
Abstract
Peat cores from Wylde Lake Bog in temperate southern Ontario, Canada, were examined to determine factors affecting Holocene ecological change and long-term rate of carbon accumulation (LORCA). By 10,250 cal. BP, a marsh was established, characterized by sediments with higher bulk density, highly decomposed material in the macrofossil record, and lower LORCA. By 8100 cal. BP, the marsh was replaced by a peat-accumulating fen dominated by herbaceous taxa and non-Sphagnum mosses. Around 4000 cal. BP, transition to a Sphagnum-dominated bog took place. These directional changes suggest a strong role for autogenesis, although periodic fluctuations in macrofossil assemblages and in LORCA suggest an important secondary role for climatic change and disturbance in explaining Holocene changes. LORCA remained fairly stable through the fen and bog zones, ~18 g C/m2/yr. In the high-resolution record spanning the most recent 1800 years, relatively higher values for LORCA and macrofossils associated with wetter conditions were recorded around 1400 cal. BP, partially coincident with the ‘Medieval Climate Anomaly’. LORCA decreases to minimum values for the record during part of the ‘Little Ice Age’ as woody macrofossils increase, suggesting a drier peat surface. The most recent portion of the record shows significant changes in LORCA and in bog vegetation associated with anthropogenic land clearance and damming. The charcoal record suggests that fire did not play an important role in peatland dynamics in the pre-industrial Holocene; however, a major anthropogenic fire in
Introduction
While not as extensive as northern peatlands, temperate peatlands should nevertheless be considered in global syntheses of carbon (C) accumulation in wetlands. Estimates of the temperate peatland C pool are poorly constrained because of spatial fragmentation and the fact that temperate wetlands have been commonly drained for agriculture and development. Extending from the extra tropical latitudes (>23.5°) to the boreal forest ecotone (between 45° and 50°), temperate peatlands are estimated to cover 0.2–0.9 million km2 (Matthews and Fung, 1987; Poulter et al., 2006). These estimates are highly uncertain and are complicated by large-scale wetland drainage across temperate regions of North America and Eurasia over recent centuries. The study region of southern Ontario, Canada, for example, has experienced >80% wetland losses over the past 150 years (Mitsch and Gosselink, 2007). More information on rates and controlling factors for C accumulation in temperate peatlands is needed for global syntheses of Holocene C accumulation, and for predicting behavior of peatlands presently situated in boreal or sub-arctic climates under climate warming scenarios. This study provides multi-proxy, medium-resolution whole Holocene and high-resolution late Holocene records from an ombrotrophic peatland in southwestern Ontario to determine how C accumulation rates vary over the Holocene and the roles of climate, autogenic succession and disturbances, including fire, in explaining that variability.
Climate is a first-order control on biological production and microbial metabolism, thus critical in understanding accumulation of peat. Numerous studies have linked Holocene climates to peatland initiation and to rates of C accumulation in peatlands. The early to mid-Holocene is characterized by high summer insolation and high temperatures in high-latitude regions, culminating in the Holocene Thermal Maximum (HTM), resulting from the Earth’s orbital variations. These conditions may have promoted initiation of peat accumulation at many arctic, sub-arctic, and boreal sites (MacDonald et al., 2006) and elevated rates of C accumulation in these northern peatlands (Yu et al., 2009). In terms of early Holocene paleoclimates in southern Ontario, pollen-based reconstructions suggest that HTM warming was associated with a transition from Picea–Pinus forest to woodlands dominated by Pinus and deciduous trees; warm and dry climates of the mid-Holocene in southern Ontario transitioned to cooler, wetter ‘Neoglacial’ climates following 3200 cal. BP (Yu, 2003; Yu et al., 1996).
In addition to broad-scale, supra-millennial changes in orbitally driven insolation anomalies, sub-millennial climatic changes driven by the dynamics of ocean-atmosphere circulation, volcanic activity, and variability in solar activity are also relevant to C accumulation in peatlands (Charman et al., 2013). The best described of the sub-millennial scale changes in the Holocene include the ‘Medieval Climate Anomaly’ (MCA) and the ‘Little Ice Age’ (LIA; Mann et al., 2009). A precisely dated pollen record from a varved lake along with pollen records from a series of sites in southern Ontario indicate the onset of LIA cooling in the region around
Uncertainties exist over how rates of C accumulation change with changes in temperature or moisture regime. Some studies suggest that high water table positions associated with cooler climates reduce vegetation decomposition, which results in higher C accumulation rates (Glaser et al., 2004; Loisel and Garneau, 2010). On the other hand, it has also been shown that low rates of C accumulation occur in cold climates where little biomass production is possible because of low temperatures and a short growing season, despite lower rates of decomposition (Mauquoy et al., 2002). Other studies indicate that higher primary productivity under warmer climates results in greater C accumulation, despite the enhanced rates of peat decomposition, as long as evaporative demand does not exceed precipitation (Cai and Yu, 2011; Jones and Yu, 2010). Cai and Yu (2011) posit that longer growing season and abundant precipitation lead to increased biomass production, while a freezing season even of moderate length decreases overall decay. The present study provides a high-resolution record of C accumulation for the past millennium, for evaluation of the effects of MCA and LIA climates on C accumulation.
While climate is a driver of the metabolic processes which ultimately control the potential for peat and C accumulation, autogenic processes have long been recognized as key drivers of peatland form and function (Jackson et al., 1988; Walker, 1970). Autogenic processes include a suite of ecological, hydrological, and biogeochemical changes associated with gradual accumulation of peat, and elevation of the peatland surface above the water table. Carbon accumulation in peatlands can be related to successional stage and vegetation community (Van Bellen et al., 2011b; Yeloff and Mauquoy, 2006). Van Bellen et al. (2011b) show that long-term rates of C accumulation (LORCAs) increase following a fen to bog transition because of the lack of decay-resistant Sphagnum spp. in fen systems; this increase takes place without an apparent climatic driver. Van Bellen et al. (2011b) also note differential decay among Sphagnum spp., as species associated with increased surface wetness tend to decay more quickly than species associated with drier surfaces. It has also long been noted that autogenic succession is frequently interrupted by disturbance, resulting in unique trajectories of ecological change at each site (Klinger, 1996). Thus, disturbance regimes must be considered when explaining long-term C dynamics. Fire is the major natural disturbance influencing net C accumulation in peatlands, as fire removes vegetation and initiates decomposition (Turetsky et al., 2002).
The objectives of the present study are to produce a multi-proxy paleoecological record, coupled with measurements of apparent rates of long-term C accumulation, for a temperate bog. The record will be used in conjunction with regional pollen-derived paleoclimate data to determine the relative importance of climatic versus autogenic drivers of change in vegetation communities and in C accumulation at the site and uses charcoal analysis to evaluate the extent to which fire impacted Holocene C accumulation.
Study site
Peat cores LB02 and LB03 (43°54.3′N; −80°24.87′W; 481 m a.s.l.) were recovered from hollows ~8 m apart in Wylde Lake Bog (WLB) within the Luther Marsh Conservation Area in the Grand River watershed of southwestern Ontario (Figure 1). The climate is humid, continental. The closest weather station, Elora, Ontario, ~25 km south of WLB, and at 376 m a.s.l., reports mean annual temperature of 6 ± 0.9°C (January: −8 ± 2.8°C; July: 19.1 ± 1.1°C) and mean annual precipitation of 892 mm (Environment Canada, 2003). At ~500 ha in area, WLB is the largest and least disturbed bog in southern Ontario (Sandilands, 1984). The Grand River watershed is situated on the Dundalk till plain, which overlies carbonate-bearing Paleozoic sedimentary bedrock (Chapman and Putnam, 1984). Lacustrine organic-rich sediments overlying clay-rich sediments are found above the till; these materials were likely deposited as glacial spillway sediments (Chapman and Putnam, 1984; Givelet et al., 2003). Above those sediments lie several meters of peat, with maximum depths exceeding 6 m (Sandilands, 1984).
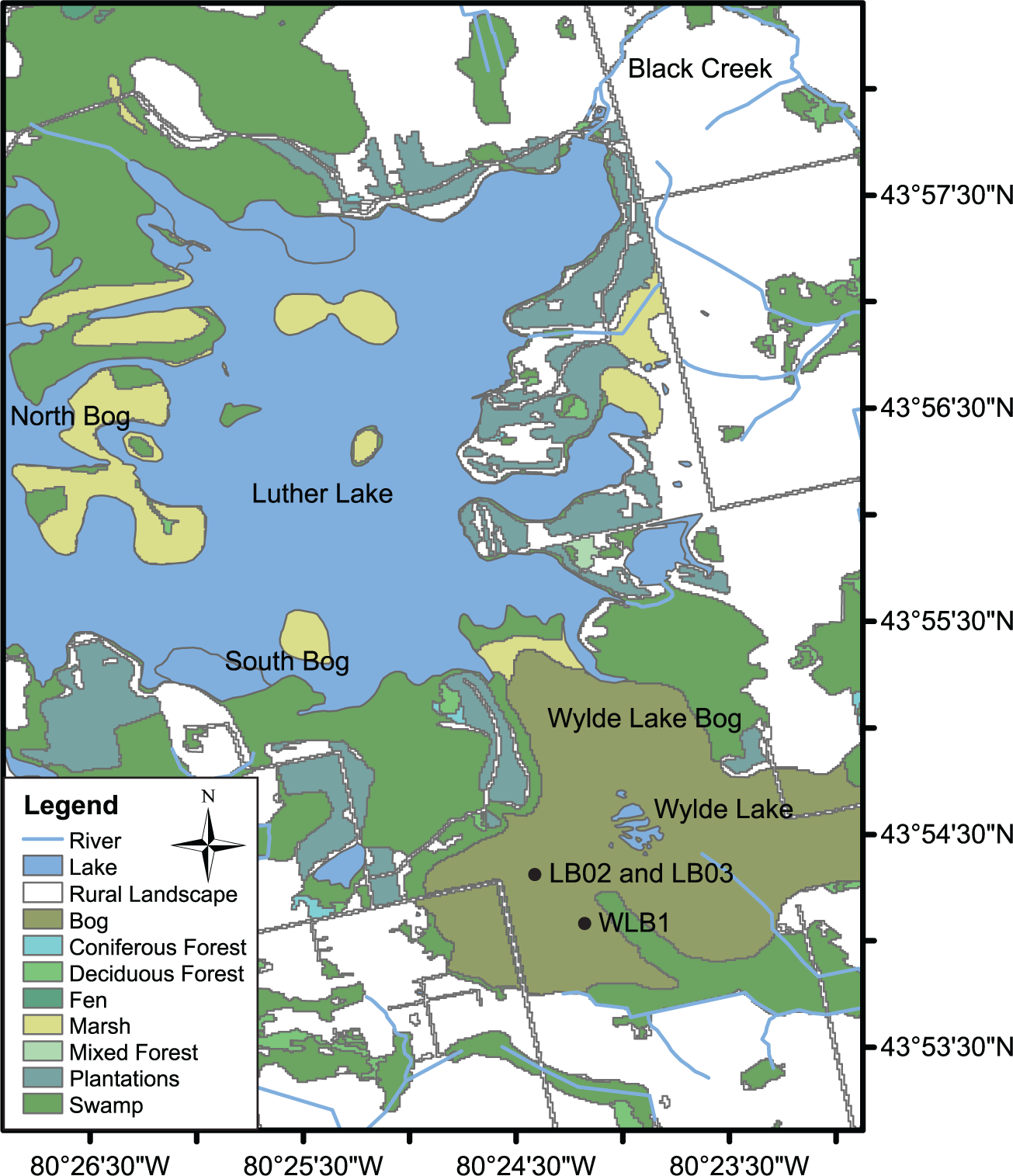
Location map showing core sites LB02 and LB03 within Wylde Lake Bog in Southern Ontario, Canada.
Because of the relatively high elevation of the Dundalk till plain, WLB exhibits some boreal-like characteristics despite its temperate latitude, including high abundance of plant taxa more commonly associated with the boreal forest to the north (Chapman and Putnam, 1984). WLB is dominated by Larix laricina and some Picea mariana, with an understory comprising moss, sedges, and shrubs. Common bryophytes include Sphagnum capillifolium, Sphagnum flexuosum, Sphagnum cuspidatum, Atrichum spp., and Warnstorfia fluitans var. fluitans; common sedges include Carex oligosperma, Carex paupercula, Carex trisperma, Eriophorum spissum, and Rhynchospora alba; and common shrubs include Andromeda polifolia, Chamaedaphne calyculata, Kalmia polifolia, Ledum groenlandicum, Myrica gale, Vaccinium myrtilloides, and Vaccinium oxycoccos (Sandilands, 1984).
Historically, Luther Marsh comprised ombrotrophic bogs, marshes, fens, two small lakes, and several streams draining into the Grand River and ultimately into Lake Erie. Following settlement around
Material and methods
Two peat cores (LB02 and LB03) were recovered from WLB in November 2011 using a Wardenaar corer (of cross-sectional area 100 cm2) for the top 30 cm of peat, and a Russian peat sampler for the remainder of the sequence. Coring refusal depth was reached at an inorganic clay. Total core lengths were 609 and 601 cm for LB02 and LB03, respectively. The cores were placed in plastic split tubes, wrapped in plastic wrap then foil, and stored in dark refrigeration until analysis.
A total of 10 samples from the two cores were submitted to Beta Analytic (Miami, USA) or the NERC Radiocarbon Facility (East Kilbride, UK) for AMS radiocarbon dating (Table 1). Samples for dating were boiled in 5% KOH, rinsed with distilled water through a 90-µm mesh, and examined under a stereomicroscope. Sphagnum stems were preferentially selected, although for depths below 225 cm, unidentifiable plant fragments were dated. Dates were calibrated using CALIB 6.1.0 with the IntCal09 calibration dataset (Reimer et al., 2009). Age–depth models were developed using linear interpolation between median calibrated radiocarbon dates using the clam package for R (Blaauw, 2010). All ages reported throughout the text are calibrated, calendar years before
AMS radiocarbon dates on macrofossils in peat cores LB02 and LB03, Wylde Lake Bog. Dates were calibrated using CALIB 6.1.0 with the IntCal09 calibration dataset (Reimer et al., 2009).

The upper sections of cores LB02 and LB03 were subsampled at contiguous 1-cm intervals for pollen analysis to document the timing of the Ambrosia rise, a regional chronostratigraphic horizon. Samples were prepared under modified standard procedures with the addition of exotic Lycopodium spore tablets to quantify pollen concentration, followed by 10% HCl, 10% KOH, concentrated HF, and acetolysis (Fægri and Iversen, 1989), and sieving at 150 µm and 10 µm. Pollen residues were stored in silicone oil and examined on the microscope at 400× magnification. Three categories were used for enumeration: Ambrosia pollen, exotic Lycopodium spores, and all other pollen and spores.
Plant macrofossil analyses were performed at contiguous 1- and 3-cm intervals on LB02 and LB03, respectively. Samples of 5 cm3 in volume were heated for ~45 min in 5% KOH and rinsed with distilled water on a 100-µm sieve (Mauquoy et al., 2010). A stereomicroscope with a 2 × 2 mm2 grid was used to identify the major peat components (Sphagnum spp., herbaceous, ligneous, ligneous roots, other mosses, and unidentifiable organic material). The amounts of these six components were estimated as percentage cover on the square grid with a categorical scale (1 = 0–5%, 2 = 5–25%, 3 = 25–50%, 4 = 50–75%, and 5 = 75–100%). Percentage cover of the six peat components was estimated for plotting purposes using the sum of the midpoint values (0–5% = 2.5, 5–25% = 15, 25–50% = 37.5, 50–75% = 62.5, and 75–100% = 87.5). Charcoal fragments were recorded in three size categories (100–250 µm, 250–400 µm, and >400 µm). Percentage peat components and charcoal fragment sizes for each sample were averaged from 15 random stereomicroscope views per pour (2–4 pours per sample). Plates from Laine et al. (2011), Levesque et al. (1988) and Mauquoy and Van Geel (2007), and a moss reference collection housed at the paleoecology laboratory at the University of Toronto were used as reference material to identify macrofossil remains. A medium-resolution pollen record from a site in WLB within ~1 km of the coring sites (site WLB1; Figure 1) was used to add insight to the macrofossil-based paleovegetation reconstruction of this study (raw data archived in Neotoma Database and produced by McAndrews (1996)). The raw pollen data along with two available radiocarbon dates for the WLB1 sequence were downloaded from the database, and an age model for that record was constructed using linear interpolation between the two dates and the clam package (Blaauw, 2010). Pollen sums for stratigraphic plotting were calculated based on a tree sum for tree pollen types, and a sum containing all taxa for non-tree pollen types.
Humification analysis proceeded based on the protocol outlined by Blackford and Chambers (1993). Subsamples (4 cm3) were taken at contiguous 1-cm intervals from the surface to 149 cm depth and 3-cm intervals below 149 cm depth in LB03. In LB02, subsamples (4 cm3) were taken at contiguous 3-cm intervals from the surface to 100 cm depth. Subsamples were dried to a constant weight at 60°C for 48 h and ground to a fine powder. Powdered peat subsamples were mixed with 8% NaOH, heated, and filtered through a 25-µm mesh. Absorbance was measured in triplicate using a UV-visible spectrophotometer at a wavelength of 540 nm. The absorbance errors for LB02 and LB03 were calculated using the mean differences in absorbance between samples and replicates. High-frequency changes in absorbance values in LB02 and LB03 were smoothed with moving averages in sets of four. Absorbance results for LB02 and LB03 were not detrended as there was no systematic increase in absorbance with depth. High absorbance values indicate higher concentrations of humic acid, found in more highly humified peat samples, while low values indicate peat samples that are less well decomposed.
For bulk density and analysis of C and nitrogen (N) content, LB02 was subsampled at contiguous 1-cm intervals, and LB03 was subsampled at contiguous 1-cm intervals from the surface to 149 cm depth and at 3-cm intervals from 149 to 601 cm depths. Samples of known volume were dried to a constant weight at 60°C for 48 h and weighed (Loisel and Garneau, 2010). Accurate volumetric measurements were made using a custom-built block and blade tool. Ground samples were analyzed on an elemental analyzer at the Department of Ecology and Evolutionary Biology at the University of Toronto, Canada (ESC 4010 Elemental Combustion System; Costech Analytical Technologies, Valencia, CA). C:N is plotted as a mass ratio. Apparent LORCAs were calculated using the C density for each sample and vertical accumulation rate generated from the age–depth models (Tolonen and Turunen, 1996).
Results
Chronologies
Peat core LB03 was 601 cm in length, and the full length of the core was analyzed for Holocene-scale interpretations. The chronology for the LB03 core was obtained through a combination of 210Pb and 14C dating, along with a charcoal horizon at depths 18–20 cm corresponding to a local fire documented in historical records in
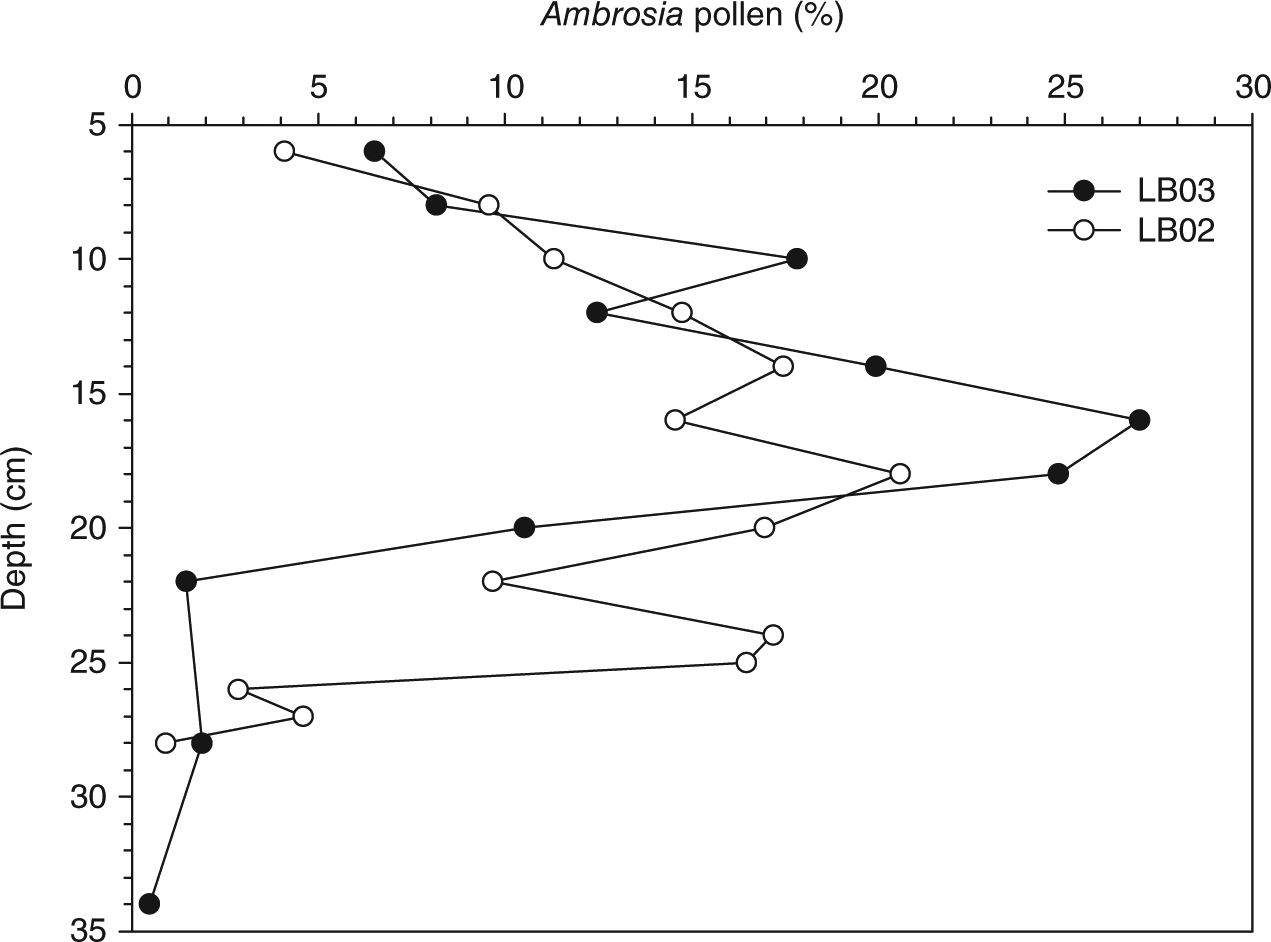
Depth (in cm) of the Ambrosia pollen rise signaling Euro-Canadian settlement, ~
The relationship between ln [excess 210Pb] and the cumulative dry mass of sediment in the peat core LB03 is approximately linear for the middle five data points (Figure 3), suggesting that ages can be modeled with some degree of confidence from 210Pb activities in the core. The upper three samples analyzed for dating show lower than expected activities, likely because of the fact that these samples included living vegetation at the surface of the peatland, and low-density acrotelm peat. Below 7 cm, the peat is more compact and similar in texture and porosity to the lower portions of the core. The break in the slope of the 210Pb activity curve (Figure 3) at depth 18 cm may be related to the peat loss caused by the local fire of
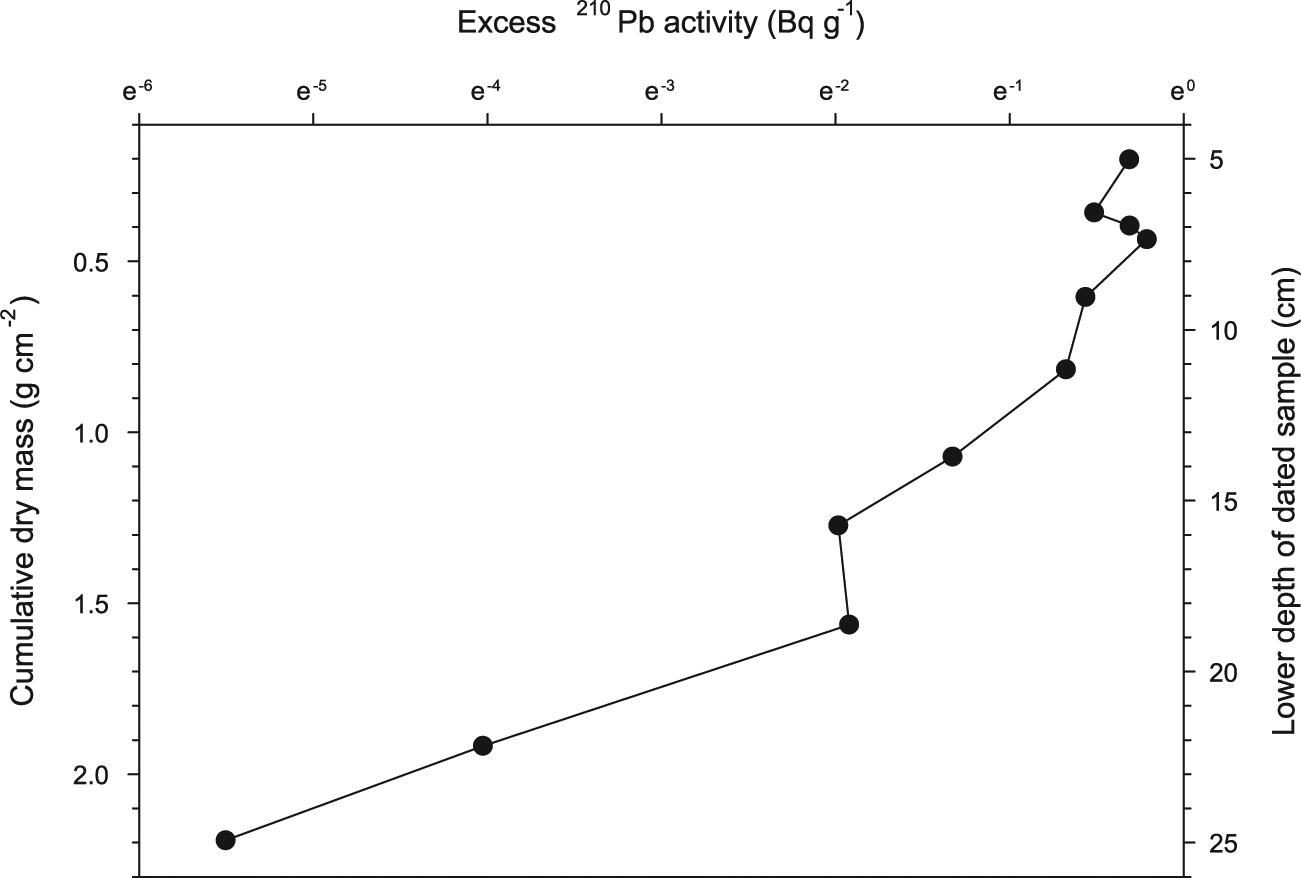
Excess 210Pb activity (ln scale), as a function of cumulative dry peat mass (g/cm2) and depth (cm) in core LB03.

Age–depth models produced by linear interpolation between dating points for (a) LB03 and (b) LB02. Radiocarbon dates are shown with error ranges associated with the probability distributions resulting from the calibrations.
The basal date for core LB03 (10,250 cal. yr BP) confirms an early Holocene initiation for the peatland, and a mean Holocene peat accumulation rate of 0.06 cm/yr. Other studies of WLB returned similar basal dates and/or mean rates of peat accumulation (Givelet et al., 2003; Irwin, 1988; McAndrews, 1996). The LB03 age–depth model shows some variability in peat accumulation, with slower rates in the early Holocene, and again in the late Holocene, after 2400 cal. BP. It is acknowledged that the low number of radiocarbon dates for a whole Holocene sequence precludes any detailed discussion of rates of change in rates of peat accumulation. The most recent apparent increase in rate of peat accumulation in the age–depth model (Figure 4) reflects the lack of peat compaction in the upper acrotelm portion of the core; therefore, the recent rate is not directly comparable to Holocene rates.
Only the top 100 cm of the second core, LB02, was analyzed. An age–depth model for this section was constructed using the charcoal horizon corresponding to the
The charcoal horizon corresponding to the

Macrofossil and humification record for (a) core LB03 and (b) core LB02. Abundances of macrofossil are expressed in terms of five categories and a percent cover scale (see text for details). Charcoal and sclerotia are plotted as numbers of fragments. Round symbols denote samples where the number of charcoal fragments is below 2. Humification is plotted as absorbance values (ABS). Major transitions in macrofossil assemblages/wetland types, the onset of MCA and LIA climate changes based on regional pollen records, and the depths corresponding to the water table (WTD) and the construction of a nearby dam (
Paleovegetation
Macrofossil analysis provides information on local paleovegetation during the period of record. At site LB03, peat accumulation begins at ~10,250 cal. BP (Figure 5a). Highly decomposed organic remains dominate the early Holocene, and this period coincides with low abundances of all other macrofossil materials; small numbers of charcoal fragments are present and peat humification is low. Around 9100 cal. BP, non-Sphagnum mosses and herbaceous peat begin to dominate macrofossil assemblages, while peat humification values increase markedly. Around 5400 cal. BP, non-Sphagnum mosses and herbaceous peat macrofossils slowly begin to be replaced by remains of Sphagnum mosses. By 4000 cal. BP, the transition from fen-like indicators to bog indicators is complete, and peat humification values increase somewhat relative to lower sections of the core (Figure 5a). The peat humification curve is variable through most of the core, and small changes in absorbance are often coincident with small shifts in the macrofossil record. The core contains some significant changes in peat-forming vegetation; thus, peat humification reflects mainly these vegetation changes, as opposed to actual variations in surface wetness (Bunbury et al., 2012; Yeloff and Mauquoy, 2006).
Sphagnum mosses are usually well preserved in the portion of the core above the fen to bog transition. This Sphagnum-dominated zone is also characterized by episodic fluctuations in the abundances of macrofossils derived from woody vegetation, while non-Sphagnum mosses are virtually absent. Charcoal fragments and sclerotia of the ectomycorrhizal fungus Cenococcum geophilum syn graniforme (Spatafora and Owensby, 2012) are occasionally observed (Figure 5a) and are interpreted as fire indicators (Benedict, 2011). The peak in charcoal fragments at 18–20 cm is associated with the
The changes documented in the LB03 Holocene macrofossil record are well supported by available pollen data from the WLB1 site (Figure 6). The Wylde Bog (WLB1) pollen record shows the presence of a number of local fen indicators from peatland initiation until about 5000 cal. BP, including Cyperaceae, Equisetum, and Larix; Sphagnum spores become abundant also after 5000 cal. BP, confirming this timing for the fen to bog transition (McAndrews, 1996).
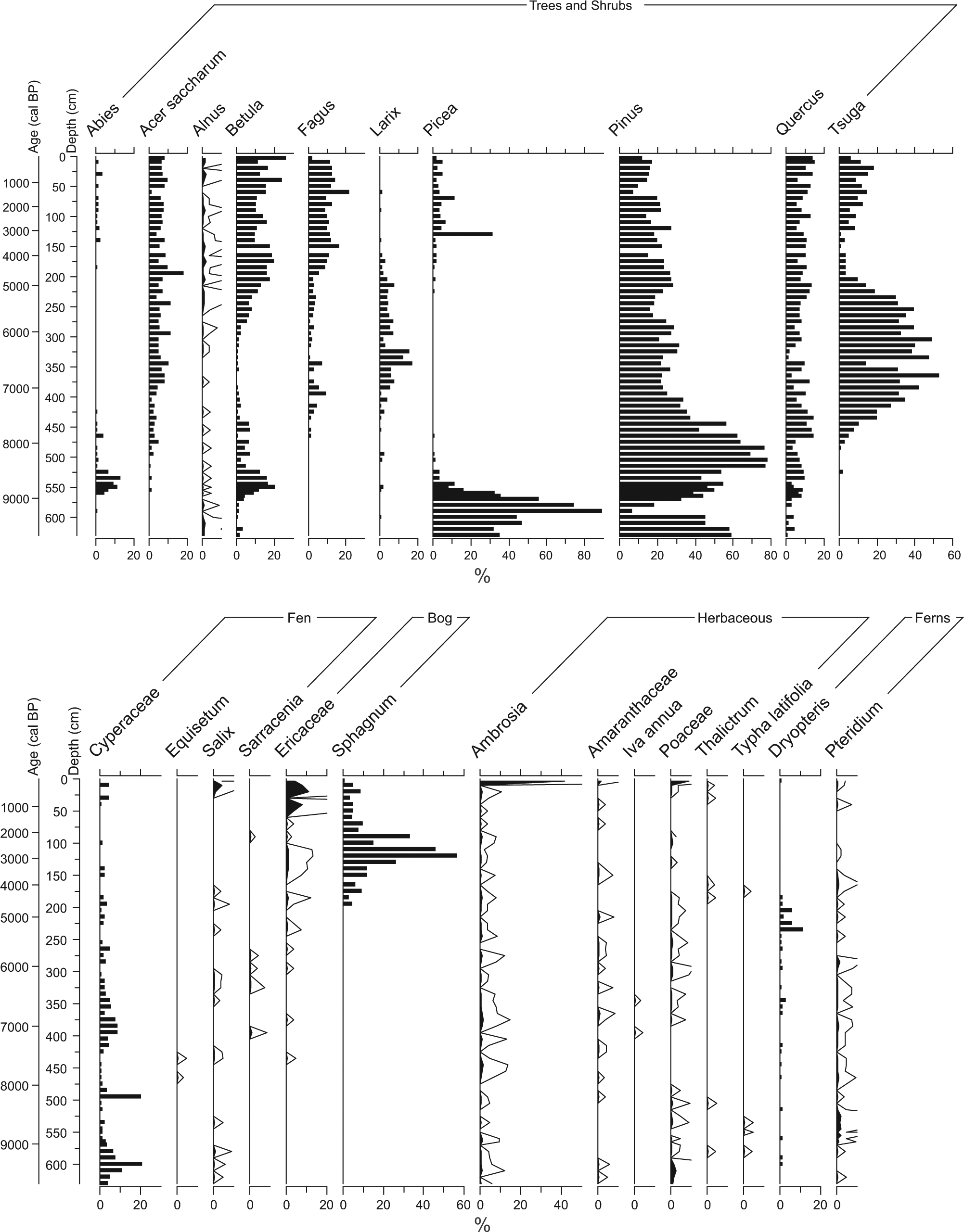
Percent pollen diagram for the WLB1 site (diagram drawn from data of McAndrews (1996)). The age model was determined from two available radiocarbon dates, and standard pollen sums were used for calculating percentages (see text for details). The upper panel shows tree and shrub pollen types; the lower panel shows aquatic, wetland, and herbaceous pollen types.
The macrofossil record from the shorter, late Holocene sequence (core LB02; Figure 5b) reflects the trends shown in the LB03 record, but the shorter sequence provides greater temporal resolution with an apparently more complete record through the most recent 1500 years (Figure 3). LB02 comprises mostly well-preserved peat-forming vegetation. Sphagnum is the dominant macrofossil type; however, the record shows three short episodes in which Sphagnum abundance decreases to <10%, while the proportions of macrofossils of herbaceous peat, ligneous peat, or roots increases. As in LB03, sclerotia of C. geophilum are recorded periodically. The significant peak in charcoal abundance at 21 cm is associated with the
C accumulation
Mean bulk density for both WLB cores is 0.07 g/cm3, mean C content for the two cores is 48.6%, and mean N content is 1.5%. Measurements of bulk density and C content were combined with the age–depth models to calculate apparent LORCA (Figure 7). In the LB03 Holocene macrofossil sequence, the oldest section of the core is dominated by highly decomposed organic matter (Figure 5a); higher values of bulk density and lower C:N ratio at that time (Figure 7a) suggest limnic sediments, associated with low values for LORCAs. By 8100 cal. BP, bulk density decreases and the macrofossil record becomes dominated by herbaceous peat; C:N ratio and LORCA begin to increase. At the fen to bog transition, beginning around 4100 cal. BP, C:N ratio increases again, leading to marginally higher values for LORCA, particularly between 2400 and 3000 cal. BP. After 2400 cal. BP, bulk density increases, and C:N ratio begins to drop as ligneous and herbaceous peat become more important relative to Sphagnum peat in the macrofossil record. Decreases in bulk density and the inferred decrease in the rate of peat accumulation since 2400 cal. BP (Figure 4) result in very low values for LORCA in the most recent two millennia in the LB03 core; this decrease, however, is related to the age model in which radiocarbon dates were likely ‘too old’ relative to the depths dated because of up to 20 cm in lost peat following the
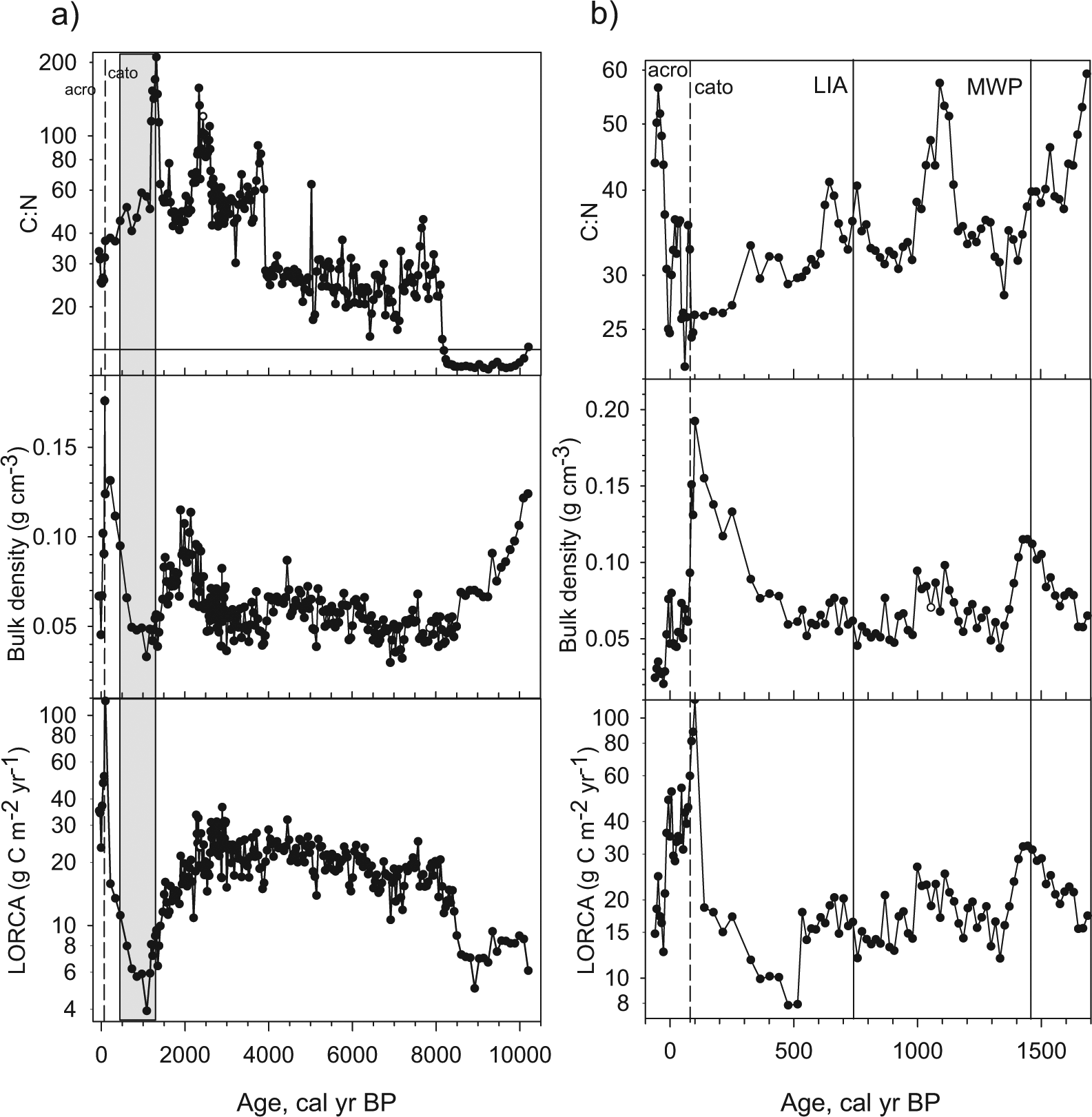
C:N (mass) ratios, bulk density and calculated LORCA for (a) core LB03 (whole Holocene) and (b) core LB02 (most recent 2000 years). The shaded bar on the LB03 curves indicates the section of the core affected by combustion and overly low rates of calculated peat accumulation because of peat losses resulting from the
In the LB02 late Holocene sequence, where we do not suspect significant peat loss, C:N ratio fluctuates with shifts in the relative abundance of Sphagnum versus ligneous macrofossils, with higher C:N ratios occurring when Sphagnum is more dominant. Lower C:N values since 700 cal. BP contribute to the lower LORCA recorded for that time (Figure 7b). The highest values for bulk density in both sequences are recorded at the charcoal horizons associated with the fire of
Discussion
Explaining variability in Holocene C accumulation
Low C:N ratios and high sediment bulk densities, coupled with highly decomposed organic remains in the macrofossil record (Figure 5a and b) and abundant Cyperaceae pollen (Figure 6; McAndrews, 1996), suggest marsh-like conditions in the early Holocene at the site of core LB03. While some higher latitude peatlands show elevated rates of apparent C accumulation in the early Holocene in response to orbitally mediated increases in summer insolation and warmer temperatures associated with the HTM (Jones and Yu, 2010; Van Bellen et al., 2011a), LORCA in the early Holocene is low at LB03. Early peatland development at this site is controlled more by post-glacial geomorphic setting than climate. Furthermore, peatlands at lower latitudes experienced insolation anomalies of lower magnitude and less pronounced thermal maxima in the early Holocene. C accumulation in temperate peatlands is therefore likely less sensitive to orbitally mediated climate changes. Other studies of temperate peatlands similarly show lower values for LORCA in early successional marsh-like phases (Cai and Yu, 2011).
Around 8100 cal. BP, C:N ratio and peat humification increase in the LB03 record, coincident with a decrease in bulk density, and a shift from highly decomposed organic remains to herbaceous peat, ligneous remains, and brown mosses in the macrofossil record, indicating a transition to a peat-accumulating fen ecosystem. LORCA values increase at this point. The WLB1 pollen record reveals an increase in fern taxa and Larix at this time (Figure 6; McAndrews, 1996), further supporting the interpretation of a fen-like environment. A shift from a shallow water marsh to fen is frequently linked to terrestrialization because of infilling by organic matter (Cai and Yu, 2011; Muller et al., 2003).
By about 4000 cal. BP, C:N ratio had increased markedly, along with Sphagnum remains in the macrofossil and pollen records, suggesting the completion of the fen to bog transition. The bog zone in the core is associated with variable LORCA; maximum LORCA values for the catotelm portion of the LB03 record occur between 2500 and 3000 cal. BP. Marginally higher values within a portion of bog zone for LORCA suggest some possibility of climatic controls, since once the bog becomes established, directional autogenic processes cannot explain episodic changes, unless nonlinear dynamics are invoked (Belyea and Baird, 2006). The pollen record shows the decline in Tsuga pollen between 5000 and 3000 cal. BP. Among other factors, drought is considered important in explaining the Tsuga decline (Haas and McAndrews, 1999). The re-initiation of wetter conditions after 3000 cal. BP, coincident with the onset of ‘Neoglacial climates’, as shown here through the re-establishment of Tsuga at that time, suggests a wetter climate which may be promoting higher values for LORCA, either by enhancing production or by reducing rates of decomposition. There is a notable decrease in LORCA following ~2270 cal. BP, which reflects the reduction in the rate of peat accumulation inferred from the age model (Figure 4). Because of peat losses of perhaps up to 20 cm related to a local fire in
The LB02 record provides an opportunity to examine changes in LORCA over the past two millennia in a sequence in which peat was likely not lost to combustion (Figure 7b). Calculated values for LORCA are variable through this period. There has been considerable interest in linking C accumulation in peatlands to the climate changes associated with the MCA and the LIA (Charman et al., 2013; Finkelstein and Cowling, 2011), observed to have been time transgressive across the northern hemisphere. To evaluate those relationships in the LB02 record, we use local pollen datasets to assign timing for MCA and LIA initiation at WLB. Campbell and McAndrews (1991) summarize pollen data from a series of records in Ontario and show using cluster analysis that pollen assemblages begin to change in response to LIA cooling after
Because of the importance of wetland pollen types in the WLB1 pollen record (McAndrews, 1996) and the relatively low sampling resolution of that pollen record (samples are on average ~150 years apart; Figure 6), it is not possible to extract detailed information from the record on the timing of the LIA and the MCA. Despite these limitations, small changes in the abundances of Fagus and Pinus pollen are noted in the WLB1 pollen data (McAndrews, 1996). The dip in Pinus pollen and concomitant increase in Fagus pollen between 1220 and 1470 cal. BP (equivalent to
LORCA as reconstructed from the LB02 core is variable over the past two millennia, and a strong contrast between values during the MCA versus LIA is not observed (Figure 7b). The MCA, ~1450 to ~750 cal. BP at the study site, however, contains periods of elevated LORCA and somewhat higher C:N ratios. The LIA phase at WLB is characterized by minimum LORCA values for the record, and lower C:N ratios. Lower values for LORCA during the LIA in the LB02 core are associated with a vegetation shift from Sphagnum spp. to more N-rich ligneous and some herbaceous remains in the macrofossil record, suggesting decreases in surface wetness during this time (Loisel and Garneau, 2010), which were likely climatically controlled (Cai and Yu, 2011). This interpretation suggests that LIA climates in the region may have been cool and dry, whereas other studies have suggested cool/wet LIA climates (Finkelstein and Davis, 2006; Pederson et al., 2005). Syntheses of multiple multi-proxy records from the region are required to resolve the LIA climate history for this region.
Role of disturbance
Similar to other studies on temperate peatland fire regimes (Lavoie and Pellerin, 2007), fires are infrequent in the LB02 and LB03 Holocene macrofossil records. Lavoie and Pellerin (2007) suggest that temperate peatlands are typically surrounded by deciduous trees or mixed forest which are not fire-prone and thus burn less frequently than boreal peatlands. In the humid climate of southwestern Ontario, natural fires are rare and, thus, do not play a large role in peatland vegetation dynamics or carbon accumulation. Charcoal fragments > 400 µm in size and sclerotia of C. geophilum are only evident in high-enough abundance to confirm the presence of one local fire for the Holocene at WLB. This is the fire of
Other recent disturbances, such as local deforestation, drainage, and damming, may also have impacted LORCA in the LB02 record; however, values for LORCA in the more aerated acrotelm portion of the peat cores should not be compared directly with results for the waterlogged catotelm, owing to different bulk densities and C composition. Additionally, the acrotelm–catotelm boundary, the location of the charcoal horizon deposited in the
While considering the above caveats, it is still clear that the LORCAs following the
Allogenic and autogenic controls on peatland development
The Holocene record of peatland development shows that this temperate wetland site in southern Ontario underwent a series of ecological changes in transitioning from a marsh to a fen to a bog; these changes mirror those reported from numerous other peatland sites. These changes in some cases were promoted by directional autogenic processes, namely, the accumulation of organic matter and the eventual segregation of the peatland surface from the water table. However, there is also evidence for climatic impacts on peatland ecology and C accumulation. For example, the drier climate inferred during the Tsuga decline may explain the periodic increases in woody vegetation on the bog surface and fluctuations in LORCA in the latter portion of the Holocene. During the past millennium, climate changes associated with the MCA were at times associated with somewhat higher values of LORCA, along with changes in peatland vegetation indicative of a wetter bog surface. The LIA was characterized by the lowest values of LORCA for the whole record, which may have been related to drier conditions, as suggested by increases in woody vegetation. Nevertheless, the difference in LORCA recorded for the MCA compared with the LIA is subtle. The climate of the study region is unlikely to have been moisture-limiting at any time during the Holocene from the perspective of peatland plant communities, thus autogenic processes are likely major factors at this site, as was also reported for other temperate wetlands (Muller et al., 2003). While in humid climates allogenic controls are less important in explaining peatland dynamics, there is evidence from the LB02 and LB03 records to suggest that periodically drier conditions did change peatland vegetation and consequently rates of apparent C accumulation.
Whereas during the Holocene there is little evidence for effects on peatland structure or function of fire or other disturbances, the LB02 and LB03 peat cores record major changes associated with a suite of disturbances since Euro-Canadian settlement in the area. The comparison of two cores, LB02 and LB03, taken in close proximity also shows the heterogeneous nature of peat accumulation across small spatial gradients and emphasizes the caution which must be used in making whole-peatland inferences from a single core. Despite this, studies such as ours indicate the potential role that temperate peatlands could have held in Holocene C dynamics. In some temperate regions, >80% of peatlands have been destroyed because of human land use change. An estimation of the total area once covered by temperate wetlands prior to European colonization might reveal that although LORCA are generally lower in temperate relative to boreal wetlands, the areal size of temperate peatlands may have been so great that they were an important sink of C for much of the Holocene. Continued research in reconstructions of C balance in Holocene peatlands of the temperate, boreal, and high-latitude regions will likely show that peatlands may have exceeded Holocene vegetation as C sinks and that their modern-day representation severely under-represents their potential for C storage over most of the past 10,000 years.
Footnotes
Acknowledgements
We thank Angela Gallego-Sala for providing two radiocarbon dates as part of the ‘Peatlands in the Global Carbon Cycle During the Past Millennium’ project. We also thank Kristen Beck, Maara Packalen, and Magadalena Sobol for field and laboratory assistance, and Tony Davis and Nathan Basiliko for helpful discussions. Pollen data were obtained from the Neotoma Paleoecology Database (![]() ); the work of Jock McAndrews on the palynology of Wylde Bog and of the Neotoma community is gratefully acknowledged.
); the work of Jock McAndrews on the palynology of Wylde Bog and of the Neotoma community is gratefully acknowledged.
Funding
This project was supported by a University of Toronto graduate fellowship to Shiller, a grant from the Natural Sciences and Engineering Research Council (NSERC) of Canada to Finkelstein, and a grant from the Canadian Institute for Advanced Research (CIFAR) Earth System Evolution Program to Cowling. The ‘Peatlands in the Global Carbon Cycle During the Past Millennium’ project was funded by NERC Grant NE/I012915/1.