
Abstract
Transhumance agriculture formed a key component of subsistence strategies in the Norse economies of the North Atlantic, with evidence of shielings or sæters found in Norway, Scotland, the Faroe Islands and Iceland. It is frequently assumed to have played a role in Norse Greenland, yet little enquiry has been made into such activity. This paper seeks to address this deficit, presenting the first palaeoenvironmental study of a suspected Greenlandic shieling site in the uplands of the former Norse Eastern Settlement. Pollen analysis, 14C and associated proxies are used to date and assess the environmental and landscape impact of shieling activity. Evidence for vegetation disturbance associated with Norse settlement is indicated from c.
Introduction
The Norse expansion across the North Atlantic Ocean began in the early 9th century
The Norse established a pastoral agricultural system in what can be considered a ‘pristine’ landscape (Dugmore et al., 2005) with the primary unit of settlement being the farm (Buckland, 2008; Kaland and Martens, 2000). Hay was grown during the summer months for the overwintering of their livestock through the severe Greenlandic winters (Berglund, 1986). This was supplemented with the hunting of wild game, seabirds and marine mammals (McGovern, 1985). These aspects of the Norse subsistence system have been discussed and studied extensively (e.g. Edwards et al., 2008; McGovern, 1985; Schofield et al., 2008), yet there has been only limited discussion of the more peripheral elements of Norse farming such as transhumance (see Albrethsen and Keller, 1986). Known to have been practiced in the wider North Atlantic and Norway (Lucas, 2008; Magnus, 1986; Mahler, 2007; Reinton, 1969; Skrede, 2005; Sveinbjarnardóttir et al., 2011), transhumance is a system whereby farmers move animals during summer, exploiting more remote pastures while at the same time reducing the pressure on pastures close to the main farm (Albrethsen and Keller, 1986). Such activity is characterized by small isolated buildings (shielings or sæters) often at altitude, but relatively near to a main farm (Albrethsen and Keller, 1986; Reinton, 1969). These structures are common throughout southern Greenland (Albrethsen and Keller, 1986; Guldager, 2002), suggesting that transhumance formed an important part of Norse subsistence strategies.
This paper presents data from a presumed shieling – Norse ruin group Ø70 (Bruun, 1896), widely referred to as the ‘Mountain Farm’ (Vebæk, 1992) – in the uplands of Vatnahverfi (‘land of lakes/waters’; Ingstad, 1966) in southern Greenland, and aims to address the deficit of knowledge relating to transhumance activity in Norse Greenland. Primarily the paper seeks to establish how land use developed at this site, and from this evidence to establish whether the Mountain Farm was utilised as a shieling. To date, no such site from Greenland has been examined in this manner, whereas in Iceland, the Faroes and Norway, pollen-analytical data have furnished important insights into the practice of transhumance in the Norse cultural sphere (Borthwick et al., 2006; Kvamme, 1988; Sveinbjarnardóttir et al., 2011).
The study site
Ruin group Ø70 comprises eight relatively small structures located in the Vatnahverfi region of the Eastern Settlement. Named ‘The Mountain Farm’ by Christian Vebæk (1992) because of its location at 260 m a.s.l. (Figure 1) the site was first recorded by Daniel Bruun (1896). The nearest modern settlements are the lower-lying sheep farms of Timerliit 1 km to the northwest, and Igaliku Kujalleq 5 km to the north. Nearby Norse structures are ruin groups Ø69 (located at Timerliit) and Ø167, a large farm complex approximately 3 km to the southwest. The ruins at Ø70 are situated on a low hill overlooking two lakes fringed with Carex rostrata reedswamp and surrounded by Sphagnum-Cyperaceae mire (plant nomenclature follows Böcher et al., 1968). The vegetation in the wider vicinity of the ruins comprises extensive upland grassland with patches of Salix glauca scrub.
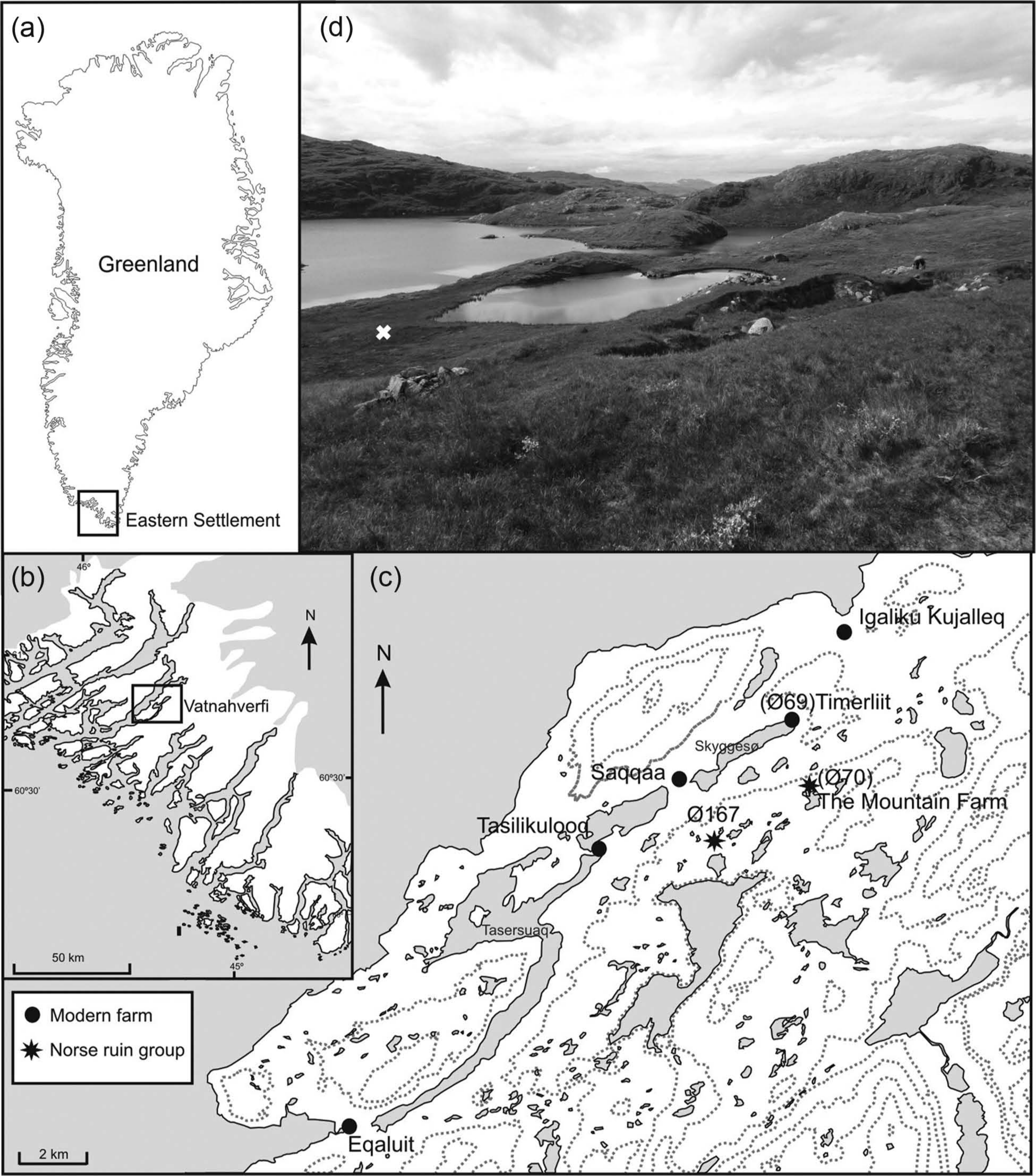
Location maps: (a) the Norse Eastern Settlement in Greenland; (b) Vatnahverfi within the Eastern Settlement; (c) the Mountain Farm and other sites mentioned in the text; (d) photograph looking southwest across the site. The ruin group can be seen in the foreground and the white cross denotes the sampling location.
Archaeology
Limited excavation of the ruin group was undertaken by Vebæk in 1950. This yielded a surprisingly large quantity and variety of finds relative to the site’s size and presumed function as a shieling (Vebæk, 1992). A total of 76 artefacts were recorded, amost which was a semi-manufactured iron bar and iron slag. This perhaps indicates that iron working was a supplementary activity practised at the site.
Physical geography
The study area is characterized by rounded mountain topography ranging from sea level to 600 m a.s.l. and interspersed with numerous lakes. Local geology is uncomplicated and dominated by a suite of granites (Allaart, 1976). The surficial drift cover comprises glacial and glaciofluvial deposits of Quaternary age (Feilberg, 1984), which are overlain in places by aeolian sediment sourced from the sandur of the Jespersens Dø glacier 15 km to the northeast (Jakobsen, 1991). The dominant soils are podzols. Climatically the site straddles the boundary of the subcontinental, subarctic and suboceanic zones of southern Greenland (Feilberg, 1984). The nearest observational climate data comes from Narsarsuaq (35 km to the north) which experiences mean summer (July) temperatures and annual precipitation of 9.8°C and 696 mm respectively (Feilberg, 1984).
Methodology
Fieldwork and sediment sampling
Fieldwork was undertaken in July 2010. Shallow peat deposits were identified at a small (110 m × 46 m) Sphagnum-Cyperaceae mire approximately 50 m south of ruin group Ø70 (60°50.762′N, 45°17.478′W; Figure 1). The total depth of organic deposits was 155 cm. This was sampled and analysed between 60 and 110 cm using an 8 cm diameter Russian corer (Jowsey, 1966) and the location was recorded using a portable Garmin GPS handset. The core section was protected in plastic guttering and wrapped in polythene before being returned to the University of Aberdeen where it was placed in cold storage (4°C). Subsampling and Troels-Smith (1955) recording of the lithostratigraphy were undertaken in the laboratory. Loss-on-ignition (LOI) was measured following 3 h combustion at 550°C (Bengtsson and Enell, 1986).
Pollen analysis
Samples for pollen analysis were prepared using standard NaOH, sieving, acetolysis and floatation (solution density 1.88 g/ml) procedures (Moore et al., 1991; Nakagawa et al., 1998). Lycopodium tablets were added to each sample to allow the calculation of concentration and influx data (Stockmarr, 1971). Samples were suspended in silicone oil, mounted on slides and examined using a Nikon E400 light microscope at ×400 magnification and counted until a sum of 500 total land pollen (TLP) had been achieved.
Pollen and spores were identified with the aid of Moore et al. (1991) and reference material held at the University of Aberdeen. Nomenclature follows Bennett et al. (1994) and Bennett (2012a). Taxa absent from these lists follow the nomenclature in Moore et al. (1991). The separation of Betula pollen into tree (Betula pubescens) and shrub (Betula glandulosa) varieties was undertaken using grain size diameter measurements (cf. Fredskild, 1973; Mäkelä and Hyvärinen, 1998; Schofield and Edwards, 2011). Where diameter was <20 µm, Betula was classified as B. glandulosa. Cereal-type pollen grains were identified and categorised following Andersen (1978). Coprophilous fungal spores were identified using van Geel et al. (2003). These spores are derived from fungi that grow on animal dung and have been demonstrated to increase at landnám following the introduction of grazing herbivores (Schofield and Edwards, 2011). Care must be taken in the interpretation of this proxy as small numbers have also been shown to be present both before and after Norse occupation (Gauthier et al., 2010). Pollen diagrams were constructed using TILIA and TGView software (Grimm, 1993, 2012) with percentages based upon the TLP sum.
Charcoal analysis
The area covered by microscopic charcoal in pollen residues was measured using a microscope eyepiece graticule at a magnification of ×400. Owing to the frequently high concentrations of charcoal, only the first 50 fragments larger than 50 µm2 were included in the analysis (cf. Edwards et al., 2008; Schofield et al., 2008). Between 93 and 83 cm where charcoal was particularly abundant, the point count method was employed (Clark, 1982). 8800 points were applied to achieve a <5% error in charcoal estimates. Techniques employed for determining pollen concentrations were used to calculate charcoal influx (cf. Swain, 1978). Additionally, charcoal to pollen (C:P) ratios were calculated to check whether charcoal abundances were changing as a consequence of sedimentation effects (Patterson et al., 1987).
Numerical analysis
CONISS (Grimm, 1987) was used to assist with the biostratigraphic zonation of the pollen diagram following square root transformation of percentage data. Rarefaction analysis (Birks and Line, 1992) was performed in psimpoll (Bennett, 2012b) and plotted using TILIA. Principal components analysis (PCA (Figure 2)) was undertaken using CANOCO 4.5 (ter Braak and Šmilauer, 2002) after detrended correspondence analysis (DCA) indicated a linear rather than unimodal pattern to the data set (Lepš and Šmilauer, 2003).

(a) PCA of pollen types and associated proxies from the Mountain Farm (selected taxa only). Key to taxon labels: Api: Apiaceae; BetG: Betula glandulosa; BetP: Betula pubescens; Brass: Brassicaceae; Cary: Caryophyllaceae; Cer: Cerastium-type; C:P: charcoal to pollen ratio; Cyp: Cyperaceae; Emp: Empetrum nigrum; Eq: Equisetum; Gal: Galium-type; Gent: Gentianaceae; Hor: Hordeum-type; Lac: Lactuceae; Lyco: Lycopodium annotinum; Poa: Poaceae; Pot: Potentilla-type; Ptero: Pteropsida monolete indet.; Ranu: Ranunculus acris-type; Rose: Roseaceae; Rsella: Rumex acetosella; Rtosa: Rumex acetosa; Sal: Salix; SaxO: Saxifraga oppositifolia-type; Spha: Sphagnum; T55a: Sordaria-type; T113: Sporormiella-type; Unk: unknown and indeterminable; Vacc:Vaccinium-type. (b) PCA plot of sample scores for the Mountain Farm pollen and spore data set.
Radiocarbon dating
Five plant macrofossil samples (Table 1) were submitted for AMS 14C dating. Sediment samples were gently disaggregated in weak NaOH and washed through a 125 µm sieve using distilled water. Residues were examined under a low power binocular microscope and plant macrofossils picked for dating. Seeds were identified using Cappers et al. (2006) and modern reference material held at the University of Aberdeen. Measurements were undertaken on terrestrial plant macrofossils where possible, with the humic acid fraction of small (1 cm3) peat samples only used where suitable macrofossils for dating were absent. AMS samples were processed and measured at SUERC, East Kilbride, and calibration was undertaken using the IntCal09 calibration curve (Reimer et al., 2009) and CALIB v6.0. Two approaches were explored to generate a chronology: Bacon, which employs a Bayesian methodology (Blaauw and Christen, 2011) and Clam, which offers a ‘classical’ approach (Blaauw, 2010).
Radiocarbon dates for the Mountain Farm profile.

Results
Chronology
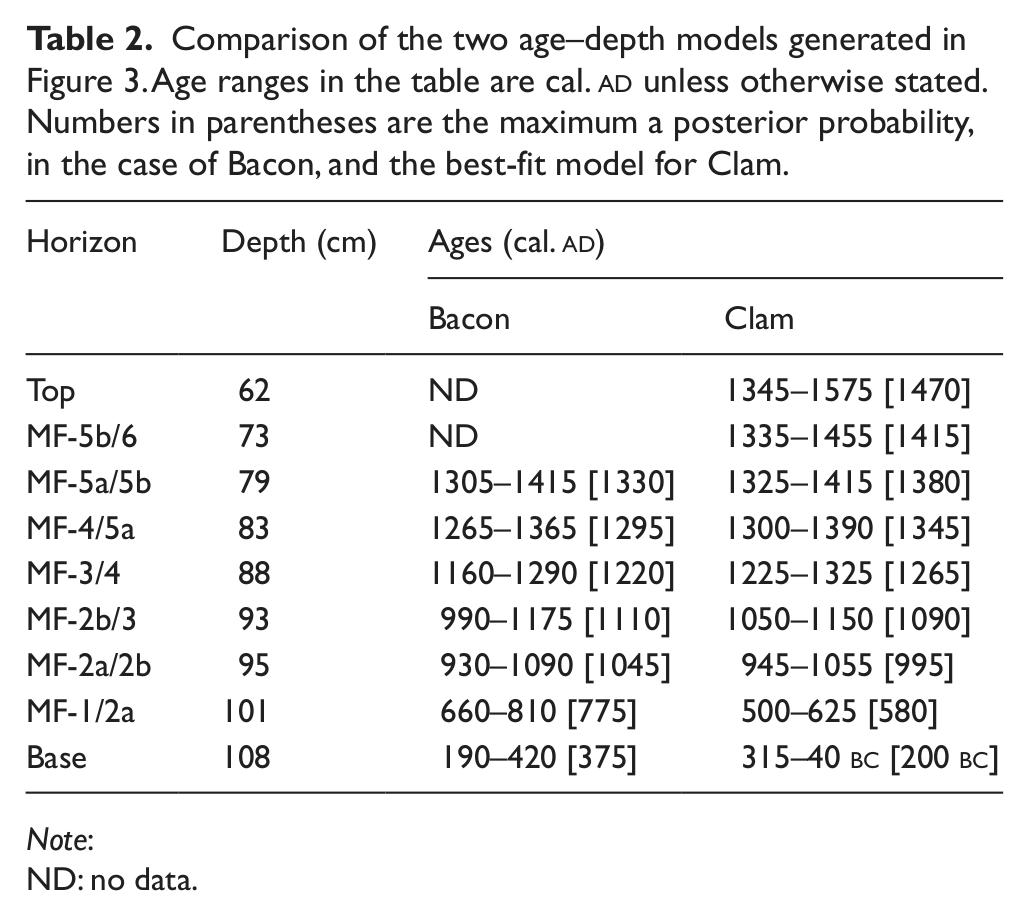
The results of 14C dating are presented in Table 1. All the dates, except the uppermost, form a clear stratigraphic sequence with no age inversions. The date at 64–61 cm (SUERC-33441) on a Betula twig provided a modern radiocarbon age and is unquestionably not in situ. The most likely explanation is that this fossil was moved down the profile during sampling and it has therefore been omitted from model runs. The two age–depth models are presented in Figure 3 and the 2σ age ranges for zone boundaries and the base and top of the analysed sequence are compared in Table 2. The two models compare favourably and there is much overlap in the 2σ age envelopes (Table 2). The main differences are that Bacon provides: (1) a model with more conservative confidence intervals; (2) marginally older dates for events postdating the 12th century

Age–depth models for the Mountain Farm profile generated using Bacon (Blaauw and Christen, 2011) and Clam (Blaauw, 2010) software. For Bacon, the mean deposition time was set at 30 yr/cm and memory strength and mean at 50 and 0.8, respectively to allow a high degree of flexibility in the accumulation curve. The shaded area in the model indicates all possible age–depth models and the dotted line reflects the 2σ age range; darker areas indicate increased certainty. Shapes show calibrated calendar age distributions of individual dates. For Clam, a smoothed spline was fitted through the dates and extrapolated from 73 cm to the top of the pollen analysed sequence at 61 cm. The black line indicates the best-fit model and the grey shaded regions indicate the 2σ age range.
Comparison of the two age–depth models generated in Figure 3. Age ranges in the table are cal.
Note:
ND: no data.
Lithostratigraphy
The mire stratigraphy is broadly homogeneous through the analysed sequence and comprised a brown to dark brown sandy and humified peat. Humified Sphagnum remains were abundant between 96 and 93 cm. Sandy peat continued below the analysed sequence, grading into sand at 155 cm. Above the analysed sequence the sandy peat continued to 34 cm where there was a sharp contact onto poorly humified Sphagnum peat. LOI is low throughout at ≤ 40%. The core stratigraphy is presented alongside pollen-analytical data in Figure 4.

Percentage pollen and spore diagram for the Mountain Farm displaying selected taxa (minimum sum = 500 TLP). Also shown are the modelled calendar ages, modelled 2-sigma calendar range, uncalibrated 14C dates, lithology, loss-on-ignition, microscopic charcoal, pollen concentration and the rarefaction index . + indicates < 1% TLP.
Palynology
The environmental baseline (LPAZs MF-1)
Local pollen assemblage zone (LPAZ) MF-1 opens cal. 315–40
The pre-landnám environment (LPAZ MF-2a) and regional landnám (LPAZ MF-2b)
The MF-1/2a zone boundary dates to c. cal.
Similar palynological changes to those witnessed in MF-2a have been noted before landnám in the Faroe Islands where an expansion of herbaceous types was interpreted as potentially representative of an early farm (Hannon and Bradshaw, 2000). Such an explanation seems unlikely at the Mountain Farm given the lack of a full suite of Norse palynological indicators (Edwards et al., 2011a) and an absence of microscopic charcoal.
It seems more plausible that development in LPAZ MF-2a relate to grazing by wild herbivores such as caribou (reindeer; Rangifer tarandus), which were previously common across the region (Ingstad, 1966). The appearance and steady increase of spores from coprophilous fungi (Sporormiella-type and Sordaria-type) support this view. The situation may have been similar to that around Kangerlussuaq, western Greenland (67°0′N, 50°41′W). Here sustained grazing by caribou has led to the creation of patches of grassland (or ‘greens’) with a near-uniform cover of non-flowering Poa pratensis (Fredskild and Holt, 1993).
Subtle changes separate LPAZ MF-2a from MF-2b, the onset of which dates to cal.
Local landnám (LPAZ MF-3)
If LPAZ MF-2b represents a ‘regional’ landnám, then the opening of MF-3 (dating to cal.
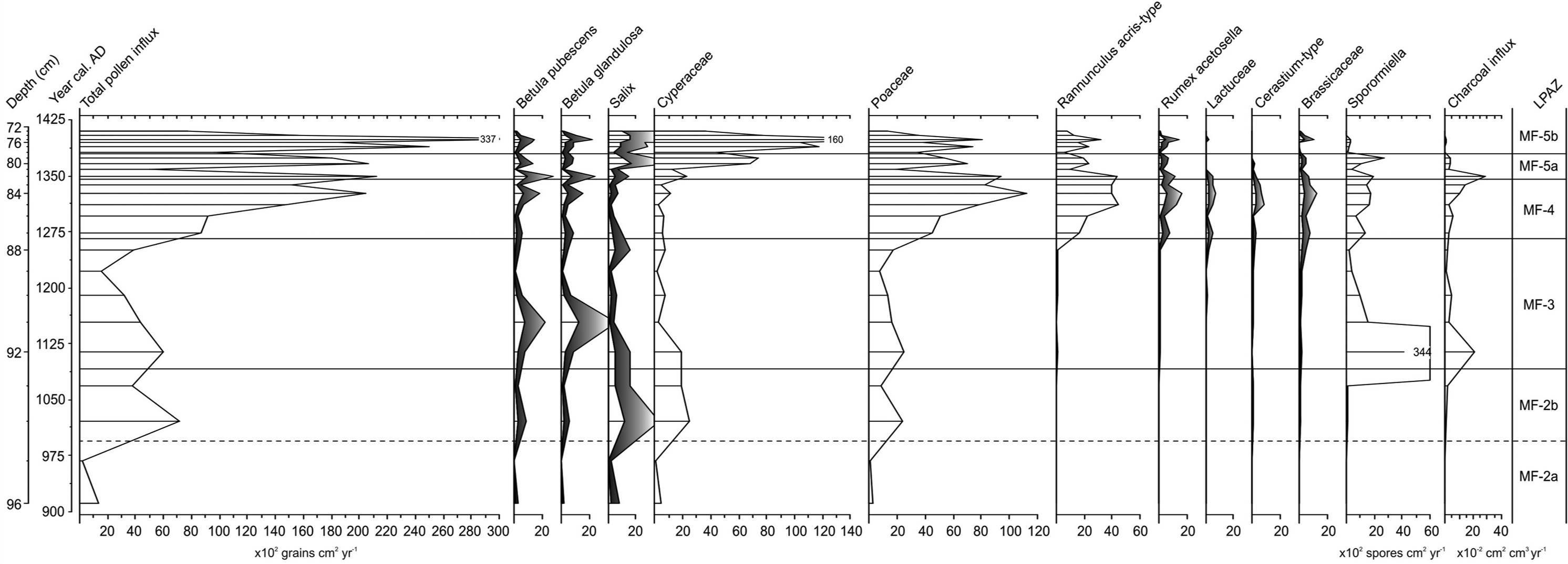
Influx diagram displaying selected pollen types, spores and charcoal (exaggeration curves where present are ×5).
The continued rise in Poaceae to c. 40–45%, alongside a general decline in Cyperaceae from c. 35% to 20% and fluctuations in Salix and Betula, suggest the expansion of grassland pasture and perhaps some hayfields at the expense of mire and dwarf-shrub heath. Aside from these broader-scale changes in the pollen assemblage, an increasing rarefaction index, rising to a maximum value of 26, apparently confirms a diversification in the habitats within the pollen catchment area. The expansion of pollen from ruderal plants such as Brassicaceae (cabbage family; c. 1–4%), possibly representing Capsella bursa-pastoris (shepherd’s purse) (cf. Fredskild, 1978), Polygonum aviculare (knotgrass; trace) and Lactuceae (up to 1%) probably indicates disturbed ground around buildings and areas of habitation (Schofield et al., 2007). Cerastium-type pollen (c. 1%) conceivably reflects Stellaria media (common chickweed) (Fredskild, 1978) growing in a similar habitat, or one of a number of herbs from the same genus growing in pasture with Ranunculus acris-type (meadow buttercup; c. 2–8%). Rumex acetosella (c. 1–3%) points to an expansion of grazed heaths, and an abundance of coprophilous spores (Sporormiella-type) at the opening of MF-3, with consistently high values thereafter (c. 30–40%), may imply high stocking densities and heavy grazing (cf. Graf and Chmura, 2006).
A major decline in LOI from c. 35% to 20% and a sharp peak in undifferentiated fern spores (c. 4%) in the middle of the zone suggest widespread landscape disturbance linked to grazing. Erosion of catchment soils and a consequent inwash of secondary pollen may also account for the anomalously high Betula pollen counts (c. 45%) near the opening of the zone (cf. Gathorne-Hardy et al., 2009). Over 75% of the Betula grains in this sample are damaged compared with c. 20–35% of the grains in samples immediately above and below, suggesting some bias within the assemblage. MF-3 seems to reflect a mosaic of heavily grazed natural vegetation communities including mire, dwarf-shrub heath and grassland, along with the emergence of culturally related pasture and ruderal communities.
Intensification (LPAZ MF-4)
The opening of LPAZ MF-4 (cal.
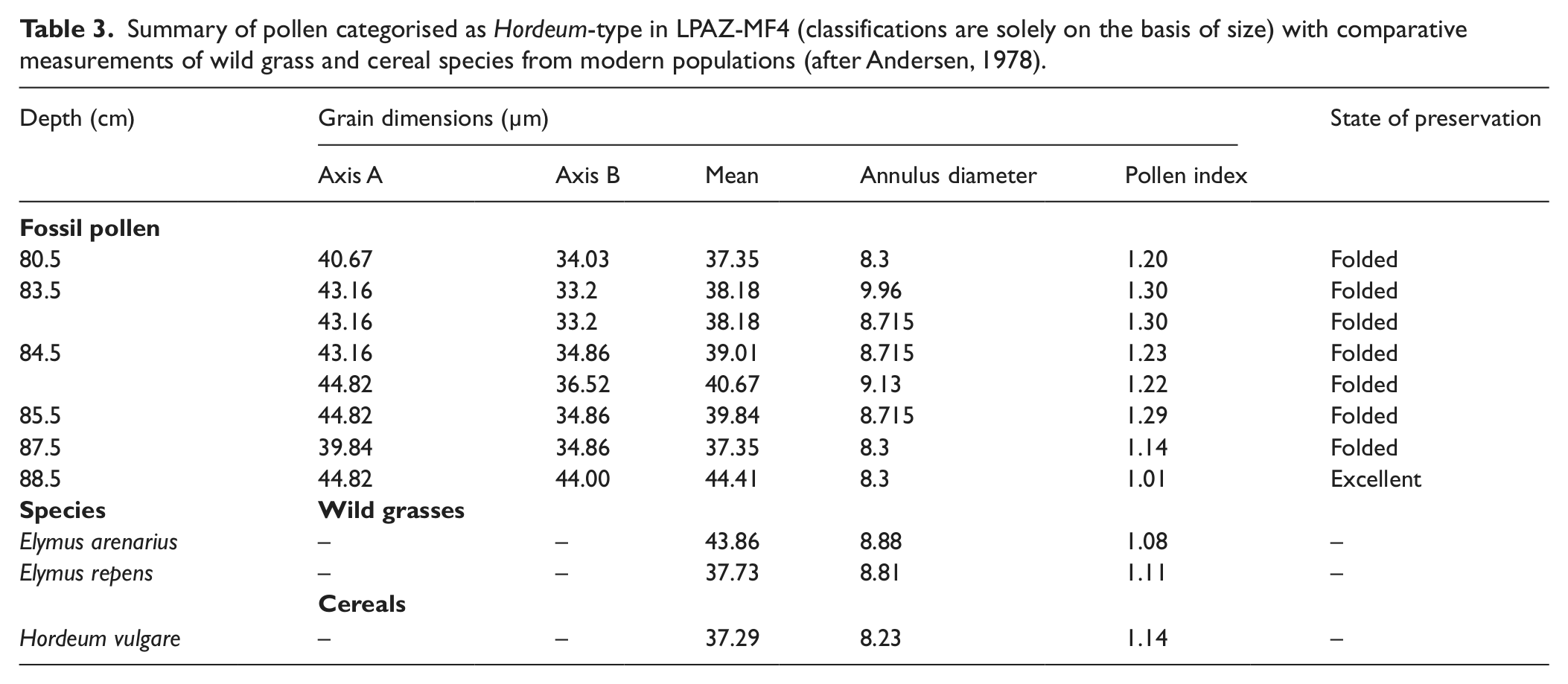
A further development is the appearance of Poaceae in the Hordeum-type size range (Andersen, 1978). Whether these large pollen grains (Table 3) reflect actual attempts to grow barley is uncertain. On the basis of annuli and pore measurements recommended by Tweddle et al. (2005), it seems more likely that they reflect Elymus arenarius (lyme grass), which generally grows on sandy or stony ground close to the coast and in river valleys (Böcher et al., 1968). Although neither of these environments is widely represented in the pollen catchment of the Mountain Farm today, it is possible that disturbance from grazing created patches of sandy ground suitable fro the plant to invade. Alternatively, Elymus pollen may have blown into the deposit from elsewhere in Vatnahverfi, such as from the sandur area to the north. There are clear indications that the pollen assemblage from LPAZ MF-4 records a significant shift in land use. PCA (Figure 2) shows samples from MF-4 plotting in a distinctly different ordination space to those from MF-3. Given the consistently high Poaceae pollen percentages (c. 50–55%) in MF-4, it is possible that this shift reflects the initiation, or a significant expansion, of seasonal hay growing at the site. In a modern study of pollen vegetation relationships in southern Greenland, Schofield et al. (2007) recorded Poaceae values ranging from 37% to 96% in the margins of hayfields. Considering that modern farms benefit from the use of pesticides and industrial fertilizers, the values of 50–55% in MF-4 could be viewed as abundant. Influx data (Figure 5) also points towards a significant increase in the productivity of vegetation communities with both Poaceae and Ranunculus acris-type rising sharply. Although warmer climate shifts can be responsible for increased pollen influx, such an explanation seems unlikely in this case. Influx values change little amongst the other taxa, implicating the influence of a factor other than climate.
Summary of pollen categorised as Hordeum-type in LPAZ-MF4 (classifications are solely on the basis of size) with comparative measurements of wild grass and cereal species from modern populations (after Andersen, 1978).
Abandonment (LPAZs MF-5a and b)
A reduction of human activity is recorded in MF-5a (beginning cal.
The opening of LPAZ subzone MF-5a appears to reflect the abandonment of hayfields, although use of the site continued, perhaps on a seasonal basis only, as microfossil signatures for grazing and burning are still evident. The opening of subzone MF-5a (cal.
The post-Norse landscape (LPAZ MF-6)
A large increase in Salix pollen (c. 20–25%) reflects the re-emergence of substantial areas of dwarf-shrub heath at the opening of LPAZ MF-6 (cal.
Discussion
Was the Mountain Farm a shieling?
Previous authors are of the opinion that the Mountain Farm was occupied temporarily as a summer shieling (Bruun, 1896; Vebæk, 1992), but does the palaeoenvironmental evidence support this claim? Traditionally, sæters in the Norse tradition have been divided into full, haymaking and dairy shielings (Reinton, 1969), an idea developed further by Albrethsen and Keller (1986) who suggested a model that may have worked in a Greenlandic context. This is summarised in Table 4 along with our hypothesized palynological signatures for each of the three stages associated with such activity.
Expected activities and associated structure for different types of shieling (summarised from Albrethsen and Keller, 1986). A ‘signature’ is proposed for each activity which may be tested against the palynological record.
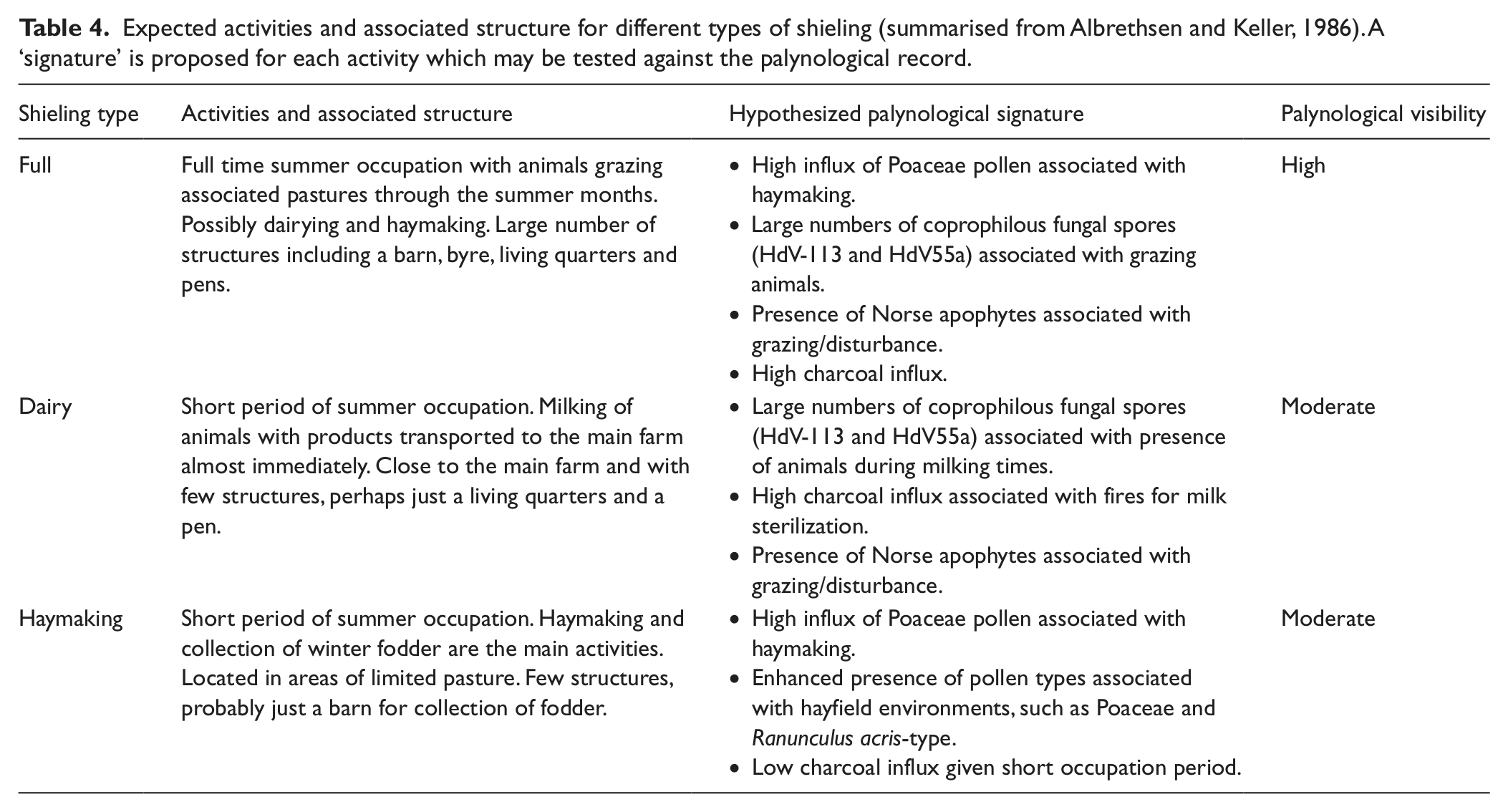
Assuming that LPAZ MF-3 reflects local landnám, this can be used as a point of departure for examining whether palaeoenvironmental evidence can classify the shieling at Ø70. Evidence for the grazing of animals is present in the form of high numbers of spores from the coprophilous Sporormiella-type (HdV-113) and perhaps also in the declining LOI values which may indicate soil erosion initiated by grazing and trampling. A diverse pollen assemblage including the Norse apophyte Rumex acetosella, suspected to respond to grazing, provides further evidence for grazed areas (Schofield and Edwards, 2011). This discounts the possibility of the site having been a simple haymaking shieling. In this context a general decline in pollen influx may be an indication of grazing-suppressed pollen production (cf. Waateringe, 1993).
Palaeoenvironmental evidence can therefore eliminate one of the possible models, but determining whether the Mountain Farm operated as a dairy or full shieling on the basis of such evidence is more difficult. Dairy shielings might be expected to have similar impacts to a full shieling, with grazing occurring for both categories alongside increased burning associated with milk sterilization (Albrethsen and Keller, 1986). There is evidence for both activities in MF-3, yet the strength of the pollen, charcoal and coprophilous fungal spore evidence seems to point towards the site having been occupied for extended periods. Dairy shielings would have been occupied for only short periods of perhaps up to one month (Albrethsen and Keller, 1986), which would be unlikely to produce the strong grazing and burning signals seen in the palynological record for Ø70. It therefore seems improbable that the site was being used as a dairy shieling.
Archaeological evidence would also support this conclusion. Eight ruins are present, amongst which is a large dwelling (29 m × 12 m) containing five rooms and a large (21 m × 9 m) structure of uncertain use which was probably a byre (Vebæk, 1992). This number of structures would almost certainly not have been present at a dairy shieling (Albrethsen and Keller, 1986). A note of caution is necessary as the chronology of the ruins is highly uncertain (Roussell, 1941; Vebaek, 1992) and they may relate to a later period than that represented in MF-3.
On the basis of the palaeoenvironmental evidence alone, it would seem reasonable to assume that the Mountain Farm operated as a full shieling during the period represented by MF-3, with the grazing of livestock perhaps just one of a wider range of activities (cf. Lucas, 2008; Reinton, 1969). Palynology can offer little insight into the nature of supplementary activities except to suggest whether haymaking may have been one of them. High Poaceae counts of c. 40–45% during MF-3 infer hay production, but declining influx of Poaceae pollen suggests otherwise. When the associated taxa are examined there are no real indications of hayfields. The contribution from Ranunculus acris-type is low (in this LPAZ, at least) as are other representatives of modern assemblages from hayfields in southern Greenland (Schofield et al., 2007).
The beginning of LPAZ MF-3 and the inferred commencement of shieling activity dates within the interval cal.
A change in the usage of the site appears to be indicated from the opening of MF-4 (cal.
The Mountain Farm and shielings in the Norse cultural sphere
Shielings, which can arguably be traced back to at least the 1st millennium
In Iceland the model has been found to be of limited utility (Sveinbjarnardottir, 1991). Some shielings were abandoned as early as the 11th century (Lucas, 2008) whilst others in northern Iceland developed into tenanted farms (Sveinbjarnardóttir, 1991) and some perhaps alternated between the two (Sveinbjarnardóttir, 1991; Sveinbjarnardóttir et al., 2011). Indeed, Lucas (2008) proposes that some Icelandic ‘shielings’ may never have been intended for such use, and were markers of land claims in an evolving cultural landscape. Conversely, and unlike those in Iceland and Norway, Faroese shielings gradually disappear following changing land management practices from the 12th century onwards (Borthwick et al., 2006; Mahler, 1993).
Despite the varying trajectories of the system in each area, archaeological excavations do suggest similarities between each region. For instance Norðungusel in Kjarardalur, Iceland (Sveinbjarnardóttir et al., 2011) is comparable with Arigsbrekka in the Faroes (Mahler, 1991, 2007). Both sites show strong similarities to those in Norway (cf. Magnus, 1986), being located within 3 km of core settlement areas or neighbouring farms and comprised around six to ten structures, some of which contained internal divisions (Magnus, 1986; Mahler, 1991; Skrede, 2005). The material culture is also similar, pointing to textile and iron working being subsidiary activities. However, there is again much variety in the Icelandic types. For instance, at Pálstóftir in eastern Iceland there are only four structures located over 3 km from the nearest farm and hunting appears to have been the primary focus (Lucas, 2008).
The Mountain Farm is therefore unremarkable, on the basis of geography and material culture, from the majority of shielings in the Norse-settled North Atlantic, suggesting that Albrethsen and Keller’s (1986) assumptions are sound. The site is c. 3 km from the nearest farm, had a similar material culture and comprised eight ruins with internal divisions (Vebæk, 1992). Pollen analyses in Norway and parts of Iceland, where there is strong evidence for grazing-induced erosion and the clearance of scrub/woodland to allow hay production, also point to a similar conclusion (Kvamme, 1988; Moe et al., 1988; Sveinbjarnardóttir et al., 2011). Yet the pollen data from the Mountain Farm also imply a phased development of the site, with a full shieling initially, followed by a small farm c. cal.
Conclusions
Shielings and transhumance agriculture have long been suspected in Greenland on the basis of their appearance at other North Atlantic islands colonized by Norse settlers. Albrethsen and Keller (1986) were the first to provide a framework for understanding how they might relate to this wider evidence. Prior to the current paper, no attempt has been made to test the assumptions of their model. The high resolution pollen-analytical approach adopted here has allowed for a greater understanding of these sites. Primarily the data show a palynological signature comparable with other farm sites in the Norse Eastern Settlement, but with tentative indications of fire having been used for landscape management and evidence for extensive herbivore grazing. A shieling system is indicated in which a full shieling is established cal.
Footnotes
Acknowledgements
We are grateful to Dawn Mooney, Dmitri Mauquoy and Andy McMullen for wood, moss and botanical identifications, respectively. Sikuu Motzfeld provided impressive hospitality at Timerliit during our fieldwork. The comments of referees are gratefully acknowledged.
Funding
The Leverhulme Trust is thanked for financial support and Gordon Cook provided radiocarbon dates.