
Abstract
Large-scale atmospheric pressure centers, such as the Aleutian and Icelandic Low, have a demonstrated relationship with physical lake characteristics in contemporary monitoring studies, but the responses to these phenomena are rarely observed in lake records. We observe coherent changes in the stratification patterns of three deep (>30 m) lakes inferred from fossil diatom assemblages as a response to shifts in the location and intensity of the Aleutian Low and compare these changes with similar long-term changes observed in the δ18O record from the Yukon. Specifically, these records indicate that between 3.2 and 1.4 ka, the Aleutian Low shifted westward, resulting in an increased frequency of storm tracks across the Pacific Northwest during winter and spring. This change in atmospheric circulation ultimately produced deeper mixing in the upper waters of these three lake systems. Enhanced stratification between 4.5 and 3.3 ka and from 1.3 ka to present suggests a strengthened Aleutian Low and more meridional circulation.
Introduction
Wintertime atmospheric circulation in the northern hemisphere is profoundly influenced by the Aleutian Low (AL) and Icelandic Low. These low-pressure cells are seasonally interdependent atmospheric processes, with an inter-annual oscillation pattern that is seesaw-like in nature (Honda et al., 2001; Overland et al., 1999; Rodionov et al., 2007). Because of the influence of these semi-persistent low-pressure cells on stationary flow patterns and the position of migratory major storm tracks, they have a significant impact on long-term climate patterns. The closely associated teleconnective climate indices of the Pacific-North America (PNA) pattern and the North Atlantic Oscillation (NAO) are linked to ecological phenology in modern lakes (Adrian et al., 2006; Gerten and Adrian, 2001; Straile et al., 2003) and are often attributed to paleolimnological patterns sharing similar spectral frequencies (Ólafsdóttir et al., 2013; Stevens et al., 2006). However, the response among North American lakes is mixed, showing limited regional coherence (Barron and Anderson, 2011; Jones et al., 2001; Katsuki et al., 2009; Nõges et al., 2010) although they are influenced by the same atmospheric processes.
The intensity and position of the Aleutian low-pressure center exerts a major influence on the climate of the North Pacific and western North America (Overland et al., 1999; Rodionov et al., 2005). The AL is a seasonally persistent Northern Hemisphere atmospheric circulation pattern that forms over the North Pacific during the cool season (September–May). The unstable mean flow extending from the AL generates transient storm systems, which are carried across North America along the axis of the westerly jet core or the prevailing storm track (Rodionov et al., 2007; Wise, 2012). On decadal and longer time scales, the intensity and position of the AL shifts, which influences the routing of winter season storm tracks across western North America. This is reflected in the long-term variability of the North Pacific Index (NPI) and in the sign of the Pacific Decadal Oscillation (PDO) (Chhak et al., 2009; Schneider and Cornulle, 2005). When shifted eastward and intensified, the AL enhances meridional flow, which routes storms along the North American coast and northward, into the Gulf of Alaska (Figure 1). During weakened (westward) shifts in the AL, meridional flow is weakened, zonal flow intensifies, and storms are more commonly routed into the continental interior (Rodionov et al., 2005). Decadal variability in the AL has been linked to changes in sea surface temperatures and thermocline variability in the North Pacific and, as such, has important implications for ocean ecosystems (Katsuki et al., 2009; Miller et al., 2004; Minobe, 1999), as well as patterns of drought, snowpack variability, and continental terrestrial and aquatic ecosystems (Barron and Anderson, 2011; Cayan et al., 1998; Gan, 2000; Pederson et al., 2013).
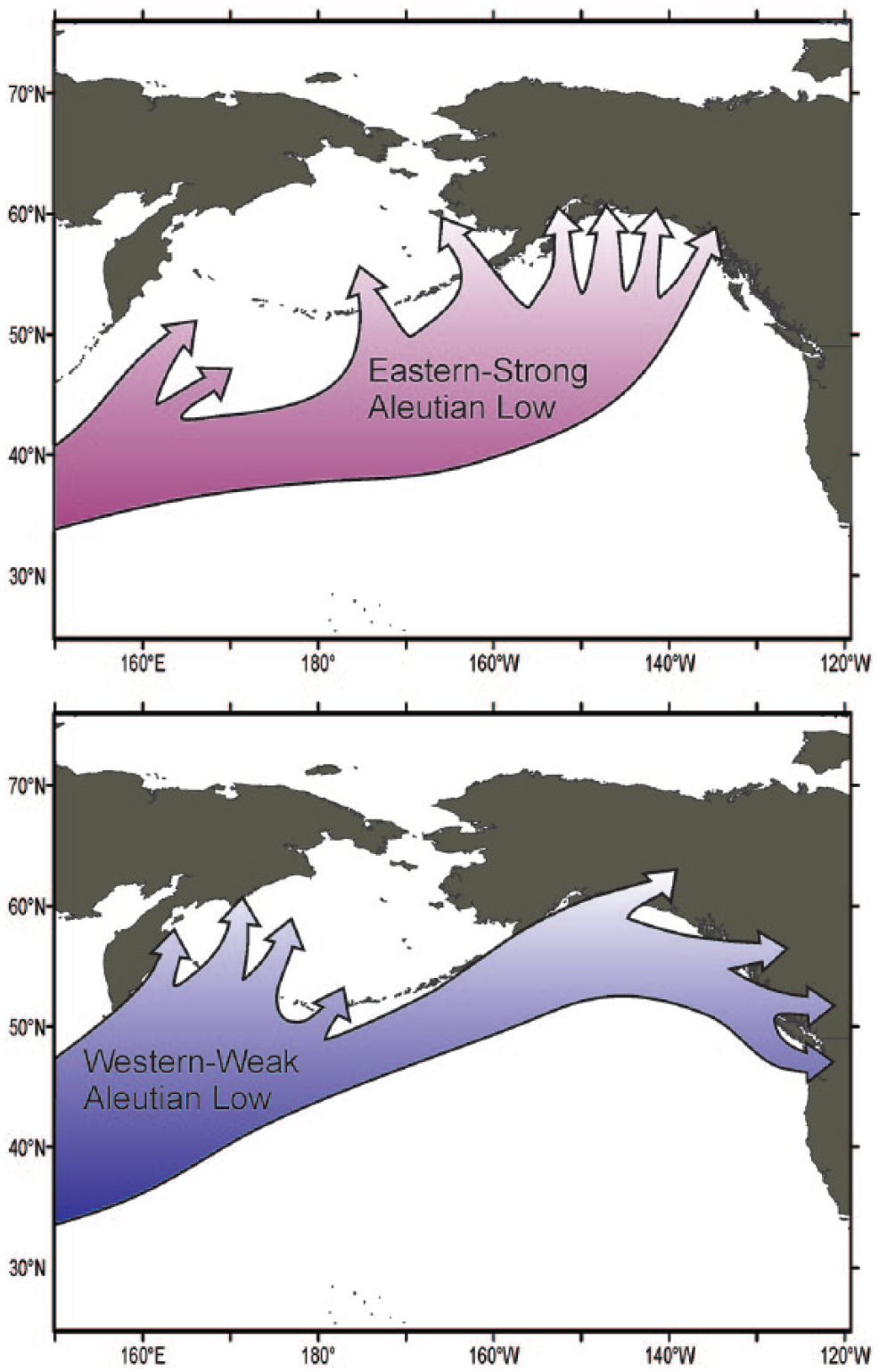
Influence of the position and intensity of the Aleutian Low (AL) on the routing of winter storms (after Rodionov et al., 2005). Upper figure shows the eastern/strong AL, with typical storms routing mostly north into Alaska. Lower figure shows western/weak AL with storms routing into mostly west into the southern Canada and northwestern United States.
Large-scale atmospheric climate patterns have a demonstrated relationship with physical and biological processes in lakes (Blenckner, 2005; Blenckner et al., 2007; Straile and Adrian, 2000; Straile et al., 2003). Regionally coherent lake changes in response to atmospheric circulation patterns are evident in the records of surface water temperature, lake-ice phenology, and lake level in long-term lake monitoring studies (Magnuson et al., 1990, 2004, 2006). This is because physical processes in lakes often respond to solar irradiation, air temperature, relative humidity, and wind speed, which express a high level of spatial coherence due to large-scale atmospheric circulation patterns (Jones et al., 1997; Livingstone, 2008; Magnuson et al., 1990). These observations have led to a shift in perspective, from lakes primarily acting as recorders of local climate to a new paradigm of lakes serving as archives of large-scale climate forcings modified to different extents by local climate and individual lake characteristics (Adrian et al., 2009; Blenckner, 2005; Livingstone, 2008; Williamson et al., 2009).
Potential multi-decadal state changes in the AL have been identified from several paleoclimate records from the northwestern Pacific region (Anderson et al., 2005; Barron and Anderson, 2011; Chipman et al., 2012; Edwards et al., 2001; Fisher et al., 2004, 2008; Latif and Barnett, 1994). However, despite the broad region where climate is influenced by the AL, few paleolimnological records outside of the Pacific Rim display a synchronous or coherent late Holocene response. In North America, abrupt, high-magnitude climate events observed at low resolution in Holocene lake records may share broadly similar sequential patterns, but upon closer inspection are time-transgressive in nature (Walker and Pellatt, 2008; Williams et al., 2010). Long-term, high-resolution paleolimnologic archives with a high degree of regional coherence are relatively uncommon (Fritz, 2008; Fritz et al., 2000; Laird et al., 2003, 2007). This has in part been attributed to the complex and commonly non-linear ecological response of the biological indicators that are used to reconstruct past conditions (Fritz, 1996). But many other factors, such as differences in the physical, hydrological, chemical, or trophic setting of lakes; quality of age control; landscape position; and local climate influences can contribute to the difficulty of interpreting environmental history from multiple sites (Blenckner, 2005; Fritz, 2008; Webster et al., 2000; Williams et al., 2010). Interannually, climatic changes related to the strength and position of the AL likely influence the thermal structure of lakes, primarily by changing the timing and nature of lake mixing and stratification patterns (Ragotzkie, 1978). For example, when the AL intensifies and shifts eastward (e.g. a +PDO), the Pacific Northwest and northern Rockies regions experience warmer winter temperatures (Wise, 2012), which can result in fewer freezing days, more precipitation falling as rain instead of snow, decreased snow cover, and warmer spring air temperatures. Because the intensity and position of the AL also influences storm track position (Rodionov et al., 2007; Wise, 2012), increased wind frequency and intensity from storms may directly influence the convective mixing energy of the epilimnion affecting the depth of the thermocline and the timing of destratification (Ragotzkie, 1978).
The ecology of lake plankton is ‘tuned’ to these seasonal changes because altering the thermal structure of lakes affects nutrient cycling, physiological processes, light availability, and trophic interactions (Blenckner et al., 2007; Dröscher et al., 2009; Gerten and Adrian, 2001; Straile and Adrian, 2000). As a result, over longer periods we expect changes in the thermal structure of a lake to have an observable influence on the structure and composition of lake plankton – as recorded in their sediment records as shifts in fossil assemblages. Diatoms are single-celled golden-brown algae, with a siliceous cell wall that commonly preserves in lake sediment, found in most aquatic systems. Diatoms are highly sensitive to environmental variables and exhibit substantial capacity for exploiting ecological niches in an exceptional range of aquatic habitats (Smol and Stoermer, 2010). In particular, in lakes with low phosphorus concentrations and elevated alkalinities (Saros and Anderson, 2015), small centric diatom species of the genus Lindavia (Cyclotella) (and closely related genera, such as Discostella) are inferred to have an ecological advantage over larger diameter congeners and dense colonial plankton with higher nutrient requirements, such as Aulacoseira, when a lake is strongly stratified (Saros and Anderson, 2015; Smol et al., 2005; Wang et al., 2008). In this paper, we use several common diatom species from lakes in the northern and central US Rocky Mountain region to investigate changes in lake stratification through the late Holocene. Using fossil diatom records from three lakes located in the US Rocky Mountains, we explore long-term changes in the AL as a possible explanation for coherent regional responses observed in lake thermal structures and ecosystems of these lakes over the last 4000 years.
Regional setting
The lakes for this study were selected based on the abundance of cyclotelloid diatom species in the plankton (and fossil records), which were the focus of the neoecological experiments discussed in Saros et al. (2012). They can be grossly characterized as deep (>30 m) subalpine to alpine lakes with low nutrient concentrations and low sediment turbidity (lacking substantial glacial inputs). When possible, the sites were selected to optimize the influence of wind intensities (large surface areas, long axes along major wind directions, few wind shelters). All three lakes have low sedimentation rates, averaging ~0.015 cm/yr.
Beauty Lake (44°58′12.12″N, 109°34′21.08″W, 2877 m a.s.l.) and Emerald Lake (44°59′49.31″N, 109°31′33.81″W, 2975 m a.s.l.) are deep (>30 m) alpine lakes with relatively small surface areas. Both lakes are located within the Beartooth Mountain Range, just south of the Wyoming–Montana border (Figure 2). The lake catchments are characterized by sparse vegetation, shallow soils, and large expanses of exposed Precambrian granitic bedrock. Because of slow weathering rates, lakes in this region are typically dilute with low silicon (Si) and phosphorus (P) concentrations. These lakes are usually ice-free from early July through October (Saros et al., 2003), similar to most of the lakes in this region. Upper Kintla Lake (48°57′14.25″N, 114°19′9.50″W, 1219 m a.s.l.) is located in the northern-most part of Glacier National Park, very near the US–Canada border (Figure 2). Upper Kintla Lake sits at a lower elevation and has a greater surface area, with a long, linear basin characteristic of a drowned river valley. Upper Kintla Lake has nutrient and water chemistry characteristics that are typical of many lower elevation lakes in the northern part of Glacier National Park (see supplemental information, available online; Saros et al., 2010).
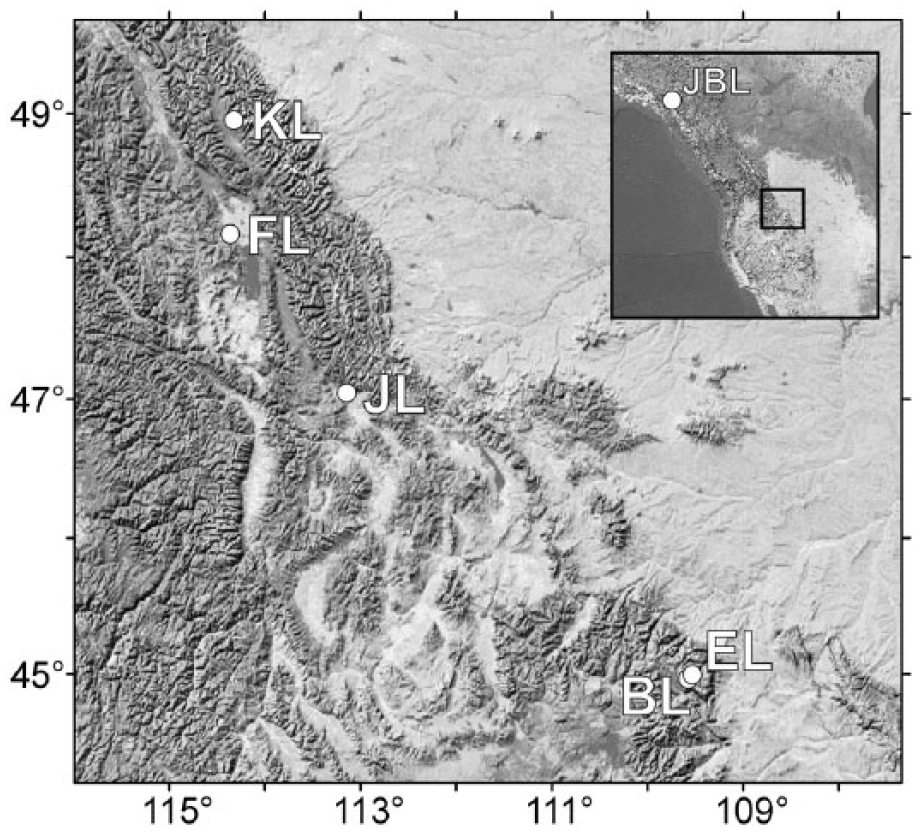
Map showing the location of the study lakes: Emerald (EL), Beauty (BL), and Upper Kintla (KL). Additional lake sites marked on the map and inset are discussed in the text: Foy (FL), Jones (JL), and Jellybean (JBL).
Methods
Age model
Sediment cores from Upper Kintla, Beauty, and Emerald Lakes were collected from ~30 m below the lake surface with a simple piston (Griffith) corer during the summer of 2007. Because Upper Kintla Lake is approximately 56 m at the deepest point, sediment cores were collected from a region nearer to the margin of the lake, whereas the other sediment cores were collected nearer to the center. Each core captures a continuous sequence of sediment ranging from 0.75 to 1.35 m in length (Table 1). Cores were sub-sampled in the field in contiguous 0.5-cm increments from the sediment–water interface to the bottom of the core. Age models (Figure 3) for each core were constructed using 210Pb-dating (Appleby and Oldfield, 1978) of sediment (except Upper Kintla), and 14C-dating of wood fragments and pine needles was recovered from the cores (Table 1) using the program BACON (2.2) (Blaauw and Christen, 2011). All radiocarbon dates were calibrated to calendar years using CALIB 6.0 (Stuiver and Reimer, 1993). All ages reported herein as ‘ka’ are calibrated ages in thousands of years before 1950.
Chronology data and materials used to create the BACON age models. 210Pb ages and error presented in calendar years (before 1950); 14C ages and error presented in uncalibrated radiocarbon years.


BACON age–depth models for Beauty, Emerald, and Upper Kintla Lakes. Gray envelope indicates 95% confidence.
Fossil diatom analysis
Sub-samples were treated with 10% HCl and 35% H2O2 at room temperature to digest carbonate and organic material. Known quantities of polystyrene microspheres were added to estimate diatom concentrations (Battarbee, 2003); diatom extractions were dried onto coverslips and mounted in Zrax, a permanent high-refractive-index medium. Diatom frustules were identified to the most specific taxonomic level possible with 1000× magnification light microscopy using a Zeiss Axioskop 2 with Differential Interference Contrast optical illumination. When possible, at least 300 diatom valves were identified from each sample interval. The sediment core from Upper Kintla Lake was collected nearer to the shoreline, so planktic diatoms were typically a smaller component of the fossil assemblage. To adjust for this, a stratified counting technique was employed, where after 300 diatoms were identified, enumeration continued, ignoring benthic diatom forms until at least 100 planktic diatom valves were identified. All relative abundance data present herein from Upper Kintla Lake are reported as percentage of plankton. Data from Emerald and Beauty Lakes are presented as percentage of the total diatom assemblage. However, converting the datasets for these two lakes to percentage of plankton produced no significant changes in the diatom stratigraphy.
Diatom-inferred stratification index
Most Aulacoseira employ meroplanktic life strategies; they form long, heavy threadlike colonies that sink rapidly and encyst during stratified lake conditions (Jewson, 1992; Kilham, 1990). Aulacoseira lirata and Aulacoseira alpigena are a common component of deeper alpine and subalpine lakes with limited phosphorus concentrations and relatively high dissolved silica concentrations. As a result, these species typically are more successful in very well-mixed lake settings, where convective mixing in stratified lakes enhances the advection of nutrients from the hypolimnion and helps them to maintain their position in the photic zone (Carrick et al., 1993; Lund, 1954; Rühland et al., 2003).
Emerald and Beauty Lakes had fossil plankton diatom assemblages comprising the cyclotelloid species (Lindavia bodanica, Lindavia comensis, Discostella stelligera) that were the subject of experiments in Saros et al. (2012) and abundant populations of Aulacoseira species (A. lirata, Aulacoseira italica, A. alpigena). The only large-diameter cyclotelloid found in the fossil assemblages was L. bodanica; we inferred a relatively deep thermocline (~14 m) from the presence of this species (Noble et al., 2013; Saros and Anderson, 2015; Saros et al., 2012). Small-diameter cyclotelloids, including the species D. stelligera, Lindavia ocellata, and L. comensis were present in the fossil diatom assemblages of these three lakes (Figure 4). Evidence suggests that in temperate regions these small-sized cyclotelloid species often out-compete other plankton when lakes are strongly stratified (Rühland et al., 2008; Saros and Anderson, 2015; Saros et al., 2012; Smol et al., 2005; Winder et al., 2009), and thus, we infer shallowest mixing depths when these species are abundant in fossil diatom assemblages.

Stratigraphic profiles of the fossil diatom plankton of Emerald, Beauty, and Upper Kintla Lakes, plotted as relative abundances, for the past 4.5 ka. Stratigraphic columns are shaded from dark to light based on the relative inferred mixing depth of the diatom species. Note that Upper Kintla Lake values are based on stratified counts of planktic diatoms.
Using this information together with the spectrum of optimal mixing depths (depth to the thermocline) resulting from ecological assay experiments for lakes that fit the appropriate nutrient criteria (Saros and Anderson, 2015; Saros et al., 2012), we created a Diatom-Inferred Stratification Index (DI-SI) that ranges from well mixed to strongly stratified for each of our three lakes. The DI-SI is based upon the concept that Aulacoseira species indicate a deeper state of mixing and a clearly higher trophic state than Lindavia (Cyclotella) in the instances where they occur in these lakes and that abundant L. bodanica indicates a deeper mixing state than when smaller cyclotelloid species such as L. comensis or L. ocellata are abundant. In the sediment record, the remains of these organisms occur together as assemblages, but taken as a whole they indicate the average stratification status of the lake through time.
The DI-SI we applied was a simple ratio expressed by the equation:

where Mk is the sum of the relative abundances of diatom species in the assemblage with the deepest inferred mixing depths and Mj is the sum of the species with the shallowest inferred mixing depths. The indices from all three records were standardized to average values over the record presented (Figure 5). A similar rationale and index (Smol et al., 2005; Wang et al., 2008, 2012) has been used previously to infer changes in the thermal structure of lakes. Non-parametric correlations between lakes were analyzed on resampled stratification index data using Spearman’s rank correlation coefficient.
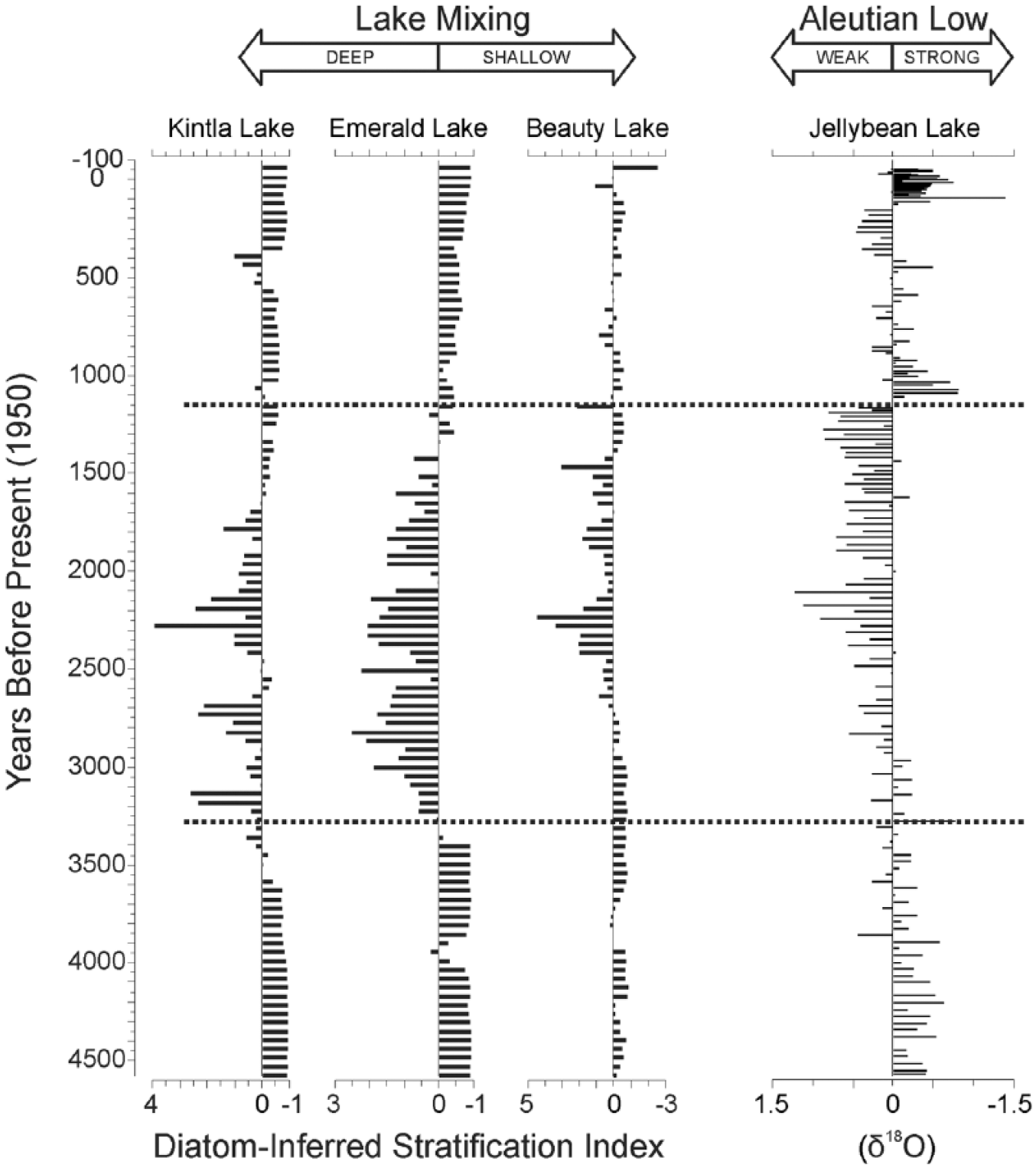
Diatom-Inferred Stratification Index (DI-SI) for Emerald, Beauty, and Upper Kintla Lakes plotted as standardized values (0 = average mixing depth for each lake) compared with the δ18O anomaly for Jellybean Lake (Anderson et al. 2005). The dotted lines mark major transitions in the inferred relative depth of mixing and corresponding changes in the position of the Aleutian Low as inferred from changes in the δ18O from the Jellybean Lake record. There appears to be a strong relationship between the weakened (westward positioned) Aleutian Low and deep mixing of the lakes between 3.2 and 1.4 ka.
Results
The long-term patterns of the DI-SI are coherent between the lakes (Figure 5). Prior to 3.2 ka, the DI-SI was consistently less than 1 for all three lakes, indicating that species that fare better under stronger stratification dominated the plankton assemblages of each lake. In Emerald and Beauty Lakes, this is represented by the dominance of D. stelligera and in Upper Kintla Lake by elevated percentages of L. ocellata. Lower abundances of species preferring deeper mixing (Aulacoseira spp. and L. bodanica) are present throughout the records of all three lakes (Figure 4). Pair-wise non-parametric correlations between each of the DI-SI records are statistically significant (Upper Kintla/Beauty: ρ = 0.31, p = 0.0013; Upper Kintla/Emerald: ρ = 0.78, p < 0.0001; Beauty/Emerald: ρ = 0.44, p < 0.0001).
Between 3.2 and ~2.3 ka, the records from all three lakes indicate periods of increasingly deeper lake mixing (Figure 5). Some minor discrepancies exist between the timing and degree of persistence in mixing depth state, which may be related to age model calculations, but the overall trend of increasing mixing depths (with respect to the previous period) exists for the records of all three lakes throughout this period (Figure 5). In the records from Emerald and Beauty Lakes, this change is represented by an increase in Aulacoseira species, replacing D. stelligera, whereas in the record from Upper Kintla Lake, L. ocellata is replaced by L. bodanica (Figure 4). From 2.3 to 1.4 ka, this trend gradually reverses for the records from all three lakes, when the relative abundances of the major planktic diatoms in the fossil assemblages show unilateral changes that are opposite to the prior transition.
After 1.4 ka all lake records show a return to enhanced stratification. With the exception of a few brief intervals, the DI-SI values return to values similar to the period prior to 3.2 ka (Figure 5). For each record, modern DI-SI values match or exceed the lowest values from any time in the entire 4.5 ka record. Overall, the DI-SI between the three lakes for the period after 1.4 ka can be characterized as shallower mixing depths with slightly more variability. In all our lake records, the transitions observed in the DI-SI represent a shift in the dominance of the few species we selected, but it is worth noting that particularly after 1.4 ka, the records from Upper Kintla and Beauty Lakes also include the introduction of high relative abundances (up to ~40%) of some intermediate-sized Lindavia (Cyclotella) species (L. comensis and Lindavia rossii/tripartita), neither of which are a major constituent of the fossil diatom assemblages spanning back to 4.5 ka (Figure 4). At Emerald Lake, there is a subtle, but important change as D. stelligera increases while Aulacoseira spp. decline, similar to the period prior to 3000 BP, but without A. italica.
Discussion
In all three lakes, the DI-SI at 4.5 ka is fairly similar to the 20th-century index within the same lake. Thus, with respect to the average depth of the upper mixed layer, the thermal structure of the lakes at 4.5 ka was fairly similar to today. Between approximately 3.2 and 1.4 ka, an excursion toward significantly deeper mixing occurs in all three records (Figure 5). We interpret the regional coherence, in both timing and direction from these three lakes to indicate a shift in large-scale atmospheric circulation, specifically changes in the AL, as the driving force behind these changes. The elevation, basin morphometry, vegetation and soil development, aspect, and surface area between these three lakes differ substantially, but the lakes show remarkably similar patterns of inferred lake mixing over the late Holocene, suggesting a large-scale forcing mechanism.
Since few studies have explicitly explored changes in lake thermal structure over the Holocene, it is unclear exactly how these changes compare against other records in western North America. However, the general patterns of inferred changes in thermal structure of Upper Kintla, Beauty, and Emerald Lakes closely follow the pattern of the oxygen isotopic record of Jellybean Lake in western Canada (Anderson et al., 2005), which is inferred to track changes in moisture source associated with changes in the position and intensity of the AL through the Holocene (Figure 5). Pair-wise non-parametric correlations between the Rocky Mountain lake records and the record from Jellybean Lake indicate coherence in all cases (ρ = 0.47–0.57 for pairwise comparisons between Jellybean Lake and each lake in this study, p < 0.0001 in all cases). Anderson et al. (2005) suggested the period 4.5–3 ka represented a persistent intense state of the AL, similar to that experienced today, and that the period ~3–1.2 ka represented a weaker (or more westward position) AL. After ~1.2 ka, the AL returned to a more eastward/intense state, but exhibited greater overall variability than previous periods. Our lake records show a change in thermal structure that may be driven by these same changes in the intensity and position of the AL through the Holocene, and they demonstrate that the influence of the AL may be responsible for broadly coherent Holocene patterns in lake records across parts of western North America.
Based on modern atmospheric circulation patterns, a persistently weakened AL would enhance upper atmosphere westerly flow into the Rocky Mountain region throughout winter and spring, which would result in increased cool season precipitation (Overland et al., 1999; Rodionov et al., 2005). We expect this would have two compound and synergistic effects on the thermal structure of lakes in this region. First, enhanced storm activity through the winter months would increase the total snowfall. A thicker snow bed over lake ice results in a higher albedo for the lake surface and typically acts to extend the ice cover duration (Ragotzkie, 1978). Over long periods this would result in a net loss of heat storage in the lake and act to weaken the average stratification strength by decreasing the total seasonal irradiance to which epilimnetic waters are exposed. For phytoplankton, whose seasonal succession patterns are substantially influenced by stratification and its influence on the nutrient cycle in lakes, this probably means a truncation of the stratification cycle, with fewer days of highly stratified epilimnetic water in the late summer. Second, increased storm activity through the spring would increase average near-surface wind intensity. Since stratification depth is primarily controlled by a combination of irradiance, light penetration, and wind intensity (Blenckner et al., 2007; Kirillin, 2003; Kirillin et al., 2012; Tanentzap et al., 2008), these changes should result in a change in how deeply the lakes mix. More intense wind in the spring would delay onset of lake stratification, creating a prolonged spring-like condition for the lake. Aulacoseira have a demonstrated relationship between wind speed and total production (Carrick et al., 1993), and enhanced near-surface wind speeds would also likely result in greater total productivity for deeper mixing species.
In clear lakes, with a small surface area (<500 ha), locally driven changes in transparency resulting from variability in dissolved organic matter can have a large influence on thermal structure (Fee et al., 1996; Houser, 2006; Snucins and Gunn, 2000). Since temperature and precipitation can impact the influx of dissolved organic matter, this may explain some of the minor differences between the timing of transitions between lakes. However, Upper Kintla Lake is considerably larger, deeper, and at a substantially lower elevation than Beauty and Emerald Lakes, yet shows the same general pattern over the last 4.5 ka. There appears to be a stronger correlation between Emerald and Upper Kintla Lakes than between Emerald and Beauty Lakes (Figure 5.), which are much closer to each other. We believe this is explained by the differences in wind exposure between the three sites; Upper Kintla is located in a long narrow valley where wind intensity can be quite high along the long axis of the lake, and Emerald Lake is located high in the Beartooth Mountains with few trees to limit exposure to wind. In contrast, Beauty Lake is located in a more protected valley site.
Over the last century, the multi-decadal frequency of the AL, as expressed by the NPI, is nearly identical to frequency of the PDO (Miller et al., 2004; Minobe, 1999), which has been cited as a major influence on effective moisture in the region. Evidence of greatly increased effective moisture matching the timing of the inferred weakening of the AL (~3 ka) from the Jellybean Lake record exists in the lake records from Foy and Jones Lakes (Figure 2), located near our study sites (Shapley et al., 2009; Shuman et al., 2009). These studies infer major changes in water chemistry and rising lake levels after 3 ka, with conditions close to those of the modern climate after 1.4 ka. Throughout the Pacific Northwest, widespread glacial advances in alpine settings begin around 3 ka (Osborn and Luckman, 1988; Walker and Pellatt, 2008). Enhanced winter and spring precipitation in the Rocky Mountain region, as might be expected from a weakened AL, would create ideal conditions for glacial advances.
Conclusion
Our findings demonstrate the utility and strength of pairing diatom neoecological experiments with paleoecological studies within the same lake to improve our ability to interpret paleolimnological records. Despite significant differences in basin morphology and elevation, we have used these techniques to extract a simple, yet robust index from a complex diatom paleoecological dataset, which has allowed us to identify three distinct, regionally coherent periods in the late Holocene thermal structure. These periods broadly correspond to reconstructed patterns of the AL from independent records and suggest a widespread influence of the AL over winter wind intensity and precipitation patterns in the late Holocene throughout the Rocky Mountain region. This technique likely holds great promise for using phytoplankton from lake records to reconstruct past changes in lake thermal structure and potentially other major modes of atmospheric circulation. Through careful site selection, this approach has the potential to reconstruct long-term changes in wind intensity, air temperature, and lake-ice phenology.
Footnotes
Acknowledgements
We thank the following people for field assistance: KS Westover, AB Nurse, A Bamberg, E Wilcox, C Kissman, and CR Wigdahl. S Wooden assisted with lab processing of the diatom slides. Lab preparations and microscope analyses were performed at the Fritz Lab (University of Nebraska-Lincoln). We thank the following people for discussion and comments that improved the manuscript: CR Wigdahl, KE Strock, and K Slemmons. This draft manuscript is distributed solely for purposes of scientific peer review. Its content is deliberative and predecisional, so it must not be disclosed or released by reviewers. Because the manuscript has not yet been approved for publication by the U.S. Geological Survey (USGS), it does not represent any official USGS finding or policy.
Funding
Funding for this work was provided by the US National Science Foundation Division of Environmental Biology Program (Grant #DEB-0734277). Any use of trade, firm, or product names is for descriptive purposes only and does not imply endorsement by the US Government.