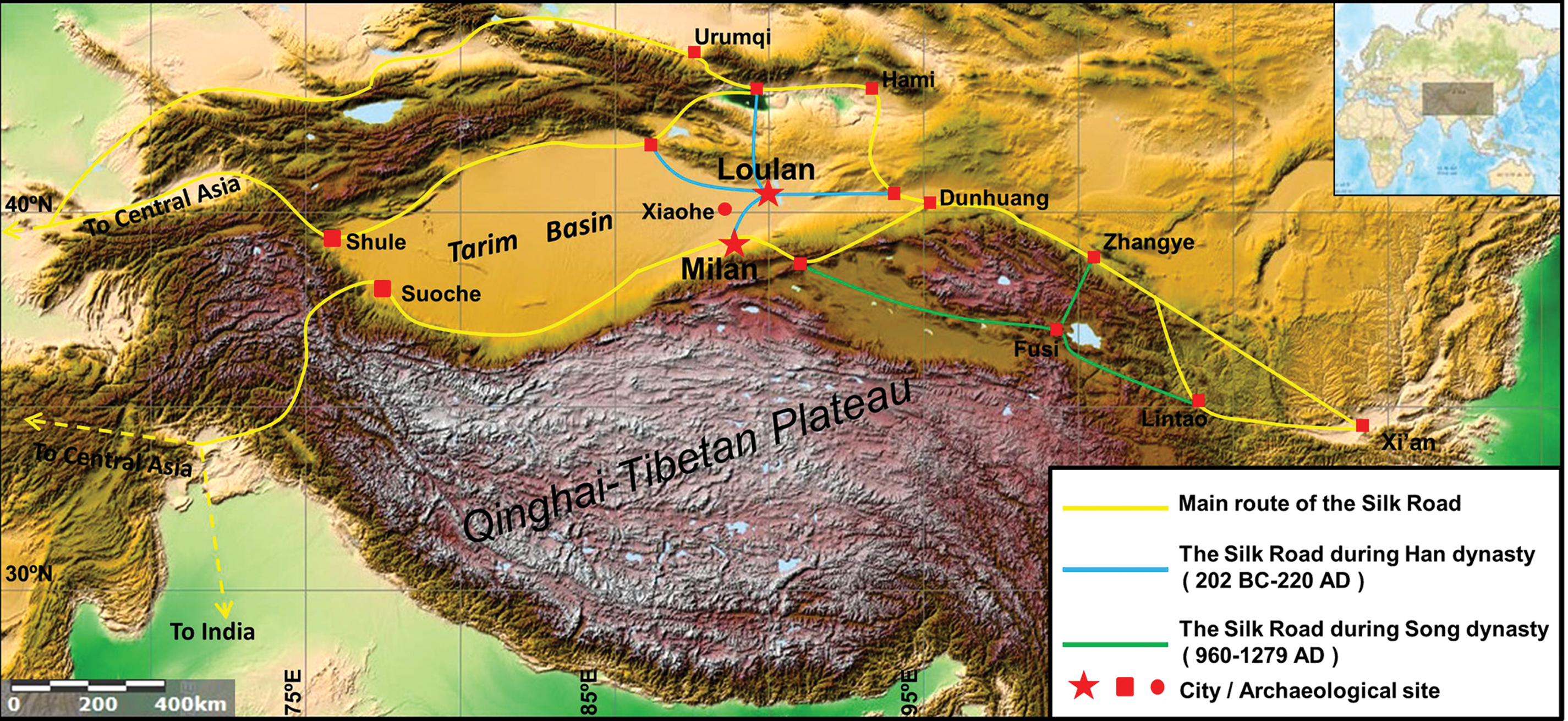
Locations of the ancient cities of Loulan and Milan and the Silk Road in different periods. The base map was from http://maps.ngdc.noaa.gov/viewers/bathymetry. After Lan (2002) and Wu (2004).
In the past century, scholars from different disciplines have carried out a wide range of investigations on causes for the decline of Loulan (Cheng, 1936; Huntington, 1907; Village, 1955; Xia et al., 2007) and proposed several hypotheses, such as climate aridification (Huntington, 1907) and river diversion (Stein, 1928). Despite efforts over 100 years, environment and vegetation (especially crop) changes are still little investigated because of the difficulty of acquiring samples (access to this area) and the lack of accurate age data (Jia et al., 2010; Wang et al., 2008; Wu, 1994); and as a result environmental conditions and agricultural activities in ancient Loulan City thus remain ambiguous.
Active agriculture in ancient Loulan was documented in the historical literature (Ban, 92), and common millet, naked barley, and wheat were also found there (Hou, 1985; Xia et al., 2007). However, few studies focused on agriculture in ancient Loulan, and they did not provide substantial diagnostic features for these crops. Therefore, more evidence, especially paleobotanical evidence, is essential for this issue. At present, it is very difficult to find suitable research materials with paleoenvironmental information on ancient Loulan because of extreme aridity, wind erosion, and salinization. For addressing this issue, camel coprolites and some archaeological samples buried in the cultural layer of ancient Loulan provide us with a unique opportunity to reveal the paleoenvironment and agriculture of that time.
Camels were the principal means of locomotion on the Silk Road in the past (Potts, 2005). The forage they ate can be considered, to some extent, a reflection of local vegetation and thus environmental conditions by inference. This information can be obtained from microfossils, such as phytoliths and diatoms from camels’ coprolites. Since phytoliths are replicas of plant cell bodies, phytolith analysis is a useful technique to identify certain genera or species of plants (Lu et al., 2005; Pearsall, 2000; Piperno, 2006). For example, many staple crops, e.g. maize (Piperno, 1984), wheat and barley (Ball et al., 1999, 2009; Rosen, 1992; Rosen and Weiner, 1994), common millet, foxtail millet, and related wild grasses (Lu et al., 2009; Zhang et al., 2011) can be specifically identified by phytoliths. Diatoms are microscopic aquatic and subaquatic algae common in a range of aquatic environments, including streams, ponds, lakes, and estuaries (Round et al., 1990; Werner, 1977), and their preserved remains are composed of silica, as are phytoliths. Phytoliths and diatoms can be extracted from many materials including coprolites (Piperno, 1988), and provide information on local vegetation, environment, and diet (Bryant and Williams-Dean, 1975; Horrocks et al., 2003; Pearsall, 2000; Piperno, 1988).
We conducted phytolith and diatom analysis on 15 fossil samples (two samples of camel coprolites and 13 archaeological samples dated from c. ad 50 to 770) and one modern sample of camel dung from two archaeological sites, Loulan (LA) and Milan (ML), in an attempt to reveal the profile of environment and agriculture of ancient Loulan and Milan on the Silk Road.
Materials and methods
Field investigations on the ruins of ancient Loulan and Milan were carried out in November and December 2008. A total of 16 samples were collected from the two archaeological sites.
It is generally accepted that a close relationship between the Loulan Kingdom and the central government of China had been established during the Han Dynasty (202 bc–ad 220) (Romgard, 2008; Xia et al., 2007; Xiao, 2006). Ancient Loulan city (LA site) (40°30′58.73″N, 89°54′51.73″E), located at the eastern end of the Tarim Basin, was a political and military center and the east–west cultural exchange point in the western region of China (Investigation Team of Cultural Relics in Luolan, 1988; Romgard, 2008; Wang, 1996; Xia et al., 2007; Xiao, 2006; Yidilisi and Zhang, 1997). The main constructions in the northeastern part of the LA site include the stupa (sacred Buddhist site) tower and several other buildings (Figure 2a). Three samples (LL3, LL4, LL5) were collected from the tower base (Figure 2b), which is square in shape. The charcoals from samples LL3 and LL5 were dated to ad 185 ± 43 and 230 ± 66, respectively (Lu et al., 2010). In the southwestern part is the famous ‘Three Rooms’ site. Three samples (LA-R1, LA-R2, LA-R3) were taken from a rubbish area 50 m from its southwest side (Figure 2c). The wood from the west room was dated to ad 66 ± 146 (Lu et al., 2010). Two samples (LA-C1 and LA-C2) of camel coprolites were taken from the livestock stall at the west side of the ‘Three Rooms’ (Figure 3). They were dated to ad 89 ± 118 (Lu et al., 2010). For comparison, a sample (LA-C3) of modern wild camel excrement, composed of four individual dung balls, was also collected from the location 2 km northeast of the Loulan site.
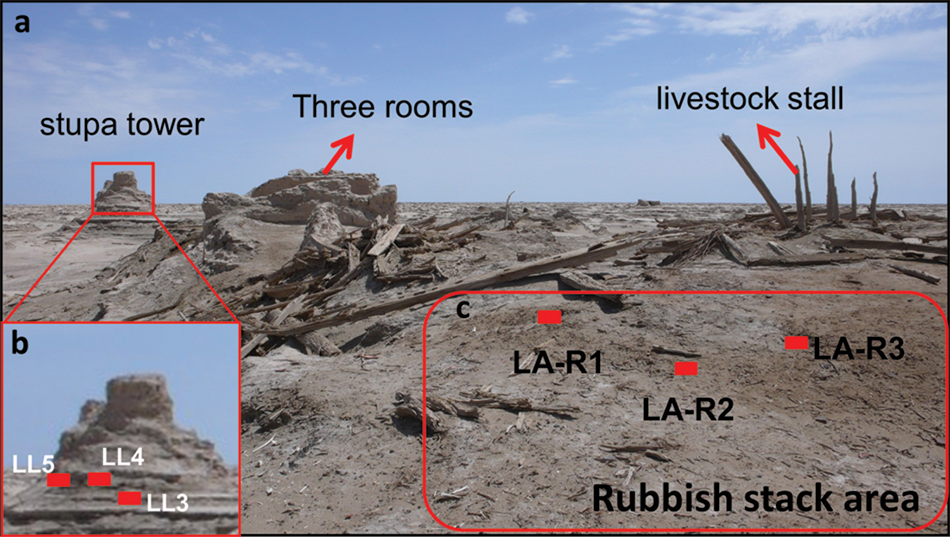
(a) The main remains of ancient Loulan City; (b) sampling locations at the stupa tower; (c) sampling locations at the rubbish stack area.

(a) Sampling location of camel coprolites relative to the ‘Three Rooms’; (b) zoom view of sampling location; (c) samples of camel coprolites.
In addition, three samples (LL2: 40°31.027′N, 89°57.344′E, altitude 781 m; LL6: 40°30.924′N, 89°57.344′E, altitude 774 m; and LL7: 40°31.052′N, 89°58.315′E, altitude 776 m) were collected from probable farmlands of ancient Loulan City which were abandoned at least at 1500 years ago (c. ad 500) (Qin et al., 2011), and one sample (LL12: 40°31.938′N, 89°59.025′E, altitude 789 m) was collected from an ancient tomb dated to ad 50 ± 37 (Lu et al., 2010). LL1 (40°31.973′N, 90°00.823′E, altitude 790 m) is a soil sample collected from the Loulan culture layer (Figure 4).
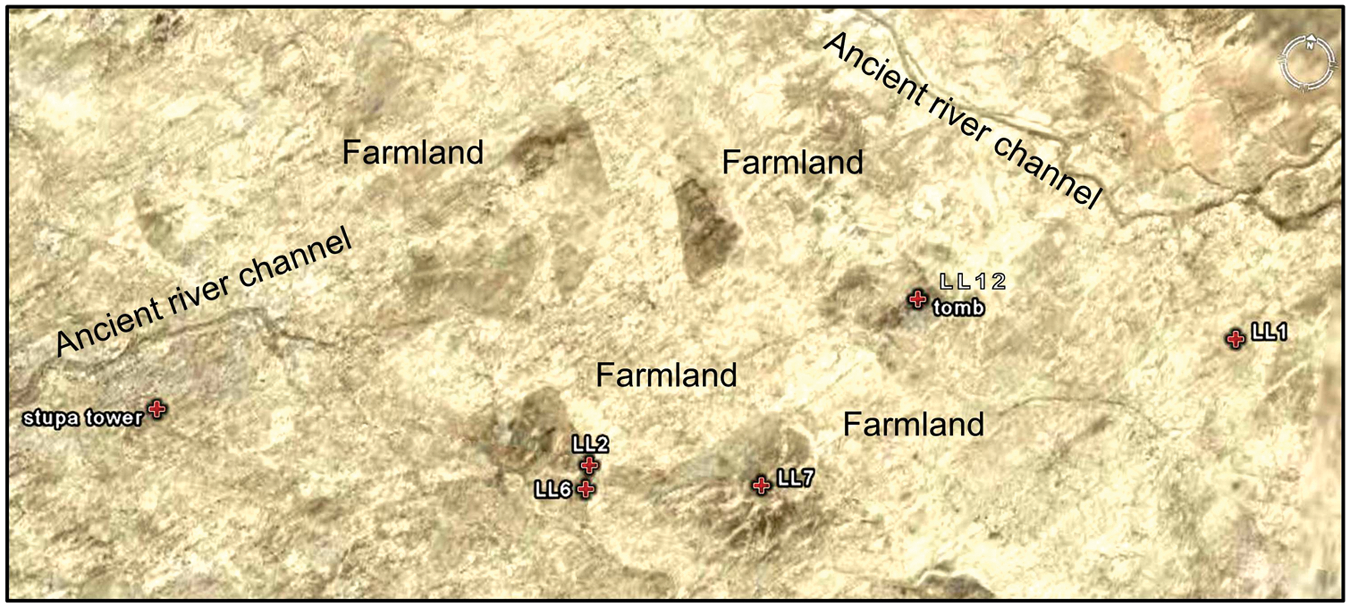
Location of possible farmlands in ancient Loulan city and sampling sites. Image from map.Google.com. After Qin et al. (2011).
The Milan site, located c. 160 km south of the Loulan site, has been considered to be the ancient city of Yixun in the Loulan Kingdom. The current conservation area of the Milan historical site is about 44.5 km2. It consists of eight stupas, three Buddhist temples, two beacons, the water protection facilities, and the ancient fortress. The ancient fortress (39°13′36.23″N, 88°58′14.83″E, altitude 926 m) presents an irregular square. The 1 m thick wall base of the north side at its west entrance is composed of plant debris, charcoal, and soil. Two samples (ML1, ML2) were taken from the levels at c. 20 and 15 cm above the bottom of the wall base (Figure 5). The 14C age at 20 cm level is ad 777 ± 65 (Lu et al., 2010).

(a) The wall base at the north side of ancient Milan city; (b) sampling locations. After Lu et al. (2010).
As both phytolith and diatom remains are composed of silica nearly with the same physical properties, they are extracted using the same process. Each sample (~2 g) was prepared using the technique described by Pearsall (2000), Piperno (1988), and Runge et al. (1999) with slight modifications. The procedure consists of sodium pyrophosphate (Na4P2O7) deflocculation, treatment with 30% hydrogen peroxide (H2O2) and cold 15% hydrochloric acid (HCl), separation with zinc bromide (ZnBr2, density 2.35 g/cm3) heavy liquid, and mounting on a slide with Canada Balsam. Phytolith and diatom identification and counting were performed using a Leica microscope with phase-contrast at 400× magnification. Identification was aided by the use of reference materials (Lu, 1998; Lu and Liu, 2003a, 2003b; Lu et al., 2006) and published keys (Denys, 1991; Kondo et al., 1994; Mulholland and Rapp, 1992; Pearsall, 2000; Piperno, 1988; Piperno and Pearsall, 1998; Runge et al., 1999). The phytoliths were named according to
In order to verify the identification of wheat husk phytoliths in archaeological samples, modern wheat (Triticum aestivum L.) (collected at 36°31′14.00″N, 107°15′35.00″E, altitude 1220 m, Shannxi Province, China), barley (Hordeum vulgare L.) (donated by the Chinese Academy of Agricultural Sciences), and naked barley (Hordeum vulgare var. nudum) grain husks were also prepared for comparison. The procedure used for modern phytolith analysis is the technique described by Lu et al. (2009).
Results
Three fossil samples (LL1, LL2, LL4), and one modern sample of wild camel excrement (LA-C3) do not contain any phytoliths. Other fossil samples have sufficient phytoliths for correct identification. More than 400 phytoliths were counted in the most of samples except samples LL5 (87) and LL6 (314). Our results show that crop phytoliths are common millet and foxtail millet (Figure 6). The common non-crop phytolith types are scutiform-bulliform (reed bulliform), middle saddle, rondel, bilobate, trapeziform sinuate (tooth), smooth elongate, elongate-echinate, acicular hair cell (point shaped), trapeziform (square). These samples also contain some wood and unidentified phytoliths (Figure 7) as well as some dendritic phytoliths that need further comparative study (Figure 8). Diatoms are found in just seven samples, but they are especially abundant in camel coprolites (Figure 9).

Crop phytoliths of fossil samples from ancient Loulan and Milan: (a)–(c) phytoliths from common millet husk; (d) phytolith from foxtail millet husk.
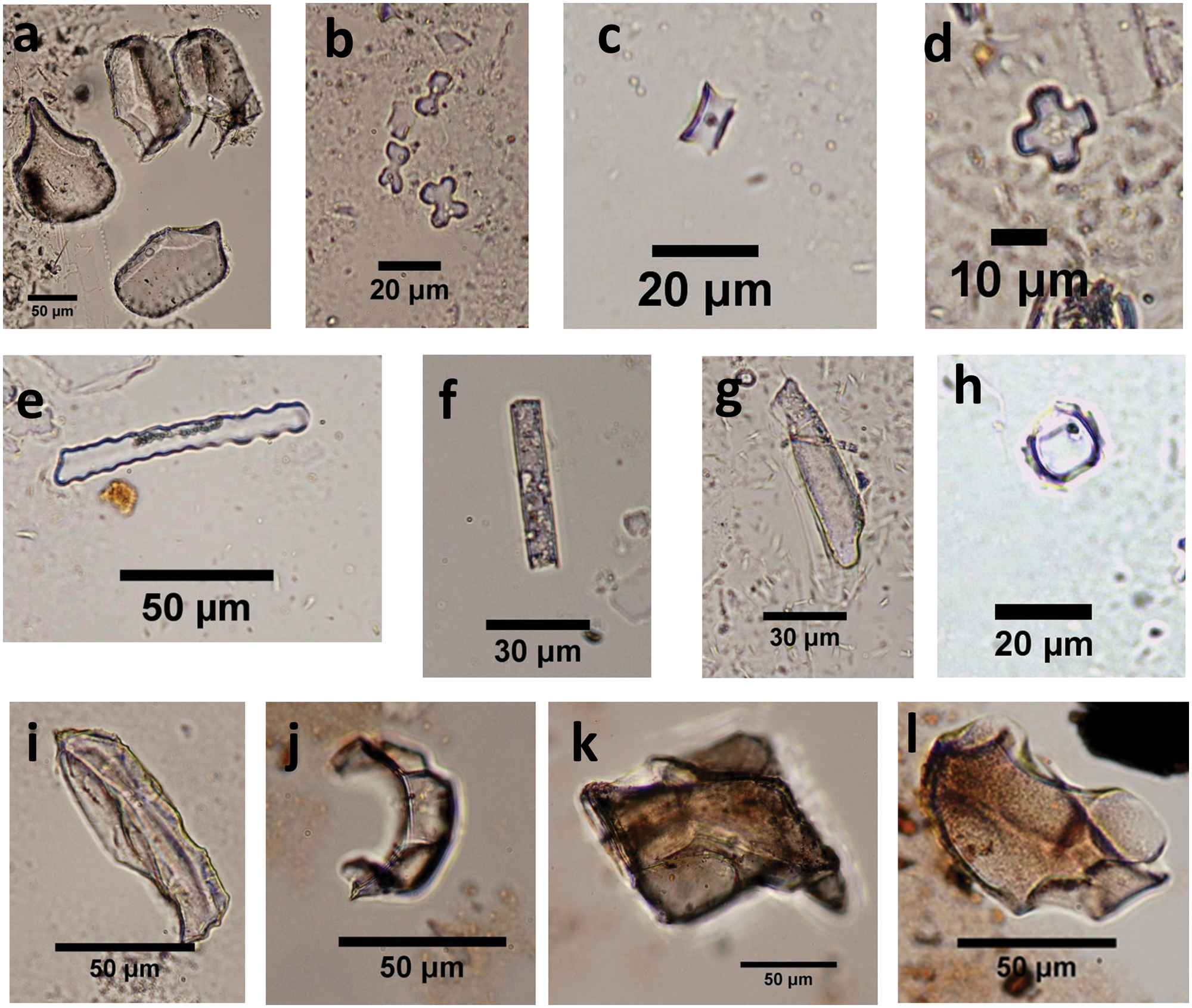
Common phytolith types of fossil samples from ancient Loulan and Milan: (a) scutiform-bulliform; (b) bilobate; (c) rondel; (d) cross; (e) trapeziform sinuate; (f) smooth elongate; (g) acicular hair cell; (h) middle saddle; (i)–(l) phytoliths from wood.
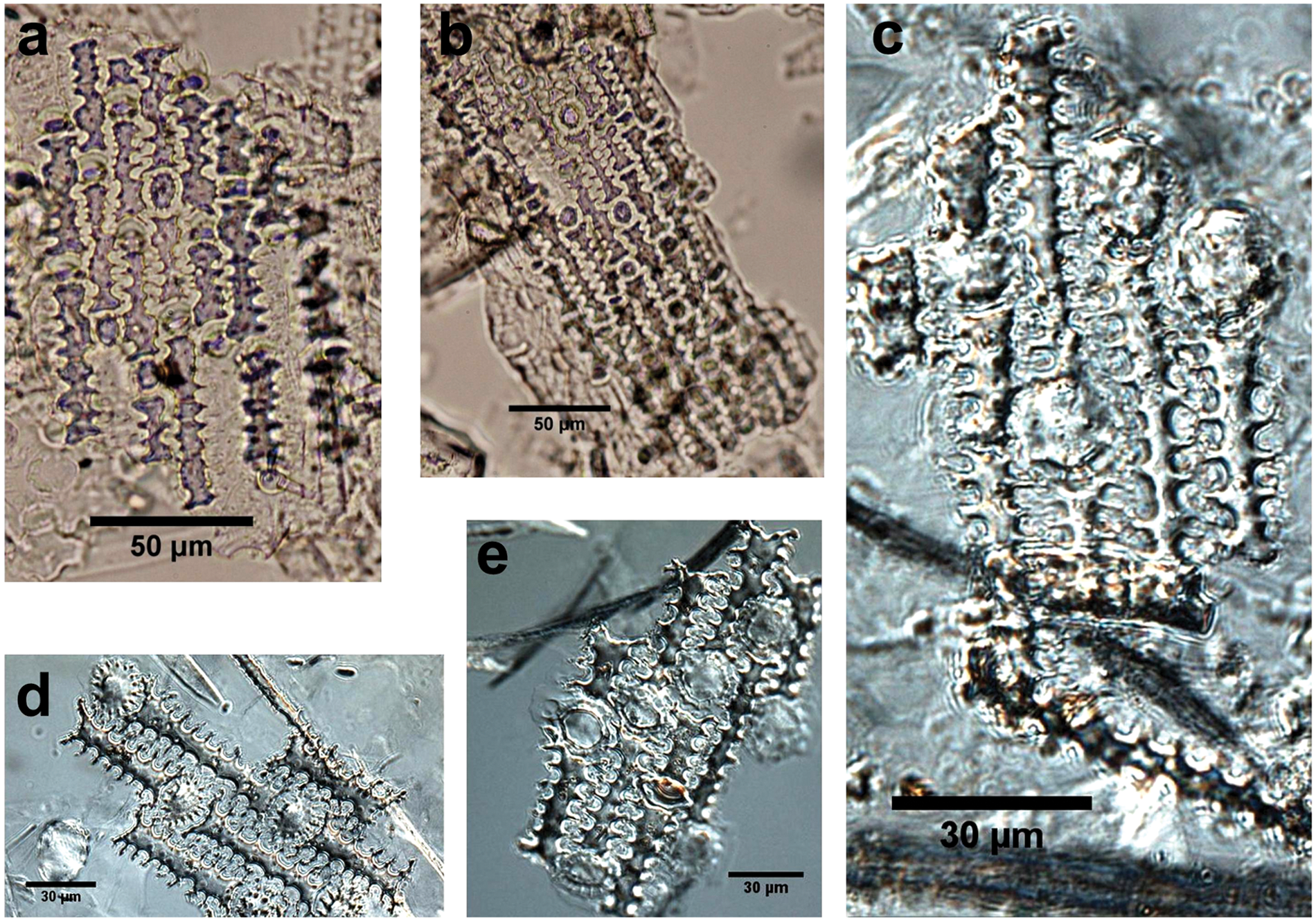
Fossil dendritic phytoliths from ancient Loulan and modern dendritic phytoliths: (a), (b) dendritic phytoliths from Loulan site; (c) phytoliths from modern naked barley (Hordeum vulgare var. nudum) husk; (d) phytolith from modern wheat (Triticum aestivum L.) husk; (e) phytolith from modern barley (Hordeum vulgare L.) husk.
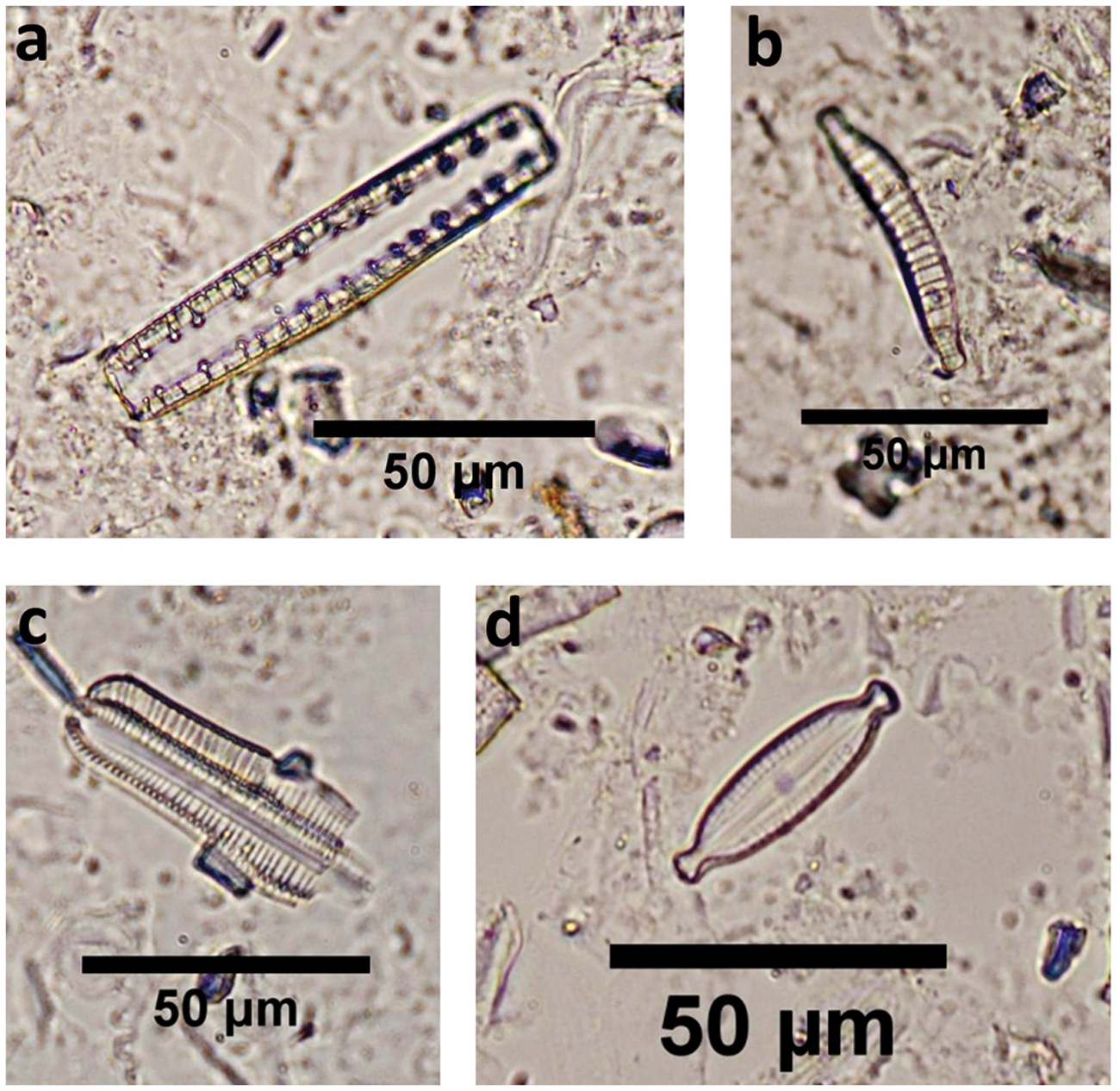
Diatoms from camel coprolites and archeological samples of ancient Loulan and Milan: (a), (b) Epithemia cf. adanata; (c) Rophalodia gibba; (d) Mastogloia smithii var. amphicephala.
According to the reference of phytolith taxa in China (Lu et al., 2006; Wang and Lu, 1993), specific phytolith types are predominantly (although not restricted to) produced by certain plants. For example, scutiform-bulliform and middle saddle are from reed, trapeziform sinuate and rondel from Pooideae, and bilobate from Paniceae. Therefore, we also use plant names in the following description of phytolith assemblages from these fossil samples (Figure 10a, b). Phytolith types without any systematic significance, i.e. smooth elongate and elongate echinate, are not shown in Figure 10.
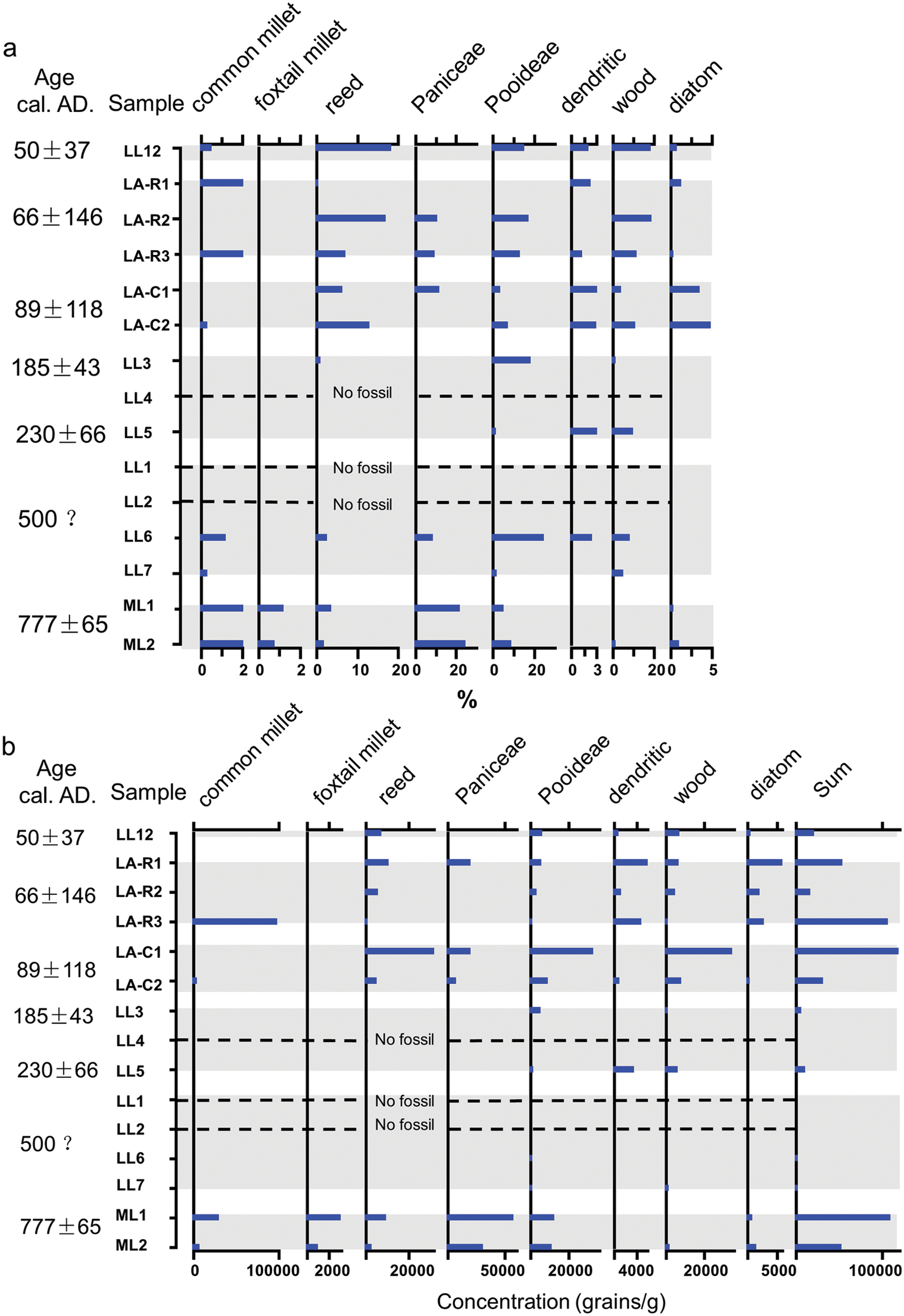
(a) Percentages of main phytoliths in fossil samples from ancient Loulan and Milan; (b) concentrations of main phytoliths in fossil samples from ancient Loulan and Milan.
Phytoliths from the ancient tomb (sample LL12; c. ad 50): The percentage of reed phytolith is highest (13.2% of scutiform-bulliform and 5.0% of middle saddle) among all samples. Pooideae (14.6%) and wood (17.8%) phytoliths are also dominant. Only a few common millet husk phytoliths (0.5%) are present in this sample. The phytolith concentration is low.
Phytoliths from the west side of the ‘Three Rooms’ at the Loulan site (samples LA-R1, LA-R2, and LA-R3; c. ad 70): Three samples have relatively high phytolith concentration, especially samples LA-R1 and LA-R2. The percentage of common millet husk phytoliths in sample LA-R1 is highest (45.6%) among all samples, whereas its percentage is low (4.1%) in sample LA-R3 and it even does not show up in sample LA-R2. In samples LA-R2 and LA-R3, reed, Paniceae, Pooideae, and woody phytoliths are present frequently. The percentages of scutiform-bulliform and middle saddle from reed are as high as 10.9% and 5.8%, respectively. In the three samples, no foxtail millet, typical wheat and barley are found.
Phytoliths from camel coprolites (samples LA-C1 and LA-C2; c. ad 90): The phytolith assemblages of two samples are characterized by relatively high percentages of reed phylitoths. The scutiform-bulliform accounts for 6.0% and 12.9%, and the middle saddle accounts for 1.4% and 3.1% of total phytoliths, respectively. Other relatively abundant phytoliths are three specific types, i.e. Paniceae (11.3% and 0%), Pooideae (3.0% and 6.7%), and wood (3.7% and 10.8%). The phytolith concentrations of two samples are relatively low.
Phytoliths from the stupa tower base (samples LL3, LL4 and LL5; c. ad 185–230): No phytolith is found in sample LL4, and the phytolith concentrations of samples LL3 and LL5 are relatively low. Phytolith content of two samples with fossils is relatively simple, mainly including Pooideae, wood and dendritic phytolith. Pooideae phytoliths decrease sharply from 17.8% in sample LL3 to 1.3% in sample LL5, whereas wood phytoliths increase from 1.0% in sample LL3 to 9.6% in sample LL5. At this site, reed phytoliths are negligible, only sample LL3 contains 0.8% of scutiform- bulliform. Some types without any systematic significance such as smooth elongate and elongate echinate are also present.
Phytoliths from possible ancient farmland and the Loulan culture layer (samples LL2, LL6, LL7 and LL1; c. ad 500?): No phytolith is found in sample LL1 and LL2, and the phytolith concentrations of samples LL6 and LL7 are very low. Common millets are quite scarce (1%), only found in sample LL6. The dominant types are Pooideae (24.1% and 1.7% in samples LL6 and LL7), Paniceae (8.0% and 0%), wood (8.0% and 4.6%), and reed (2.3% and 0%).
Phytoliths from the Milan site (samples ML1 and ML2; c. ad 780): Paniceae dominates the phytolith assemblages (21.5% and 24.0%). Remarkably, these two samples contain not only abundant common millet husks (10.9% and 5.0%) but also a few foxtail millet husk (1.2% and 0.7%). Their phytolith concentrations are high.
Dendritic phytoliths are found in most of the samples at the Loulan site, especially in samples LL5, LA-C1, LA-C2 (5.7%, 3.5% and 2.9%). However, no dendritic phytolith is present in samples at the Milan site.
Diatoms in ancient camel coprolites (LA-C1, LA-C2) have relatively high percentages at 3.5% and 4.8% (the percentage expressed as all phytoliths counted), respectively. In other samples, diatoms are few (c. 1% or less). All diatoms belong to brackish water species, including Rophalodia gibba, Epithemia cf. adanata, and Mastogloia smithii var. amphicephala.
Discussion
Vegetation and environment in ancient Loulan City
Many phytolith types, including bilobate, scutiform-bulliform, rondel, middle saddle, smooth elongate, trapeziform, trapeziform sinuate, wood, and dendritic are found in coprolites and archaeological samples. Most of these phytoliths are from grasses in terms of the reference of phytolith types in China (Lu et al., 2006). The scutiform-bulliform type is probably from Phragmites australis (reed), as its dimension (about 90 μm in length and 60 μm in width) is distinct from other bulliforms from Oryzoideae, Bambusoideae, and Eragrostidoideae in China (Wang and Lu, 1993). And this type is also present along with middle saddles (about 12 μm in length), which occurs predominantly in Phragmites australis (up to about 10% in abundance) (Wang and Lu, 1993). Middle saddle can be distinguished from long saddle (longer than 18 μm) and short saddle (10 μm in length on average). The former predominates in phytoliths from Bambusoideae (Li et al., 2005), whereas the latter is common in phytoliths from Eragrostidoideae (Wang and Lu, 1993).
Two types of bilobates occur in these samples. One type has smooth rounded lobes, and another type has branched outer margins at the ends of the lobes. These two types of bilobates predominantly represent C4 grasses of Paniceae, which grow in warm and arid to semi-arid regions with low soil moisture (Lu and Liu, 2003a). Trapeziform sinuate and rondel types mostly occur in phytoliths of Pooideae that grow in dry and cold regions with a mean annual precipitation less than 450 mm (Lu et al., 2006). Wood phytoliths with irregular shapes are from broadleaved trees (Kondo et al., 1994; Lu et al., 2006; Piperno, 1988; Runge et al., 1999). From the above discussion, it can be speculated that reeds, grasses of Paniceae and Pooideae, and shrubs might occupy the landscape around the ancient Loulan City during its early period from AD 50 ± 37 to 230 ± 66.
The investigation of vegetation in the present Loulan region is difficult because of the harsh natural environment. Fortunately, modern wild camel dung can provide some valuable information. Modern wild camels are mainly found in the Jiashun gobi area of northern Lop Nor, northern Altun Mountains, and eastern Taklimakan Desert (Gao et al., 2003). The composition of their food depends on the types and content of vegetation in each region (Zhang and Yuan, 1997). In Jiashun gobi, plants of Calligonum sp., Halogeton sp., Halostachys sp., Nitraria sp., and reeds are the selections of choice for camels (Zhang and Yuan, 1997). All these plants, except reeds, rarely produce phytoliths (Piperno, 1988). Moreover, the dry environment and high soil pH around Lop Nor reduce the dissolution of silica and thus limit the assimilation of silica by plants (Piperno, 1988, 2006; Xia et al., 2007). Therefore, it is not surprising that no phytolith is found in modern wild camel excrement. However, this fact at least indicates that Calligonum sp., Halogeton sp., Halostachys sp., and Nitraria sp. seldomly grow and reeds do not grow in the present Luolan region, since contemporary wild camels are found there. The footprints of wild camel are found in the present Loulan site (not shown).
A comparison of paleovegetation inferred from fossil phytolith assemblages from ancient wild camel excrement with modern vegetation (Figure 11) shows significant differences. It is evident that vegetation consisting of reeds, grasses of Paniceae and Pooideae, and shrubs during the early period of ancient Loulan City was eventually replaced by desert. This vegetation change reflects the deterioration of environmental conditions around the ancient Loulan City. However, this study cannot provide substantial evidence to determine the timing of this replacement, the temporal pattern of this replacement (abrupt or gradual), and the association of this replacement with the decline of the ancient Luolan City because of the discontinuity in sampling and short temporal coverage of samples, although our study reveals the occurrence of such a replacement from a typical oasis to desert during the last 2000 years. Continuous and high-resolution records of vegetation and climate covering the last 2000 years are necessary to solve these issues. Although no such record is available in this region, some records in adjacent regions (Shao et al., 2005; Xu, 1998; Zhao et al., 2007, 2009, 2010) might provide some insights into these issues. A high resolution pollen record from Hurleg Lake in the Qaidam Basin shows the occurrence of a major drought between ad 650 and 800, indicated by very low values of the A/C (Artemisia/Chenopodiaceae) ratio, an index of regional vegetation and effective moisture changes (Zhao et al., 2010). This persistent major drought also exhibits in a 1437 yr record of annual precipitation derived from tree rings in the northeastern Qaidam Basin (Shao et al., 2005). It seems reasonable to speculate that this persistent major drought might be one of the factors causing vegetation replacement and decline in the ancient Loulan area.
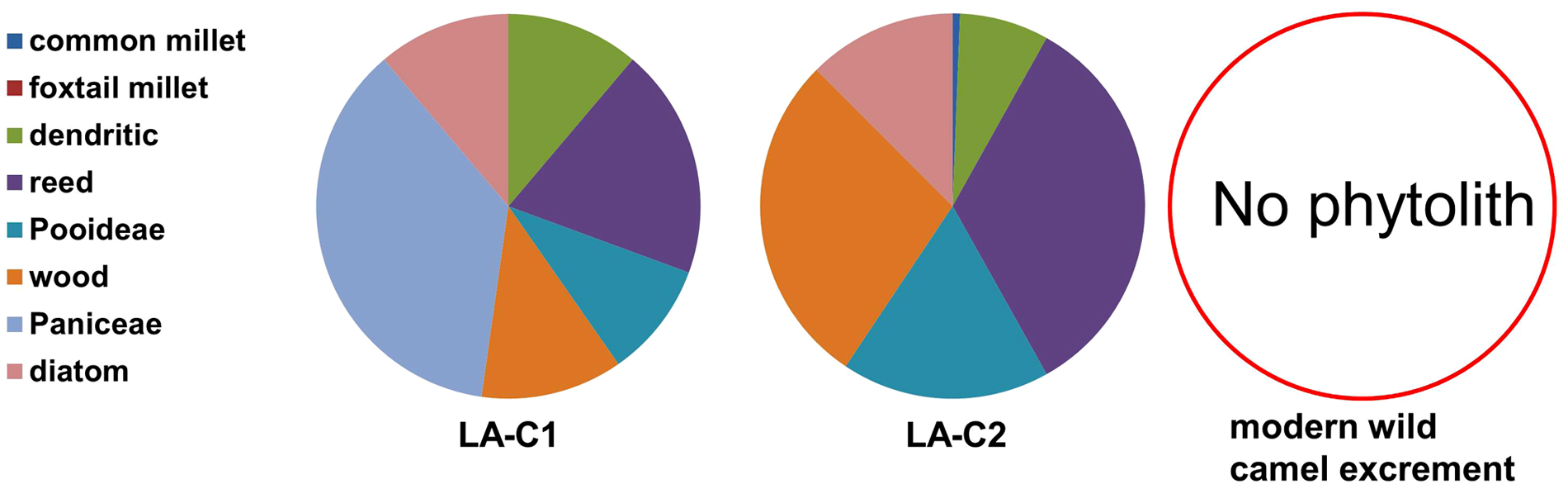
A comparison of phytolith assemblages from ancient and modern wild camel excrement.
Brackish water diatoms, including Rophalodia gibba, Epithemia cf. adanata, and Mastogloia smithii var. amphicephala, are found in samples LA-C1 and LA-C2. One possible explanation of this diatom result is that camels mainly drank brackish water and reeds might also have grown in brackish water around ancient Loulan city as far back as ad 89 ± 118. As camels ate reeds or drank water, diatoms growing in the brackish water or adhering to the reeds were absorbed and subsequently excreted as coprolites by them.
Crops in ancient Loulan and Milan
Phytolith results indicate that common millet was the dominant food crop for the residents (45.6% in LA-R1, 10.9% in ML1) in the ancient cities of Loulan and Milan. Interestingly, foxtail millet was present in quantities of about 1% from ML1 and ML2 in ancient Milan. This fact suggests that Milan in c. ad 780 had relatively greater water resources than Loulan at the period of ad 50–230, since foxtail millet requires more water than common millet to grow well (Li and Wu, 1996; Nesbitt, 2005; You, 1993). An alternative explanation is that this crop was introduced into Milan after the disappearance of the ancient city of Loulan.
The origin of the small quantity of dendritic phytoliths in fossil samples is unknown. A comparison of them with modern references and published keys (Ball et al., 1996, 1999, 2009; Berlin et al., 2003; Rosen, 1992; Rosen and Weiner, 1994; Tubb et al., 1993) shows that the dendritic phytoliths from the Loulan site do not have typical diagnostic features of wheat or barley based on Rosen’s criteria (Figure 8) (Rosen, 1992; Rosen and Weiner, 1994). However, their shapes are very similar to those of modern naked barley, e.g. fewer pits around the margins of papilla in the silica epidermis and smaller papilla diameter (Figure 8). We thus speculate these phytoliths probably come from naked barley. Grains of naked barley found at the Loulan site (Hou, 1985) seem to support our speculation.
Although further study is needed to confirm the original species of these dendritic phytoliths, the results at least indicate that typical wheat was not a staple food for Loulan and Milan residents between ad 50 and 770. Their main crops were common millet with occasional foxtail millet and possibly naked barley. This conclusion is consistent with the records from historical literature (Hou, 1985; Xia et al., 2007).
Conclusions
Phytoliths and diatoms were retrieved from camel coprolites, fresh camel dung, an ancient tomb, and possible farmlands and soils in archaeological sites at the ancient cities of Loulan and Milan on the Silk Road. Diatom results suggest that some water bodies probably were brackish in the catchment of an ancient lake, Lop Nor. Phytolith results indicate that reeds and grasses of Paniceae and Pooideae along with some shrubs occupied the study area from ad 50 to 770. This landscape of the ancient Loulan was a typical oasis that could support human habitation. Our results also suggest that common millet was a staple crop, and foxtail millet and possibly naked barley were non-staple crops for ancient Loulan and Milan residents.
It should be pointed out that more work is needed to reveal the complete picture of paleoenvironment and agriculture of ancient Loulan and Milan because only limited samples and two types of microfossil are used in this preliminary study. Well-dated and high-resolution records of other vegetation and environmental proxy data such as pollen, plant macrofossil, and geochemical index are needed in future studies.
Footnotes
Acknowledgements
We thank Margaret Joyner and Caiming Shen from the University of Florida and State University of New York, respectively, for their work in improving the English and for valuable suggestions.
Funding
This work was supported by the National Natural Science Foundation of China (Grant No. 41102111), ‘Strategic Priority Research Program’ of the Chinese Academy of Sciences, Grant No. XDA05130401, XDA05130602, National Science Foundation of China (No. 41071131) and the National Basic Research Program of China (2010CB950201).