
Abstract
A paleoecological analysis of a 6-m-long sedimentary sequence from the La Encrucijada Biosphere Reserve (RBLE) from southern Pacific Mexico provides detailed information about past vegetation changes between ~6200 and 700 cal. yr BP. Highest percentages of mangrove pollen in pollen zone I (from ~6200 to 5400 cal. yr BP) suggest moister and warmer conditions than at present related with global rates of sea-level rise and northward position of the Intertropical Convergence Zone (ITCZ), suppressing El Niño-Southern Oscillation (ENSO) events. Drier conditions and decreasing rates of sea-level rise at ~5400–4500 cal. yr BP are evidenced in pollen zone II by the lowest percentages of mangrove pollen and highest presence of disturbance pollen. Recovery of mangroves and tropical forests occurred from ~4500 cal. yr BP, suggesting moister conditions. Agricultural activities are recorded around ~4900, 4700 and 2400 cal. yr BP, although human occupation was scarce in the mangrove area. ENSO variability correlates to mangrove pollen variation between 3600 and 2400 cal. yr BP. A protracted drought is observed at ~1300–700 cal. yr BP characterized by proportions in montane forest pollen and may be related with southern oscillation of the ITCZ and the Classical Mayan Demise.
Introduction
Mangroves are complex plant communities fringing coastal areas (Lugo and Snedaker, 1974) that develop in tidal zones of the tropics and sub-tropics, where soils are generally water saturated and the salinity can be equal to (or higher than) that of the sea (Hogarth, 1999). Mangroves are distributed along the Pacific Coast of Mexico (Rzedowski, 2006) and are considered one of the most diverse, rich and productive ecosystems in the country (Instituto Nacional de Ecología (INE), 1999a). In Chiapas, Mexico, mangroves provide environmental services such as soil retention and protection from floods during high tides, rain storms (INE, 1999a) and tsunamis (Alongi, 2008). Mangrove swamps also act as refuges and nesting areas for a number of bird species and as nursery habitats for commercially important fisheries (Tovilla-Hernández et al., 2009), making mangrove ecosystems significant areas for conservation as well as the main food source for people living within these areas (INE, 1999a). However, the mangroves along the coast of Chiapas are threatened by growing agricultural frontiers and inefficient management of natural resources in the high basins (Tovilla-Hernández et al., 2009), coupled with the predicted consequences of global climate change (Gilman et al., 2008).
Climate conditions during the Holocene have not been stable (Mayewski et al., 2004), and plant communities react according to the intensity and length of climate events (Williams et al., 2002). The local occurrence of mangroves depends on abiotic (i.e. winter sea-temperatures higher than 20°C, availability of soil, special geomorphologic conditions and fluctuating hydrologic conditions) and biotic factors (i.e. plant succession and propagule sorting; Hogarth, 1999; Tomlinson, 1986). The link between the climate and the ecological conditions affecting mangroves makes the areas where they grow ideal sites for tracking past climate changes (Torrescano-Valle and Islebe, 2012) because mangrove deposits (peat) provide the wet and anaerobic conditions required for the long-term preservation of environmental indicators (proxies; Ellison, 2008).
Pollen has been widely used as a proxy in sediment records because of the possibility of identifying pollen at several taxonomic levels. Thus, pollen records provide a reliable snapshot of the vegetation in a particular space and time (Bennett and Willis, 2001). Shifts in past climate conditions have been inferred from the changes in vegetation observed in the pollen record in mangrove peat worldwide (Ellison, 2008). Records from New Zealand, Oman and West Africa have revealed that during the early to middle-Holocene, increasing sea levels and warm global conditions favoured the development of mangroves at higher latitudes where they no longer exist (Mildenhall, 2001), or allowed more extensive distributions of these species than are observed today (Lézine, 1997; Lézine et al., 2002). Stanley and Hait (2000) found that mangrove peat records and sedimentation rates in the Great Sundarbans (Bangladesh and India) are best explained by subsidence processes occurring during the Holocene. While rising sea levels in the Bay of Bengal caused brackish and mangrove environments to occur intermittently from 7500 to 5500 calibrated years before present (cal. yr BP), later (~4870–4780 cal. yr BP) freshwater dominance resulted as decreased sea levels (Rashid et al., 2013). A similar conclusion has been proposed by Cohen et al. (2012) for northern Amazonia in Brazil, where rising sea levels, tectonic subsidence and low river discharge were responsible for the development of a major mangrove belt along the coast during early and middle Holocene. Subsequently, in the late Holocene, increased precipitation disrupted the mangrove belt at estuaries, turning them into freshwater systems.
Precipitation along the Pacific coast of Chiapas is related to the climate conditions of the circum-Caribbean region, with both areas being influenced by the Intertropical Convergence Zone (ITCZ). North–south movements of the ITCZ determine the amount of moisture transported from the Caribbean sea to the Pacific Ocean via the Isthmus of Panama (Leduc et al., 2007). Paleoenvironmental records from Caribbean Central America indicate the existence of increased precipitation, temperatures and sea levels from ~8000 to ~4000 cal. yr BP (Islebe and Sánchez, 2002; Monacci et al., 2011; Mueller et al., 2009), interrupted by minor dry episodes (Carrillo-Bastos et al., 2010; Haug et al., 2001) and reduced sea level (Wooller et al., 2007). Beginning at ~4000 cal. yr BP, the same records indicate reductions in available moisture. At ~3000 cal. yr BP, the records show discrepancies in the timing, length and strength of climate conditions: humid conditions have been reported for Puerto Morelos (Islebe and Sánchez, 2002), Punta Laguna (Curtis et al., 1996) and the Lacandon rainforest (Domínguez-Vázquez and Islebe, 2007), while dry conditions prevailed in the Cariaco basin (Haug et al., 2001) and Lake Tzib (Carrillo-Bastos et al., 2010), and intense human activities occurred during a humid episode in the lowlands of northern Guatemala (Mueller et al., 2009). Despite these contrasting conditions, these records show a drying trend towards ~1000 cal. yr BP coinciding with the Classical Mayan Demise.
Surveys of past environmental conditions carried out along the Pacific coast of Central America remain scarce. Major introgression events have been recorded in the Panama Basin from 11,300 to 5600 cal. yr BP, coupled with higher precipitation and warmer temperatures, except for a period of reduced moisture from 7000 to 5640 cal. yr BP (González et al., 2006). Blake et al. (1995) present data on human occupation on the Pacific coast of south-eastern Mesoamerica, evidencing Early Formative period ceramics. The 1550 BC first sedentary communities are recorded for coastal Chiapas, suggesting fishing and hunting as main activity, but with presence of cultivated plants (Blake et al., 1995). Chisholm and Blake (2006) suggest horticulture since ~10,000–4000 cal. yr BP. Neff et al. (2006a) present evidence of Zea phytoliths from Guatemala around 5500 cal. yr BP. Kennett and Voorhies (2002) record sedentary fishing farming between 3500 and 2800 cal. yr BP. Records from Guatemala (Neff et al., 2006a, 2006b, 2006c) and Chiapas (Kennett et al., 2010; Kennett and Voorhies, 1996; Voorhies et al., 2002) provide important insights about early human occupation in the area. These studies revealed the first significant information about climatic factors that affected Pacific Central America in the Holocene. However, the low resolution in these records and the interpretation of the results (mainly focused on human occurrence) restrict the available information related to the responses of vegetation to past climate changes in the area.
This study refers to the responses of vegetation to environmental changes along the coast of Chiapas during the middle and late Holocene using pollen as a proxy. The pollen records provide useful first insights into the events that influenced coastal environments during the Holocene (Leyden, 2002). The purpose of this study is to elucidate the climate and human contributions producing vegetation changes in mangrove areas, considering past climate variability (e.g. El Niño-Southern Oscillation (ENSO) variability) and human activity.
Study area
The sampling site (15°8.98′N, 92°45.06′W) was located within La Encrucijada Biosphere Reserve (RBLE) in the Mexican state of Chiapas (Figure 1). The climate of the area is tropical wet/dry (Aw), and the mean annual temperature is 28°C, with annual precipitation ranging between 1300 and 3000 mm. The rainy season occurs from May to November, with a mid-summer drought in July and August. The dry season is from December to April, with occasional precipitation being observed in March and April (INE, 1999a). The amount of precipitation in the area is determined by annual movements of the ITCZ, creating contrasting conditions between the rainy (northern position of the ITCZ) and dry seasons (southern position of the ITCZ). Drought conditions prevail in the rainy season during ENSO events, when the ITCZ does not reach its normal northern position.
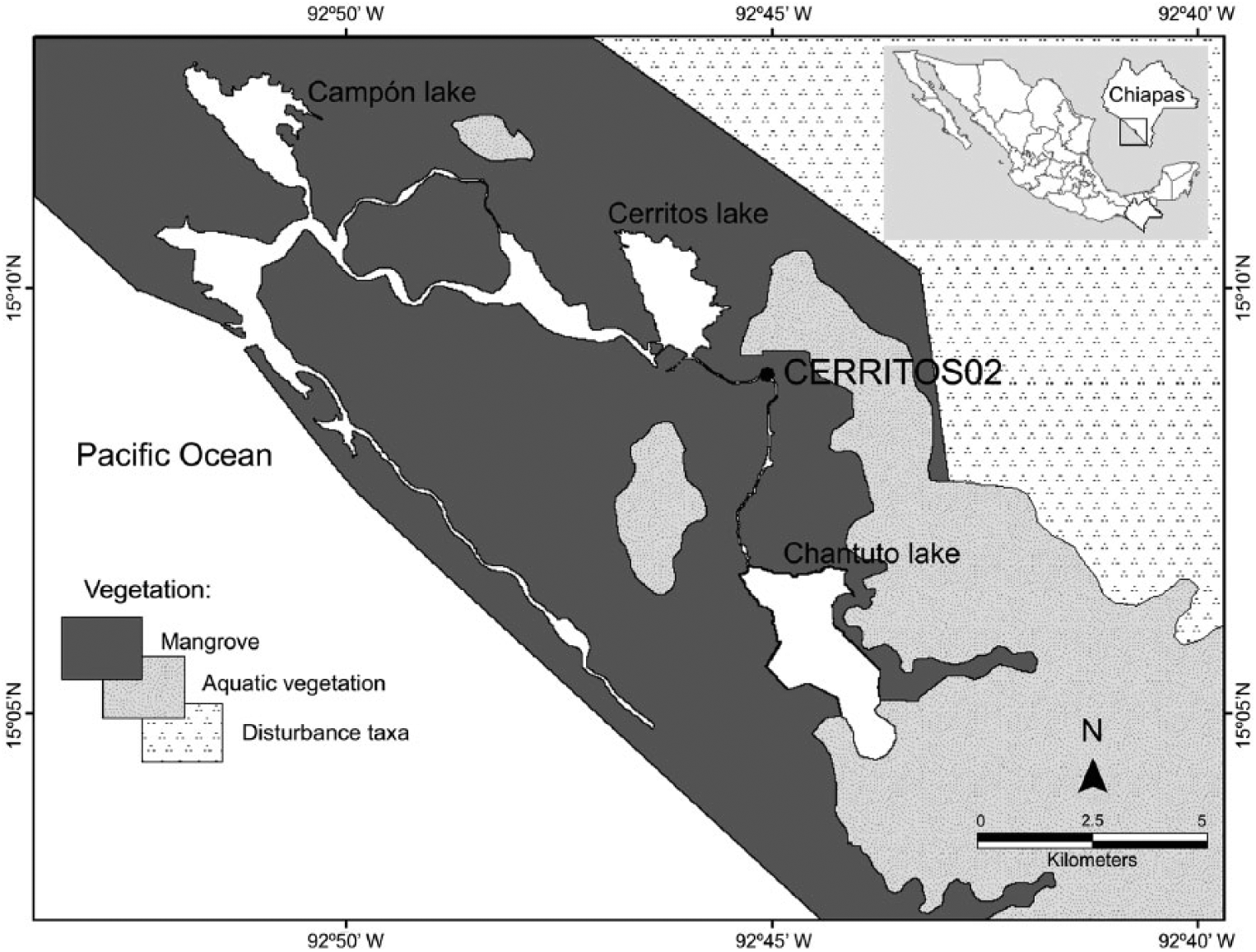
Location of the coring site, the main water bodies and the main vegetation types in the area.
The physiographic conditions of the area correspond to the Guatemalan Coastal Plain, an intricate network of debris fans along the Pacific coast consisting of volcanic deposits that accumulated during the Quaternary (Marshall, 2007). The soils are mainly alluvial and lacustrine, although palustrine and littoral soils may occur. The main soil groups are solonchaks, regosols, cambisols, gleysols, phaeozems and fluvisols. Complex lagoon systems are created by the infilling of rivers flowing from the Sierra Madre de Chiapas and tides from the Pacific Ocean (INE, 1999a).
The principal vegetation types within RBLE are mangroves, high tropical forests, gallery forests, salt marshes, coastal dune vegetation and Pachira aquatica Aubl. forests. Rhizophora mangle (L)., R. harrisonii Leechm., Laguncularia racemosa (L.) C.F. Gaertn., Avicennia germinans (L.) Stearn and Conocarpus erectus L. represent the species of mangroves in the area, showing medium heights ranging from 5.9 to 7.9 m (C. erectus) to 22–27.6 m (R. mangle and R. harrisonii; Tovilla-Hernández et al., 2009). The vegetation surrounding the sample site consists of a low-stature pure stand of R. mangle, 8–10 m in height, situated 8 km inland from the shoreline. The sediment at the coring site consists of muddy organic peat.
Materials and methods
The selection of the coring site was focused on areas with little or no influence of streams and apparently high sedimentation. Core CERRITOS02 was extracted using a Russian corer in 50-cm-long sections, down to a total depth of 600 cm, with no overlapping. Each core section was wrapped, labelled and transported to the ECOSUR palynology laboratory. Sediment samples were taken at 5-cm intervals for pollen extraction. A total of 100 pollen samples were obtained using standard HCl, KOH and acetolysis methods (Erdtman, 1943; Faegri et al., 1989). Exotic Lycopodium spores were added to each pollen sample to assess the pollen concentration and pollen influx. Identification of the pollen taxa in the samples was possible using the keys of Hooghiemstra (1984), Palacios-Chávez et al. (1991), Roubik and Moreno (1991) and Sánchez-Dzib et al. (2009) and the pollen reference collection of ECOSUR – Unit Chetumal.
At least 350 pollen grains were counted per sample using a 400×-magnification light microscope. A minimum of 100 pollen grains was counted for samples with low pollen counts. The pollen taxa were grouped according to their ecological affinities into mangrove, tropical forest, montane forest, disturbance, crop plants and aquatic taxa. The aquatic elements (Cyperaceae, Nymphaceae and Typha) and fungal and fern spores were counted, but not added to the total pollen count because of over-representation from local elements. To recognize additional environmental features, Foraminifera and charcoal were also recorded in the presence/absence counts.
The integrated data were presented in percentages diagrams of the pollen taxa generated in the TILIA 1.17.16 software package (Grimm, 2001). Pollen zones were delimited according with composition and abundance of the pollen taxa using Constrained Incremental Sum of Squares (CONISS) in TILIA.
Four radiocarbon (14C) ages were obtained via accelerator mass spectrometry (AMS) from wood, plant remains and organic sediment at Beta Analytic Inc., Miami, Florida. Calibration was performed with the Calib Radiocarbon Calibration Program 7.0.0 (Stuiver and Reimer, 1986–2013) using the IntCal13 curve (Reimer et al., 2013) with 2σ probability intervals. Chronology was calculated via linear regression between the four calibrated ages and extrapolation of the regression model. Sedimentation rates and sample resolution were calculated based on chronology, and hence changes in sedimentation rates outside the range of the calibrated ages were not considered.
Results
Lithology and time control
Fine dark brown/black peat with abundant fine remains of plants (i.e. leaves) is representative at the bottom of the core from 600 to 417.5 cm interrupted by 5-cm sections of dark grey organic-rich clay at 575 and 540 cm. A dark grey organic-rich clay is observed from depth 417.5 to 355 cm, and organic detritus are almost absent in this horizon, although fine leaves are noticed at 370 cm. From 355 to 147 cm, the sediments turn into a dark brown/black peat with dark grey organic-rich clay intersected at 290–300 and 200–194 cm; wood fragments are present at 292 cm. Fine grey clay is found from 147 to 60 cm interrupted by dark brown/black peat at 108–92 cm; organic detritus are absent in this section. A dark brown peat without organic detritus is noticed at the upper horizon of the core from 60 to 0 cm.
The four AMS 14C ages (Table 1) and chronology show that core CERRITOS02 represents the last 6200 cal. yr BP, with a mean annual sedimentation rate of 0.8317 mm/yr. No significant changes were observed in the sedimentation rate (Figure 2). The average resolution was ~60 years per sample. As observed by Neff et al. (2006b) in a nearby study site, sand layers were not found in the stratigraphy, discarding the occurrence of intense rain storms and tsunami events.
Radiocarbon (AMS) dates and calibrated dates for core CERRITOS02.
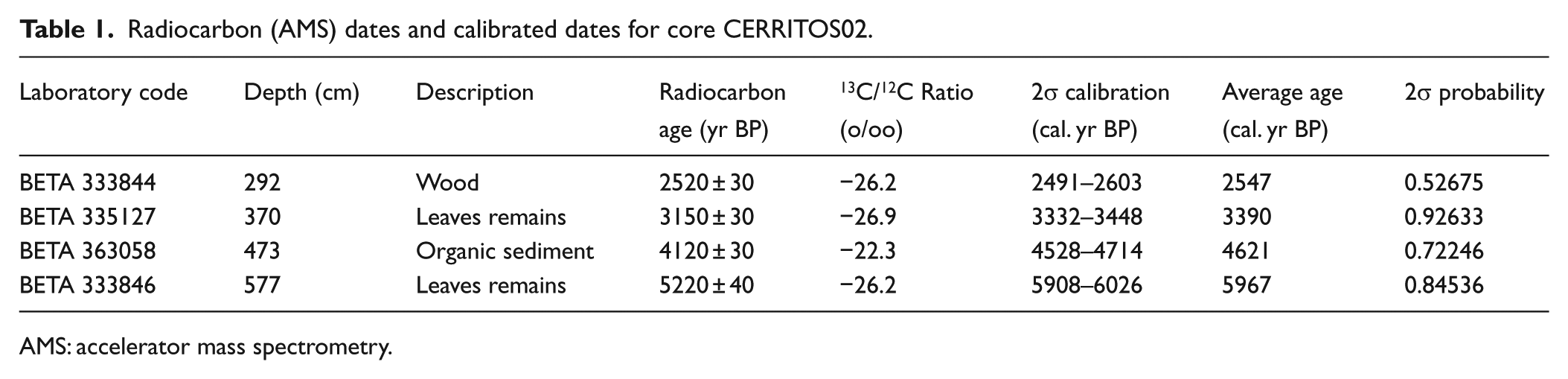
AMS: accelerator mass spectrometry.
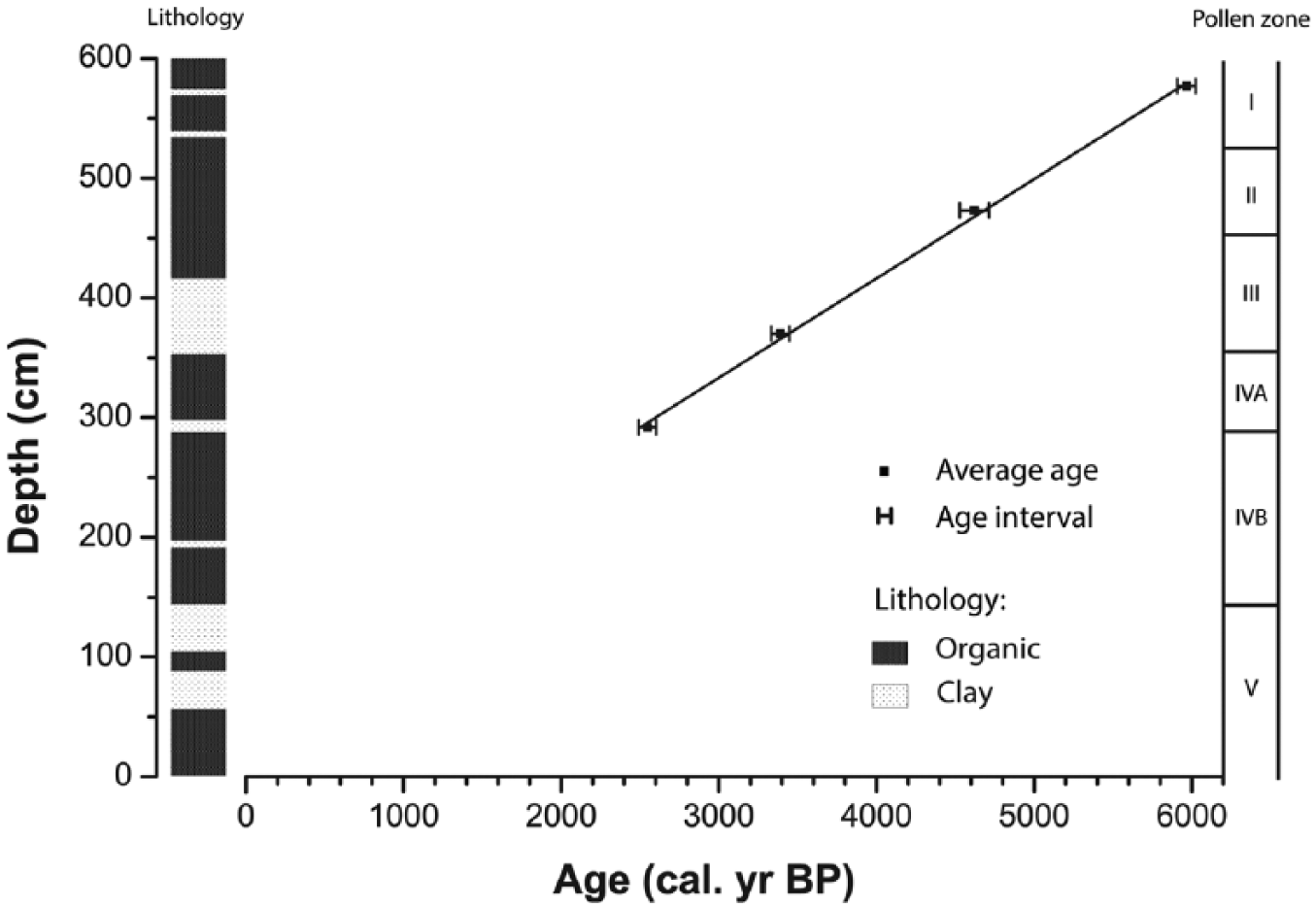
Age–depth model of calibrated radiocarbon ages.
A total of 62 pollen taxa were identified in the analysis, but only taxa reaching a percentage of at least 1% are represented in the pollen diagram (Figure 3). Changes in composition and abundances allowed CONISS to delimit five pollen zones. However, pollen zone V (145–0 cm) presented a low pollen concentration and high concentrations of silicates, which highly reduced the observation of pollen grains. Thus, changes in the pollen composition of zone V could not be inferred.
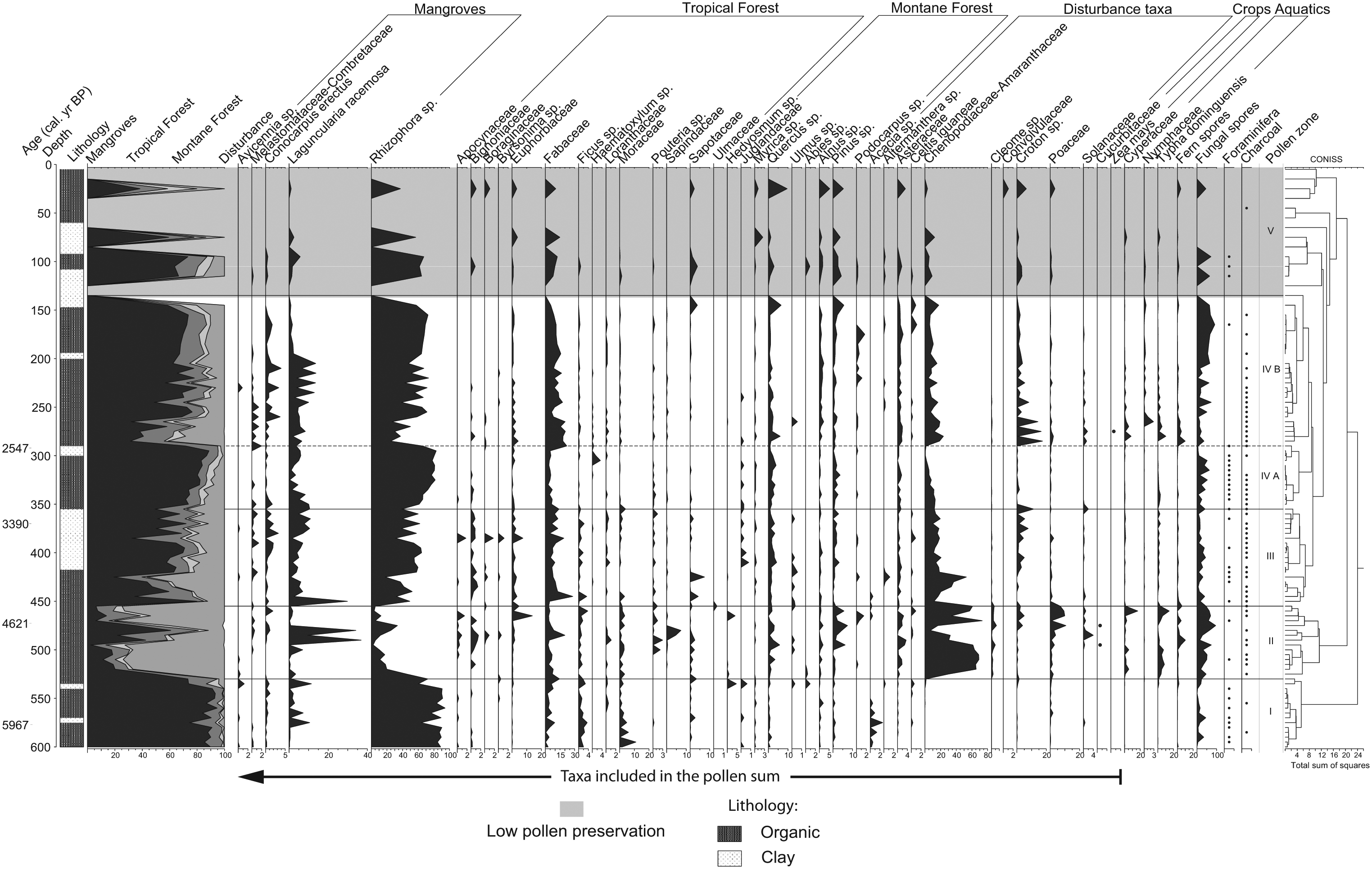
Pollen diagram and average calibrated ages for core CERRITOS02.
Zone I (600–530 cm; ~6200–5400 cal. yr BP)
Pollen from mangrove elements dominates this zone (>85%), with R. mangle constituting the dominant species (>80%). L. racemosa presents low percentages (<5%), but high peaks (7–11%) are present at 575, 565 and 535 cm. The percentages of A. germinans, C. erectus and other Melastomataceae/Combretaceae (Melasto/Comb) are low through this zone.
The pollen percentages of tropical forests taxa are relatively low (4–8%) but rises (10–18%), as shown by the counts obtained for Fabaceae, Moraceae and Ficus, at the same intervals as L. racemosa. Montane forest pollen is scarce (0–3%), being represented by the Quercus, Alnus and Pinus genera. Disturbance taxa show the lowest percentages in the entire record (0–4%). At the end of the zone, mangroves sharply decline (91–73%), with R. mangle showing the largest decrease (90–60%), while increments of tropical forest (4–12%), montane forest (0–8%) and aquatic taxa (0–1.4%) are observed.
Zone II (530–455 cm; ~5400–4500 cal. yr BP)
The initial stage of this period shows the continuing trend of declining mangroves that started in the uppermost section of zone I. The remainder of zone II presents fluctuations in pollen percentages, and presents the lowest counts for mangroves found in the entire record (2–35%; R. mangle 2–34%), except at 490 and 480 cm, where high percentages of L. racemosa occur (36% and 34%, respectively).
The high diversity and abundance of tropical forest taxa explain the higher pollen percentages observed (9–34%), where Fabaceae, Moraceae and Euphorbiaceae make the greatest contributions, along with Bignoniaceae, Ficus, Sapotaceae and Pouteria. Montane elements show slight increases in pollen concentration (up to 13%), with Quercus and Pinus occurring at higher percentages (up to 5% and 7%, respectively), while Abies occurs with low pollen counts at the onset of the zone.
Disturbance taxa present the highest percentages for the entire record (80%). Chenopodiaceae/Amaranthaceae (Cheno/Ams, up to 73%), Poaceae (up to 10%) and Cleome (up to 6%) exhibit greater contributions, although Asteraceae and Croton are also often represented. Cucurbitaceae pollen is present at 495 and 475 cm, coinciding with high percentages of disturbance taxa. Cyperaceae and Typha are present throughout all of zone II, showing fluctuating, higher percentages than in zone I (0.09–16% and 1–14%, respectively). Charcoal is presents almost continuously during this zone.
Zone III (455–355 cm; ~4500–3200 cal. yr BP)
High fluctuations in mangrove pollen percentages characterize this period. Pollen of R. mangle is represented with great variability (30–52%) along the zone, except for the period between 420 and 390 cm, where there are high stable percentages. Small oscillations in pollen percentages of L. racemosa (1–10%), Combretaceae (<2%) and C. erectus (<3%) are noted throughout this horizon, except at 450 cm, when a peak of L. racemosa pollen is observed. A. germinans is absent in this period, except at 425 and 375 cm. Tropical forest elements also exhibit high variability in pollen percentages (5–42.5%), showing the highest values in the record contrary to decreasing mangroves. The main representatives of this group are Fabaceae, Euphorbiaceae and Ficus, while Moraceae, Sapotaceae and Bignoniaceae are also frequent, but at lower percentages. Two peaks of pollen from tropical forest taxa are observed at 445 and 385 cm, in contrast to the patterns found for both mangrove elements and disturbance taxa at the same depths.
Pollen from montane forest taxa show steady low counts in this zone. Quercus is found more often and at higher percentages (up to 4%) than in zone II, while Pinus pollen is less frequent and abundant (up to 3%). Alnus pollen is present at high frequency but at low percentages. Representation of disturbance taxa shows lower and more variable values (11–32%) than in the previous zone. Cheno/Ams pollen shows a sudden increase (52%) at 425 cm, corresponding to the lowest values found for pollen from mangroves (19%). Aquatic taxa are scarce in the pollen record, but dominance of Typha over Cyperaceae is noted. Foraminifera are present at 430–415 cm, and charcoal occurs frequently.
Zone IV A (355–290 cm; ~3300–2500 cal. yr BP)
The pollen percentages of mangrove taxa increase steadily in this zone, showing higher percentages (73–88%). R. mangle is predominant (61–82%), and the percentages of L. racemosa are high from 350 to 340 cm (8–10%) but remain low (0–5%) in the rest of zone IVA. C. erectus shows a declining trend, while A. germinans disappears from the pollen record.
The percentages of pollen from tropical forests are low (4–26%) throughout the entire zone, with Fabaceae and Euphorbiaceae showing the highest representation. Moist tropical forest elements (Moraceae, Ficus and Sapotaceae) are reduced, except in a short section at 305–300 cm where Haematoxylum pollen occurs up to 3%. Pollen percentages of montane forests decline in this part of the record (2–6%), with grains of Quercus and Pinus (up to 3%, both) representing the dominant species, while Alnus pollen is found throughout this zone but at low percentages as in zone III.
A reduced abundance and frequency of disturbance taxa pollen is observed in this zone. Cheno/Ams (0.5–13%) and Asteraceae (0.5–1.5%) are the most frequent taxa, with a continuous reduction trend being found for the former group. Representation of aquatic taxa is highly reduced in this section (<3%), with alternate dominance between Typha and Cyperaceae being found. Foraminifera are recognized at depth 345–300 cm, while charcoal occurs from 350 to 320 cm.
Zone IV B (290–145 cm; ~2500–700 cal. yr BP)
A sudden decrease in mangrove elements distinguishes the lower phase (2420–265 cm) of this zone. Then, percentages of mangrove taxa fluctuate towards higher values (73%) until the end of the zone, when this variability declines. Rhizophora mangle is the dominant species in the pollen sum during this section (24–72%), and its percentages counter-oscillate with those of L. racemosa (0.5–13%). Conocarpus erectus and Melasto/Comb pollen occur during this part of the record at low percentages (<4% and <2%, respectively), coinciding with high percentages of R. mangle. Avicennia germinans is present only at 230 cm.
Tropical forest taxa display higher and more consistent percentages (9–25%) than in the previous zone. Fabaceae is the dominant species (up to 20%) and shows a declining trend towards the end of the zone. Euphorbiaceae is the second most abundant taxon, presenting higher percentages (up to 4%) in the lower part. The less frequent pollen taxa include Moraceae, Ficus, Sapotaceae, Pouteria and Loranthaceae (<1%). Pollen of montane forest taxa maintain low percentages (up to 9%), with three-mode dominance being observed among Quercus (up to 6%, 290–230 cm), Alnus (up to 2%, 225–205 cm) and Pinus (up to 2%, 195–730 cm).
High variability in pollen counts of disturbance taxa associated with relative high percentages (up to 41%) are observed from 285 to 265 cm, corresponding to high percentages of pollen from tropical forests, but contrasting with the abundance of mangroves. This variability then decreases at the upper sections. The representative taxa are Cheno/Ams, Asteraceae and Poaceae. Significant increases in counts of Croton pollen are observed at 285, 275 and 265 cm. Only a single pollen grain of Zea mays is found in the entire record, at 275 cm. Pollen grains of Cyperaceae and Typha are present in this zone, showing a decreasing trend from the onset of the zone until 235 cm, after which they are found at low percentages. Charcoal particles are observed continuously from the onset of the zone to 230 cm. Foraminifera are absent in this zone, except at 165 cm.
Environmental reconstruction
Zone I (~6200–5400 cal. yr BP; 600–530 cm) is characterized by the dominance of Rhizophora, and low percentages of tropical and montane forest elements and reduced disturbance taxa in the pollen record are indicative of a Rhizophora pure stand forest in the surroundings of the coring site. Zone II (~5400–4500 cal. yr BP; 530–455 cm) features a sharp drop in mangrove pollen percentages, dominance of disturbance taxa and increased tropical forest elements in the pollen sum, pointing out the presence of secondary vegetation associated with tropical dry-forest elements. In Zone III (~4500–3300 cal. yr BP; 455–355 cm), the recovery of mangroves is noticed, albeit of fluctuating percentages observed in the pollen record for both mangrove elements and tropical forest taxa, indicative of a highly unstable environment. Zone IVA (~3300–2500 cal. yr BP; 355–290 cm) represents the development of a Rhizophora pure stand forest denoted by steady increase in dominance of Rhizophora and low percentages of tropical forest elements and disturbance taxa in the pollen record. Zone IVB (~2500–700 cal. yr BP; 290–145 cm) is characterized in the onset by a strong descend in pollen from mangrove elements and occurrence of Zea mays, suggesting the mangrove forest was disturbed by human occupation, while unstable conditions are noticed for the rest of Zone IVB observed in the fluctuating pollen percentages of mangroves, tropical taxa and disturbance taxa.
Discussion
Middle Holocene (6200–4500 cal. yr BP; 600–455 cm)
The percentages of mangrove pollen and occurrence of Foraminifera found from ~6200 to 5400 cal. yr BP (600–530 cm) indicate a fringe mangrove forest around the coring site, dominated by pure R. mangle stands, with a high tidal influence (Medeanic et al., 2008; Pool et al., 1977; Urrego et al., 2009). The presence of pollen of C. erectus and L. racemosa may be explained by water transportation because both species are most commonly found in riverine and inland environments (Pool et al., 1977; Tomlinson, 1986; Urrego et al., 2009). Despite the low pollen percentages obtained for A. germinans, we cannot rule this species out as a main taxa simply through pollen analysis, as this species presents low pollen production (Tomlinson, 1986) and often occurs in a patchy distribution (Urrego et al., 2009).
This episode of ecosystems favouring mangroves corresponds to episodes of rising sea levels, high precipitation and warm temperatures reported for the Panama Basin (11,300–5600 cal. yr BP; González et al., 2006), the Cariaco Basin (10,500–5400 cal. yr BP; Haug et al., 2001), northern Mexico (middle-Holocene to ~4000 cal. yr BP; Metcalfe et al., 2000) and the lowlands of northern Guatemala (~8000–4500 cal. yr BP; Mueller et al., 2009). In southern Chiapas, dominance of mangrove forests is observed from 6600 to 4100 cal. yr BP (Kennett et al., 2010). Neff et al. (2006b) reported that high precipitation and high forest cover occurred at ~6000 cal. yr BP in the north Pacific region of Guatemala, with a drying trend being observed from ~5850 to 5000 cal. yr BP, although conditions were sufficiently humid to support mature rainforests. The increased humidity and temperature during this period are related to the increased insolation reported for the Caribbean region (Leyden, 2002), forcing the ITCZ to move northward, increasing precipitation over Mesoamerica (Haug et al., 2001) and ultimately suppressing ENSO events (Roy et al., 2010). The latitudinal extent of this events points to an orbital-induced insolation event as the causative factor underlying the warm-humid events of the middle-Holocene (Clement et al., 2000).
The low percentages of pollen from montane forests and the dominance of Quercus pollen in these forests from ~6200 to 5360 cal. yr BP (600–530 cm) support the existence of regional warm and moist conditions. Rodgers and Horn (1996) and Bush (2000) warn about the misinterpretation of Quercus pollen in the lowlands, as Quercus oleoides is present at a range of 50–500 m.a.s.l.; Horn, 1985). Quercus forests are reported over 700 m.a.s.l. on the Pacific slope of the Sierra Madre Oriental (INE, 1999a, 1999b; Martínez-Meléndez et al., 2008; Pérez-Farrera et al., 2012; Salas et al., 2007;). Thus, the Quercus pollen found out-of-range in our record is better explained by wind dispersion because this genera is one of the few wind-pollinated montane species (Islebe and Hooghiemstra, 1995; Jacobs, 1982).
Starting at ~5300 cal. yr BP (525 cm; zone II), the sharp decreases in pollen of mangrove elements, increases in pollen from tropical forests and lack of Foraminifera suggest that the environment rapidly changed into an inland mangrove (Urrego et al., 2009). The increment of disturbance taxa pollen could also reflect the expansion of drier environments, such as tropical sub-deciduous forests and secondary vegetation, as the pollen from these environments mainly consists of Cheno/Ams, Asteraceae and Poaceae (Jacobs, 1982). Additionally, Amaranthaceae are representative herbs of tropical sub-deciduous and thorn forests the Pacific coast of Chiapas and Oaxaca (Martínez-Meléndez et al., 2008; Pérez-Farrera et al., 2012; Salas et al., 2007). Higher counts of montane forest pollen and dominance of Pinus indicate regional reductions in temperature and most likely less precipitation, accounting for the more open vegetation (Domínguez-Vázquez et al., 2004). In open vegetation forests, erosion because of runoff is more frequent, which could explain the increased occurrence of tropical forest pollen in the sediments, coupled with higher counts of disturbance taxa. The abundance of tropical forest elements and disturbance taxa versus mangrove forests in the pollen record also suggests periods of decreasing sea-level rise, allowing succession to occur in areas that were previously occupied by mangroves (González et al., 2006). The increase in Cyperaceae and Typha pollen observed from ~5300 to 4500 cal. yr BP (525–455 cm) could be interpreted as indicating the growth of salt marshes derived from rising sea levels and increased freshwater inputs (Kennett et al., 2010; Urrego et al., 2009). However, the higher percentages of these taxa could be better explained by basin geomorphology, as the decreasing water level in response to reduced humidity and the receding sea shore could have increased the availability of environments with different salinities (Bush, 2002). The nearly continuous occurrence of charcoal and the presence of Cucurbitaceae pollen at ~4900 and ~4700 cal. yr BP (495–475 cm) evidence the occurrence of agricultural activities in the mangrove area. It seems very likely that a combination of dryer conditions, decreasing sea-level rise and human activities was responsible for the minimum percentages of mangrove taxa and maximum percentages of disturbance taxa observed in our record.
The middle Holocene was characterized by a strong decrease in the sea-level rise from ~7700 to 4800 cal. yr BP. Mangrove ecosystems impacted by this phenomenon have been reported in the Caribbean region (Toscano and Macintyre, 2003), the Sibun River in Belize (Monacci et al., 2011), the coast of Oman (Lézine et al., 2002) and Bangladesh (Rashid et al., 2013). In addition, the late middle Holocene presented periods of decreased humidity. González et al. (2006) reported a phase of decreases in mangrove taxa in the Panama Basin at 6500 cal. yr BP, coinciding with a dry event reported for South America. A dry episode occurred in Central Mexico from 6000 to 5000 cal. yr BP, although much variability exists among sites (Metcalfe et al., 2000). Precipitation decreases in the Cariaco Basin at ~5000 cal. yr BP (Haug et al., 2001). Mayewski et al. (2004) observed a global-scale ‘cold poles, dry tropics’ event at 6000–5000 cal. yr BP, likely caused by decreased solar radiation. In Lake Tzib, on the Yucatan Peninsula, lower precipitation has been inferred to have occurred from 6500 to 4700 cal. yr BP (Carrillo-Bastos et al., 2010). In North Pacific Guatemala, a higher aridity index has been recorded from 5270 to 4750 cal. yr BP, with arboreal taxa being reduced to a minimum at 4750 cal. yr BP, together with a dominance of Cheno/Ams (Neff et al., 2006b).
Farming has been conducted in Pacific Central America since the middle Holocene. Evidence suggests that Cucurbitaceae have been grown along the coast of Chiapas since ~6500 cal. yr BP, together with other crop plants (Kennett et al., 2010). The occurrence of agricultural activities was inferred from the charcoal concentrations found in swamps of Pacific Guatemala at three different sites intermittently around 5500 cal. yr BP (Neff et al., 2006c). Kennett et al. (2010) attribute the high percentages of Cheno/Ams and Asteraceae observed at 4200–3900 cal. yr BP to human activities near our study site.
Sudden increases in L. racemosa are observed at ~5900, 5800, 5400, 4900, 4800 and 4400 cal. yr BP (575, 565, 535, 490, 480 and 450 cm), suggesting short periods (~60 years) of droughts or reduced tidal influence, since L. racemosa can tolerate higher salinity and desiccation than R. mangle (Gallo and Rodríguez, 2010). These episodes are in close relation with ENSO events recorded in lake sediments of south-eastern Ecuador and the Galápagos islands (Conroy et al., 2008; Moy et al., 2002; Figure 4). High percentages of L. racemosa pollen recorded could result from this species colonizing disturbed areas, forming pure stands (Tomlinson, 1986) after ENSO-related droughts or floods, leading to the best representation of Laguncularia pollen in the sediments.

Climatic reconstruction of ENSO-related precipitation as compared with abundance of main mangrove elements in CERRITOS02. (a) Percent of sand, (b) silt and (c) clay in a core from El Junco lake, San Cristobal, Galápagos (Conroy et al., 2008). Increasing percentages of sand combined with high percentages of silt are indicative of El Niño; increasing percentages in sand content combined with high percentages of clay indicates La Niña conditions. (d) Percent of titanium in a deep-sea core from Cariaco Basin (Haug et al., 2001). (e) Red colour intensity (110-point smooth of the raw red intensity record) in a core from Laguna Pallcacocha, southern, Ecuador (Moy et al., 2002). Data downloaded from ncdc.noaa.gov/data-access. (f) Pollen percentage of main mangrove elements in CERRITOS02, this study. Light-green shade represents the ENSO-ITCZ coupling. Dark-green shade is indicative of La Niña–dominated episode. Grey shading shows El Niño–dominated state.
Rapid episodes of freshwater inputs were reported by Kennett et al. (2010) from core SOC05-2 between 6600 and 4100 cal. yr BP in a study conducted nearby, although the ages are dissimilar. Elemental data for the Pacific Coast of Guatemala indicate higher precipitation after 4940 cal. yr BP, with tropical forests recovering after 4750 cal. yr BP (Neff et al., 2006b). In Lake Tzib, 18O records show increased precipitation during a short period at ~5000 cal. yr BP within a longer dry phase (Carrillo-Bastos et al., 2010). These records demonstrate the presence of brief periods of augmented moisture availability during the middle Holocene.
Late Holocene (~4500–0 cal. yr BP; 455–84 cm)
The lack of extended variations in the pollen composition since ~4500 cal. yr BP (455 cm) suggests the establishment of the present shoreline at that date. Differences in the rates of sea-level rise among sites will depend on regional (i.e. geomorphology) and local factors (i.e. deltaic processes; Ellison and Stoddart, 1991; Soares, 2009). The recovery of Rhizophora and Laguncularia mangroves observed in the pollen record at ~4400 cal. yr BP (450 cm) indicates increased sea levels, combined with higher precipitation, enhancing sediment input to the basin and additional spaces for mangrove establishment therefore became available (González et al., 2006). The dominance of Quercus pollen and reduced percentages of Pinus and Alnus pollen from the upper slopes supports the existence of regional warm-humid conditions (Domínguez-Vázquez and Islebe, 2007; Metcalfe et al., 2000) from ~4500 to 3200 cal. yr BP (455–350 cm). At ~4300 and 3700 cal. yr BP (445–385 cm), pollen of tropical forest taxa show an opposite pattern to disturbance taxa and mangroves, suggesting increased moisture. A stable period of mangrove growth from ~4000 to 3600 cal. yr BP (420–380 cm), together with the occurrence of Foraminifera and decreases in aquatic elements, is indicative of marine introgressions.
Contrasting climate conditions have been described for the beginning of the late Holocene by different authors. A less extensive period of ‘cold poles, dry tropics’ from 4200 to 3800 cal. yr BP has been observed at several sites around the globe (Mayewski et al., 2004). In Lake Cobá, reduced fluctuations in precipitation have been reported based on the presence of pollen from mesic dry-forest taxa around 4700 cal. yr BP (Leyden, 2002). Mueller et al. (2009) suggest that a dry period occurred from 4500 to 3000 cal. yr BP in the Petén of Guatemala. At Lake Tzib, increased precipitation has been inferred from 4600 to 4100 cal. yr BP, followed by a dry episode at 3500 cal. yr BP (Carrillo-Bastos et al., 2010). Aragón-Moreno et al. (2012) reported that there was sufficient precipitation to support a moist tropical forest from 4500 to 3000 on the northern area of the Yucatan Peninsula, with a drying trend beginning at 3500 cal. yr BP. Records from Puerto Morelos show higher values of mangrove elements and tropical forest taxa, suggesting the occurrence of a sustained episode of higher humidity than at present at 3600–3300 cal. yr BP (Torrescano-Valle and Islebe, 2012). Neff et al. (2006b) reported humid conditions for northern Pacific Guatemala from 4750 to 4300 cal. yr BP, after which reductions in organic carbon and increased carbonates observed from 4200 to 4000 cal. yr BP may suggest marine introgression. These authors also reported brief dry events at 4300 and 4100 cal. yr BP, with the driest event being observed after 4000 cal. yr BP (Neff et al., 2006b). The results are also corroborated by records from southern Chiapas, where a change from a brackish- to a freshwater-dominated environment is observed at 4700–4200 cal. yr BP, and the recovery of mangroves at 4200–3900 cal. yr BP suggests slightly marine-influenced conditions (Kennett et al., 2010).
A transition towards steady and more favourable conditions for mangrove development was established at ~3200 cal. yr BP (350 cm). Increasing percentages of R. mangle pollen and the presence of Foraminifera suggest a transition to brackish environments. A slight reduction of tropical forest pollen is observed, suggesting the existence of reduced moisture with sufficient precipitation to allow the persistence of tropical forests. Reductions in pollen of montane elements suggest increasing temperatures, although elements from colder areas (Pinus and Alnus) persist at low values, indicating the presence of these forests at higher altitudes.
The almost continuous occurrence of charcoal particles from ~4400 to 2800 cal. yr BP (450–320 cm) in the sedimentary record strengthens the evidence that agricultural activities were carried out in the area (Kennett et al., 2010). Nevertheless, the lack of crops pollen during this time demonstrates that such human activities were conducted outside the mangrove area. The absence of human disturbance during a long period could allow mangrove ecosystems to recover.
At 2400–2200 cal. yr BP (285–265 cm), the decreases in mangrove pollen and increased percentages of pollen from disturbance taxa and moist tropical forests could suggest decreased precipitation. However, the presence of Zea mays pollen at ~2300 cal. yr BP (275 cm), together with the continuous occurrence of charcoal (~2500–1800 cal. yr BP, 290–230 cm) and increments in the pollen of disturbance taxa such as Croton, are indicative of agricultural activities near the mangrove area because disturbance taxa are well represented in human environments (Leyden, 2002). The dominance of oak pollen from montane forests suggests the existence of a regional warm-humid climate over a longer period (3200–2500 cal. yr BP, 350–295 cm).
Our results correspond to those reported for the Gulf of Mexico, the Yucatan Peninsula and the Pacific Coast of Chiapas and Guatemala. Leyden (2002) reported a mangrove–brackish–freshwater transition from 3400 to 3150 cal. yr BP in coastal Tabasco, related to increased moisture and a reduced sea-level rise. The pollen record from Lake Tzib indicates higher precipitation at 3000–1900 cal. yr BP and the development of a medium-stature tropical forest (Carrillo-Bastos et al., 2010). In a study performed near our site, Kennett and Voorhies (1996) observed stable temperatures and precipitation from 3000 to 1800 cal. yr BP, similar to present-day conditions. Stable humid conditions are reported from 2800 to 1900 cal. yr BP for Pacific Guatemala, with reduced moisture being observed from 2600 to 2400 cal. yr BP (Neff et al., 2006b). This short episode of reduced moisture during a longer humid episode could have facilitated agricultural activities (Mueller et al., 2009).
Regional shifts towards lower temperatures and slightly dryer conditions are inferred from ~1600 to 1500 cal. yr BP (220–205 cm) based on changes in pollen abundance of montane forests from Quercus- to Alnus-dominated. Alnus occurs in colder conditions than Quercus at upper latitudes (Islebe and Hooghiemstra, 1995) and is an opportunistic species in disturbed montane forests (Weng et al., 2004). At 1500–1000 cal. yr BP (210–165 cm), the pollen record suggests that the montane forest became Pinus dominated, indicating cooler and dryer conditions. Additionally from ~1500 to 700 cal. yr BP (205–145 cm), the percentages of Rhizophora pollen were lower than in the two previous zones with reductions also in pollen of L. racemosa along with tropical forest elements, and a sustained presence of pollen from disturbance taxa can be explained as a result of decreasing moisture availability in the lowlands.
The declining frequency of charcoal starting at ~1700 cal. yr BP (225 cm) indicates human activities progressively reduced in the region. Climate instability could be responsible for the abandonment of human settlements in the mangrove area, although agricultural activities could still have occurred on the coastal plain. As regional drought conditions persisted after ~1460 cal. yr BP (205 cm), abandonment of human settlements occurred in more extensive areas.
Dry conditions at 1200 cal. yr BP have been reported in tropical and subtropical areas (Mayewski et al., 2004), coinciding with the Classic Mayan Collapse. In Lake Chichancanab, a dry trend has been observed from 3800 to 1400 cal. yr BP, although date corrections had to be applied because of hard water effects (Leyden, 2002). A drying trend has been suggested at Lake Tzib from 1300 to 1200 cal. BP, with a higher peak occurring at 1200 cal. yr BP, although dry conditions occurred until 850 cal. yr BP (Carrillo-Bastos et al., 2010). Records from Punta Laguna indicate a dry event from ~1700 to 840 cal. yr BP, with a higher intensity being observed from ~1170 to 840 cal. yr BP (Curtis et al., 1996). Significant dry events have been reported for Puerto Morelos from ~1380 to 1180 cal. yr BP, after which humid conditions were observed from ~1180 to 890 cal. yr BP. From ~890 cal. yr BP to the present, dry conditions have been suggested (Islebe and Sánchez, 2002). The record from San José Chulchaca on the Yucatan peninsula shows the driest period reported for this area occurring at 1500–900 cal. yr BP (Metcalfe et al., 2000).
Several peaks in pollen from tropical forest taxa coincide with high percentages of disturbance taxa pollen, suggesting short periods (~60 years) of reduced humidity within the periods of ~4500–3200 cal. yr BP and ~2400–700 cal. yr BP. These rapid dry events are coincident with ENSO events reported for either Laguna Pallcacocha (Moy et al., 2002) or El Junco lake (Conroy et al., 2008). We suggest that ENSO-related droughts are responsible for these events.
Two oscillating phases with decreasing percentages of mangrove pollen are observed in CERRITOS02. The first phase, at ~3700–3200 cal. yr BP (Zone III), could be explained by the increased variability and frequency of ENSO events started after ~4000 cal. yr BP (Moy et al., 2002) combined with a drying trend initiated earlier at ~4200 cal. yr BP (Haug et al., 2001). Pollen and geochemical proxies from Lake Petén Itzá record a drying trend from ~4500 cal. yr BP (Mueller et al., 2009). A La Niña–dominated period is observed in El Junco lake at ~3800–2800 cal. yr BP (Conroy et al., 2008). This interval of La Niña–dominated conditions may have stopped reef accretion at ~3800–3200 cal. yr BP in the Tropical Eastern Pacific (Toth et al., 2012).
The second phase, observed at 1900–1500 cal. yr BP (Zone IVB), can be interpreted as a result of unstable conditions during an El Niño–dominated period reported by Conroy et al., (2008). Reduced frequency of ENSO events at this period (Moy et al., 2002) in conjunction with a stable northern position of the ITCZ (Haug et al., 2001) relative to Zone III (this study) could be responsible for lesser reduction in mangrove elements observed in CERRITOS02. However, sedimentological proxies evidence the occurrence of stronger ENSO events starting at circa 2000 cal. yr BP in the Peruvian coast and El Junco lake (Conroy et al., 2008; Rein, 2007). Thus, we suggest that intense and dry/warm and cold/humid phases characteristic of strong ENSO events may be responsible for increased variability observed in mangroves abundance and composition in CERRITOS02 (Figure 4). Clay deposits found at Lake Petén Itzá indicate unstable climate conditions around 1800–1000 cal. yr BP (Mueller et al., 2009). The coast of Pacific Guatemala presented dry and variable conditions from 1300 to 480 cal. yr BP in the north (Neff et al., 2006b) and from 1100 to 600 cal. yr BP in the south (Neff et al., 2006a). All of these records demonstrate the high variability of climate in the region during the late Holocene.
Causes of environmental change on the Pacific coast
According to Curtis et al. (1996), changes in precipitation have been the dominant factor during the Holocene. These changes are related to the movements of the ITCZ, which is found in northern latitudes during humid periods of the boreal hemisphere and southern locations in arid periods (Haug et al., 2001). Simulation models associate a reduction in the frequency and intensity of ENSO events in the middle- to late Holocene with reduced insolation during the boreal summer as a result of orbitally induced changes (Clement et al., 2000). According to Mayewski et al. (2004), decreased insolation does not account for the dry events reported from 4300 to 3800 and from 1200 to 1000 cal. yr BP in the tropics. However, wide oscillations of the ITCZ during increased frequency and higher strength of ENSO events suggest a coupling of both systems at ~3800–2800 cal. yr BP (Conroy et al., 2008), traduced in extreme dry and floods during this period. After ~2500 cal. yr BP, the increased frequency and strength of ENSO events and the lack of variability in the ITCZ suggest a decoupling of the ITCZ variability with ENSO events (Conroy et al., 2008). Maximum frequency and high strength of ENSO events after ~2000 cal. yr BP are evidenced in different records from Andean Ecuador, the Galápagos and the Peruvian Coast (Conroy et al., 2008; Moy et al., 2002; Rein, 2007). These records agree with observations of the Zebiak and Cane model, reinforcing the notion that highest frequency of ENSO events (and associated extreme droughts and floods) could be a result of orbitally induced changes in insolation (Clement et al., 2000). The dry episodes during the late Holocene may also depend on the strength and location of the Bermudas High (Metcalfe et al., 2000), which restricts the northern movement of the ITCZ, and on variations in the atmospheric and oceanic circulation (Curtis et al., 1996).
Agriculture in Mesoamerica evolved from simple ‘peasant’ agriculture to complex ‘stratified’ agriculture, permitting the development of stratified societies based on maize cultivation. The removal of forests and slash and burn activities are associated with agriculture and urban construction and have heavily impacted natural landscapes during the Holocene (Roberts, 1998). Evidence from pollen and charcoal found in CERRITOS02 suggests that to increase the crop area, earlier settlers modified the mangrove landscape during different episodes in South Pacific Mexico. The detection of past agricultural activities in the mangroves of coastal Chiapas is difficult because of various factors. First, evidence from different sites in the mangroves of the Pacific coast of Guatemala suggests that human occupation was extensive in the area, but at low population levels, movements to another area occurred when resources were depleted (Neff et al., 2006c). Second, few areas near mangroves are able to sustain crop cultivation at present (Kennett and Voorhies, 1996), mainly because of a lack of hydrological control (Prager, 2006). Third, the morphology and weight of pollen from crops (Zea mays and Cucurbitaceae) restrict the representation of these taxa in sediments, even in the proximity of crop fields (Kennett et al., 2010). The scarcity of direct evidence of farming activities at our study site also agrees with the statement that the extremely high variability in the environment explains the occurrence of small populations and the lack of permanent establishment in the mangrove area (Neff et al., 2006c).
Conclusion
The lack of major river flows and intense tides at our coring site is responsible for the high sedimentation and lack of significant changes in sedimentation rates at the site, leading to a good depth–age correlation in the record. Core CERRITOS02 indicates changes in vegetation for the time frame between ~6200 and 700 cal. yr BP and provides evidence of environmental changes affecting mangrove vegetation on the coast of Chiapas. The wettest and warmest phase is observed for the middle Holocene, related to high rates of sea-level rise, favouring the development of R. mangle forests. In contrast, intense effects of reduced precipitation, an abrupt slowing of the sea-level rise and human impacts related to agriculture may account for the lowest proportion of mangrove pollen observed in our record at ~5400–4500 cal. yr BP. The late Holocene is recognized by the recovery of mangroves in pollen taxa at ~4400 cal. yr BP, suggesting increased moisture and the establishment of the current coast line. Beginning at 3200 cal. yr BP, a dry phase is observed, allowing marine introgression and the development of a brackish environment. The absence of strong human activities associated to ENSO variability in the mangrove area from ~4400 to 2900 cal. yr BP could have facilitated the recovery of mangroves. Intense agricultural activities within the mangrove area are suggested for the period from ~2500 to 1600 cal. yr BP, while regional moist conditions are inferred from the presence of pollen from montane taxa. Drying trends are observed from ~1600 to 700 cal. yr BP, characteristic of late Holocene conditions. These events are observed throughout the entire record, although a trend towards more frequent and less intense events is noted starting at the beginning of the late Holocene. A rapidly changing environment could have prevented high concentrations of human settlement in the area (Neff et al., 2006c), thus facilitating the interpretation of climate events in our results.
In general, our data indicate that mangroves from coastal Chiapas are a good indicator of past environmental and climate changes at local and regional scales. However, modern pollen rain studies are desirable for better interpretation of the vegetation types coupled with different sets of environmental conditions. High-resolution multi-proxy surveys are suggested for corresponding regional and local climate events that have occurred in the area as well as for the precise recognition of human activities at different sites in the upper basin.
Footnotes
Acknowledgements
We thank Henry Hooghiemstra and an anonymous reviewer for helpful criticism of an earlier version of this paper. We also thank Cristian Tovilla-Hernández, Alicia Carrillo-Bastos, Luz Verónica Gutiérrez Ayala, Alejandro Antonio Aragón-Moreno and Alejandro Vela Peláez for their support and observations. We would also like to thank to Margarito Tuz-Novelo and Gerardo De la Cruz Montes for their help during the field work. Finally, we thank ECOSUR – Chetumal for providing facilities.
Funding
This work was supported by the CONACYT grants program (CVU 384724); and the CONACYT research program ‘Variabilidad climática y paleoecológica de los últimos 1,500 años del sureste de México’.