
Abstract
Major Mediterranean deltas began to develop during a period between 8000 and 6000 yr BP when the rate of fluvial sediment input overtook the declining rate of sea-level rise. However, different authors have argued that the Ebro Delta primarily formed during the late Middle Ages as a consequence of increased anthropogenic pressure on its river basin and these arguments are supported by the scarcity of previous geological studies and available radiocarbon dates. To reconstruct the environmental evolution of the Ebro Delta during the Holocene, we used micropalaeontological analysis of continuous boreholes drilled in two different locations (Carlet and Sant Jaume) on the central delta plain. Different lithofacies distributions and associated environments of deposition were defined based on diagnostic foraminiferal assemblages and the application of a palaeowater-depth transfer function. The more landward Carlet sequence shows an older and more proximal progradational delta with a sedimentary record composed of inner bay, lagoonal and beach materials deposited between 7600 and >2000 yr BP under rising sea-level and highstand conditions. This phase was followed by a series of delta plain environments reflected in part by the Carlet deposits that formed before 2000 yr BP. The Sant Jaume borehole is located closer to the present coastline and contains a much younger sequence that accumulated in the last 2.0 ka during the development of three different deltaic lobes under highstand sea-level conditions. The results of this study reinforce the idea that the Ebro Delta dates to the early Holocene, similar to other large Mediterranean deltas.
Keywords
Introduction
Deltas are the largest coastal landforms in the world (Evans, 2012). These coastal depositional environments result from the interaction between river and marine forces. The strength of both elements determines the dominant processes governing the evolution of deltaic systems through time (Jiménez et al., 1997). The morphology and sedimentary architecture of deltas depend on the relative magnitude of tides, waves and currents (Galloway, 1975; Wright and Coleman, 1973). The morphological evolution of a delta is also controlled by relative sea-level changes, which in turn depend on the eustatic sea-level rise and local subsidence or uplift (Galloway, 1975). Subsidence results naturally from the compaction of deltaic sediments, degassing of peats and growth faults developed at the base of deltaic sediments; however, subsidence can be additionally increased by human activities, such as the extraction of groundwater. Relative sea-level rise (RSLR) can be compensated by vertical accretion processes taking place in the delta plain. The accretion rates depend on fluvial sediment inputs and sea-level rise itself through feedback mechanisms (Day et al., 2011; Ibáñez et al., 2014).
Deltas are considered to be highly vulnerable to even minor changes in relative sea level, particularly because most modern deltas are actively subsiding and their sediment supply has been curtailed (Giosan et al., 2014; IPCC, 2014). Moreover, global warming is accelerating sea-level rise, which intensifies coastal erosion and land loss due to marine inundation (Fatorić and Chelleri, 2012). To implement science-based coastal protection measures in these sensitive areas, a precise definition of the relationship between sea-level change and delta evolution is critical. Consequently, it is essential to understand the relationships between delta development and sea-level changes during the Holocene (Stanley and Warne, 1994). Under natural conditions, deltas have mechanisms to enhance vertical accretion and gain land as a response to RSLR, especially via the increased river avulsion and delta lobe formation in shallow areas and the increased accretion in coastal marshes and beaches connected to marine environments (Ibáñez et al., 2014). This suggests that river-dominated deltas can be resilient to changes in sea level.
Previous analysis of major modern deltas in the Mediterranean Sea showed that these environments started to form between 8000 and 6000 yr BP. Overlying upper Pleistocene fluvial gravels, Holocene deltaic deposits consist of variable aggradational and progradational lithologies (Anthony et al., 2014; Stanley and Warne, 1994, 1997; Vella et al., 2005). These previous works showed that the deceleration in sea-level rise was the key to the initiation of delta formation and that Holocene deltaic sequences began to accumulate as the rate of fluvial sediment input overtook the declining rate of sea-level rise along the coasts. For the northwestern Mediterranean area, Lambeck and Purcell (2005) and Pirazzoli (2005) found a rapid sea-level rise until 6.0 ka, followed by a more gradual increase in sea level with a slight deceleration during the last 1.5 ka, with stabilization for the last 0.5 ka.
Recently, Maselli and Trincardi (2013) supported the idea that the onset of the northern Mediterranean deltas followed an ancestral phase dominated by estuary fill and relatively slow delta growth at approximately 6000 yr BP. Furthermore, they suggested that these deltas formed almost synchronously during two short intervals of enhanced anthropogenic pressure on the landscape during the Roman Empire and the ‘Little Ice Age’.
Mediterranean deltas contain a widespread and generally consistent Holocene stratigraphic succession composed of peat, lagoonal and other organic-rich facies that accumulated as delta plain deposits at or near sea level (Stanley and Warne, 1994). These resource-rich ecosystems were used by humans soon after their development. Documented archaeological sites dated to 7000 yr BP or earlier are positioned on or adjacent to deltas, such as those of the Rhône and the Nile (Stanley and Warne, 1997).
Benthic foraminifera have been long and widely used as indicators of past environmental conditions (salinity, temperature, oxygen content, etc.) in open marine and coastal areas (Murray, 2006) and are a valuable tool, in combination with the sedimentological analysis of the stratigraphic succession and other palaeontological and geochemical proxies, for palaeoenvironmental reconstruction. However, the use of foraminifera for an accurate reconstruction of coastal habitats is not straightforward because of the enormous complexity and variability of these ecosystems. This is of paramount importance in the case of deltas because a small delta plain such as the Ebro (320 km2) contains at least four different habitats with particular foraminiferal assemblages (Benito et al. (2015) and Tables 1 and 2). Moreover, the existing literature shows that previous palaeoenvironmental reconstructions of Mediterranean deltaic sequences have used modern analogues only from open marine environments (Amorosi et al., 1999, 2013; Carboni et al., 2010; Curzi et al., 2006; Dinelli et al., 2012; Milli et al., 2013; Rossi and Horton, 2009) and not from delta plain habitats (coastal lagoons, inner bays, salt marshes, etc.). Thus, the present study represents the first palaeoreconstruction based on both open marine and delta plain assemblages, thereby providing the possibility for a new and sounder interpretation of the fossil record in deltaic sedimentary sequences.
Living foraminiferal species found in and off the Ebro Delta. Above: quantitative summary of living foraminiferal assemblages composition in different environmental settings of the modern Ebro Delta. Figures represent relative abundance (%) unless otherwise indicated. The single value represents the average and those in parentheses give the range. Below: reference list of marine species found in the modern foraminiferal assemblages of the Ebro Delta.

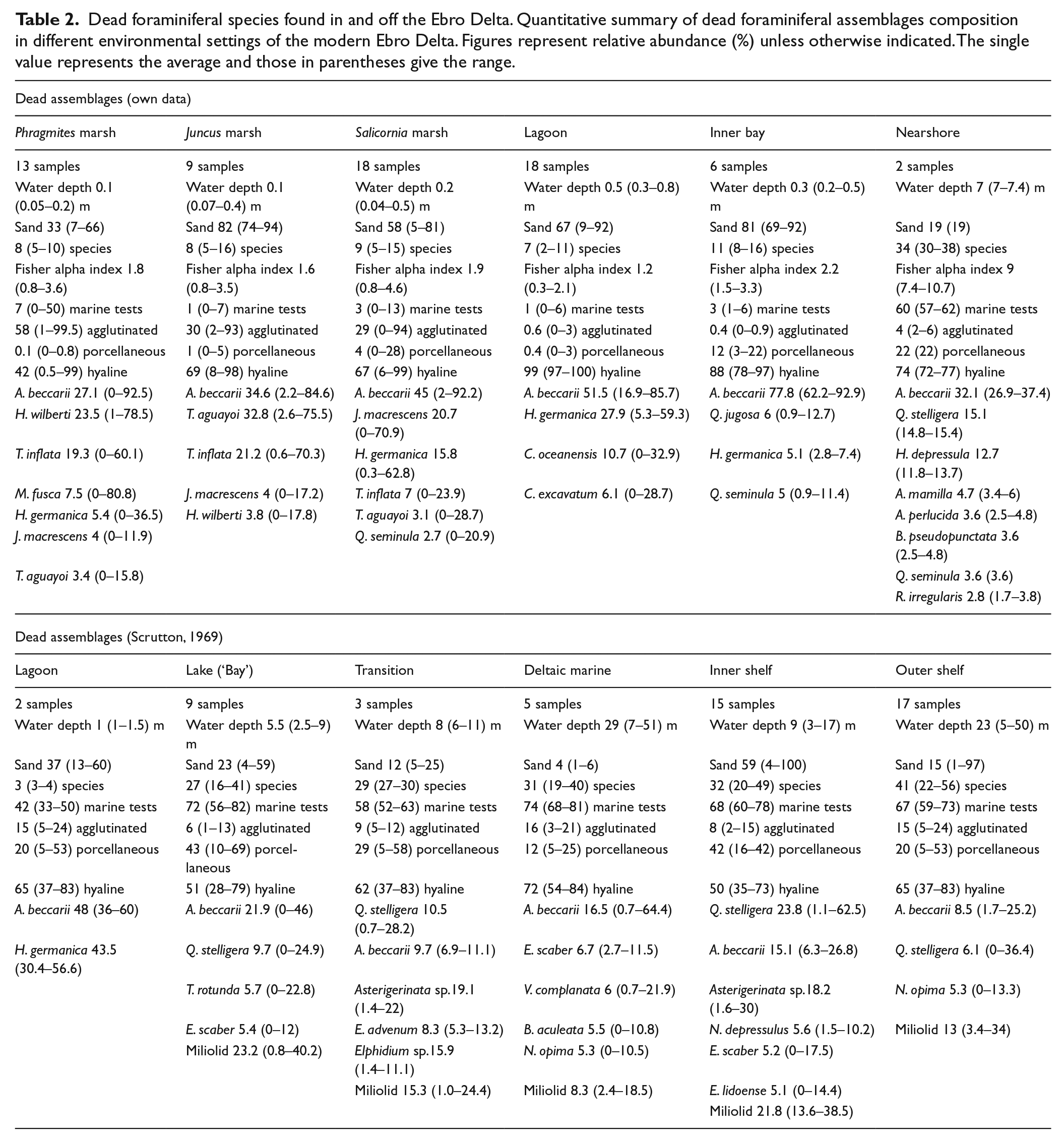
Dead foraminiferal species found in and off the Ebro Delta. Quantitative summary of dead foraminiferal assemblages composition in different environmental settings of the modern Ebro Delta. Figures represent relative abundance (%) unless otherwise indicated. The single value represents the average and those in parentheses give the range.
Previous work
Earlier geological studies of the Ebro Delta and its Holocene sedimentary archives and evolution are scarce and most of them are supported by just a few radiocarbon dates, especially initial works carried out in 1960s and 1970s (e.g. Macau, 1961; Maldonado, 1972; Maldonado and Riba, 1971; Solé et al., 1961). According to Maldonado and Murray (1975) and a sedimentological and palaeontological comparison between borehole sedimentary successions and recent environments, after a temporary stabilization of sea level at ca. −10 m extensive deltaic progradation started, and the delta plain formed over the last 8.0 ka (based on peat material radiocarbon dated by Solé et al. (1965) to 7680 yr BP). The fluvial supply of sediment was sufficient to prevent extensive transgression over the delta plain during this time interval, which was concurrent with the slowing of the sea-level rise. Maldonado and Murray (1975) concluded that the entire development of the Ebro Delta was governed mainly by the interaction between the rate of sea-level rise and the rate of sediment delivery by the river. Other factors, such as river floods and coastal processes caused the diversion of the distributaries as well as changes in the delta morphology, with river avulsions being responsible for evolution of the delta through sequential progradation and abandonment of different deltaic lobes, which were subsequently modified by rapid subsidence. Thus, the geometry of Ebro Delta was created by the advance of successive deltaic lobes that prograded radially seawards from an avulsion point usually located close to Gracia Island (Maldonado and Riba, 1971). These processes were studied by Maldonado (1977) who found evidence for five different major avulsions during the last centuries (Díaz et al., 1990).
The Holocene deposits of the delta present a thickness ranging from 20 m on the landward side to 52 m on the delta front (Maestro et al., 2002; Maldonado, 1972). Radiocarbon ages indicate that deposition of the prodelta on the shelf began at approximately 11,000–10,000 yr BP (Díaz et al., 1990). After the pioneering work of Maldonado (1972), Somoza et al. (1998) published the most comprehensive study of the Holocene depositional units of the Ebro Delta based on high-resolution seismic profiles and analysis of 11 existing boreholes (20–60 m long) drilled on the modern delta (3 in the alluvial valley, 7 in the delta plain and 1 in the prodelta). The Holocene deposits of the delta were interpreted as a depositional sequence composed of a transgressive systems tract (TST), composed mainly of a basal mollusc-shell lag and marine grey or black clays overlying the upper Pleistocene gravels, and a highstand systems tract (HST). The top of the maximum flooding surface (MFS) separating the TST from the HST was dated to 6900 yr BP based on peat material from the inner delta area previously published by Arasa (1994), as they did not obtain any direct dates from their sedimentary sequences. The HST, which overlies the MFS, is reported to include a total of five progradational units made of prodelta clays, delta-front sands or delta plain silty sands, depending on the location within the deltaic three-dimensional architecture. Only their ITGE-6 borehole was drilled in the central area of the delta plain near Gracia Island (Figure 1). The ITGE-6 borehole was approximately 30 m long, of which approximately 27 m was Holocene in age. The Holocene stack was interpreted to be composed of five progradational units (with assigned ages in Somoza and Rodríguez-Santalla, 2014). Basal unit d0 is composed of bioclastic coarse sand containing marine molluscs and deposited during the TST before 7000 yr BP. Units d1 and d2 are composed of coarse and medium sands that accumulated above the MFS and are dated to between 6150 and 3600 yr BP. These three lower units are considered to be delta-front and nearshore deposits. Finally, units d3 and d4 are characterized by sands with scattered pebbles and silty sands, which are defined as delta plain deposits younger than 2700 yr BP. These progradational units were interpreted to have been deposited as a succession of prograding delta lobes with frequencies of on the order of thousands of years.
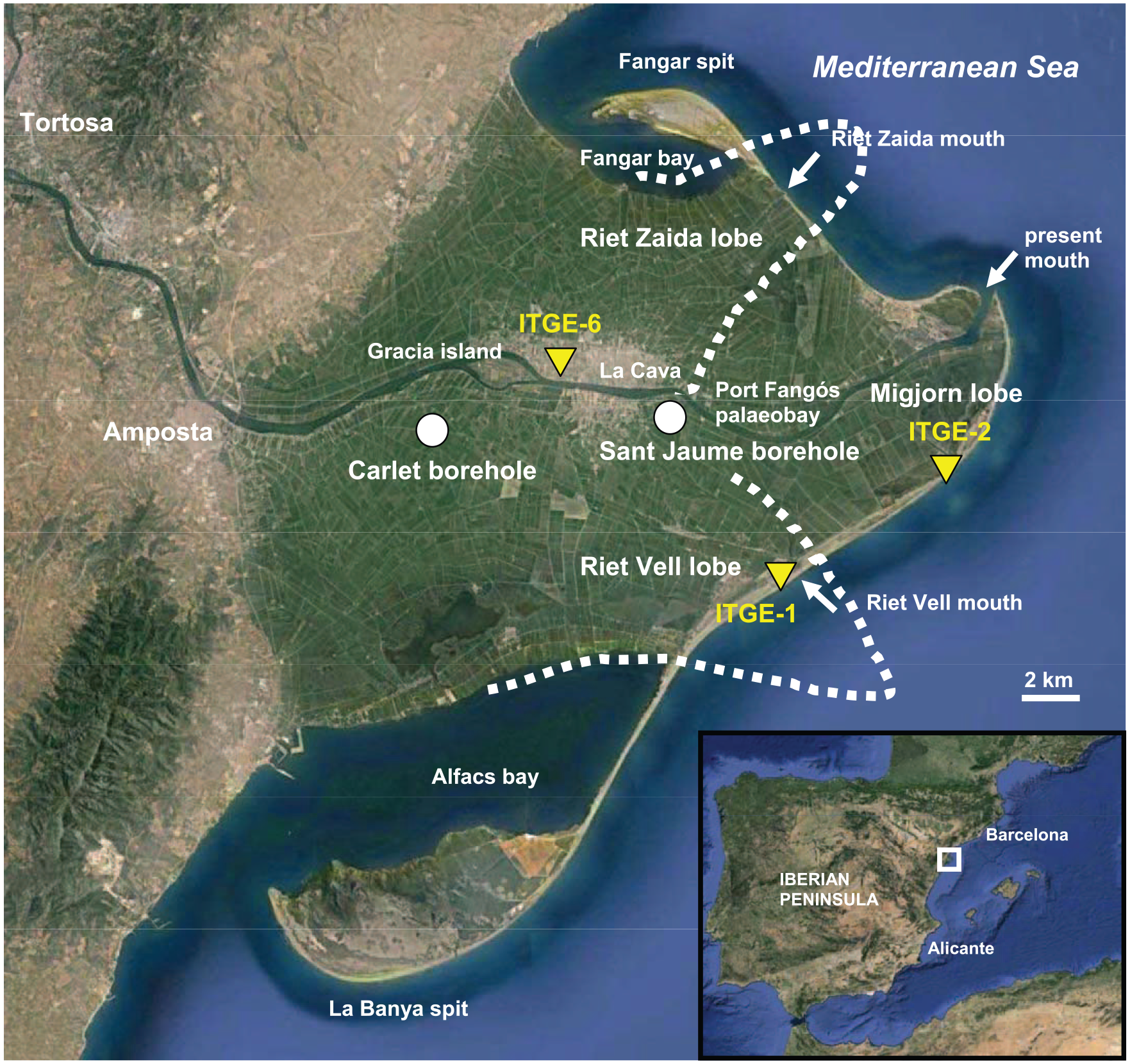
Geographical location of the Ebro Delta in the western Mediterranean Sea, boreholes and places referred to in the text. Dashed line represents the approximate morphology and position of the two historical deltaic lobes (from Maldonado, 1972). Images taken from Google Earth.
Canicio and Ibáñez (1999) identified several coastal barriers separating thick peat bodies in the landward limit of the present delta plain dated to 3050 ± 45 radiocarbon years in the northern hemidelta and 5745 ± 50 radiocarbon years in the southern hemidelta. They concluded that the orientation of the barriers suggests that at approximately 6000 yr BP the mouth of the delta was near the present fluvial island of Gracia (Figure 1).
Recently, in contrast to the above-mentioned studies, Maselli and Trincardi (2013) supported the idea that Amposta, a town now located at the inland margin of the delta, had a marine harbour during Roman times. This erroneous idea began with a personal interpretation of Roman texts by Bayerri (1934) who considered the mention of a ‘sea port’ in Tortosa (12 km upstream of Amposta; Figure 1) to be proof of the existence of an estuary. The concept of ‘sea port’, however, could also refer to fluvial ports that harbour marine vessels. As noted by Canicio and Ibáñez (1999), the same misinterpretation led some other authors to consider that the Ebro Delta formed very recently (mostly during the Islamic Period in the 14th and 15th centuries) and that it was an estuary during Roman times. For example, Guillén and Palanques (1997) and Palanques and Guillén (1998) considered that the Holocene sea-level rise caused the flooding of the river mouth, which became an estuary that evolved into a delta only during the last 2.0 ka. Some publications even concluded that the delta plain began to form during the 12th century downstream from the town of Amposta (Serra, 1997; Somoza and Rodríguez-Santalla, 2014).
Successive delta lobe progradation has been identified during the last millennium in the Ebro Delta through the recognition of three main lobes: the Riet Vell, Riet de Zaida and Migjorn lobes (Ibáñez et al., 1997) (Figure 1). The oldest map showing a relatively detailed and reliable configuration of the Ebro Delta is the Mercator–Hondius Atlas, which dates from 1580 CE (Ibáñez et al., 1997). The southeastern Riet Vell lobe was the main active mouth of the delta in 1149 CE and was probably abandoned in 1362 CE (Somoza and Rodríguez-Santalla, 2014). Modern bathymetric configuration suggests that partial marine destruction of this abandoned lobe provoked an 8-km retreat of the old headland and the subsequent growth of the southern La Banya spit (Canicio and Ibáñez, 1999). After this period, the main mouth of the Ebro Delta moved northward to the Riet de Zaida lobe, which was already active in 1575 CE (Somoza and Rodríguez-Santalla, 2014). This lobe was separated from the old Riet Vell lobe by a large palaeobay (Port Fangós), and it developed from the proximal zone of the delta, suggesting that it was built by the switching of the river near the Gracia Island (Figure 1). This new channel, shorter and with a larger hydraulic gradient to the sea, provoked a quick decay of the old Riet Vell main channel. The next detailed maps include a navigation chart of the Ebro Delta coast (Plan Des Rades de Sausa, 1733 CE) and the map of Miguel Marín (1749 CE) (Canicio and Ibáñez, 1999). The main differences with the previous situation at the end of the 16th century were the complete filling of the palaeobay that separated the two previous deltaic lobes and the rapid progradation of the new and central Migjorn lobe. This last active lobe was the result of a river switching event that took place in approximately 1666 CE at a location called La Cava near Gracia Island (Figure 1) as a consequence of a reported anthropogenic excavation in the outer levee of a pronounced meander (Ribas, 1996). The retreat of the Riet de Zaida lobe by marine erosion led to the subsequent development of the Fangar spit that started to form in approximately 1739 CE, as indicated on the Miguel Marín map (Canicio and Ibáñez, 1999). Modern maps show a rapid progradation of the central Migjorn lobe until 1880 CE because of the filling of the shallow inner palaeobay of Port Fangós, followed by a shift of the delta mouth during the 1930s and a quick retreat during the last decades because of sediment retention in the various dams located along the Ebro River watercourse (Ibáñez et al., 1997).
Another controversial issue related to the hypothesis of a recent and rapid growth of the Ebro Delta is the impact of land use changes in the river basin on the progradation rates. The 13th century is considered to be the beginning of intense deforestation in the Ebro basin because of changes in land use from forest to agricultural activities. These changes favoured sediment erosion and may have caused significant progradation of the Ebro River mouth (Maselli and Trincardi, 2013; Palanques and Guillén, 1998; Somoza and Rodríguez-Santalla, 2014). However, a recent study modelling sediment transport in the Ebro River during the last 4.0 ka (Xing et al., 2014) shows that sediment load was already high (30.5 Mt/yr) before any significant human intervention and that the increase in sediment load because of land use change was up to a maximum of 47.2 Mt/yr.
Modern foraminifera (live and dead assemblages) off the Ebro Delta were studied extensively by Scrutton (1969), who quantitatively defined the different species that characterized mainly the open marine environments. In contrast, the study of delta plain habitats was very limited (only six samples from a coastal lagoon). Afterwards, Maldonado (1972) published qualitative results on the presence of benthic foraminifera in several surface (total assemblages) and borehole samples from the Ebro Delta (from both delta plain and open marine environments). More recently, an exhaustive analysis of delta plain and nearshore foraminiferal assemblages of the Ebro Delta was carried out by the authors (see Tables 1 and 2), which complements very well the study by Scrutton (1969). Data from both studies are the basis for the palaeoreconstruction conducted in the present paper, in combination with the interpretation of stratigraphic sequences and radiocarbon dates. The geological analysis of continuous borings from deltas and their lateral correlations can define the associated environments of deposition. Such analyses provide a context for interpreting both regional palaeogeography and site-specific environmental settings.
Objectives
The present work is focused on the methodological contribution of foraminiferal assemblages to reconstructing the sequence of palaeoenvironments that characterize the Holocene evolution of the central Ebro Delta plain. We supply new data to pinpoint the age of the delta, as it has been questioned in recent years (Maselli and Trincardi, 2013). We contribute substantial new environmental and chronological information to complete and improve previous interpretations (e.g. Somoza et al., 1998).
The main aim of the current work is to provide new insights into the Holocene evolution of the Ebro Delta using micropalaeontological proxies (benthic foraminifera) based on modern analogues covering the whole range of deltaic environments (from the inner delta plain to the outer prodelta).
The combination of the fossil foraminiferal data obtained from two new boreholes with the available geological knowledge (mainly published by Somoza et al., 1998) and the existing data of modern foraminifera allowed a more precise reconstruction of deltaic environments and their evolution through time in the central delta plain area. From this knowledge, a new understanding of some controversial points regarding the origin and evolution of the delta has been obtained. In addition, numerous radiocarbon dates allowed a detailed chronology of the deltaic successions in the two boreholes to be made.
Materials and methods
Study area
The Ebro Delta is one of the largest modern deltas in the Mediterranean after those of the Nile, Rhône and Po (Barnolas et al., 1996). It is located on the western Mediterranean coast, approximately 200 km southwest of Barcelona, and it extends over an area of 320 km2. It has an outer sandy shoreline 50 km long and an estimated sedimentary volume of 28 km3 (Figure 1). The maximum tidal amplitude is 0.25 m (astronomical) and 1 m (meteorological) (Sánchez-Arcilla et al., 1996; Somoza and Rodríguez-Santalla, 2014).
The main morphological features of this microtidal delta are two spits, Fangar and La Banya, which partially close two adjacent bays, Fangar and Alfacs (Figure 1). Most of the surface area of the modern Ebro Delta has been devoted to agriculture since the construction in 1860 CE of the first irrigation canal, which transformed most wetlands and some lagoons into rice fields, which occupy 65% of the delta plain (Cardoch et al., 2002; Ibáñez et al., 2010; Roca and Villares, 2012). Natural delta habitats account for only 20% of the surface area and include fresh-water, brackish and saline lagoons, salt marshes and coastal and sandy dune systems (Valdemoro et al., 2007). Several national and international designations (RAMSAR, Natura 2000 and Natural Park) currently protect the natural delta habitats.
At present, RSLR and coastal erosion by wave action cannot be compensated by river sediment input (Ibáñez et al., 1997; Jiménez et al., 1997). A series of dams were built along the Ebro River watercourse mainly in the 1960s to support a variety of intensive water uses (Ibáñez and Prat, 2003). Irrigation and damming are responsible for a 30% decrease in the water discharge, and the reservoirs retain approximately 99% of the sediment input that would otherwise be partially deposited in the Ebro Delta, creating a severe sediment deficit (Ibáñez et al., 1996). As a result, the delta has ceased to grow, erosive processes are dominant, and it has changed from progradational to a storm wave–dominated coast that is being morphologically reshaped (Guillén and Palanques, 1992; Jiménez et al., 1997; Jiménez and Sánchez-Arcilla, 1993).
At different temporal scales, Somoza et al. (1998) estimated subsidence rates of approximately 1.75 mm/yr for the Ebro Delta during the last 7.0 ka, and Ibáñez et al. (1997) considered a subsidence of 2 mm/yr for the last 0.3 ka and recent subsidence rates to be 1–3.2 mm/yr. Recent research works combining subsidence and sea-level rise data along the Ebro Delta coast estimate a variable RSLR ranging between 2 and 6 mm/yr (Jiménez et al., 1997), and from 4 to 6 mm/yr (Ibáñez et al., 1997).
Sampling
The Carlet and Sant Jaume boreholes were drilled in 2011 in reclaimed areas of the central modern delta plain (Figure 1). The Carlet borehole is located at X 303479/Y 4508218, approximately 18 km from the modern delta mouth, and it is Z 2.33 m above the national ordnance datum and 19.27 m long. The Sant Jaume borehole is located at X 310438/Y 4508070, approximately 10.5 km from the modern delta mouth, and it is Z 1.02 m above the national ordnance datum and 21.95 m long. The depths are always referred to the Spanish national ordnance datum (mean sea level at Alicante recorded between 1870 and 1882 CE). The boreholes did not reach the basal Pleistocene gravels, although some gravels were recovered in Carlet, suggesting their proximity. They were drilled using a percussion/rotary drill that produced a core approximately 8 cm in diameter. The cores comprise alternating sands, sandy muds, muddy sands and muds with plant remains and mollusc-shell fragments in the muddy and sandy intervals.
Analyses
Foraminifera
Samples for micropalaeontological analysis were taken from the boreholes at approximately 25-cm (Carlet) and 20-cm (Sant Jaume) intervals. They were dried in an oven at 50°C and weighed. The target weight was 70 g per sample. Samples were wet sieved through 63-micron and 2-mm meshes to retain sand and gravel respectively. The samples were then dried and weighed again to determine the proportion of sand. The foraminifera were concentrated using trichloroethylene. Samples were split into fractions using a splitter, and tests were picked until a representative amount of more than 300 individuals for each assemblage was obtained. Otherwise, all the available tests were picked and studied under a stereoscopic binocular microscope using reflected light. Only assemblages with more than 100 tests were used for calculations. Altogether, 143 samples were studied (Table S1; the data table can be found online on the Holocene website http://hol.sagepub.com/), and more than 21,850 foraminifera grouped into 113 different species were identified (Appendix 1).
The species were divided into deltaic and marine forms based on modern distributions and abundance of living foraminiferal assemblages in the Ebro Delta environments based on a quantitative study by Scrutton (1969) and our own data (Table 1). Dead foraminiferal assemblages were also characterized from modern samples and the results used as modern analogues for the interpretation of the fossil foraminiferal record (Table 2).
Radiocarbon dating
A total of 39 samples of shell fragments and three samples of wood were radiocarbon dated. Radiometric analyses were carried out by Beta Analytic Inc. (Miami, US) and NOSAMS (Woods Hole, US) using Accelerator Mass Spectrometry (AMS). The radiocarbon ages of shells were adjusted for the marine reservoir with a local deltaR correction, and conversion of all dates into calendar years was performed using 2013 calibration databases (Reimer et al., 2013) (Table 3).
Radiocarbon dates from the Carlet and Sant Jaume boreholes (Ebro Delta).
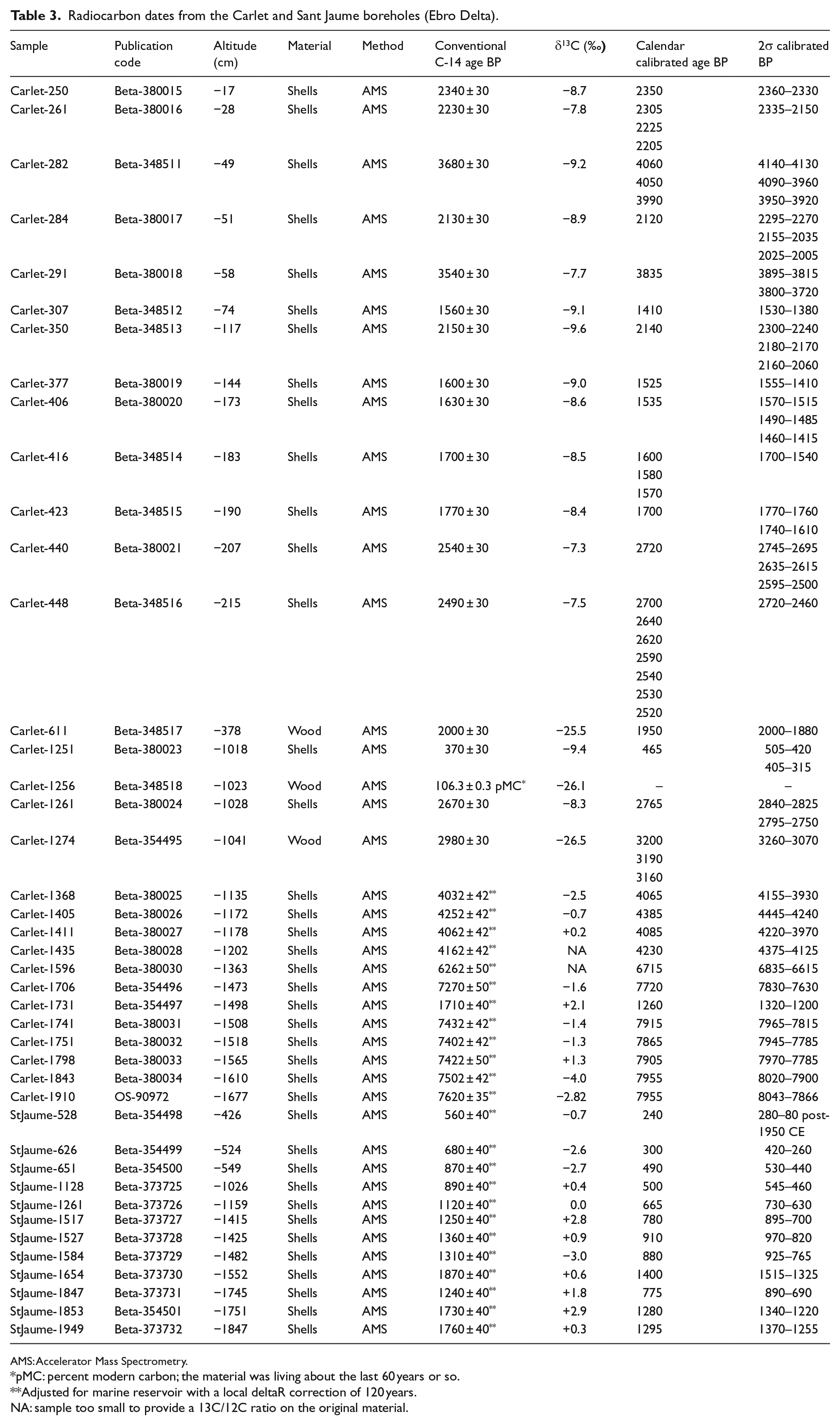
AMS: Accelerator Mass Spectrometry.
pMC: percent modern carbon; the material was living about the last 60 years or so.
Adjusted for marine reservoir with a local deltaR correction of 120 years.
NA: sample too small to provide a 13C/12C ratio on the original material.
Statistical analyses
Fisher’s alpha index was calculated for foraminiferal assemblages with >100 tests (Murray, 2006) to explore their diversity. Based on the alpha values, a clear boundary can be drawn between normal marine environments (alpha > 5) and restricted marginal marine environments (alpha < 5).
A Detrended Correspondence Analysis (DCA) was used to determine whether fossil foraminiferal assemblages were represented by the modern Ebro Delta habitats. This was performed by passively plotting onto the same unconstrained ordination space modern (dead) foraminiferal samples along with the borehole samples. Analyses were performed using the ‘vegan’ package of R (Oksanen et al., 2013).
The Linear Discriminant Function (LDF) technique was used to statistically assign each fossil sample to the most likely modern habitat group. A total of four modern habitat groups based on cluster analyses of dead foraminiferal samples were identified in the Ebro Delta: (1) offshore, (2) nearshore and outer bays, (3) coastal lagoons and inner bays, and (4) salt and brackish marshes. The LDFs estimated the probability (0–1) that a borehole sample should be classified into each of the four modern habitat groups by means of discriminant functions. The relative abundances of the foraminiferal data were square root transformed prior to the analyses to stabilize their variance. Following Kemp et al. (2012), samples with probabilities of >0.95 are considered exclusive to one habitat group, whereas those with probabilities of <0.95 samples can be assigned to more than one group. The LDF analyses were carried out using the ‘MASS’ package of R (Venables and Ripley, 2002).
The Modern Analogue Technique (MAT) was used to test the reliability of the palaeowater-depth reconstructions based on the transfer function developed by the authors. This technique evaluates the degree of similarity (or dissimilarity) in the foraminiferal assemblages between each fossil sample and the modern assemblages. The squared chord distance (SCD) was used as dissimilarity coefficient. Using the largest dissimilarity coefficient among all the modern foraminiferal samples as a critical threshold (Woodroffe, 2009), we identified fossil samples with close modern analogues. Samples with SCDs of ⩽0.271 were considered similar. The MAT analyses were calculated using the ‘analogue’ package of R (Simpson and Oksanen, 2014).
Results
Based on general sedimentological features (sand content), foraminiferal test abundance, and species diversity and dominance, the microfossil assemblages present in the two boreholes can be divided into different depth intervals (DIs). Table 4 and Figures 2 and 3 summarize the main borehole and microfaunal data. Interpretation of these DIs in terms of different habitats or subenvironments that evolved through time in this central area of the Ebro Delta is based on a palaeowater-depth transfer function developed by the authors. This transfer function compares the buried Holocene assemblages with dead foraminiferal assemblages in various settings of the modern delta based on data previously obtained by Scrutton (1969) and our own data (Table 2 and Figure 4).
Summary of core and microfaunal data from the Carlet and Sant Jaume boreholes (Ebro Delta). Figures represent relative abundance (%) unless otherwise indicated. The single value represents the average and those in parentheses give the range.

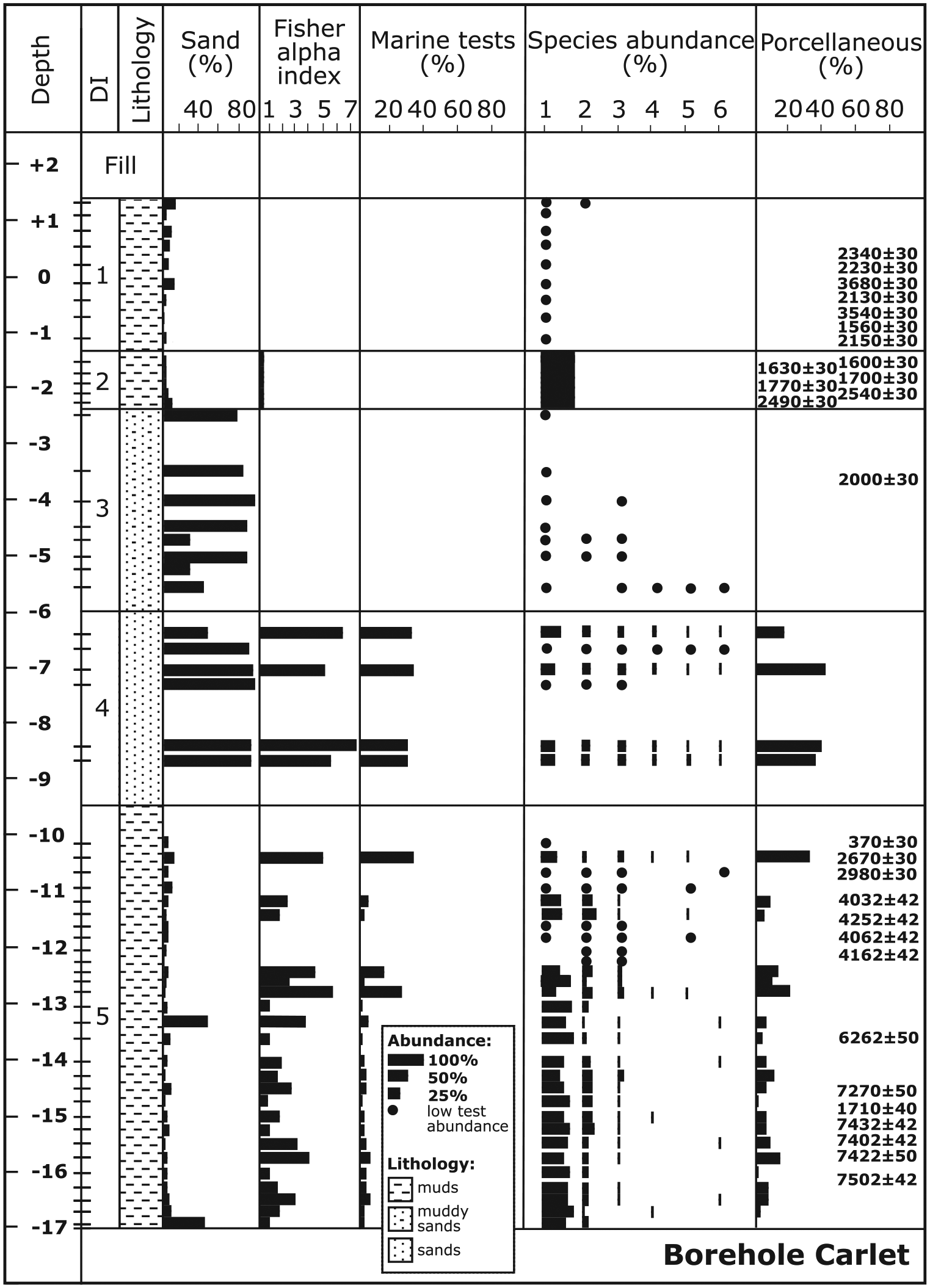
Sedimentary sequence, sand content, general micropalaeontological data, distribution and relative abundance of the main foraminiferal species (1: A. beccarii; 2: C. selseyense; 3: Q. seminula; 4: R. anomala; 5: T. marioni; 6: C. lobatulus), and porcellaneous wall–type content with depth (m) in the borehole Carlet (Ebro Delta). Foraminiferal depth intervals (DIs), radiocarbon dates (conventional years BP) and sample levels are also indicated. Black dots indicate presence of the species in assemblages with less than 100 foraminiferal tests.
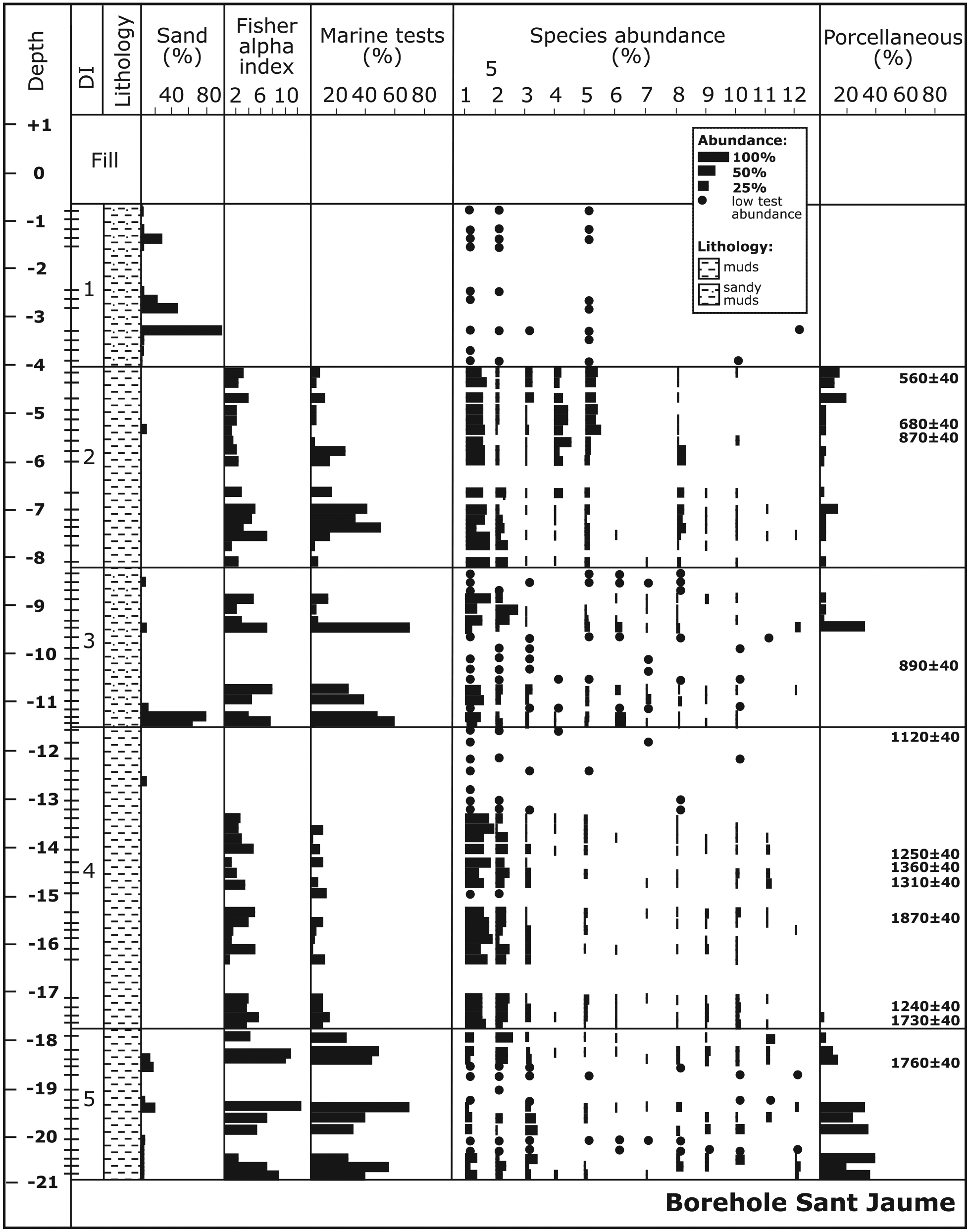
Sedimentary sequence, sand content, general micropalaeontological data, distribution and relative abundance of the main foraminiferal species (1: A. beccarii; 2: C. selseyense; 3: Q. seminula; 4: C. oceanensis; 5: H. germanica; 6: A. mamilla; 7: Q. oblonga; 8: B. variabilis; 9: B. gibba; 10: C. poeyanum; 11: V. bradyiana; 12: R. irregularis), and porcellaneous wall–type content with depth (m) in the borehole Sant Jaume (Ebro Delta). Foraminiferal depth intervals (DIs), radiocarbon dates (conventional years BP) and sample levels are also indicated. Black dots indicate presence of the species in assemblages with less than 100 foraminiferal tests.
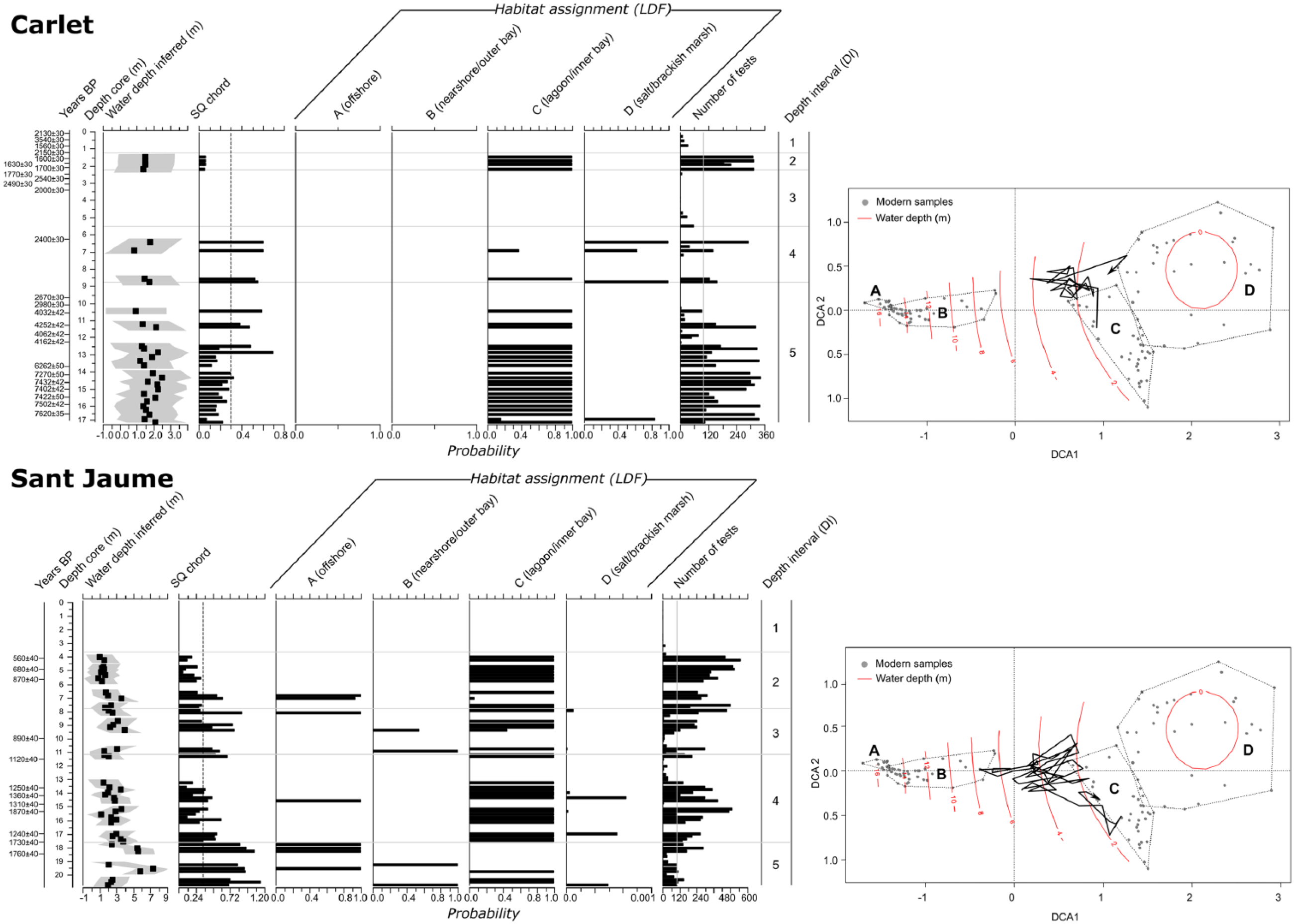
Reconstruction of palaeowater depth (associated errors as grey envelope) and different environmental settings through time in the Carlet and Sant Jaume boreholes (Ebro Delta) based on the transfer function developed by the authors. Habitat assignments using Linear Discriminant Functions (LDFs) are also shown. The foraminiferal depth intervals (DIs), number of tests and radiocarbon dates (conventional years BP) are also indicated. On the right, core trajectories projected onto DCA along with modern (dead) foraminiferal assemblages are shown. Dashed lines in DCA plots encompass modern habitat samples of each habitat type identified in the Ebro Delta: (A) offshore, (B) nearshore and outer bays, (C) coastal lagoons and inner bays, and (D) salt and brackish marshes. Water depth (m) intervals are marked for each DCA plot.
At the base of the Carlet borehole, DI5 is composed of at least 7.5 m of muds and is characterized by an assemblage highly dominated by Ammonia beccarii (average 74%) and Cribroelphidium selseyense (13%), with minor Quinqueloculina seminula (4%). The number of species is moderate (average 10), and the contents of marine and porcellaneous tests are low (average 7% and 8%, respectively). The comparison with modern assemblages suggests a lagoonal or shallow inner bay environment (0.5- to 1.5-m depth) for this interval, which developed during a long time span from 7600 to 2600 yr BP. Above DI5, DI4 is composed of 3.5 m of sands with variable abundances of foraminifera, and a high number of species (21), marine tests (34%) and porcellaneous forms (35%). The assemblages are composed mainly of A. beccarii (36%), Q. seminula (18%) and C. selseyense (13%), with secondary Rosalina anomala (6%), Triloculina marioni (6%) and Cibicides lobatulus (4%). All these features indicate a sandy environment with a mixture of deltaic (inshore) and marine (offshore) species, similar to a beach or back-barrier setting around the deltaic fringe. The transfer function did not find a close modern analogue for the assemblages in this interval, although the assemblages do indicate very shallow palaeowater depths (ca. 0–1 m). No age has been obtained in DI4 but radiocarbon ages from the underlying and overlying intervals suggest it developed after 2600 and before 2000 yr BP. Consequently, the coastline was located close to this location around this time. The overlying 3.7 m of muddy sands (DI3) contain very few foraminiferal tests and may indicate the shift to a higher elevation and less flooded habitat (an alluvial environment closer to the river levee) in the area of Carlet approximately 2000 yr BP. Subsequent conditions suggest the presence of a fresh to brackish marsh environment (less than 0.5-m depth) in which 1 m of muddy sediments (DI2) was deposited at approximately 1700 yr BP. These sediments are characterized exclusively by an abundant A. beccarii assemblage, and the presence of terrestrial gastropods and oogonia of characeae algae is indicative of very low salinity conditions in this area. Finally, DI1, with 2.7 m of muds, also features terrestrial gastropods and characeae oogonial, but the foraminiferal content is very low (only a few tests of A. beccarii), suggesting a lacustrine environment. Radiocarbon dates show a great variety of ages and inverted dates. This reinforces the idea that, during the last 2.0 ka (DI2–DI1), this area was a fresh to brackish aquatic environment located close to the river thus featuring a relatively high elevation and occasional marine flooding conditions.
In contrast, the Sant Jaume borehole shows much more recent and deeper materials than the Carlet borehole. The lower part of the sequence initiates with DI5 and more than 3 m of muds containing a variable abundance of foraminifera characterized by a high number of species (22), marine tests (43%) and porcellaneous forms (24%). The assemblage is composed mainly of C. selseyense (18%), A. beccarii (13%) and Q. seminula (11%), together with Cribroelphidium poeyanum (6%), Brizalina variabilis (5%), Bulimina gibba (4%), Valvulineria bradyana (3%) and Rosalina irregularis (3%). A radiocarbon age of 1760 yr BP for the upper part of this interval indicates that in this period the sediment accumulated in the nearshore environment (approximately 7-m depth). The following unit (DI4) is represented by 6.3 m of muddy sediment deposited in an inshore more proximal setting (lagoon or shallow inner bay environment; 0.5- to 1.5-m depth) dominated by A. beccarii (65%) and C. selseyense (19%), with Q. seminula (3%) and C. poeyanum (3%). The number of species is moderate (13) and the marine tests (8%) and porcellaneous content (5%) are low. This interval developed between 1700 and 1100 yr BP. Above this unit, a 3.4-m unit of sandy muds with a variable abundance of foraminiferal tests exhibits an increase in the number of species (18) and marine (34%) and porcellaneous (16%) tests (DI3). An assemblage composed of A. beccarii (38%), C. selseyense (18%) and Asterigerinata mamilla (11%), with minor Quinqueloculina oblonga (3%), Q. seminula (3%) and Haynesina germanica (3%), suggests a sandier nearshore habitat (approximately 7-m depth) that developed approximately 1.0 ka ago. The overlying interval DI2 (4 m of muds) shows a decrease in the open marine influence (15% marine tests and 5% porcellaneous forms), a moderate number of species (14) and the dominance of more deltaic (inshore) forms, such as A. beccarii (52%), H. germanica (11%), Cribroelphidium oceanensis (11%) and C. selseyense (8%), with B. variabilis (6%). These features are indicative of a lagoonal or shallow inner bay environment (0.5- to 1.5-m depth) that developed in this area between 900 and 600 yr BP. The final 3.3 m of sandy muds (DI1) are younger than 0.5 ka, contain few foraminiferal tests (A. beccarii, H. germanica and C. selseyense) and may represent an emerged fresh-water environment with occasional marine flooding located in a delta plain.
Discussion
Palaeoenvironmental evolution of the Ebro Delta
A comparison of the materials, microfossil assemblages, radiocarbon ages and palaeoenvironmental evolution of the two boreholes clearly indicates that the more landward Carlet sedimentary sequence exhibits sandier, older and more continental conditions, whereas the Sant Jaume geological record exhibits muddier, younger and more marine environmental conditions characteristic of a more seaward setting within the Holocene architecture of the Ebro Delta. These distinctive stratigraphic and foraminiferal sequences, with an older and more proximal progradational record on one side and a younger and more distal progradational record on the other side, are also found in other Mediterranean deltas, such as the Rhône (France) and the Po (Italy) (Amorosi et al., 2005, 2013; Boyer et al., 2005; Dinelli et al., 2012; Rossi and Vaiani, 2008). However, the interpretation of the environments in some cases is different than in the present study because a wider range of foraminiferal assemblages, including open marine and delta plain environments, is considered in this study (see discussion on this topic in the following section).
The palaeoenvironmental interpretation of the sedimentary sequences in the Carlet and Sant Jaume boreholes is based on diagnostic foraminiferal assemblages and is shown in Figure 5. At Carlet, apart from the lowermost part of DI5 deposited before 7.0 ka during a TST under rising sea-level conditions, the muddy DI5 at the base and the sandy DI4 are interpreted to be mainly a part of a sequence composed of inner bay–lagoonal–beach deposits in this central area under sea-level highstand conditions between 7000 and ~2000 yr BP. Then, further sediment accumulation led to the formation of a series of non-marine deposits represented during the last 2.0 ka, initially represented by continental sands in DI3. These deposits were followed by a very low salinity marsh environment with occasional marine flooding at approximately 1700 yr BP (DI2), which was finally replaced by a fresh-water muddy environment (higher elevation and closer to the river levee) containing a mixture of materials with variable radiocarbon ages (DI1).
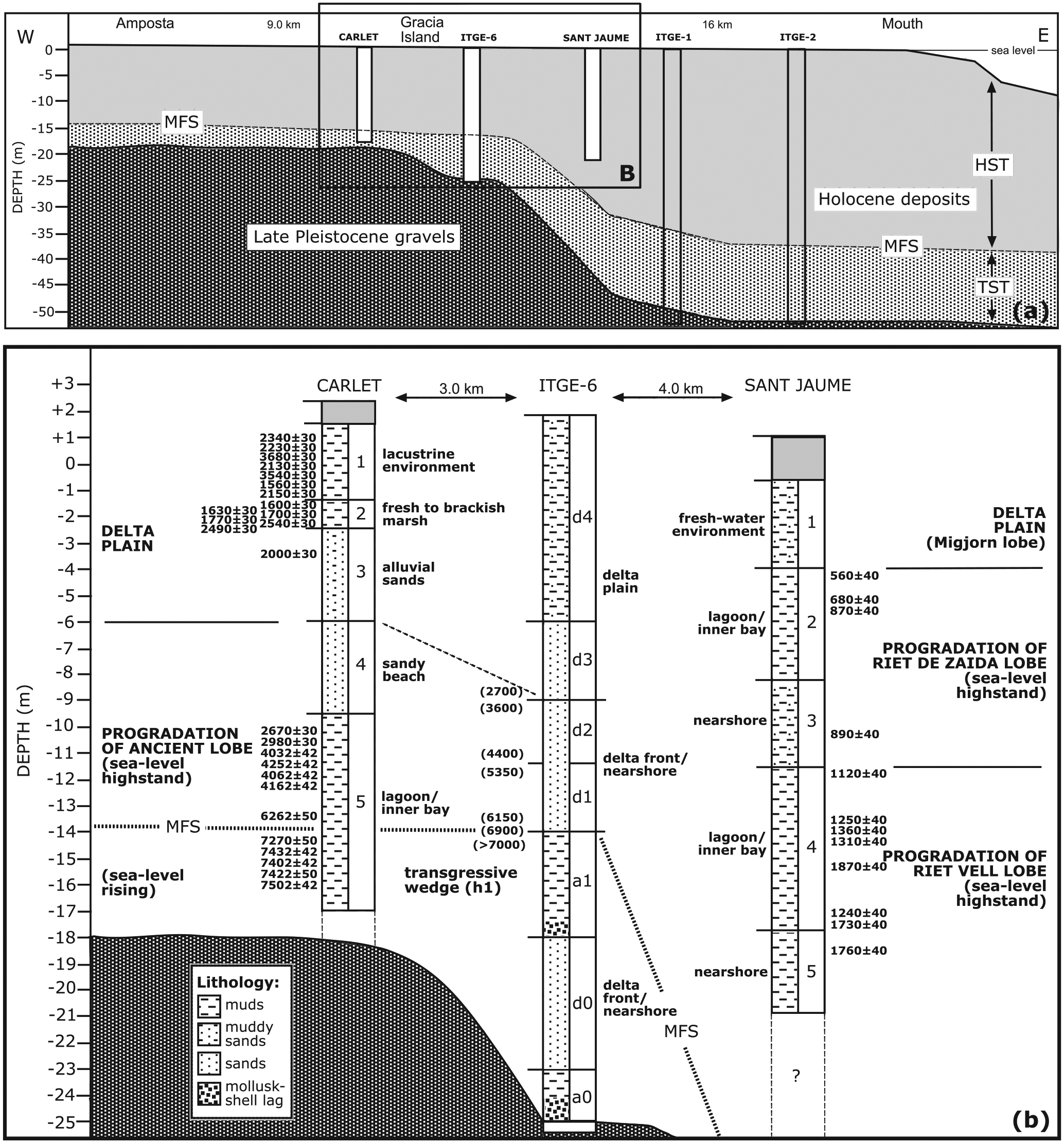
(a) General location of the analysed boreholes in the framework of the Ebro Delta architecture proposed by Somoza et al. (1998) for the late Quaternary; and (b) palaeoenvironmental interpretation of the Carlet and Sant Jaume boreholes based on foraminiferal assemblages. Foraminiferal depth intervals (DIs), lithology and radiocarbon dates (conventional years BP) are also indicated. Interpretation of the ITGE-6 borehole is from Somoza et al. (1998) with indication of the aggradational marine (a units) and progradational deltaic (d units) deposits. Dates in parentheses were assigned by Somoza and Rodríguez-Santalla (2014) but were not obtained from materials of this borehole.
A comparison of the Carlet results with the previous ITGE-6 borehole sequence of Somoza et al. (1998) indicates the partial absence in our record of the most basal units of the Holocene sequence that were deposited under rapidly rising sea-level conditions (TST) (Figure 5). There are similarities but also important quantitative and qualitative differences between the two records in terms of palaeoenvironmental reconstructions and the timing of events. The lower part of our muddy DI5 interval (deposited between 7.6 and 7.0 ka) could be correlated to the muddy aggradational unit (a1) in ITGE-6 (deposited as the transgressive wedge h1 before 7.0 ka) and represents the final record of the TST. After the MFS, suggested by Somoza and Rodríguez-Santalla (2014) to have occurred at approximately 6900 yr BP, the remainder of the muddy DI5 and sandy DI4 intervals (deposited between 6.9 and 2.5 ka) could be correlated to the sandy units d1 and d2 (delta front/nearshore) in ITGE-6 (deposited between 6.1 and 3.6 ka under sea-level highstand conditions). Then, the series of delta plain deposits (the sandy and muddy intervals DI3–DI1) during the last 2.5 ka in Carlet very likely correspond to the sandy and silty d3 and d4 delta plain units (considered to be younger than 2.7 ka) in the ITGE-6 sequence. However, the information provided by the foraminiferal assemblages in our study suggests that the maximum Holocene marine transgression did not reach the inner part of the Ebro Delta in the form of open-sea environments but rather via lagoon and/or shallow bay development. This is an important difference with the interpretation provided by Somoza et al. (1998) in relation to the existence of a transgressive wedge (h1) with marine clayey sediments corresponding to the MFS. Our results clearly show the existence of restricted and brackish environments, which is more compatible with the presence of fresh-water peat deposits in the innermost part of the delta during that time (Arasa, 1994; Solé et al., 1961), because fresh-water peat cannot form during high-salinity conditions. This reasoning also applies to the other highstand sea-level events (h2, h3 and h4) postulated by Somoza et al. (1998) in the innermost part of the delta (upstream of the city of Amposta in the alluvial valley) and next to the Ebro River, where each period of fresh-water peat accumulation would coincide with the presence of the salt-water conditions. We do not find any evidence for this interpretation, and an alternative explanation is a succession of fresh-water peat deposits and alluvial or lagoonal deposits as a function of the changing distance between the borehole location and the Ebro River through time (due to the migration of the river course), as well as changes in the frequency and magnitude of river floods.
This alternating pattern of peat and alluvial or lagoonal clay deposits is also found in the innermost part of other large Mediterranean deltas. In the Po Delta plain, Amorosi et al. (2005) describe a similar sequence in the innermost boreholes (204-S17, 204-S5, 204-S6) with the presence of peat layers in the middle of fresh-water (swamp) clays or brackish-water (lagoonal) clays, whereas the marine (bay) clays are only found in the outermost boreholes (204-S7, 205-S5) and never coincide in time with peat layers. This study also included the analysis of microfossils (foraminifera and ostracoda) and similar foraminiferal assemblages (Ba–Bd) with few species that were interpreted as low-energy brackish-water back-barrier environments. In the Rhône Delta, Boyer et al. (2005) also describe the presence of clay (including layers of alluvial sand) with brackish to fresh-water fauna in the innermost boreholes (109, 108, 126), whereas clays with marine fauna are only found in the outermost boreholes (125 to 106).
The Sant Jaume borehole is located in a more seaward position than Carlet and contains a longer, deeper and much younger sequence that accumulated during the last 2.0 ka. Due to the available historical data discussed above, it is possible to interpret its sedimentary record as deposition during the formation and development of three different deltaic lobes under highstand sea-level conditions. The muddy intervals DI5 and DI4 represent a partial record of the formation and development of the Riet Vell lobe that prograded towards the southeast and deposited progressively shallower (from nearshore to more proximal inner bay) materials in this area between 2000 and 1100 yr BP. Then, deposition of a new muddy progradational shallowing-upward succession between 1100 and 500 yr BP, represented by DI3 and DI2, characterizes the development of a new deltaic lobe (Riet de Zaida) that developed towards the northeast and deposited nearshore–inner bay–lagoonal sediments in this area. A more recent switch in the river course because of human intervention created the newest delta lobe (Migjorn) represented by lacustrine (fresh-water) deposition (DI1) in this area during the last 0.5 ka.
Two external boreholes (ITGE-1 and ITGE-2; Figure 1) were studied by Somoza et al. (1998), although their descriptions are less detailed than the central ITGE-6 borehole. Both sedimentary successions reached the Pleistocene gravels, were approximately 50 m thick, and included TST and HST deposits. The HST was composed of four progradational units (dl, d2, d3 and d4) and three aggradational units (a2, a3 and a4). The boreholes were located at the apices of two main delta lobes identified in historical records: ITGE-l over the Riet Vell lobe, and ITGE-2 over the most recent Migjorn lobe. Their final progradational unit (d4) corresponds to the formation of their respective deltaic lobes, but no information on the origin, sedimentary characteristics and chronology of the other earlier progradational units is presented. Our Sant Jaume borehole is located in the central delta plain at the confluence area of the last three delta lobes: Riet Vell, Riet de Zaida and Migjorn (Figure 1). Distinct marine and delta plain habitat successions have characterized the formation and development of those three lobes, and these habitat successions were associated with particular foraminiferal palaeoassemblages. First, progradation of the Riet Vell lobe (2.0–1.1 ka) resulted in progressively shallower muddy environments from nearshore (DI5) to more proximal inner bay (DI4). The Riet de Zaida lobe (1.1–0.5 ka) exhibits a similar environmental succession with nearshore (DI3) and inner bay–lagoonal sediments (DI2). Finally, the most recent Migjorn lobe (0.5 ka) is represented by a fresh-water environment (DI1) in a delta plain setting.
As we have shown, important differences exist in the scale of resolution between the Carlet and Sant Jaume boreholes. Carlet provides information at a temporal resolution of thousands of years, whereas San Jaume provides data at the historical scale. A similar thickness of sediments represents 7.6 ka in Carlet and circa 2 ka in San Jaume. The Carlet borehole correlates with the progradational d1, d2, d3 and d4 of Somoza et al. (1998), which correspond to four different fifth-order cycles. In contrast, the entire record of the San Jaume borehole can be assigned to the prograding portion of one complete cycle of higher rank, corresponding to the last d4 unit of the historical lobes Riet Vell, Riet de Zaida and partially the d5 unit of Migjorn. In addition to the processes of compaction, preservation and possible time-averaging experienced by the older Carlet sedimentary sequence, the shorter time interval and deeper environments represented by the longer Sant Jaume sedimentary sequence can be understood as a consequence of the greater accommodation space available in the delta seaward from Gracia Island. As shown in Figure 5a, the geometry of the Holocene materials has a thickness ranging from 20 m on the landward side (Amposta) to 52 m at the delta front. The erosional unconformity between the upper Pleistocene gravels and the Holocene deposits exhibits a more pronounced slope in the external delta just beyond Gracia Island and was formed initially by marine erosion during rapid sea-level rise during the first stages of the Holocene transgression (Maestro et al., 2002). These authors also indicated the presence of extensional tectonics that affected the Quaternary deltaic deposits via the formation of large-scale faulting that generated differential subsidence beneath the Ebro Delta and increased the slope angle. The principal process involved in fault development is considered to be differential compaction resulting from the prograding deltaic lobes that overlie aggradational clay deposits. Growth faults increase subsidence, which in turn generates accommodation space for subsequent prograding deltaic sediment. This arc-shaped topographic depression acted as a trap for distributary channels meandering across the delta plain, such as the abandoned Riet Vell and Riet de Zaida (Maestro et al., 2002).
Implications for the Holocene evolution of Mediterranean deltas
In terms of their palaeoenvironmental significance, when modern foraminiferal assemblages from the delta plain habitats are included in the analysis, the interpretation is more robust compared with other studies that only consider open marine (offshore) assemblages. The existing literature shows that foraminiferal assemblages living in coastal marginal environments, such as coastal lagoons, bays or salt marshes, differ greatly from those in adjacent offshore habitats (Murray, 2006). This also applies to some worldwide and Mediterranean deltas, such as the Mississippi, US (Lankford, 1959), Mahakam, Indonesia (Lambert, 2003), Rhône, France (Fanget et al., 2012; Vangerow, 1974), Nile, Egypt (Arbouille and Stanley, 1991) and Ebro, Spain (Scrutton, 1969). Previous studies often interpreted the occurrence of shallow brackish-marine species, such as Ammonia spp. (A. tepida, A. beccarii, A. parkinsoniana), Cribroelphidium spp. and/or H. germanica, as indicators of past offshore habitats within Holocene deltaic sequences (Amorosi et al., 2008, 2013; Milli et al., 2013; Rossi and Vaiani, 2008). However, it is well known that these species are mainly indicative of particular marginal coastal habitats, unless other ecological requirements based on the study of their living populations are actually identified (Guillem, 2007; Usera et al., 2002). Otherwise, the lack of close modern analogues may lead to weaker interpretations of the foraminiferal fossil record.
The present study has demonstrated the similarity between most modern and fossil samples in the Ebro Delta by applying the MAT and LDFs. Hence, a finer-grained habitat reconstruction can be achieved. The interpretation of results would have been different if foraminifera from deltaic (inshore) environments had not been included, especially for the Carlet sequence, which features the presence of shallower habitats typical of delta plain environments, in comparison with the Sant Jaume sequence, which features deeper environments typical of the prodelta and delta front. To our knowledge, this is the first attempt to include modern samples from both delta plain (coastal lagoons, marshes and inner bays) and open marine (prodelta and delta front) habitats of a Mediterranean delta to provide adequate analogues for all borehole samples. Thus, the Ebro Delta data set could be used in other Mediterranean deltas with the aim to provide new interpretations of their depositional environments because all of these deltas share a similar Holocene evolution (Stanley and Warne, 1994).
The application of a water depth transfer function to fossil foraminiferal assemblages has complemented the palaeoenvironmental reconstruction of the Ebro Delta. The only comparable work is by Rossi and Horton (2009), who applied the Northern Adriatic Transfer Function (NATF) to reconstruct the evolution of the Holocene palaeobathymetry of the Po Delta. These authors concluded the existence of a shallowing-upward trend following the progradational succession of this delta during the last 5.5 ka. In turn, the palaeowater depths were considered reliable according to MAT results. For the Ebro Delta, very similar results were found in the Sant Jaume sequence, where two deeper-to-shallower successions were detected (DI5–DI4 and DI3–DI2) with inferred water depths from 7 to 1 m (Figure 5).These findings suggest the progradation of delta lobes in this distal part of the delta during the last centuries, although some samples did not contain close modern analogues. In contrast, the palaeowater depths of the Carlet sequence revealed very shallow conditions throughout all of the recorded depositional environments, with water depths ranging from 1 to 2 m. The validity of these reconstructions is supported by the MAT and LDF results, although palaeowater depths must be taken with caution because of the complex relationships between water depth and foraminiferal distributions, particularly in highly dynamic systems such as deltas. Further research on modern foraminiferal assemblages in inshore habitats of other Mediterranean deltas could allow a more reliable interpretation of their Holocene evolution.
Conclusion
The scarcity of previous geological studies and the few available radiocarbon dates from Holocene sedimentary archives, together with misinterpretations of historical documents, have permitted the idea that the present Ebro Delta plain formed mostly during the 14th and 15th centuries and that it was an estuary during Roman times. Our results support an early Holocene start for the Ebro Delta, proving that a deltaic depositional system was present throughout the Holocene and that the Ebro Valley never became an estuary. This brings the Ebro evolution story in line with other major deltas along the Mediterranean Sea and worldwide, which initiated 8.0–6.0 ka ago when the rate of fluvial sediment input overtook the decreasing rate of sea-level rise.
The palaeonvironmental evolution of the central plain of the Ebro Delta during the Holocene was reconstructed using micropalaeontological analysis of two continuous boreholes (Carlet and Sant Jaume). Diagnostic foraminiferal assemblages and the application of a palaeowater-depth transfer function allowed the definition of various lithofacies and associated environments of deposition. The geometry of the Ebro Delta was created by the advance of successive deltaic lobes, which prograded radially across the inner shelf up to 25 km seaward during the Holocene. Avulsion and channel abandonment processes are considered to be the main delta constructional processes, with the resulting deposits subsequently being modified by rapid subsidence.
The similarity between most modern and fossil samples in the Ebro Delta has been demonstrated by applying the MAT and LDFs, allowing a much better habitat reconstruction to be achieved. The importance of extensive characterization of modern foraminiferal assemblages from both deltaic inshore and offshore environments in order to provide adequate analogues for the interpretation of borehole samples is demonstrated particularly by the more landward Carlet sequence, which only contains shallow and inshore deltaic habitats. The palaeowater depths revealed very shallow conditions for all depositional environments, ranging from 1 to 2 m during the last 7.6 ka under salty, brackish and fresh-water conditions.
Footnotes
Appendix 1
Acknowledgements
Microfossil samples from the Carlet borehole were prepared and analysed initially by Alfonso Palazuelos as part of his MSc dissertation (academic year 2012/2013) at the University of the Basque Country UPV/EHU. Dr Francisco Fatela (University of Lisbon, Portugal) and an anonymous reviewer greatly improved the original manuscript with their critical comments and suggestions. It represents a contribution to the INQUA Commission of Coastal and Marine Processes and Contribution #24 of the Geo-Q Zentroa Research Unit (Joaquín Gómez de Llarena Laboratory).
Funding
Drilling and coring was funded by the US National Science Foundation grant EAR-0952146. Work on the cores presented in this study was partially financed by the Formation and Research Unit in Quaternary: Environmental Changes and Human Fingerprint (UPV/EHU, UFI11/09) and HAREA-Coastal Geology Research Group (Basque Government, IT767-13) projects. It was supported by an IRTA-URV-Santander fellowship to Xavier Benito through ‘BRDI Trainee Research Personnel Programme funded by University of Rovira and Virgili R + D + I projects’ and the European Community’s 7th Framework Programme through the grant to Collaborative Project RISES-AM-, Contract FP7-ENV-2013-two-stage-603396.