
Abstract
A detailed investigation of two deep-sea sediment cores from the Gardar Drift in the subpolar North Atlantic, cores MD99-2251 and MD99-2252, has been undertaken to determine the extent of Holocene climate variability reflected by changes in diatom floral abundances. Core MD99-2251 extends through the Holocene, while core MD99-2252 spans the last 7200 years. A new regional-based sea surface temperature (SST) calibration is derived using the weighted averaging partial least squares (WA-PLS) method based on 28 taxonomic categories within 53 coretops. WA-PLS-derived SST estimates indicate relatively small long-term SST changes of only 1–2°C during the Holocene. Notably, a well-defined thermal maximum is absent in the early Holocene record and the 8200-year event is not recognized as a discrete temperature minimum. However, lower SSTs between 8800 and 7800 years ago associated with the presence of sea ice and cold water species indicate an increased incursion of the subarctic water mass. A significant warming of ~2°C around 7000 years ago is the most unambiguous change in the SST records and is replicated in both cores. A brief interval of warmer sea surface conditions ~6000 years ago, associated with a Holocene maximum in North Atlantic Current indicator species, ended by 5300 years ago. Relatively muted SST variability is evident through the remainder of the Holocene.
Introduction
Diatom assemblage counts from North Atlantic deep-sea sediments have been used to reconstruct near-sea surface conditions, primarily temperature (Andersen, Koc and Moros, 2004; De Sève, 1999; Jiang et al., 2001) but also sea ice concentration (Justwan and Koç, 2008). A number of different statistical methods are employed to derive ‘transfer functions’ for use over local to regional scales in the North Atlantic. The Imbrie-Kipp (IK) method was first applied by Koç Karpuz and Schrader (1990) and has been used to reconstruct late glacial and Holocene temperature change in subsequent studies (Andersen, Koc and Moros, 2004; Koç and Jansen, 1994). More recent studies have employed techniques such as maximum-likelihood factor analysis (Witon et al., 2006), canonical correspondence analysis (Witak et al., 2005), and the weighted averaging partial least squares (WA-PLS) method (Berner et al., 2008). These studies have identified sea surface temperature (SST) as the dominant control on diatom assemblage composition.
This study explores Holocene temperature change in the subpolar North Atlantic based on diatom analyses from marine cores MD99-2251 (57°26.87′N, 27°54.47′W) and MD99-2252 (57°26.84′, 027°55.83′W) collected from the flank of the Gardar Drift (Figure 1). Unlike previous studies, our records are from within a main branch of the North Atlantic Current (NAC; Flatau et al., 2003) and hence provide a direct history of the transport of warm surface waters toward the Nordic Seas and the Irminger Current. Reconstructing past temperatures in this area is of fundamental importance because it has implications for our understanding of the large-scale meridional overturning circulation in the North Atlantic region. In this study, we use Holocene diatom census counts from core MD99-2251 (Miller et al., 2011) to reconstruct past temperature variability. In order to do this, we combine published and unpublished coretop diatom assemblage data to derive a new palaeoecological transfer function. We also apply the transfer function to unpublished diatom data from core MD99-2252, which was recovered adjacent to core MD99-2251, to provide replicated SST records over the last 7200 years. These additional records should help determine the extent to which North Atlantic diatom-based Holocene palaeotemperature records (e.g. Andersen, Koç, Jennings et al., 2004) are responding to changes in the NAC or localized climate forcing at high latitudes.
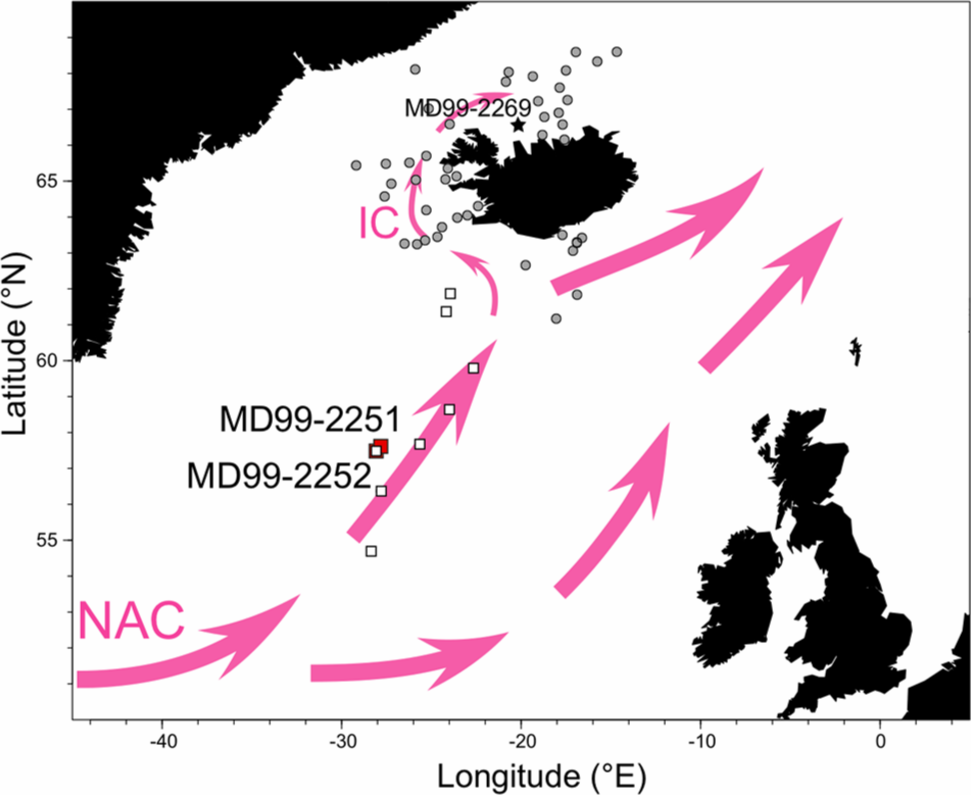
Generalized surface circulation map for the subpolar North Atlantic showing the location of cores MD99-2251 and MD99-2252 (red squares) that have been analysed to produce Holocene palaeotemperature records. NAC: North Atlantic Current; IC: Irminger Current. Also shown are the locations of the 53 coretop samples (solid circles: Jiang et al., 2001; open squares: this study) used to produce the regional SST calibration.
Methods
Core sampling and stratigraphy
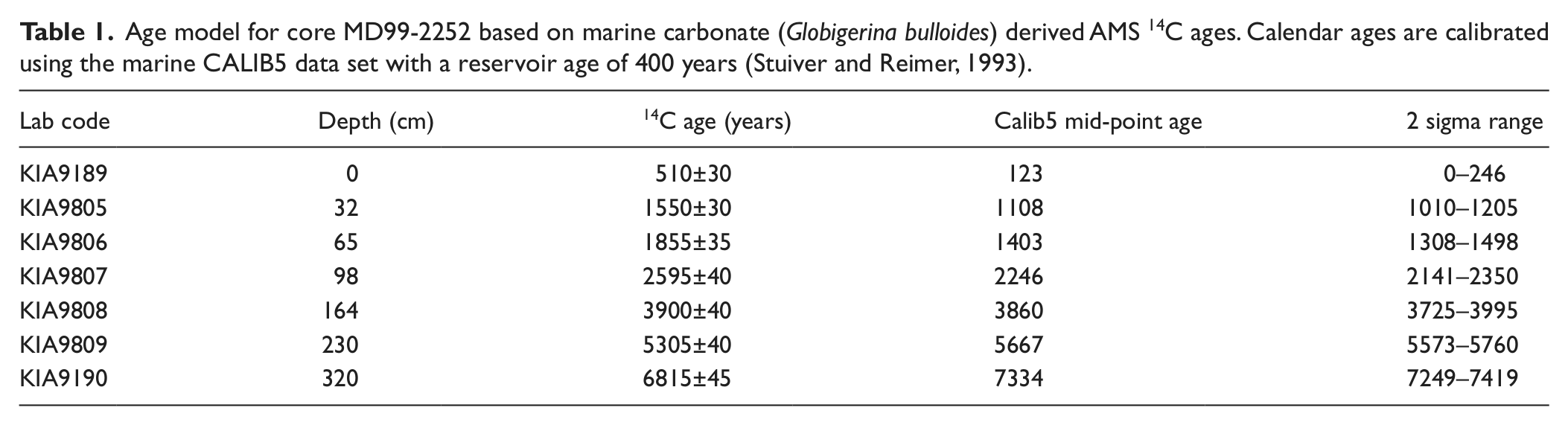
The cores investigated in this study were collected during the 1999 IMAGES V programme. Marine core MD99-2251 is a 36.6 m Calypso core. Diatoms have been analysed for the top ~16 m of sediment corresponding to the Holocene. Marine core MD99-2252 is a 3.4 m Kasten core that recovered a sedimentary record from the mid to late Holocene. The chronostratigraphy for the Holocene section of cores MD99-2251 and core MD99-2252 has been constrained using accelerator mass spectrometry (AMS) 14C dates corrected using CALIB (Stuiver and Reimer, 1993). The 23 calibrated calendrical ages used in the construction of the age model for MD99-2251 have been published previously (Ellison et al., 2006; Miller et al., 2011). Monospecific samples of Globigerina bulloides have been analysed for seven horizons in core MD99-2252; the results and mid-point calibrated ages used in the construction of the age model are shown in Table 1. Samples for diatom census counts were taken at ~250-year intervals through the Holocene for core MD99-2251 and ~180 years for core MD99-2252. Sampling density was increased to provide approximately bidecadal sample spacing around the 8200-year cooling event in core MD99-2251. For each sample more than 300 diatom frustules (excluding Chaetoceros resting spores) were counted along random transects. Further details of the sample processing procedures and taxonomy are given in Miller et al. (2011).
Age model for core MD99-2252 based on marine carbonate (Globigerina bulloides) derived AMS 14C ages. Calendar ages are calibrated using the marine CALIB5 data set with a reservoir age of 400 years (Stuiver and Reimer, 1993).
Derivation and construction of a regional diatom transfer function
Surface sample assemblage counts for seven box cores from the Iceland Basin were used to assess the performance of published transfer functions (Andersen, Koç, Jennings et al., 2004; Berner et al., 2008) at the Gardar Drift, which is not well represented in their coretop database. WA-PLS-derived SST estimates for the coretop validation samples are ~2–3°C above the modern August maximum surface ocean temperature at the respective sites (Figure 2). This discrepancy potentially poses problems for Holocene reconstructions as the scale of past temperature variability is likely to be small. The ~3°C offset of temperatures could be attributable to either the choice of statistical approach and/or limitations in the coretop calibration data set. To test this we have compiled a regional data set using assemblage data from our box coretops in the Iceland Basin, the surface sample from core MD99-2252, and 47 coretop counts from the Icelandic Shelf and northern North Atlantic collected by Jiang et al. (2001). The assemblage counts from these 53 coretops combined with summer season surface environmental data (0 m water depth) from the World Ocean Atlas (Levitus and Boyer, 1994) were used to generate transfer functions using both the Imbrie-Kipp and WA-PLS methods employing the R package Rioja (Juggins, 2009).
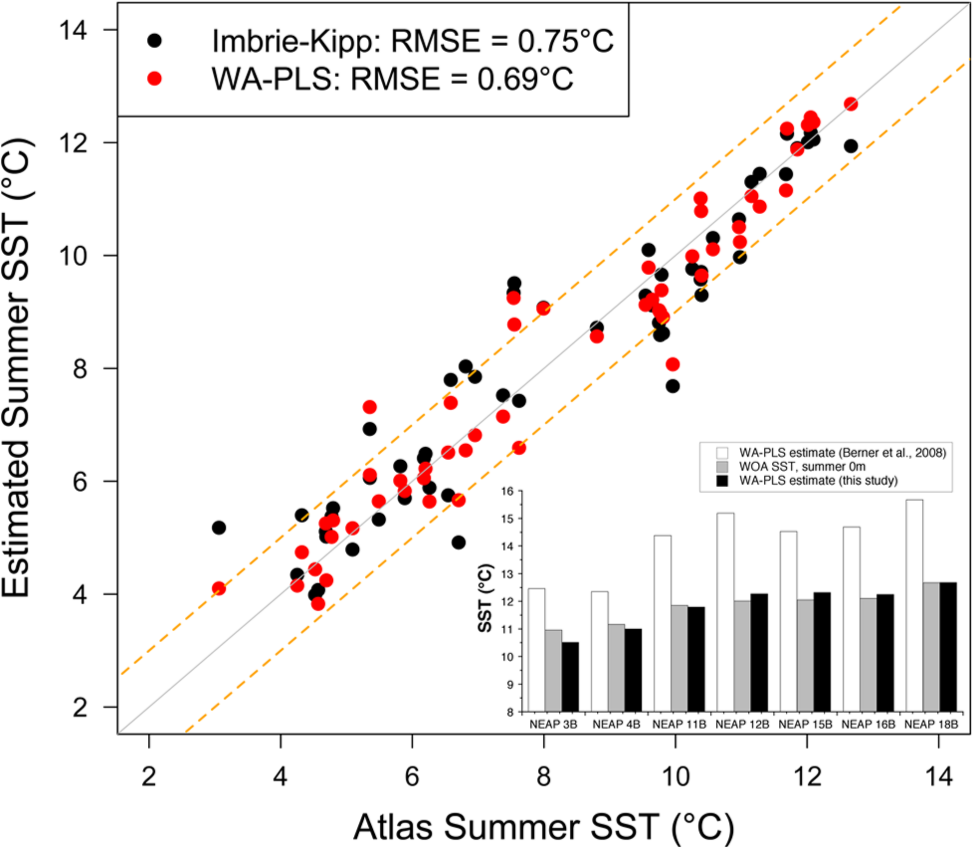
Coretop calibrations produced for the Iceland Basin calculated using the Imbrie- Kipp approach and WA-PLS technique (Juggins, 2009). Inset shows a comparison of WA-PLS-derived temperature estimates for Gardar Drift coretops produced using our calibration and the larger North Atlantic dataset of Berner et al. (2008). Atlas SSTs for the surface layer during the summer season in the World Ocean Atlas (Levitus and Boyer, 1994) are indicated.
Certain modifications were adopted in this study in order to standardize the coretop and downcore diatom datasets. Some taxonomic groupings were combined: Rhizosolenia hebetata hebetata and Rhizosolenia hebetata semispina are recorded as Rhizosolenia hebetata; Thalassiosira eccentrica and Thalassiosira spp. eccentrica group are recorded as Thalassiosira eccentrica; Rhizosolenia borealis and Rhizosolenia styliformis are combined and recorded as Rhizosolenia borealis/styliformis. The Jiang et al. (2001) study recognizes Thalassiothrix to genus level, distinguishing both Thalassiothrix sp. and Thalassiothrix? sp. These two forms, together with Thalassiothrix longissima identified at the Gardar Drift sites, were combined and recorded in the calibration data set as Thalassiothrix species. The resultant coretop database comprises 28 taxonomic categories.
Downcore diatom assemblage counts, employing the same taxonomic groupings, were used to derive palaeotemperature estimates for 173 Holocene samples. The simplified taxonomic list means that the entire diatom assemblage counts from cores MD99-2251 and MD99-2252 are not used for palaeoclimatic reconstructions. For core MD99-2251, the proportion of the diatom assemblage used ranges from 74% to 96% of the total flora (mean = 86%, standard deviation = 4.2). For core MD99-2252, the proportion of the diatom assemblage used ranges from 79% to 92% of the total flora (mean = 85%, standard deviation = 3.0). An examination of all species represented in the downcore records that are unused in the calibration dataset shows that no excluded species exceeds 5% of the total diatom flora in any sample. The omission of a key environmentally sensitive taxon therefore is unlikely to significantly impact our results.
Results
Diatom-based transfer function
The performance of the regional data set demonstrates a clear relationship between diatom assemblage composition and sea surface temperature for both the IK and WA-PLS techniques (Figure 2). The optimum configurations of the IK model (six factors, root mean square error [RMSE] = 0.75, R2 = 0.92) and WA-PLS method (two components, RMSE = 0.74, R2 = 0.92) yield equally convincing calibrations. Uncertainties estimated by leave-one-out cross-validation suggest that calibrations are accurate to ±1°C, with root mean squared error of prediction (RMSEP) values of 0.96 and 0.93 for the IK and the WA-PLS, respectively. These results compare very well to previous calibrations based on a much larger coretop data set (Andersen, Koç, Jennings et al., 2004) and also the original work of Jiang et al. (2002). The performance matches were documented in the study of Ran et al. (2011), based on a similar set of coretops but with different taxonomic groupings and environmental data, which showed a high level of consistency between reconstructed SSTs and instrumental data over the last 100 years.
The regional WA-PLS transfer function does not appear to exhibit the SST over-estimation problem evident in the larger database (Figure 2), suggesting that the elevated temperatures reflect either insufficient regional constraints within the calibration data or spatial variations in the species–environment relationships. We have used the WA-PLS approach to reconstruct Holocene summer SSTs in cores MD99-2251 and MD99-2252 on account of the slightly better calibration performance statistics. The use of WA-PLS also has been favoured in many recent palaeoenvironmental reconstructions using diatoms (e.g. Berner et al., 2011) as it can result in a reduction of the prediction error (ter Braak and Juggins, 1993). However, it should be noted that the same Holocene trends in SST are evident when the IK transfer function is applied (not shown). This high degree of consistency indicates that the calculated SST estimates are not method-specific and likely to be robust.
Holocene SST records
The range of Holocene variability in SST estimates from cores MD99-2251 and MD99-2252, derived using the regional WA-PLS transfer function, indicates only small differences from the modern temperature (Figure 3). Long-term changes in SST are only ~2°C, with the most robust features of the record being a cool phase from 9000 to 8000 years ago followed by a progressive warming from 8000 to 6000 years ago. This latter event is replicated in both cores and reflects an unambiguous change in diatom assemblage composition. It is driven by a prominent (~25%) increase in the warm-water species Thalassiosira oestrupii, whose present distribution is strongly associated with the NAC, which reaches an abundance maximum 6000 years ago (Figure 3). The cold-water indicator T. gravida vegetative also decreases significantly 7000 years ago in both cores and remains at very low abundances for the remainder of the Holocene. The overall composition of the diatom assemblages take on a modern aspect 5300 years ago, which is marked by a slight decrease in SST. Small-scale variability is evident in both records through the remainder of the Holocene where SST values, despite single-point excursions, remain close to the modern maximum temperature of 12°C (Levitus and Boyer, 1994).
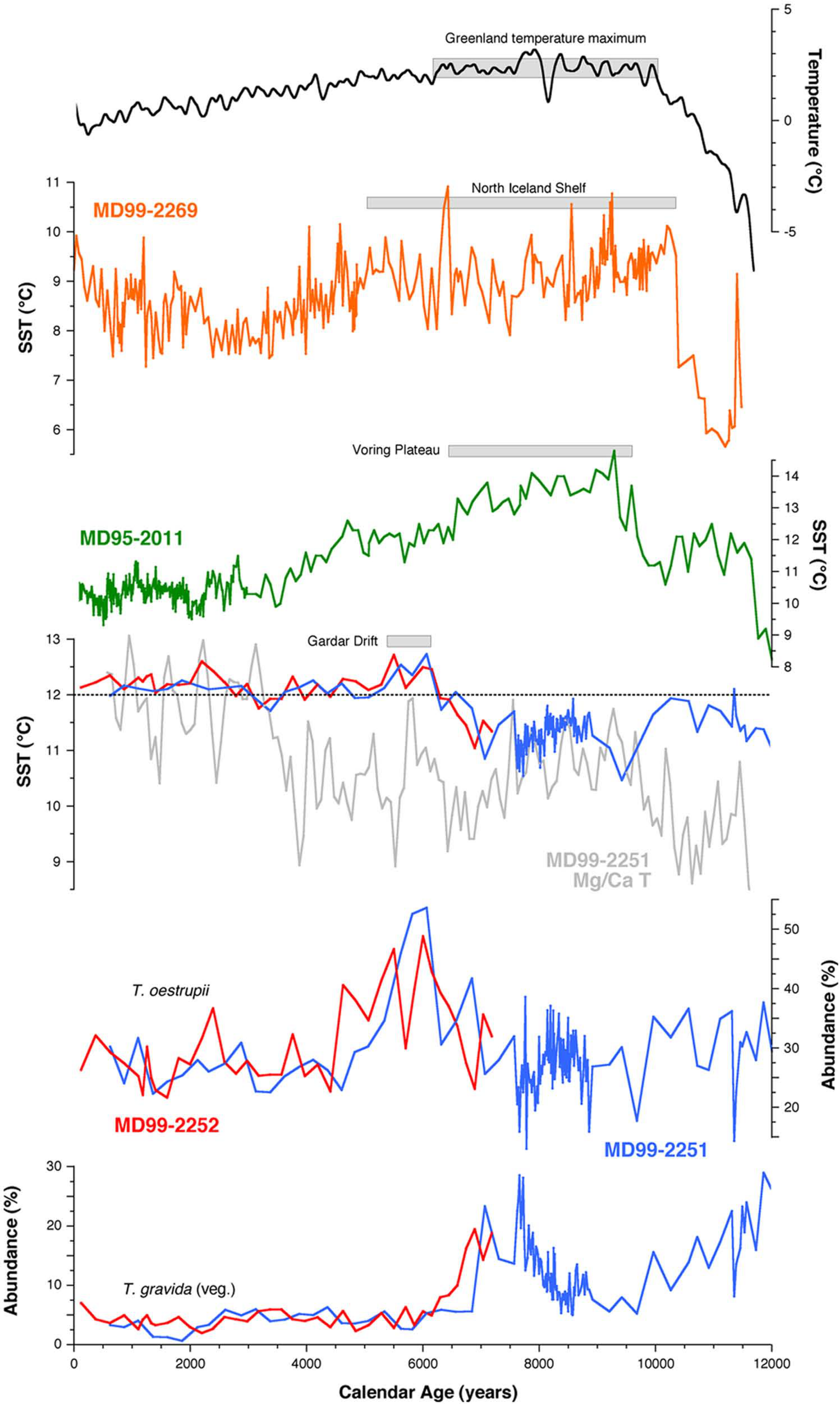
WA-PLS-generated Holocene temperature records and selected abundance data for cores MD99-2251 and MD99-2252. The Globigerina bulloides based Mg/Ca record from core MD99-2251 (Farmer et al., 2011) is plotted in grey. Present day summer sea surface temperature (SST) at the core sites is indicated by the dashed line. The diatom-based SST records (RMSEP ±1°C) and intervals of maximum SST warmth are compared with diatom-based records from the Nordic Seas (Birks and Koç, 2002) and the Icelandic Shelf (Justwan et al., 2008). Also shown is the δ18O-derived Greenland temperature history of Vinther et al. (2009) with a 200-year gaussian smoothing applied; this atmospheric record is considered to be representative of regional Holocene temperature change in and around Greenland.
One sample has been excluded from the SST reconstruction on account of the high abundances of Rhizosolenia hebetata semispina found ~9700 years ago. This assemblage is not represented in the coretop data base and cannot be interpreted robustly and most probably reflects unusual productivity conditions that existed at or near the Subarctic Front in the early Holocene (Miller et al., 2011).
Discussion
Early to late Holocene SST contrast
Previous studies have identified a well-defined thermal maximum during the early Holocene followed by a long-term cooling trend (Marchal et al., 2002). This pattern is most clearly expressed in the northern high latitudes, where regional patterns in atmospheric temperature (Vinther et al., 2009) and SST (Moros et al., 2004) are in good agreement. The diatom assemblages in core MD95-2011 (67°N, 8°E) from the Norwegian Sea illustrate the same pattern, with uniquely elevated abundances of warm water species occurring between 9200 and 6200 years ago yielding SST estimates that are 5–6°C above modern values (Berner et al., 2011; Birks and Koç, 2002). By contrast the warm floral assemblages have similar abundances across the early and late Holocene intervals in core MD99-2251, indicating that there was no pronounced early Holocene thermal maximum in the subpolar ocean. In fact the diatom SST estimates from MD99-2251, which are low and reach a minimum during the early Holocene (Figure 3), differ markedly from the pattern identified in the Nordic Seas in terms of the magnitude of temperature change and the long-term trend. The absence of a clear early Holocene thermal maximum in MD99-2251 is supported by the near-surface Mg/Ca ratios measured on Globigerina bulloides in the same core (Farmer et al., 2008, 2011), although differences exist in the timing and scale of abrupt climate events. The diatom-based SSTs indicate warmer conditions than the Mg/Ca temperatures in the early and mid Holocene but converge through the cool period between 7000 and 9000 years ago. In the late Holocene the Mg/Ca record exhibits greater variability than the diatom SSTs but both indicate warmer temperatures than in the early to mid-Holocene.
Reconstructed SST values through the early Holocene deviate from modern values by only ~1°C (close to the error associated with the WA-PLS method) but the actual composition of the diatom assemblage differs significantly. Elevated abundances of T. gravida vegetative occurred between 9500 and 7000 years ago (Figure 3). There are also minor increases in other cold-adapted taxa within this interval (Miller et al., 2011). T. gravida vegetative is indicative of cold surface conditions and strongly associated with Arctic diatom assemblages (Andersen, Koc and Moros, 2004), and in the North Atlantic is found in a belt following the Subarctic Front. The early Holocene diatom record from core DS97-2P, situated nearby on the Reykjanes Ridge (59°N, 30°W), is dominated by T. gravida vegetative, R. hebetata and R. styliformis (Witak et al., 2005). Such a well-developed cold flora is not evident in any of the core MD99-2251 samples, but the mixed cool-warm aspect of the floras found in core MD99-2251 are consistent with the Subarctic Front being situated close to the mid-Atlantic Ridge and possibly extended into the Iceland Basin where it mixed with the waters of the NAC. Although the interval of lower SSTs identified in our record can be interpreted as a period of subtle changes in the flora that reflect a relatively continuous advection of warm NAC flow over the site and periodic eastward incursion of colder waters originating from the East Greenland Current (EGC), it also may be explained by seasonal changes in surface temperatures and water masses given the likely proximity of the Subarctic Front to the site.
Diatom-derived palaeotemperature records from the western side of the Reykjanes Ridge (Andersen, Koc and Moros, 2004; Berner et al., 2008) indicate a relatively cool and highly variable early Holocene with two significant cooling periods from 10,000–9400 and 8000–7000 years ago. The WA-PLS-generated SSTs for MD99-2251 identify significant decreases in SST around 9400 and 7700 years ago. These may be the same events recorded in core LO09-14 given the uncertainties associated with chronology and the different sampling density of the cores. Dinocyst assemblage-derived summer SSTs for LO09-14 indicate significantly higher temperatures (14–17°C) around 9000–8000 years ago, compared with diatom estimates through this interval and modern summer maximum values (Solignac et al., 2008). However, dinocyst-based salinity variations, which suggest an incursion of fresher waters from the EGC in the early Holocene, exhibit a very similar pattern to the diatom SSTs on Gardar Drift. In summary, reconstructions from the subpolar ocean clearly provide evidence of a different SST history for the Holocene, irrespective of location or the specific configuration of the transfer function. The cooling pattern in mid-Atlantic diatom records suggests that Arctic waters transported via the EGC extended across much of the Irminger Sea until ~7000 years ago.
Mid-Holocene warming and the 5300 year event
A ~2°C warming commencing 7000 years ago culminated in a relatively short interval of maximum SSTs in both cores MD99-2251 and MD99-2252 at ~6000 years ago. This episode appears to have a widespread signature as it correlates with the mid-Holocene SST optimum in core LO09-14 on the Reykjanes Ridge (Andersen, Koç, Jennings et al., 2004) and it has a clear counterpart in the diatom abundance records of core DS97-2P (Witak et al., 2005). The very strong similarities between the records imply that this change represents an enhancement of the NAC/Irminger Current system of regional significance following a significant westward adjustment in the position of the Subarctic Front. An abrupt change in the hydrology of the North Iceland Shelf was identified by Ran et al. (2008) at 5300 years BP, characterized by a dramatic increase of Thalassionema nitzschioides, the predominant species of the Norwegian-Atlantic Current assemblage in the Nordic Seas, at the expense of the warm-indicator T. oestrupii, and an expansion of sea ice species associated with the EGC. This change is coincident, within dating error, with the cooling from Holocene maximum SSTs that occurred ~5300 years ago in cores MD99-2251 and MD99-2252 and the onset of more stable late Holocene conditions. The cooling in our records is relatively limited and differs from the early Holocene interval as the diatom assemblages do not reflect a switch to conditions strongly influenced by the EGC. Changes in diatom assemblages near the Faeroe Islands also suggest a reduction in the influence of the NAC and an increase in cold water influence at ~5400 years ago (Witak et al., 2005; Witon et al., 2006).
Magny and Haas (2004) have reviewed the evidence for a mid-Holocene reorganization and the onset of cooler conditions between 5600 and 5000 yr BP. In addition to their study of lake level and vegetation data from Lake Constance, they cite support for a widespread event at this time based on, amongst others, cooler Greenland ice core temperatures, a retreating timberline in Canadian high latitudes, and cooler SSTs. The diatom records from the subpolar region add further support for this being a transitional event of some significance within the Holocene. Interestingly, the sortable-silt record from MD99-2251 indicates that there was a substantial reduction in deep ocean flow speed between 5600 and 4800 years ago, with a pronounced minimum centred at 5100 years (Hoogakker et al., 2011). This correlation is significant because a reduction or weakening in the meridional overturning circulation has been suggested as a possible cause and/or mechanism for propagating millennial-scale events. In this instance, the reorganization at 5300 years appears to postdate a major change in the Holocene deep ocean circulation ~6500 years ago (Hoogakker et al., 2011), but could reflect a wider change or threshold response, such as an adjustment in the position of frontal systems in the surface circulation of the subpolar North Atlantic.
Wider correlation to diatom SST records
Diatom-derived palaeotemperature records from the North Iceland Shelf (Andersen, Koç, Jennings et al., 2004; Ran et al., 2008) and Vøring Plateau (Berner et al., 2011; Birks and Koç, 2002) indicate the establishment of warmer SSTs earlier in the Holocene followed by the onset of a general cooling trend at ~7000 years (Figure 3). To a large extent, the SST patterns observed to the south in the Iceland Basin and subpolar North Atlantic are the opposite. The exact duration of maximum Holocene warmth varies geographically, from 10,400 to 4700 years on the North Icelandic shelf (Justwan et al., 2008) to 6000 to 5300 years in our records from the Iceland Basin. The presence of distinct regional variation in diatom-generated SSTs cautions against a simple regional interpretation in terms of a single causal mechanism. One implication of our results is that maximum summer SSTs appear to have been 3–4°C warmer in the Norwegian Current than in core MD99-2251 for a large part of the early Holocene. We therefore find no evidence for a stronger/warmer NAC in the Iceland Basin as this time. The proximity of the Subarctic Front, which resulted in the combination of NAC and Arctic diatom floras, is a likely explanation for the lower SSTs. It is possible that the warm core of the NAC was positioned further to the east in the early Holocene; however, available low resolution records suggest that there were low SST gradients between the North Atlantic and Norwegian Sea over this interval (Koç et al., 1996). This supports the idea that insolation forcing is a major control of Holocene SST evolution in the Norwegian Sea, but there are complicating issues that impact even this interpretation when the records from adjacent cores are compared in detail (Birks and Koç, 2002). This complexity is further highlighted by the coincidence of the most notable feature of our SST records, the warming trend ~6500 ago, and the onset of cooling and the trend toward neoglaciation at the Vøring Plateau. The time transgressive nature of changes in Holocene diatom assemblages largely reflects the sensitivity of the core locations to readjustments and migration of local frontal systems.
Conclusions
Holocene variations in diatom assemblage composition for cores MD99-2251 and MD99-2252 from the Gardar Drift in the subpolar North Atlantic have been quantified and used to produce a regional SST history with a new regional-based temperature calibration calculated using the WA-PLS method. WA-PLS-derived SST estimates indicate long-term changes of only 1–2°C during the Holocene, but these events are marked by unambiguous changes within the diatom community (Miller et al., 2011). Significant SST changes in the subpolar North Atlantic occurred in the 12,000 to 5000 years time window. Any subsequent variability is difficult to quantify robustly given the temporal resolution of the data. Notably, a well-defined thermal maximum is absent in the early Holocene record and the 8200-year event is not recognized as a discrete SST minimum. However, SST cooling between 8800 and 7800 years ago reflects the nearby presence of the Subarctic Front and an increased incursion of Arctic waters. The mid Holocene was a time of significant changes in surface conditions. This is marked by a significant SST warming trend ~6500 ago, followed by a brief thermal maximum that was over by ~5300 years ago, when assemblages similar to the modern became established. Our records provide limited evidence of SST variability through the late Holocene.
Footnotes
Acknowledgements
We acknowledge the support of Nalan Koç and participants of the 1999 IMAGES cruise of the Marion Dufresne. We thank Nick McCave for the provision of NEAPACC box core samples and Hui Jiang and Carin Andersson for diatom data. We also thank two anonymous reviewers for their suggestions to improve the manuscript.