
Abstract
We provide the first fossil pollen and charcoal analysis from the island of Gran Canaria (Canary Islands). The pollen record obtained from Laguna de Valleseco (870 m a.s.l.) spans the late Holocene (c. 4500–1500 cal. yr BP) and thereby captures the impact of human colonization. During the earliest period, pollen composition resembled contemporary thermophilous communities, with palms (Phoenix canariensis) and junipers (Juniperus cf. turbinata) being the dominant trees, suggesting that these elements were more widespread in the past. Vegetation in Valleseco began to change at around 2300 cal. yr BP, 400 years before the earliest archaeological evidence of human presence in the island (c. 1900 cal. yr BP). Our data show an increased frequency of fires at that time, coinciding with the decline of palms and the increase of grasses, indicating that humans were present and were transforming vegetation, thus showing that the demise of Gran Canaria’s forest began at an early point in the prehistoric occupation of the island. In the following centuries, there were no signs of forest recovery. Pollen from cultivated cereals became significant, implying the introduction of agriculture in the site, by 1800 cal. yr BP. The next shift in vegetation (c. 1600 cal. yr BP) involved the decrease of grasses in favour of shrubs and trees like Morella faya, suggesting that agriculture was abandoned at the site.
Introduction
In recent decades, palaeoecological studies have been undertaken on islands worldwide. Holocene vegetation history has been outlined for some of the best-known islands, including the British Isles (e.g. Bennett, 1988), Galápagos (e.g. Froyd et al., 2014), Hawaii (e.g. Burney et al., 1995), Japan (e.g. Hayashi et al., 2010), Madagascar (e.g. Virah-Sawmy et al., 2009), New Zealand (e.g. McGlone and Bathgate, 1989) and Sicily (e.g. Sadori and Narcisi, 2001). Each island presents the opportunity to study vegetation history in the context of specific geological, climatic and human settlement conditions, providing informative comparisons across different islands within or among archipelagos. In addition, as a result of their geographic characteristics, relatively small size, short geological life cycle, and isolation, island biotas are peculiar and vulnerable, especially when it comes to oceanic islands, that is, those that have never been connected to continents (Whittaker and Fernández-Palacios, 2007), which makes the study of long-term vegetation responses in island settings of considerable practical conservation interest (Froyd and Willis, 2008). Moreover, remote islands have typically been colonized only recently relative to the continents and thus provide palaeo-records permitting analyses of climatic and geological drivers within the Holocene without the complication of anthropogenic influences, in contrast to less remote islands, which typically were colonized much earlier (Bottema and Sarpaki, 2003; Reille et al., 1999).
The Canary Islands have been recognized as a valuable model system in island ecology and biogeography (Whittaker and Fernández-Palacios, 2007), but until recently their palaeoecological history has been largely unknown. An obvious limitation is the general lack of depositional environments suitable for fossil preservation, although macrofossils such as plant imprints and charred timber can be found encased by volcanic materials and have been explored particularly in Gran Canaria (Anderson et al., 2009; García-Talavera et al., 1995; Marrero, 2014; Schmincke, 1968). Recently, our group has begun a programme of palaeoecological research analysing fossils from several former waterlogged sedimentary sequences described in historical records (e.g. Criado, 2002) or suggested by toponymy. Pollen records from one lake on Tenerife (De Nascimento et al., 2009) and a second temporary lake on La Gomera (Nogué et al., 2013) have revealed changes in forest composition on each of these islands during the Holocene, which indicate both natural and anthropogenic drivers of vegetation change over the last several thousand years. This paper contributes a third core, from a third Canarian island, providing novel insights into the environmental history of the archipelago.
On La Gomera, a major climatically driven change in vegetation occurred around 5500 years ago. Typically hygrophilous trees dominant in the vegetation around the core site (a seasonal pond within a small crater) were replaced by common species from the laurel forest, which currently dominate the area. This change was interpreted as indicating a shift to generally drier conditions, associated with a regional climatic shift across nearby parts of Africa (Nogué et al., 2013). The core from Tenerife spanned only the last 4700 years, and within it we detected a later change, about 2000 years ago, suggesting that humans had colonized and had already become major drivers of vegetation change. A transition was underway from a mixture of forest species with no modern-analogue, including trees from the laurel forest, the thermophilous woodlands, and two genera that were not considered native to the Canaries (Carpinus and Quercus), to a forest resembling the current laurel forest types (De Nascimento et al., 2009). In addition to the differences in temporal extent between both records, the disparities in the results obtained could be linked to the characteristics of each study site (e.g. elevation, wind exposure) and the history of human occupation on each island, although the timing of colonization is unfortunately poorly constrained. The former lake of La Laguna (Tenerife) was located at 550 m a.s.l. (above sea level) while the elevation of Laguna Grande (La Gomera) is 1250 m a.s.l., together representing, respectively, the lower and upper limits of the present laurel forest distribution. Moreover, radiocarbon dates from archaeological sites suggest that Tenerife could have been one of the earliest islands in the archipelago to be colonized by humans around 2800–2500 years ago (Atoche, 2008), whereas on La Gomera the oldest date obtained so far indicating human presence is 1800 years ago (Arnay-de-la-Rosa et al., 2009).
The island of Gran Canaria is one of the largest, highest, oldest and most geologically complex within the Canarian archipelago, hosting all main Canarian ecosystems from coast to peak, except for the summit scrub, which is only present on the higher islands above 2000 m a.s.l. The vegetation is not pristine, and it is estimated that all vegetation types have been reduced to less than 30% of their original area within the island (Del Arco et al., 2010). The laurel forest is the most reduced of the major ecosystem types, as less than 1% of its estimated original distribution remains, which is the lowest proportion of any Canarian island (Del Arco et al., 2010). The reduced extent of this and other vegetation types occurring on the island has generally been attributed to the intensive exploitation of natural resources occurring during the historical period (Naranjo, 2002; Parsons, 1981; Santana, 2001), that is, since the island conquest by Castilians in 1483 CE (Common Era) (Abreu Galindo, 1977). However, recent studies of archaeological sites indicate that the aboriginal inhabitants of Gran Canaria practised intense use of forest resources and that agriculture was widespread in prehistoric times (Machado, 2007; Morales et al., 2009, 2014). According to archaeological evidence and early historical descriptions, there may have been a dense population in the centuries prior to the European conquest, indicating that the island population had experienced a progressive growth (up to several tens of thousands) (Santana, 1992; Velasco, 1998). This suggests that humans may have had a significant impact on the environment long before the conquest of the 15th century, but data in support of this have hitherto been lacking. Apart from human activities, natural factors, such as climate change or volcanic activity, may also have contributed to shape the vegetation of Gran Canaria in the past. For instance, several studies have concluded that during the late Holocene, climate became more arid on several islands of the archipelago (e.g. Nogué et al., 2013; Von Suchodoletz et al., 2010; Yanes et al., 2011), while the north of Gran Canaria was affected by several Holocene volcanic eruptions, the last phase being dated to 3200–1900 years ago (Rodriguez-Gonzalez et al., 2009).
We present the first record of fossil pollen and charcoal from the island of Gran Canaria, aiming to provide a first insight into vegetation and linked environmental changes during the late Holocene. Our site, Laguna de Valleseco, was a shallow, temporary crater lake. Its location at 870 m a.s.l. is intermediate between our core sites on Tenerife and La Gomera and is at an elevation thought to be within the past natural range of the laurel forest in Gran Canaria (Del Arco et al., 2006). The resulting record spans the period of human colonization of the island and provides both a pre-human baseline and an assessment of the relative impact of anthropogenic activities in driving vegetation change prior to the 15th-century conquest. We discuss our findings in the context of our previous work from Tenerife and La Gomera, thus providing a gradually improving sketch of the Holocene vegetation history of the Canaries.
Site description
The island of Gran Canaria is in a central position within the Canary Islands. Its oldest rocks date to c. 14.5 Ma, making it the third oldest island (Carracedo et al., 2002) as well as the third in size (1561 km2), elevation (1949 m a.s.l.) and distance to the African continent (196 km). All these characteristics combine to give the island its high topographical complexity and extraordinary diversity. Gran Canaria hosts around 4800 terrestrial native species (including animals, vascular plants, bryophytes and fungi) and presents the highest percentage of endemism in the archipelago (27% and 14% of Canarian and single island endemics, respectively) (Martín, 2010). Its high elevation allows the main Canarian zonal ecosystems to be represented, except for the summit scrub. The areas of remaining more-or-less intact native vegetation cover have been significantly reduced (Table 1): the coastal shrubland is restricted to few coastal locations, ravine slopes and cliffs; the thermophilous woodlands (including Olea cerasiformis and Pistacia spp. woodlands) are significantly represented compared with other islands, but there are only a few isolated trees of Juniperus turbinata; small remnants of the evergreen laurel forest occur in the most humid northern valleys and ravines; while the Canarian pine forest is the best-represented forest, although this has in part been recovered by reforestation activities. Other characteristic non-zonal vegetation types are hygrophilous communities associated with ravine beds: Canarian willow and palm groves. However, the most widespread vegetation types are secondary communities like the Euphorbia scrubland occupying the original area of coastal shrublands and thermophilous woodlands, and the broom scrub, dominated by Teline, Adenocarpus and Chamaecytisus species, which replace the evergreen laurel forest and the pine forest where the dominant trees have been removed (Del Arco et al., 2006).
Main vegetation types present in Gran Canaria and extent of their current area, percentage remaining of their estimated potential area, elevational range and main plant species. (Based on Del Arco et al., 2006, 2010).
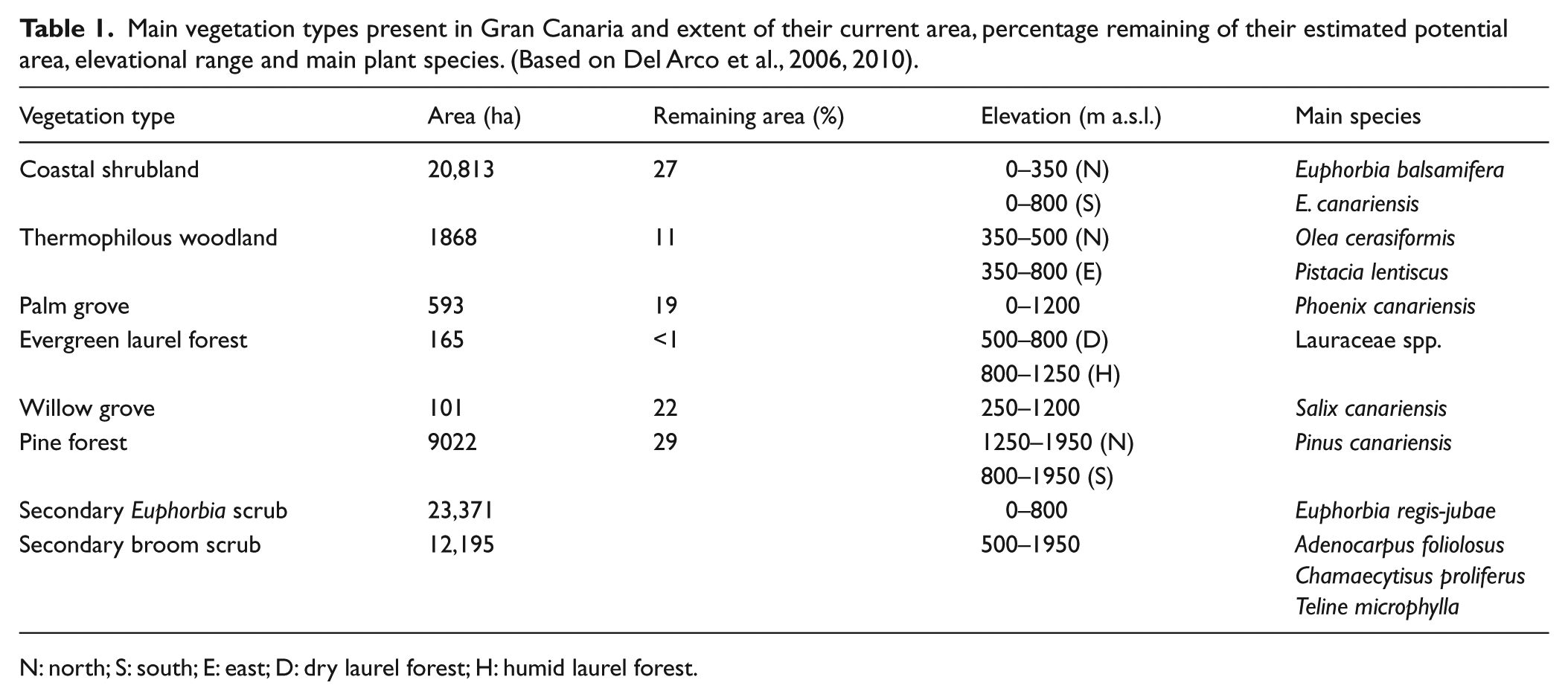
N: north; S: south; E: east; D: dry laurel forest; H: humid laurel forest.
Laguna de Valleseco is on the northern part of the island (28R 444535E, 3104573N), at an elevation of 870 m a.s.l. It is a 5-ha crater of an old volcano, probably of Pleistocene age (Guillou et al., 2004), surrounded by the ravine of Azuaje on its west side and the valley of Teror to the east (Figure 1). The site is under the influence of the ‘sea of clouds’ that forms below the trade wind inversion zone on the northern slopes of the island, where the average precipitation is around 850 mm and the annual mean temperature is 14°C (Gobierno de Canarias, 2010). Laguna de Valleseco is currently used as a recreational area, and lies within the Rural Park of Doramas, which was designated in 1994 (Canarian Network of Natural Protected Areas). The landscape has been highly transformed and the current vegetation comprises Castanea sativa and Pinus radiata plantations, Adenocarpus foliolosus and Chamaecytisus proliferus secondary scrubs, and several species from the laurel forest (Arbutus canariensis, Bencomia caudata, Erica arborea, Laurus novocanariensis and Morella faya) that have been planted as part of a restoration programme carried out in the area between 2001 and 2003 (LIFE project Laurisilva XXI), which also involved the installation of an artificial pond for bird watching within its basin (Gobierno de Canarias, 2010).
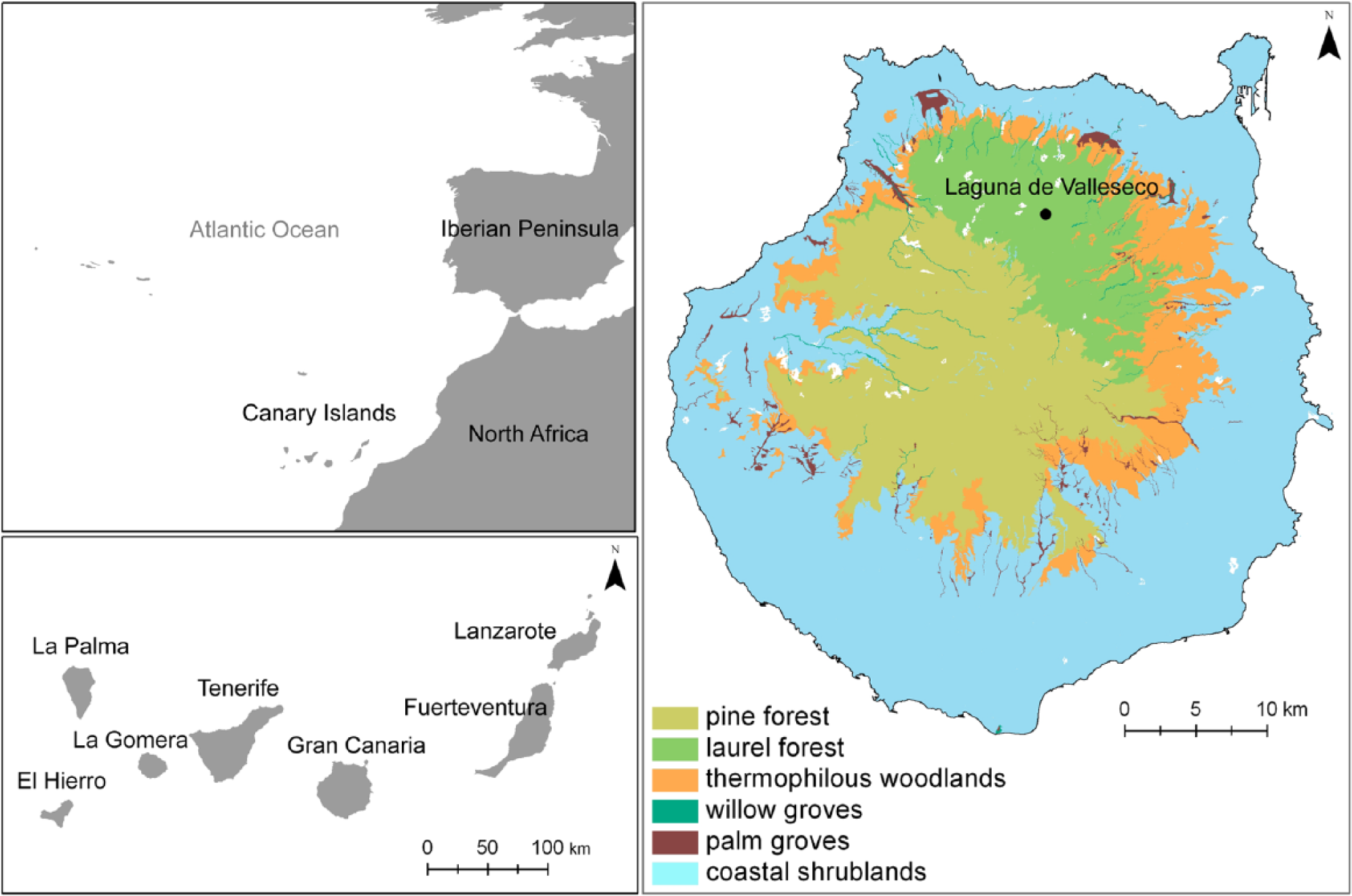
Map of Gran Canaria and the main vegetation types represented according to their potential distribution (Del Arco et al., 2006). The black dot represents the coring site in Laguna de Valleseco.
Materials and methods
A sediment core of 5.2 m long was extracted from the inside of the caldera, avoiding the area covered by the artificial lake. The extraction was performed using automatic drilling rotation equipment. Drilling stopped at 5-m depth when we found a matrix of silty-clay sediments several metres thick, including abundant and altered pyroclastic material, indicating that the slope deposits on the inner flank of the caldera had been reached. The core was kept in PVC tubes in a cold store at 4°C at the University of La Laguna. Tubes were opened for sediment description (Figure S1, available online), and an initial subsampling was carried out every 20 cm to check for pollen content. The preliminary analysis showed that only the first metre contained well-preserved pollen, thus we subsampled this section at 4-cm intervals.
A volume of 1-cm3 sediment was analysed for each level. Pollen extraction followed standard procedures described in Bennett and Willis (2001), including the addition of Lycopodium spores as an exotic marker, hot treatments with acid (hydrochloric acid) to remove carbonates, and with sodium pyrophosphate to disaggregate clay particles, and sieving at 150 µm to separate large particles and retain macro-charcoal fragments. Mineral fragments and organic particles were then removed from the fraction smaller than 150 µm using density separation with heavy liquid of specific gravity 2.1 (sodium polytungstate). The remaining sample, containing pollen grains, spores and micro-charcoal, was then acetolyzed for 3 min, stained, dehydrated and mounted in silicone oil for observation under the microscope at 400× magnification. A minimum count of 300 pollen grains was undertaken, excluding ferns, fungi and algal spores from the pollen sum. Pollen identification was based on the reference collection of Canarian native species held at the Ecology Laboratory in the University of La Laguna and photographs from several pollen atlases, including European and North African pollen types (Reille, 1992, 1995, 1998). Native and cultivated grasses were differentiated based on their pollen grain diameter, following conventions used in continental Europe (Joly et al., 2007).
The concentration of macro-charcoal (particles > 150 µm) and micro-charcoal (particles < 150 µm) per volume of sediment was estimated on each subsampled level (every 4 cm). All macro-charcoal particles retained after sieving 1 cm3 of sediment were counted using a stereomicroscope (Whitlock and Larsen, 2001). Micro-charcoal concentration (cm2 cm−3) in each sample was measured using the point count method on several microscope slides per sample, which provides an estimate of the area covered by micro-charcoal particles per unit volume of sediment (Clark, 1982).
Four samples of bulk sediment were dated using 14C Accelerator Mass Spectrometry in two different radiocarbon laboratories (14CHRONO Centre at Queen’s University Belfast and BETA Analytic). The radiocarbon dates were calibrated to years before present (cal. yr BP) using the IntCal13 database and the software Calib Rev 7.0.2 (Reimer et al., 2013; Stuiver et al., 2005). The age–depth model was obtained by linear interpolation from adjacent pairs of values assuming an unknown age for the top sample (Bennett, 1994).
Pollen percentages for each taxon, along with charcoal and total pollen concentration values, were plotted against age and depth in pollen diagrams generated using Psimpoll 4.26 (Bennett, 2008). The pollen diagram was zoned using optimal splitting by information content and the number of significant zones calculated with a broken-stick model (Bennett, 1996). Pollen types were also grouped into trees, shrubs, forbs, grasses, and ferns, and plotted against age. To detect those taxa that contributed to dissimilarities between zones, we performed a similarity percentage analysis (SIMPER) (Anderson et al., 2008). Correlations between macro- and micro-charcoal concentrations within specific pollen zones were also measured with the Spearman correlation test using the SPSS 15.0 statistical package (SPSS, 1986).
To describe local vegetation occurring in each period, we compared fossil pollen samples from Valleseco with the modern pollen rain composition and specific threshold values studied in the main Canarian vegetation types from the island of Tenerife (De Nascimento et al., 2015). Threshold values were calculated as the maximum and minimum pollen percentages for a particular taxon within a specific vegetation type. Similarities in composition between fossil and modern samples were explored with Detrended Correspondence Analysis (DCA) (Lepš and Šmilauer, 2003) using CANOCO for Windows 4.5 (Ter Braak and Šmilauer, 2002). Significant differences in composition between pollen samples grouped according to the different vegetation types (modern pollen) and pollen zones (fossil pollen) were tested by means of permutational analysis of variance (PERMANOVA) (p < 0.01), based on a Bray–Curtis distance matrix (Anderson et al., 2008). Average similarities between vegetation types and pollen zones were obtained after a pair-wise comparison using t-tests (p < 0.01) and Monte Carlo correction. PRIMER 6 with PERMANOVA+ software was used to perform permutational ANOVA and SIMPER analyses (Anderson et al., 2008).
Results
Chronology and stratigraphy
The first metre of sediments analysed for this study showed a homogeneous stratigraphy consisting of massive brown-reddish silts, resulting from alluvial fan deposition related to episodic flooding phases (details in Supplementary Material; Stratigraphic description and Figure S1, available online). According to the depth–age model, the sedimentary sequence covers a time span of c. 3000 years, from 4474 to 1506 cal. yr BP (Table 2; Figure S2 (available online)). Due to the low sedimentation rate during the first 2100 years of the record (0.005 cm yr−1), the resolution in this period, c. 4500–2300 cal. yr BP, is very poor. Higher sedimentation rates in the rest of the sequence (0.07–0.13 cm yr−1) allow for a good resolution of the subsequent period, c. 2300–1500 cal. yr BP, leaving a period of 900 years through which we are confident in interpreting changes in pollen abundance (Figure S2, available online). The relatively old date obtained at 10-cm depth (c. 1500 cal. yr BP) suggests that the soil within the caldera has been removed, very likely because of the most recent uses given to the site.
Radiocarbon ages (±SD) and calibrated ages (showing the interval ± 2SD) from Laguna de Valleseco (Gran Canaria).

SD: standard deviation.
Pollen zones
Four significant zones were determined by optimal splitting of the pollen diagram: V1 (94–82 cm), V2 (82–42 cm), V3 (42–14 cm) and V4 (14–0 cm), indicating that significant changes in vegetation occurred at c. 2300, 1800 and 1600 cal. yr BP (Figure 2).
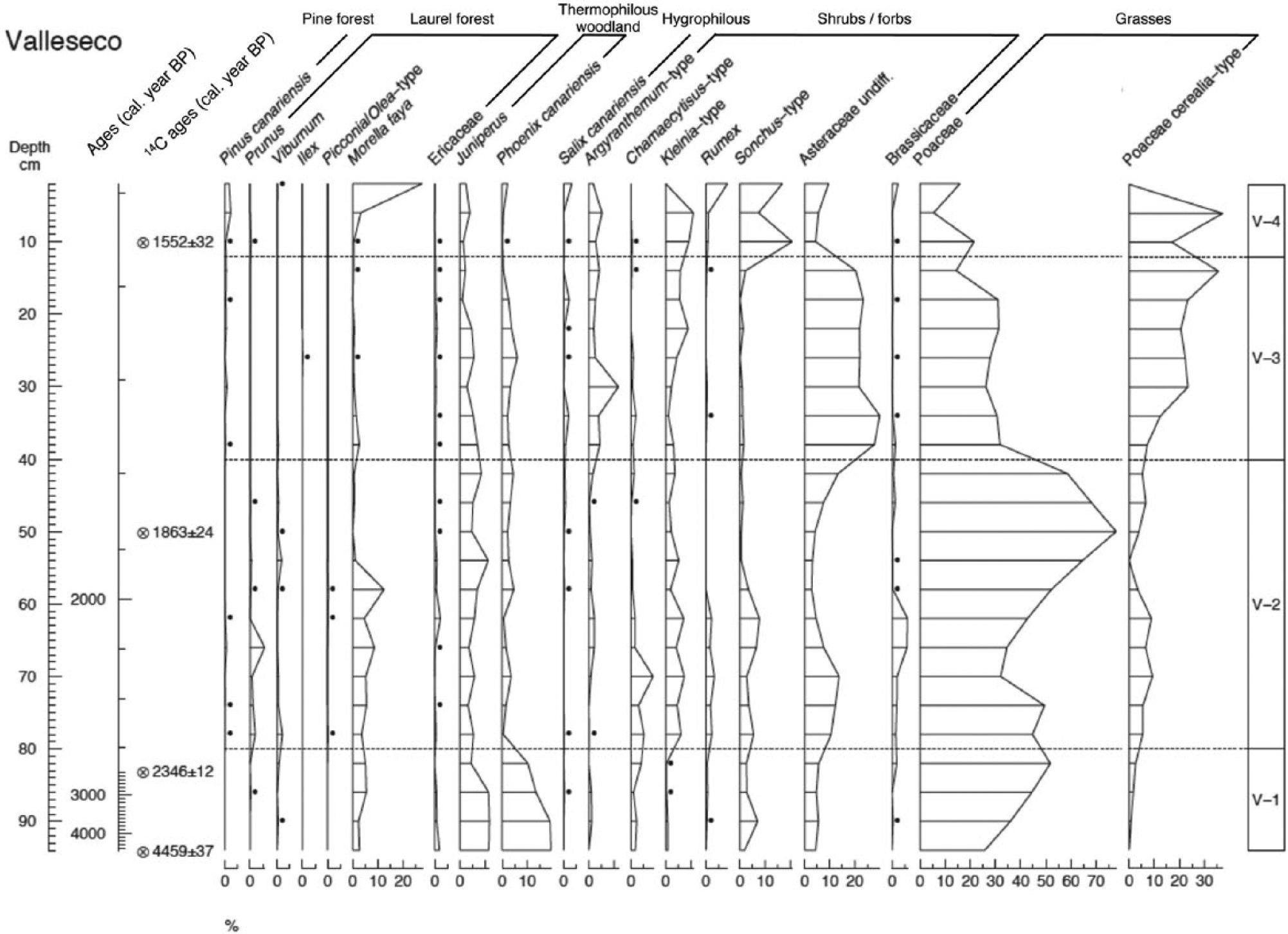
Percentage pollen diagram plotted against age (cal. yr BP) for the sedimentary sequence from Laguna de Valleseco, Gran Canaria, with selected taxa grouped in trees (arranged by forest type), shrubs and forbs, and grasses. For a complete diagram including all taxa, see Figure S1 in the Supplementary Material, available online. The dashed lines indicate the zones (V1, V2, V3 and V4) delimited by numerical zonation (optimal splitting within Psimpoll 4.25). Circles with crosses stand for 14C ages (cal. yr BP, mean ± SD). Dots represent values below 0.5%.
Zone V1: Thermophilous vegetation (c. 4500–2300 cal. yr BP)
In this zone, the sum of arboreal pollen reached its highest percentage (20–29%) apart from the uppermost sample in the sequence (37%) (Figure 3a). Phoenix canariensis (10–19%) and Juniperus (4–12%) were the two most abundant arboreal types in this period. Other tree pollen types, including Ericaceae, Prunus, Salix and Viburnum, appeared in very low proportions (<2%), while Morella reached slightly higher values (2–5%) (Figure 2). During this period, grasses rose to become the most common element (20–52%), mostly comprising native grasses. Poaceae with large pollen types were in a very low proportion (0–3%), and we assume that this signal originates from some of the native grasses presenting large pollen grains. The sum of shrubs and forbs ranged between 20% and 31% of the total pollen sum (Figure 3a): with Chamaecytisus, Globularia, Hypericum, Asteraceae, Sonchus and Urticaceae being the best-represented types (Figures 2 and S1 (available online)). Fern spores were also abundant (22%, mostly Davallia spores) in the oldest part of the diagram (Figure 3a).
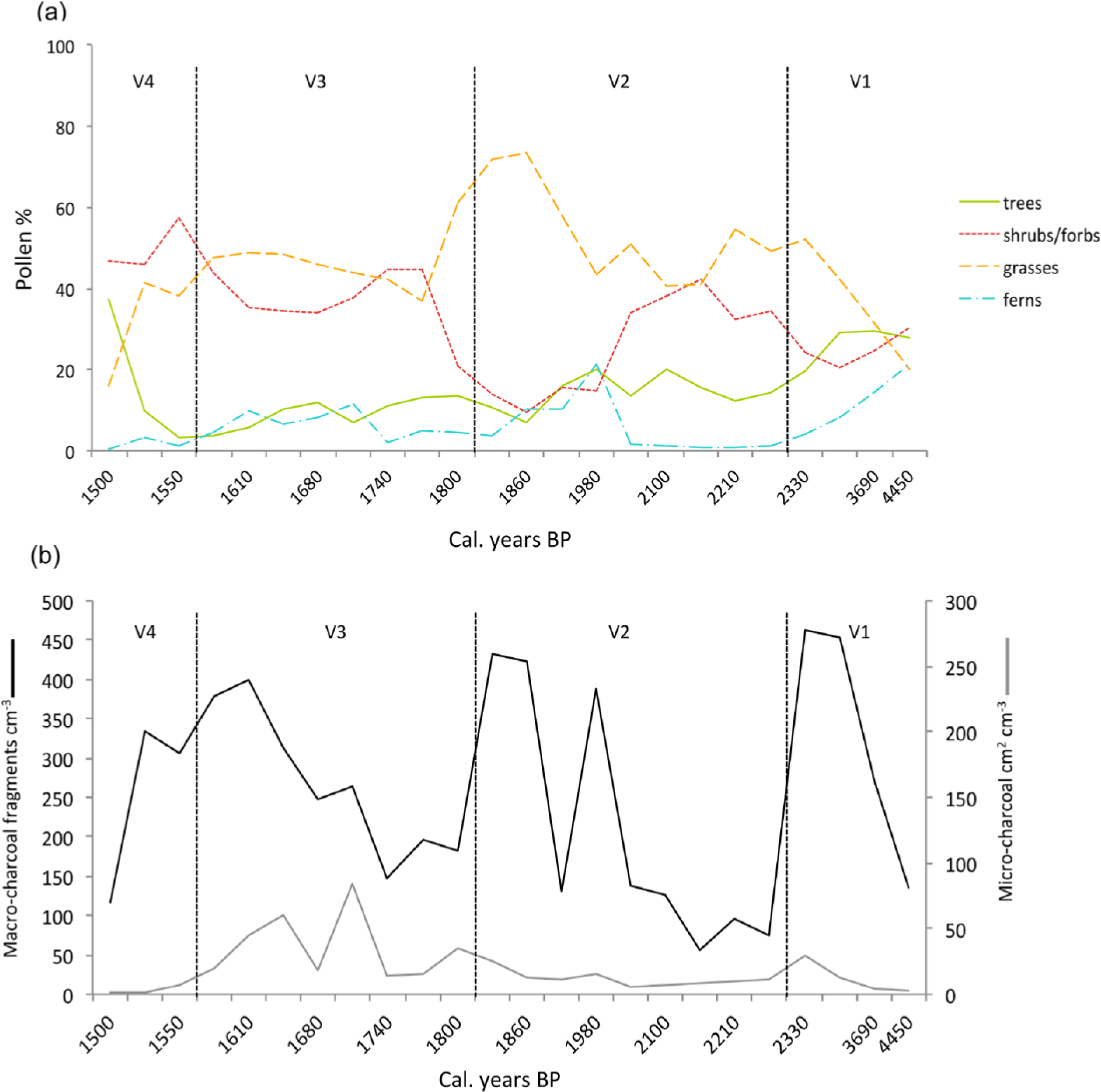
Summary of pollen sums and charcoal concentration by zones, plotted against age (cal. yr BP) for Laguna de Valleseco sediments: (a) pollen sums for trees, shrubs and forbs, grasses, and ferns; (b) macro- and micro-charcoal concentrations in particles cm−3 and cm2 cm−3, respectively. The dashed lines indicate the zones V1, V2, V3 and V4.
Zone V2: The spread of grasses (c. 2300–1800 cal. yr BP)
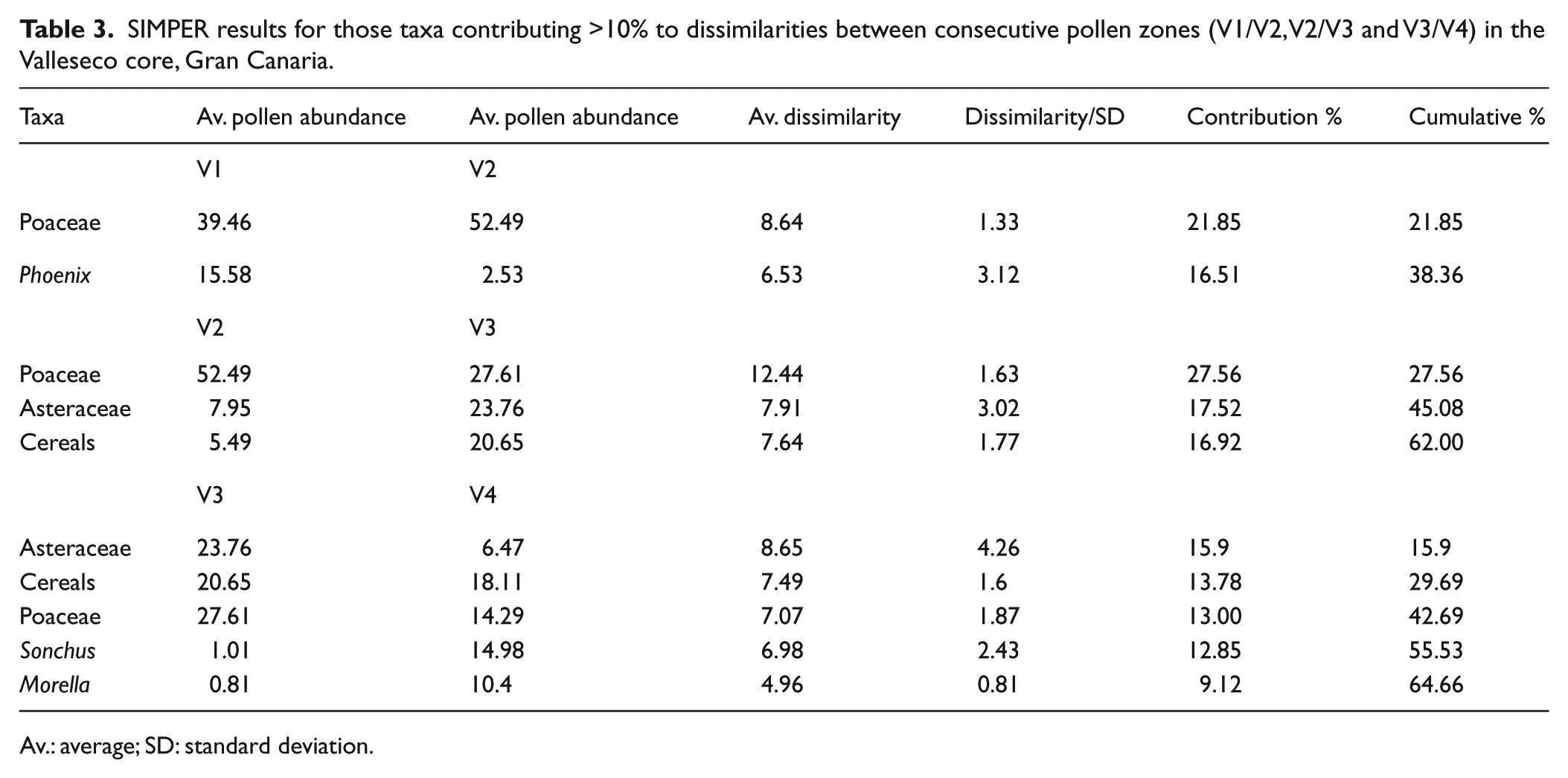
In this zone, increasing values of grasses (41–73%) started to replace trees (7–20%) and later shrubs and forbs (10–42%) (Figure 3). Native grasses, and shrubs and forbs from the Asteraceae, Brassicaceae and Fabaceae types, including Chamaecytisus, Kleinia and Sonchus, appear to have dominated the vegetation in this period (Figure 2). Among the trees, Juniperus (3–11%) and Morella (0–12%) displayed the highest values, while Phoenix canariensis decreased notably (0–5%) compared with zone V1 (Figure 2). Other trees, including Ericaceae, Oleaceae, Pinus canariensis, Prunus, Salix and Viburnum, were also recorded in this period but with a very low percentage (<2%) (Figure 2). The contribution of ferns as a group fluctuated in this zone (1–21%), but the spores of Ophioglossum increased (Figures 3a and S1 (available online)). The SIMPER results indicate that the distinction between zones V1 and V2 was marked principally by the decrease of Phoenix and the increase of Poaceae (Table 3). This is the sharpest change detected in the sequence, with an average dissimilarity between zones of 54.4%.
SIMPER results for those taxa contributing >10% to dissimilarities between consecutive pollen zones (V1/V2, V2/V3 and V3/V4) in the Valleseco core, Gran Canaria.
Av.: average; SD: standard deviation.
Zone V3: The emergence of cereals (c. 1800–1600 cal. yr BP)
The relative abundance of shrub and forb pollen increased in this zone (34–45%), with increasing representation of Asteraceae pollen, including Kleinia and Argyranthemum types (Figures 2 and 3a). Grasses (excluding cereal types) decreased slightly (37–49%), although pollen of cultivated cereals (7–36%) rose to became as significant as pollen of native grasses (14–32%) (Figures 2 and 3a). Arboreal pollen remained relatively low in abundance (4–13%). Juniperus and Phoenix provided the highest percentage (1–7%), while all other trees were below 2% (Figures 2 and 3a). Ferns, represented by Ophioglossum and Polypodium types, maintained their low values (2–12%) (Figures 3a and S1 (available online)). The change in pollen composition between zones V2 and V3 was less marked than between V1 and V2, with an average dissimilarity between zones of 45.1%. Differences were due to the increase of Asteraceae and cereals from zone V2 to V3 and the decrease of native grasses in zone V3 (Table 3).
Zone V4: Secondary vegetation (c. 1600–1500 cal. yr BP)
Within the most recent zone, arboreal pollen increased from their low representation to reach 37% by the end of the sequence, while grasses (excluding cereals) reached their lowest values (16%) (Figure 3a). Vegetation was dominated by shrubs and forbs (46–58%), including Sonchus, Kleinia, Argyranthemum and other Asteraceae, and by cereals (0–37%), which on average prevailed in their representation over the native grasses (5–21%) (Figure 2). Towards the end of the sequence, Morella became the best-represented tree (27%), while Juniperus and Phoenix reached their lowest values (0–4%) (Figure 2). We discard the possibility of pollen in these uppermost samples being of recent origin because there is no signal of pollen from high pollen-producer trees planted in the last centuries in the site (e.g. Castanea sativa and Pinus radiata). Ferns were scarcely represented in this zone (<3%) (Figure 3a). The average dissimilarity between zones V3 and V4 (39.5%) indicates a smaller change in pollen composition. Taxa contributing to the differences were Morella and Sonchus, showing an increasing trend in zone V4, and Asteraceae and Poaceae (both native and cultivated), which decreased (Table 3).
Charcoal
The mean abundance of macro-charcoal fragments over the last 4500 years is 253 fragments cm−3, with high concentrations within each zone and peaks occurring around 2300, 1900, 1800, 1600 and 1500 cal. yr BP (Figure 3b). Mean concentration of micro-charcoal was 19 cm2 cm−3 for the whole sequence, with the highest concentrations occurring in zone V3, followed by zone V1, indicating that regional fires were more important in those periods. Micro-charcoal peaks appeared at around 2300, 1800, 1700 and 1600 cal. yr BP (Figure 3b). Based on peak occurrences, both local and regional fires became more frequent after c. 1900 cal. yr BP than in the previous period (Figures 3b and S1 (available online)). Over the course of zones V1 and V2 (4500–1800 cal. yr BP), there was a significant correlation between levels of macro- and micro-charcoal (r = 0.547, p < 0.05) at each sample level.
Fossil and modern pollen rain comparison
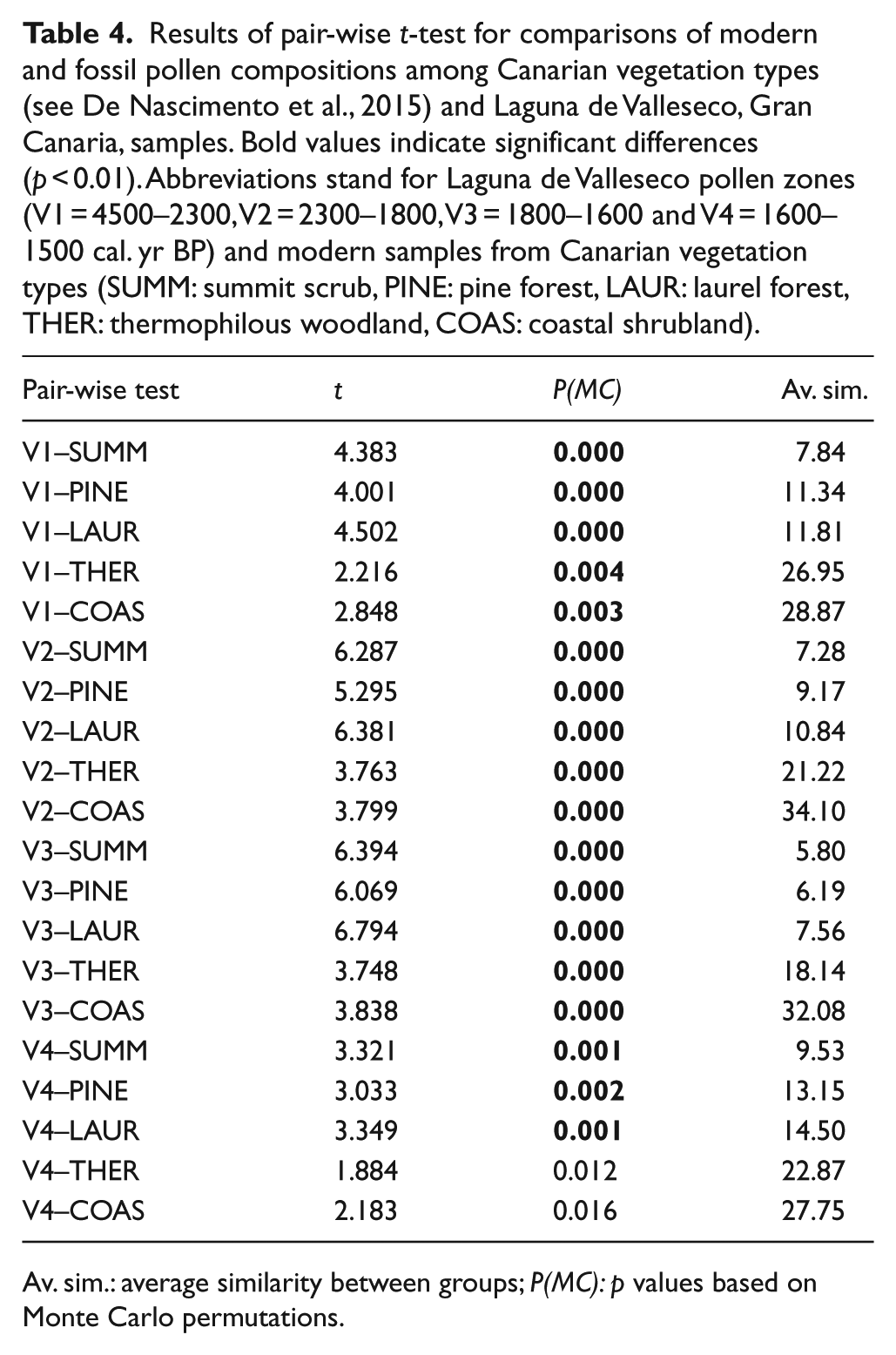
An initial DCA ordination analysis of the main Canarian vegetation types as represented in the modern pollen rain composition of Tenerife (De Nascimento et al., 2015) showed that the modern pollen composition of both the pine forest and summit scrub was highly dissimilar to that of the Valleseco fossil pollen samples. Average similarity percentages measured from the Bray–Curtis distance matrix show the lowest values (5.8–13.2%) between the four pollen zones and both vegetation types (Table 4). Accordingly, we repeated the ordination analyses including only samples from the laurel forest, thermophilous woodlands and coastal shrublands (Figure 4). The DCA diagram showed samples arranged along axis I according to different zonal vegetation types, in a gradient from the coastal shrubland to the laurel forest, while on axis II the gradient represented the variability in composition between samples of the thermophilous woodlands, with the Canarian palm groves (TW2, TL1) in the upper part of axis II and the Juniperus and Olea woodlands in the lowest part (TW3, TL3, TW1, TL2) (see De Nascimento et al., 2015). In general, fossil samples did not show a clear gradient along the axes, although samples from the oldest zone (V1 = 4500–2300 cal. yr BP) were closer in composition to the modern samples from the thermophilous woodlands (Figure 4). Based on average similarities, the best matches in composition for this zone occurred with coastal (28.9%) and thermophilous (26.9%) vegetation types (Table 4). Fossil samples from all other zones (V2 = 2300–1800, V3 = 1800–1600 and V4 = 1600–1500 cal. yr BP) were similar in composition to the modern pollen rain from coastal shrubland vegetation of Tenerife (Figure 4), especially to windward coastal sites. This was confirmed from the highest average similarities obtained between each of these zones and the coastal shrubland samples (34.1%, 32.1% and 27.7% for V2, V3 and V4, respectively) (Table 4). This is very likely because of the abundance of Poaceae and Asteraceae in these coastal sites, which are also common elements in the secondary shrubland. However, as we have no record of the modern pollen rain from secondary shrubland in the Canaries, this interpretation is tentative. The virtual absence of Euphorbia pollen from these samples also points to the occurrence of secondary vegetation, as Euphorbia is a good indicator of the coastal shrubland. Finally, the most recent fossil sample in the Valleseco sequence (c. 1500 cal. yr BP) shows an abrupt switch towards the modern pollen signal for thermophilous woodlands (compare Figures 2–4). PERMANOVA results showed that, in general, fossil pollen samples from Valleseco differed significantly from modern pollen rain composition of each of the main Canarian vegetation types (Pseudo-F = 14.563, p < 0.01), except for the zone V4, which was not different from the modern pollen rain in the thermophilous and coastal vegetation types (Table 4).
Results of pair-wise t-test for comparisons of modern and fossil pollen compositions among Canarian vegetation types (see De Nascimento et al., 2015) and Laguna de Valleseco, Gran Canaria, samples. Bold values indicate significant differences (p < 0.01). Abbreviations stand for Laguna de Valleseco pollen zones (V1 = 4500–2300, V2 = 2300–1800, V3 = 1800–1600 and V4 = 1600–1500 cal. yr BP) and modern samples from Canarian vegetation types (SUMM: summit scrub, PINE: pine forest, LAUR: laurel forest, THER: thermophilous woodland, COAS: coastal shrubland).
Av. sim.: average similarity between groups; P(MC): p values based on Monte Carlo permutations.
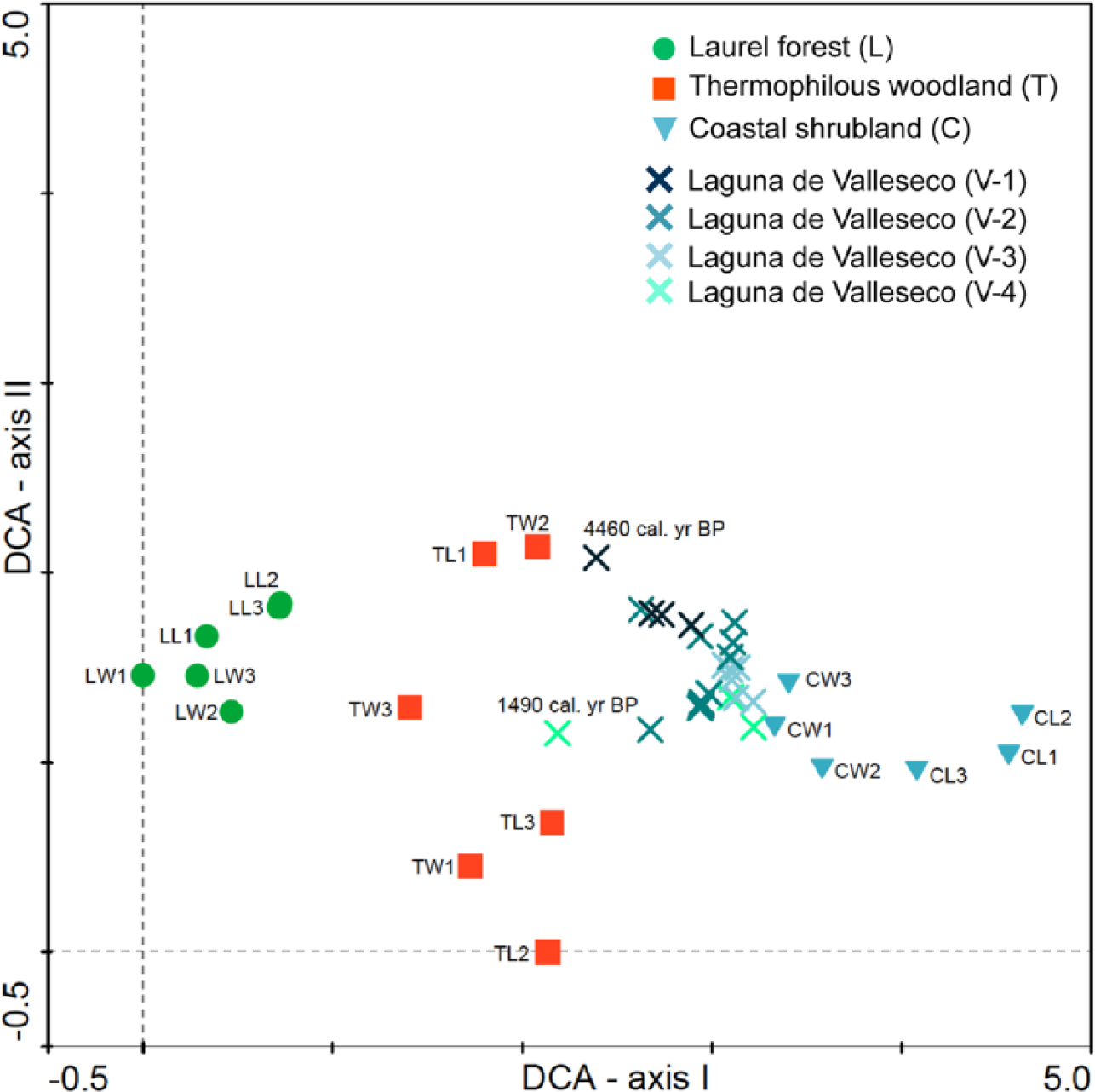
Detrended Correspondence Analysis (DCA) scatter-plot based on pollen composition (%) of modern samples from different vegetation types on Tenerife (see De Nascimento et al., 2015), and fossil pollen samples of Laguna de Valleseco, classified according to pollen zones (V1 = 4500–2300, V2 = 2300–1800, V3 = 1800–1600 and V4 = 1600–1500 cal. yr BP) (total inertia = 2.9, cumulative variance = 30.0%). Names of present pollen plots indicate vegetation type (first letter, C: coastal shrubland, T: thermophilous woodland, L: laurel forest), aspect (second letter, W: windward, L: leeward) and elevational position (1–3 from lowest to highest elevation).
We compared fossil pollen percentages of the most abundantly represented trees recorded in Valleseco (Juniperus, Phoenix and Morella) with the modern pollen threshold values obtained for each taxon within the thermophilous vegetation and laurel forest sites (Figure 5). In the oldest part of the sequence (V1 = 4500–2300 cal. yr BP), both Juniperus and Phoenix exceed the minimum modern threshold values recorded for the thermophilous vegetation, with fossil Phoenix percentages also exceeding the maximum modern threshold (Figure 5). However, in the rest of the sequence, fossil pollen percentages for both trees are continuously below the minimum modern threshold. Fossil pollen percentages for Morella through the sequence remain within the modern thresholds recorded for this taxon in the thermophilous vegetation, and only in the most recent sample (c. 1500 cal. yr BP) does Morella fossil pollen reach the minimum threshold for its modern pollen percentage in the laurel forest (Figure 5).
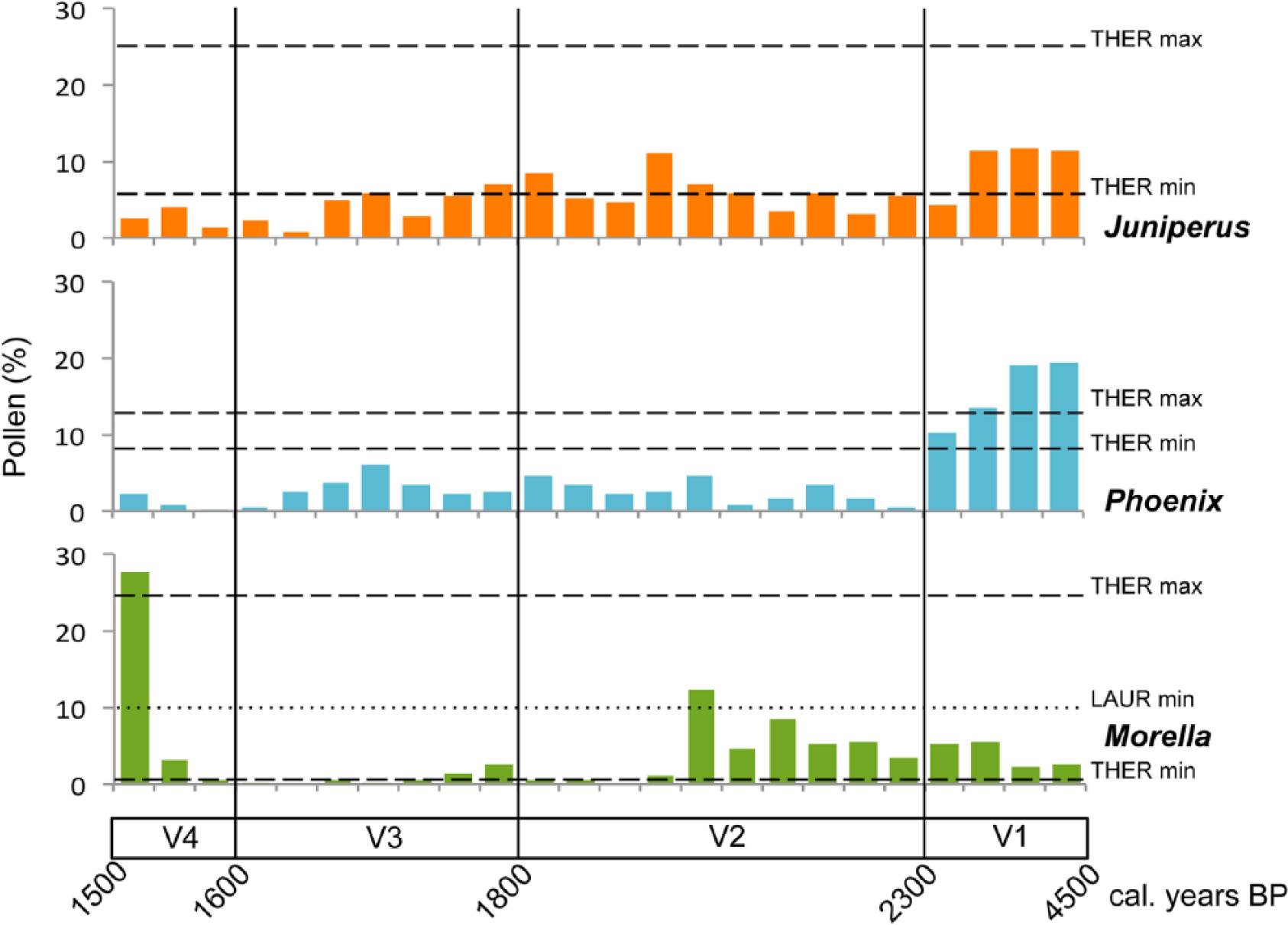
Fossil pollen percentages for the main trees recorded (Phoenix, Juniperus and Morella) in Laguna de Valleseco, Gran Canaria, plotted against age and modern pollen threshold values calculated as the percentage of pollen for each taxon within thermophilous and laurel forest vegetation types from Tenerife (see De Nascimento et al., 2015). Dashed lines represent both maximum and minimum thresholds for the pollen representation of each taxon within the thermophilous woodlands (THER) and the dotted line represents the minimum threshold for Morella pollen representation within the laurel forest (LAUR).
Discussion
Laguna de Valleseco has provided the first study of past vegetation on the island of Gran Canaria by means of pollen analysis. The sedimentary record spanned part of the late Holocene (c. 4500 cal. yr BP) but, due to irregular sedimentation rates, the best temporal resolution was restricted to the period c. 2300–1500 cal. yr BP. Apparently, the soil from the surface of the caldera has been removed in recent times, therefore the last 1500 years of vegetation history were not available from this sequence. The soil truncation can be explained by a common local practice from the 20th century, known as ‘sorribas’, which involves the removal of fertile soils from mid-elevation areas to enrich the soils of coastal export crops. Nevertheless, the record spans the period immediately before and following human colonization (occurring at least around 1900 years ago), even allowing for the uncertainty involved in dating that event. Unfortunately, it does not allow us to explore the shifts during the last 1000 years of aboriginal occupation (c. 1500–500 cal. yr BP) and the historical period after European conquest (c. 500 cal. yr BP).
Vegetation history
The predominance of grasses throughout the sequence (Figures 2 and 3a) reflects the existence of relatively open vegetation within the area since c. 4500 years ago. However, we detected three major shifts in species composition and abundance. Vegetation began to change around 2300 years ago, and experienced repeated transformations within the following centuries. Our data indicate that the vegetation of the area has changed from relatively open woodlands to grasslands, which were later replaced by cultivated cereals, followed by encroachment by secondary shrubland vegetation and a final switch to pioneers of the evergreen laurel forest (mostly Morella). Considering these vegetation transitions, we have divided the discussion into two significant periods: before and after the decline of woodlands.
The woodlands around Laguna de Valleseco (c. 4500–2300 cal. yr BP)
Within this period, there was forested vegetation around Laguna de Valleseco that resembled contemporary remnants of thermophilous woodlands (Figure 4). The tree layer was formed mainly by two taxa, Juniperus and Phoenix, while native grasses and several woody species like Globularia, Hypericum or Sonchus, which can be typically found in the thermophilous vegetation at present (Del Arco et al., 2006; González-Artiles, 2006), were important in the understory. Certainly, this is the only period where these trees were abundant enough to produce a pollen signal comparable to the modern pollen rain characteristic of communities such as the Juniperus turbinata woodlands or Phoenix canariensis palm groves that occur within the thermophilous vegetation zone (De Nascimento et al., 2015). Taking account of the modern pollen rain threshold values (Figure 5), it is evident that the palm groves were important in the area and the number of palm trees at that time around the site may even have exceeded that of palm groves occurring in the islands at present (Fernández-Palacios et al., 2008; Sosa et al., 2007).
The prevalence of these communities in the area in this period is unexpected. Considering the current climatic conditions for the site, which is located on the windward slope of the island at 870 m of elevation, the area has been included within the evergreen laurel forest potential vegetation (Del Arco et al., 2006) (Figure 1). This is in contrast with our results, since the average pollen percentage of laurel forest taxa found in the sediments of Laguna de Valleseco (Morella, Ericaceae, Prunus and Viburnum) was extremely low compared with average modern pollen rain values obtained from laurel forest stands on Tenerife (4% vs 40%, 0.5% vs 46%, 0.5% vs 4%, and 0.3% vs 1%, respectively) (De Nascimento et al., 2015). A weak pollen signal is common for entomophilous trees typical from the laurel forest, such as Prunus and Viburnum. Despite their under-representation, both pollen types are good indicators of the local occurrence of these trees. In contrast, Morella and Ericaceae trees produce abundant pollen so that they are usually over-represented in the pollen rain in relation to their abundance in the vegetation, thus we might expect higher contributions of both types if they were well-represented in the area. Furthermore, because Lauraceae species are typically under-represented in the laurel forest pollen rain, it seems reasonable to use this subset of taxa (i.e. Ilex, Ericaceae, Morella, Picconia, Prunus and Viburnum) as indicators of the presence of laurel forest in the area. Thus, considering the low pollen contribution obtained for these taxa, we infer a low or distant presence of the laurel forest elements around the site at that time. Juniperus and Phoenix pollen are, however, among the well-represented types (De Nascimento et al., 2015), confirming the presence of both trees in the area. The Phoenix type, in particular, is a good indicator of local palm occurrence because it shows lower dispersability values than Juniperus (De Nascimento et al., 2015). Given its phreatophytic ecology (Morici, 2006), the Canarian palm is capable of withstanding periodic flooding over the caldera bottom, providing a competitive advantage in relation to other trees that were limited to the drained areas. At present, Olea and Pistacia woodlands are the best-preserved and largest communities of thermophilous vegetation in Gran Canaria (Del Arco et al., 2010). However, Olea and Pistacia pollen types are scarcely found in the fossil record, so it remains unclear what the past role of Olea and Pistacia woodlands may have been within this region. Both taxa appear also in the Tenerife and La Gomera cores but always in low proportions compared with other trees of the thermophilous woodlands (De Nascimento et al., 2009; Nogué et al., 2013). Moreover, while Pistacia pollen was well-represented in the modern pollen rain and was identified as a good indicator type of the presence of its source plant, Oleaceae pollen provided only small contributions to the modern pollen rain (De Nascimento et al., 2015). Thus, we can confirm the past presence of Olea and Pistacia woodlands in the islands, but we cannot estimate their importance based on the available data.
Canarian palm groves in Gran Canaria are among the best represented in the archipelago, together with those of La Gomera (Fernández-Palacios et al., 2008). It has been variously estimated that they cover between 600 and 1200 ha (Del Arco et al., 2010; Sosa et al., 2007). They are currently distributed along ravines all over the island, from sea level to their highest locations at 1200 m a.s.l. in the south, although their optimal range, 0–600 m a.s.l., is mainly within the thermophilous vegetation zone (Sosa et al., 2007). Conversely, Juniperus turbinata is the least common tree in the relictual patches of thermophilous woodland on Gran Canaria (Del Arco et al., 2006; González-Artiles, 2006) and is currently found in isolated leeward areas in the southwestern region of the island, mainly at 500–1000 m a.s.l. (González-Artiles, 2006). However, macro-charcoal fragments of Juniperus spp. have been identified in the deposits of a northern ravine from the mid-Holocene in Gran Canaria (Hansen and Criado, 1996), while some historical quotes referred to the presence of this tree also in the north of the island (González-Artiles, 2006), which is consistent with our results. Moreover, a recent study of its occurrence within the thermophilous vegetation patches remaining today in the island suggests that Juniperus turbinata could have had a circuminsular distribution in the past (González-Artiles, 2006), as has been also noted for this species in Tenerife (Otto et al., 2012). Based on the elevation ranges for the different localities of this species, its potential distributional area has been estimated as 14,800 ha, which added to the potential area of Olea and Pistacia woodlands, make a total of 42,200 ha (González-Artiles, 2006). This surface considerably exceeds previous estimates of the original area of this major ecosystem type (Del Arco et al., 2010), therefore decreasing from 11% to 3% the percentage of the possible original distribution area of thermophilous woodlands that remains at present. Juniperus cedrus, the other species of juniper found on Gran Canaria, is extremely rare and only occurs in inaccessible cliffs far away from the study site (>20 km) (Rumeu et al., 2014).
Thermophilous woodlands, as the name suggests, are warmth-loving communities adapted to the summer hydrological and winter thermal stresses provided by the Mediterranean climate. Since the establishment of the Mediterranean climate regime c. 2.5 Ma, the Canarian laurel forest has been dependent on the constant supply of humidity provided by the NE trade winds. We may assume the laurel forest to have been restricted to the area of influence of the ‘sea of clouds’ on windward slopes, while the thermophilous woodlands and the Canarian pine forest, which are more tolerant of the summer drought, were spread, respectively, below and above the laurel forest (Fernández-Palacios et al., 2008). Palaeoecological records of several islands within the archipelago have provided evidence of a trend towards a drier climate during the late Holocene (Nogué et al., 2013; Von Suchodoletz et al., 2010; Yanes et al., 2011), which may have favoured the expansion of thermophilous woodlands within the last 5000 years, at least until human arrival.
The significance of both palm groves and juniper woodlands around the study site in the period 4500–2300 cal. yr BP may support the idea of a much more widely distributed thermophilous vegetation in the past, where not only Olea and Pistacia woodlands but also Phoenix palm groves and Juniperus cf. turbinata woodlands were important and very likely exhibited a circuminsular distribution. Consequently, the distribution of the laurel forest at that time must have been largely circumscribed to the purely north facades of the island or at least at higher elevations in the northeastern slopes (see Santana, 2001). Examples of mismatches between the natural potential vegetation, defined from modern floristic and climatic considerations, and past vegetation, inferred from palaeoecological evidence, are often found both in continents (Carrión and Fernández, 2009 and references therein) and in islands (De Boer et al., 2013; Reille et al., 1999; Tinner et al., 2009; Virah-Sawmy et al., 2009; Wilmshurst et al., 2013); accordingly, existing potential vegetation models should be reviewed. In addition, it is also necessary to continue searching for more palaeoecological archives to improve the resolution and extent of our knowledge of past vegetation.
The spread of grassland and shrubland (c. 2300–1500 cal. yr BP)
The earliest known date available of human presence on the island of Gran Canaria, based on radiocarbon dating of archaeological sites, is c. 1900 cal. yr BP (Del Arco et al., 1997; Martín, 2000). However, some authors believe that the arrival of humans on the island must have occurred earlier, because Gran Canaria, one of the biggest and highest islands in the archipelago, offered abundant resources, and is half-way between the islands of Lanzarote and Tenerife, where radiocarbon dates provide evidence of earlier human presence at c. 2900 and 2800 cal. yr BP, respectively (Atoche, 2008). The first change in vegetation identified in Laguna de Valleseco occurred 2300 years ago, about 400 years before the first archaeological evidence of human presence in the island. At that time, the thermophilous vegetation began to be replaced by grasslands, especially Phoenix palm groves, which experienced a significant decline.
This reduction of palms in favour of native grasses might be related to changes in fire regimes, as shown by the peak in both macro- and micro-charcoal concentrations occurring at that time (Figure 3b). Within this period (c. 2300–1800 cal. yr BP), the concentration of both parameters is correlated, suggesting that the area was subjected to fires that reached the site, thus leaving both macro- and micro-charcoal signals in the sediments. It is possible that fires could have been originated by volcanic activity. The only eruption occurring nearby (3 km away) at that time was that of Doramas (c. 2400 cal. yr BP) (Rodriguez-Gonzalez et al., 2009), which might have initiated fire events in the area. However, the Doramas eruption was strombolian, a type of volcanism characterized by the formation of cinder cones with small amounts of associated tephra layers and long lava flows running through the courses of ravines, thus only affecting nearby areas. This means that it is unlikely that such events had long-lasting or far-reaching impact on the vegetation.
An alternative explanation for the increase of fires between c. 2400 and 2300 cal. yr BP would be that humans had already colonized the island and consequently changed the natural fire regime. The first colonizers may have intentionally used fires to clear the vegetation in order to provide new land for agricultural practices, and they may also have felled trees to obtain timber and other plant resources. Fragments of macro-charcoal from Juniperus turbinata and Phoenix canariensis are often found on archaeological sites, indicating that both trees were used as fuel (Machado, 2007). Charcoal remains from La Cerera archaeological site (Arucas), which is some 7 km to the north of Laguna de Valleseco, but at a lower elevation (285 m a.s.l.), reveal the use of conifers, including Juniperus spp., by humans since at least c. 1600 cal. yr BP, coinciding with the last 100 years of our sequence, and persisting until the arrival of Europeans on the island at the end of the 15th century (Machado, 2009). Charcoal from Pinus canariensis is by far the most abundant element in La Cerera archaeological site, whereas identified charcoal of Juniperus was present in a lower proportion and there was no charcoal from Phoenix. This is consistent with our results since at that time, Juniperus and Phoenix had greatly reduced in abundance around Laguna de Valleseco. Despite its low elevation, La Cerera site contained charcoal of many arboreal species from all the forests in the island, but especially from the pine and the laurel forests during the most recent times (c. 1300–500 cal. yr BP), which may indicate that humans were collecting wood from farther distances at that time, probably suggesting that the nearby areas, including Laguna de Valleseco, had been deforested in earlier times. Apart from the use of wood as fuel, many other uses were made of both species, as suggested by the earliest historical descriptions. Palms specifically provided food (fruits, palm hearts, beverages), tools (sewing needles made with thorns, mats of leaves, fishing buoys made of stalks, rafts made of trunks) and fibres (clothing, sails, fishing lines), while wood from junipers was used for dwelling construction (roofs, doors) and tools (fishing rods, weapons) (Morales, 2003).
The decline, extirpation or extinction of palms following human colonization has been also recorded from Holocene palaeoecological archives in the Pacific islands, coinciding with evidence of increased fires, the spread of grasses and the introduction of cultivated plants (Prebble and Dowe, 2008). Human impact on vegetation has also been detected from pollen records in Tenerife, similarly involving the disappearance of trees, together with the increase of fires and grasses after human settlement of the island (De Nascimento et al., 2009). The decline of Phoenix and Juniperus in particular has also been noticed in both Tenerife and La Gomera fossil records. At around 2900 cal. yr BP, both Phoenix and Juniperus decreased slightly in Tenerife, but no apparent signs of human activity were detected at that time in the area; in turn, changing representation of other taxa suggests that this change was more related to climatic variability (De Nascimento et al., 2009). On La Gomera, palms were abundantly represented since at least the early Holocene, until a marked decline occurred about 5500 years ago, most probably in response to regional climatic trends (Nogué et al., 2013). In addition, the first palaeoecological study carried out in the Azores detected the collapse of Juniperus brevifolia communities after recent (European) human colonization, showing the vulnerability of this species to human impact in comparison to other natural disturbances that may occur in these islands, such as climate change or volcanic activity (Connor et al., 2012).
In the following centuries, 1800–1600 cal. yr BP, vegetation was open and dominated by Poaceae and Asteraceae species, showing no signs of woodland recovery, while cultivated cereals increased significantly, pointing to the development of agriculture in the site. Archaeological evidence confirms that humans were already on the island at that time (Del Arco et al., 1997; Martín, 2000). Agriculture was a common practice among the aborigines, although the degree of development varied among islands. In Gran Canaria, both rain-fed and irrigated agriculture coexisted, mostly relying on cereal cultivation (Velasco, 1998). Seeds recovered from archaeological sites show that Gran Canaria presented the highest variety of cultivated plants, including cereals (barley and wheat), legumes (lentils, field beans and peas) and fig trees, as well as being the only island in the archipelago where prehistoric granaries have been found (Morales et al., 2009, 2014). Within this period, fire became more frequent at the regional scale, while local fire was reduced c. 1800–1700 cal. yr BP, suggesting that humans were often burning other areas in the island but may also have lit fires within the site. The last volcanic eruption in Gran Canaria, Pico and Caldera de Bandama (c. 1900 cal. yr BP) has been dated close to this period (Rodriguez-Gonzalez et al., 2009). Caldera de Bandama, about 900 m in diameter and 350 m deep, was formed by a phreatomagmatic event involving a more explosive eruption and the production of tephra layers that covered an area of at least 11 km2. However, due to the dominant wind direction the ashes were mainly deposited towards the south east of Bandama (Hansen et al., 2008); indeed, we did not find any tephra layer within the sediments of Laguna de Valleseco, which is located several kilometres towards the west of Bandama. Consequently, Bandama might had a significant destructive effect on its nearby vegetation and could set fires in the region, contributing to the increase of the micro-charcoal signal, but whether it affected the vegetation at farther sites such as Laguna de Valleseco is difficult to assess. Probably, humans would have caused most fires occurring after this last eruption, considering that lightning strikes, which are rare, would be the only alternative cause of wildfires in the absence of volcanic activity.
The last 100 years of the sequence (c. 1600–1500 cal. yr BP) are characterized by the dominance of shrubs and a slight recovery of trees, especially of Morella faya, towards the end. The presence of cereals denotes the maintenance of agriculture at least for some time, until apparently Morella began to spread within the nearby vegetation. The increase of Morella faya following human colonization is a constant feature in each of the fossil pollen records of Macaronesian islands analysed so far (Connor et al., 2012; De Nascimento et al., 2009; Nogué et al., 2013). This tree, together with Erica spp., is a pioneer species within the laurel forest (Fernández-Palacios and Arévalo, 1998); consequently, its germination is favoured when natural or anthropogenic disturbances open the canopy and increase light conditions in the understory. While the forest is recovering, both trees dominate the canopy, forming the Erica–Morella woody heath community; in addition, adult trees of these species may persist within mid- to late-successional stages of the laurel forest (Arévalo et al., 2012). Finally, the charcoal signal also shows a reduction in fire, which could indicate the abandonment of agriculture in the area. Although archaeological inventories reveal an extensive occupation of the territory, large population nuclei and irrigated agriculture were established mostly in the fertile valleys located within the lower reach of ravines, whereas at higher elevations the settlements were of smaller size, with rain-fed agriculture and grazing as main activities (Santana, 1992; Velasco, 1998). If following this model, cultivation of cereals in Laguna de Valleseco must have been rain-fed and probably developed only temporarily, in the early stages of human settlement on the island.
Conclusion
Despite the paucity of suitable sites for pollen analysis in the Canary Islands, Laguna de Valleseco provided a first opportunity to track vegetation history during the late Holocene (c. 4500–1500 cal. yr BP) in the north of Gran Canaria. Forested vegetation was only present during the first 2000 years of the record and resembled thermophilous vegetation communities; specifically, Phoenix canariensis palm groves and Juniperus cf. turbinata woodlands were prominently represented around the site. This contradicts potential vegetation models that predict the presence of laurel forest in the area. Despite the fact that thermophilous vegetation is relatively well-represented in Gran Canaria as compared with the other islands, our findings, together with other studies of the thermophilous remnants within the island, suggest a much wider distribution of the thermophilous vegetation in the past, meaning that the percentage of the original area of this vegetation type that remains at present is much lower than current estimates suggest. Accordingly, thermophilous communities may be considered among the most reduced and thus threatened ecosystems in Gran Canaria and should also be regarded a priority in restoration plans.
Several changes in vegetation have occurred since c. 2300 cal. yr BP, mainly involving the clearance of woodlands, as indicated by a marked decline of palms and the spread of grasses. Our results could be indicating that Gran Canaria was populated earlier than previously demonstrated. Later, once the forest had been cleared from the area, agriculture was locally established, as suggested by the increase of cereals after 1800 cal. yr BP. These changes were accompanied by an increase in fire frequency, and some coincided with archaeological evidence of the earliest human presence in the island, which led us to infer that humans were mostly responsible for the landscape transformation, although we do not entirely discard the sporadic impact of recent and nearby volcanic eruptions.
Footnotes
Acknowledgements
Drilling on Laguna de Valleseco was possible thanks to the permission and logistic support provided by the Ayuntamiento de Valleseco. We are also grateful to the technical staff of Estudios del Terreno, S.L., Fernando and Miguel, for their patience and accuracy during the drilling work. Finally, we thank Francisco J Pérez Torrado for his valuable comments on an early draft of this paper.
Funding
This work was supported by the Spanish Ministry of Science and Innovation (CGL 2009-10939). SN participated through a postdoctoral fellowship from the Spanish Ministry of Education (EX2009-0669). CR has been supported by a Short Term Mobility programme granted by the National Research Council of Italy.