
Abstract
Geomorphologic, stratigraphic, faunistic, palynological and 14C analyses were carried out in the area of the mouth of the Garigliano River characterized by two strand plains that are referred to the Eutyrrhenian and the Holocene, rimming two depressed zones separated by the Garigliano River channel. This study depicts the palaeoenvironmental evolution over the last 8200 years and the landscape context at the time of Minturnae Roman colony. Between 8200 and 7500 yr BP, a wet zone occurred in the northern zone, whereas in the southern part, a lagoon developed. During the final transgression stage and the beginning of the sea level still stand (7500–5500 yr BP), a freshwater marsh formed in the northern zone, and the width of the southern lagoon decreased. Between 5500 and 3000 yr BP, the coastal barrier changed into a delta cusp, a freshwater marsh also appeared in the southern part and the river wandered between the twin marshes. Because of local uplift, previously unknown in this area, part of the floor of the southern marsh emerged, and after 4000 yr BP, both marshes became coastal ponds with prevailing clastic sedimentation. A progressive increment in anthropic forcing on the land took place after 3000 yr BP. The Marica sanctuary was built (7th century BC), and the Roman colony of Minturnae was developed beginning the 3rd century BC. The shallow depth of the ponds prevented their use as harbours, and saltwork plants can be ruled out based on the faunal and palynological data. The ongoing infilling of both ponds was never completed, and their reclamation is still in progress.
Introduction
Many accounts have reported that along the microtidal Mediterranean coasts in the late Holocene, several river mouths developed deltas of variable width according to the dynamic equilibrium between the rate of sea level rise and the solid river discharge (Arnaud-Fassetta and Provansal, 1999; Bellotti et al., 1994, 1995, 2003; Correggiari et al., 2005; Coutellier and Stanley, 1987; Gensous et al., 1993; Milli et al., 2013; Somoza et al., 1998; Vella et al., 2005).
The determinations of the evolutionary patterns of Mediterranean deltaic areas have frequently revealed the palaeoenvironments at the times of the development of ancient settlements and are a valuable aid to archaeological research (Arnaud-Fassetta et al., 2003; Bellotti et al., 2011; Bicket et al., 2009; Di Bella et al., 2011; Di Rita et al., 2010; Giraudi, 2011; Giraudi et al., 2009; Goiran et al., 2009, 2014; Marriner and Morhange, 2006, 2007; Sadori et al., 2015; Stanley, 2005; Stanley and Bernasconi, 2006, 2009). This paper addresses the palaeoenvironmental reconstruction of the delta plain of the Garigliano River in the area where the river divides two depressions that are almost parallel to the coast and are constrained between two strand plains. On the inner strand plain, which is referred to the Eutyrrhenian (Pennetta et al., 2011), the Roman colony of Minturnae was founded, whereas the outer one is part of the present Garigliano River delta. The sedimentological and environmental features of the study area have been previously investigated by Remmelzwaal (1978), Bellotti et al. (2012), Ferrari et al. (2013b) and Bellini and Trigona (2014). In this new research, multidisciplinary analyses and pollen zonation highlight the delta development phases over the last 8000 years, as well as the landscape context at the time of Minturnae. This study allows the insertion of a new element in the changing geomorphology of Mediterranean deltas where important historical settlements developed (Anthony et al., 2014). For the first time, the architecture of the local sedimentary succession is shown. The architecture reveals a conflict between the actual stratigraphic positions of sediments, of known age, with those suggested by the sea level rise curves. This conflict suggests a vertical neotectonic hitherto unknown in this area.
Geological setting
The Garigliano delta plain (Figure 1) is located on the Tyrrhenian coast between Latium and Campania. Along with the Gaeta gulf and the Campanian coastal plain, it is part of a wide Quaternary extensional basin belt that stretches from the Tyrrhenian margin to the Central-Southern Apennine chain (Casciello et al., 2006; Ferranti et al., 1996; Malinverno and Ryan, 1986).
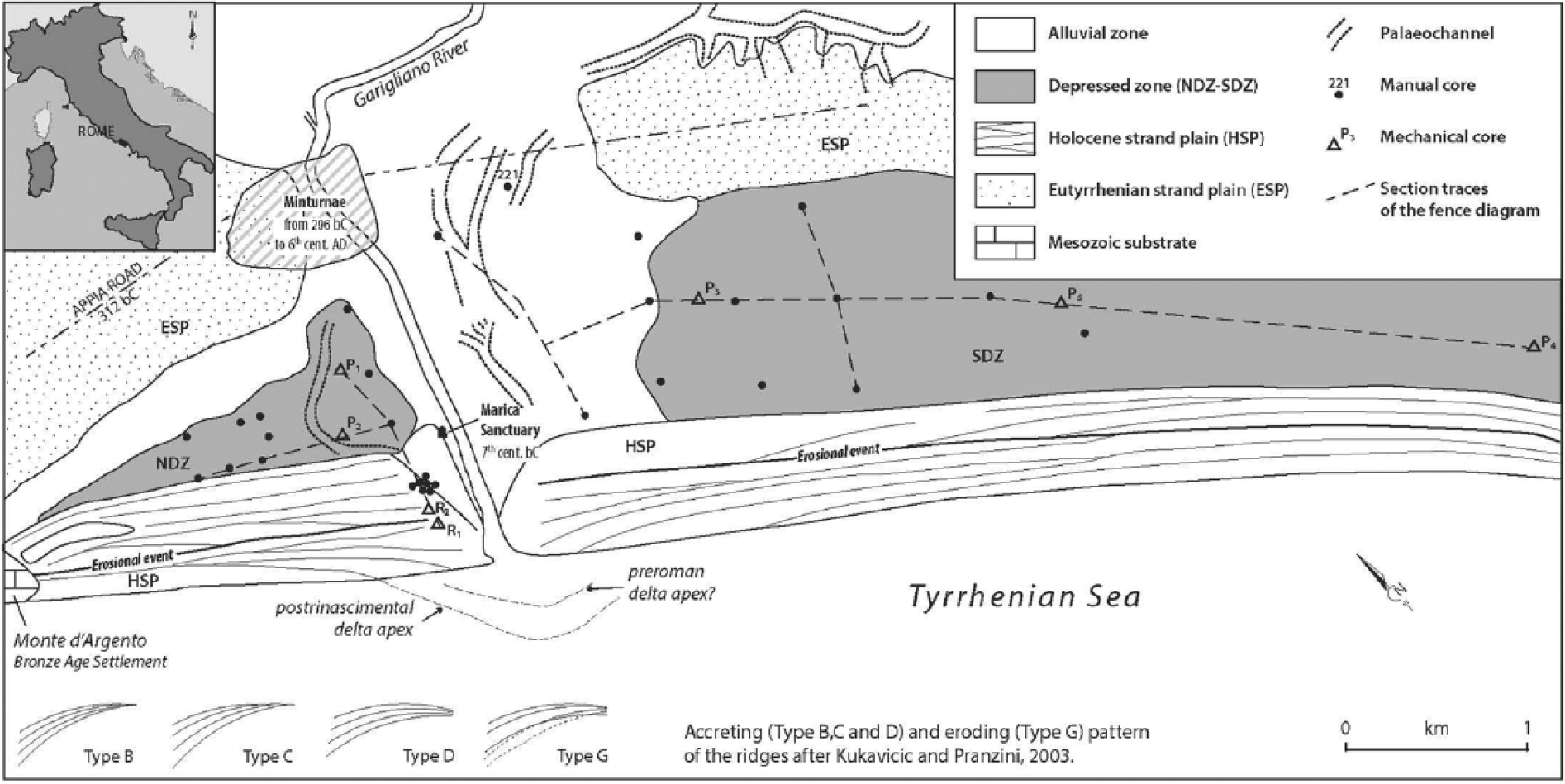
Morphological features of the Garigliano delta plain. In the HSP zone, the crests of the main beach ridges are drawn, which allowed the main progradational and erosional phases of the Holocene strand plain to be defined.
The coastal plain lies on a graben produced by a complex anti-Apennine recurrent faulting (Abate et al., 1998) from the late Miocene to approximately 125,000 yr BP. Over that time span, the graben was filled with marine Pliocene and marine-transitional Pleistocene sediments (Ippolito et al., 1973). During the latter infilling period, the area also received tephra from the Roccamonfina volcano (Remmelzwaal, 1978), whose activity only ended at approximately 50,000 yr BP (Davoli et al., 1999). The late Pleistocene–Holocene evolution depended on distinct, interacting processes that included the late glacio-eustatic cycle (125,000–6000 yr BP), solid river discharges, tephra from the Phlegrean Fields and Vesuvius and the final, smooth tectonic phases (Brancaccio et al., 1991; Di Vito et al., 1999).
During the Tyrrhenian high stand (125,000 yr BP), the coastal zone was characterized by a strand plain that bordered a wide bay. Then, in the succeeding glacio-eustatic sea level fall, the strand plain was locally eroded by the Garigliano River (Pennetta et al., 2011; Remmelzwaal, 1978). In particular, during its westward flow, the river incised a valley that was subsequently buried in the Tyrrhenian shelf (Chiocci and La Monica, 1996).
After the last glacial maximum from 6000 to 7000 yr BP, the sea level rose and approached the present level, and paralic systems with wave-dominated estuaries or lagoons developed in analogy with several delta areas in the Tyrrhenian margin (Amorosi et al., 2012; Bellotti, 2000; Milli et al., 2013; Sacchi et al., 2014).
Then, contemporary to the subsequent sea level still stand, the shoreline near the Garigliano River mouth progradated and originated, westward of the Eutyrrhenian beach ridge system, the Holocene strand plain. This latter strand plain confines seaward the relics of the Holocene coastal ponds, marshes and lagoons.
Archaeological setting
The Garigliano delta plain has been inhabited since at least the Late Bronze Age when a village was set atop the rocky promontory of Monte d’Argento, and several others were founded on the Eutyrrhenian strand plain bordering the Holocene marshes landward (Ferrari et al., 2012). In this period, the wetlands probably were exploited for fishing or for harvesting aquatic plants (Angle and Belardelli, 2007).
In the Iron Age, the coast was probably uninhabited, apart from the area near the river mouth. Near the river’s right bank, on a Holocene beach ridge between the shoreline and the marshes, there was a place of worship dedicated to the goddess Marica (a chthonian divinity with power and control over water and marshes), which was in use since at least the 7th century BC (Andreani, 2003). It was an emporic sanctuary that mediated the inland and Mediterranean trade (Bellini, 2002). The most important settlements of the Aurunci (the populations living in the Garigliano delta plain in this period) were atop the carbonate terraces in the hinterland (Ferrari et al., 2014). Because of the alliance with the Samnites during the Second Samnite War, the Auruncan towns and villages (oppida) were destroyed by the Romans in 314 BC.
The Roman reorganization of the region took place in three steps: the construction of a road network centred on the Via Appia (312 BC), the deduction of colonies (Sessa Aurunca, 313 BC; Minturnae and Sinuessa, 296 BC) and the centuriation of the territory. Minturnae, which was located atop the Eutyrrhenian strand plain on the right bank of the Garigliano, very quickly became an important town (Guidobaldi and Pesando, 1989). Many small settlements and farms were set in the countryside, and their goods were traded throughout the Mediterranean because of the accessibility of its harbour.
The riverbanks and the landing place close to the sanctuary of Marica were strengthened during the 1st century AD, but the marshes were not drained. Plutarch (Life of Marius, 37–39), Appian (De Bello Civili, 1, 7, 61–62), Cicero (Pro Sestio, 22, 50; Ad Quirites 8, 20; In Pisonem, 19, 43) and other Latin authors gave a detailed description of the landscape near Minturnae in 88 BC. The ancient written sources record wetlands with shallow water basins and muddy soils that were rich in vegetation (reeds and aquatic plants), with Salix and Quercus dominating the arboreal plants (Ferrari et al., 2013a). Epigraphs (mentioning socii salinatores, socii picarii and an architectus navalis) and written sources (especially those after Plutarch) testify the existence of an important harbour on the right bank of the river, just in front of the ancient town (Ruegg, 1995). Both Plutarch’s account and the submerged structures nearby the Marica sanctuary suggest that a further harbour or a landing place, distinct from that previously mentioned, was located near the river mouth (Bellini, 2007; Bellini and Trigona, 2014; Gregori and Nonnis, 2013).
From the 3rd century AD, the settlement pattern, based on farms and villas, declined, and around the second half of the 6th century AD, Minturnae itself was probably abandoned, following the migration of the population towards the nearby hills where the present Minturno is situated (Arthur, 1989).
In any event, between AD 881 and 915, the Garigliano River mouth was still used as a landing place until a colony of Saracens settled close to the ancient town of Minturnae. Then, following the military expulsion of the Saracens, the coast remained almost unpopulated, and the area behind the Holocene beach ridges was occupied by small lakes and marshes, as reflected in toponyms such as Pantano di Traetto and Pantano di Sessa (Pantano = Italian for marsh or pond).
Materials and methods
For the geomorphologic analysis, sets of aerial photographs taken by the Royal and Italian Air Forces were used, and ancient cartography was used to identify the palaeodrainage network, the relationships among the beach ridge crests of the Holocene strand plain and the features of the coastal basins between the Holocene and Eutyrrhenian strand plains.
Elements for the stratigraphic analysis were acquired from 31 manual auger cores and five continuous mechanical drillings. In addition, the logs from two mechanical cores drilled on behalf of the Latium Region, just north of the Garigliano River mouth, were utilized. The manual core logs yielded the lithology of the uppermost 350 cm of the sedimentary suite. The mechanical cores provided undisturbed samples that were reliable for proxies and biological analyses. The sediment textures were determined using mechanical sieving and laser diffractometry for >62 µm and <62 µm fractions, respectively.
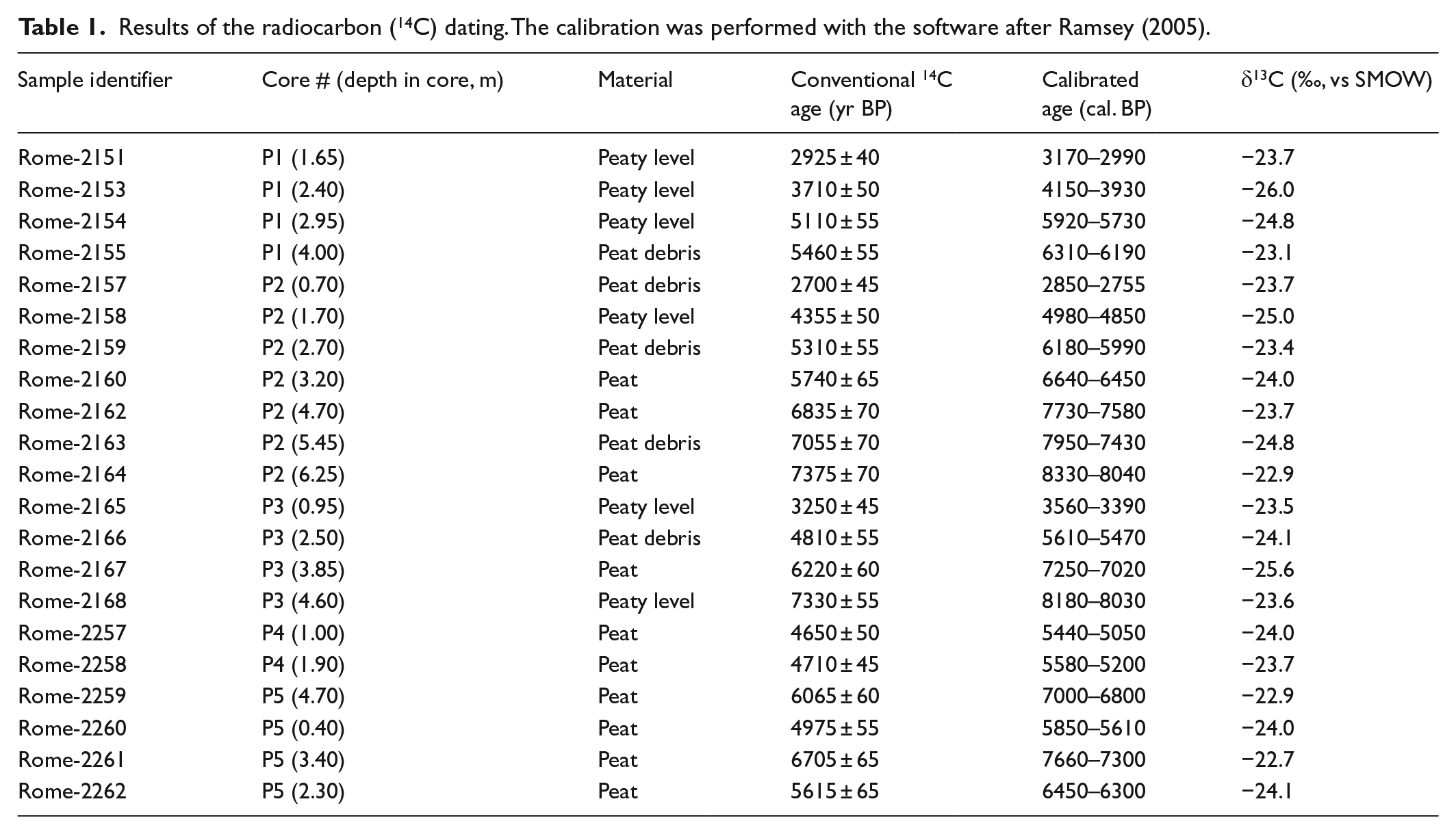
The radiocarbon dating was performed on plant remains ranging from herb filaments to variably altered wood debris that had been previously decontaminated to remove carbonates, as well as acid- and alkali-soluble organics. The residues were then burned, and the resulting CO2 was used for the synthesis of benzene, the medium for the 14C analysis using Liquid Scintillation Counting (LSC). The conventional ages, corrected for the C isotope fractionation to δ13C = −25‰ and calculated according to Stuiver and Polach (1977), are reported in yr BP (present time set at 1950). The conventional ages were calibrated according to Ramsey (2005) and are given as cal. yr BP time-spans. The uncertainties of both the conventional and calibrated ages are at the level of ±1σ.
Malacological analyses were performed on 76 sediment samples (3 from core P1, 7 from P2, 3 from P3, 19 from P4 and 44 from P5). The shells were counted and identified according to the WoRMS database for marine and brackish taxa and Welter-Schultes (2012) for non-marine taxa. The ecology inferences were according to Pérès and Picard (1964) for the marine and brackish species and after Welter-Schultes (2012) for the non-marine species. The foraminiferal analyses were based on 74 sediment samples (2 from core P1, 9 from P2, 1 from P3, 20 from P4 and 42 from P5). The sediments were wet-sieved through a 63-µm sieve, dried and split into aliquots carrying some 300 well-preserved specimens that were classified according to Loeblich and Tappan (1987), Cimerman and Langer (1991) and Sgarrella and Moncharmont-Zei (1993). The percentage distribution of the specimens in each species yielded the Faunas Association Zone (FAZ).
Pollen analyses were performed on cores P1, P2 and P3, and the sampling frequency ranged from 5 to 10 cm. Based on the stratigraphy versus 14C data readings 45 sediment samples were correlated and analysed: 14 from P1 (from 180 to 10 cm deep), 28 from P2 (625–75 cm) and 3 from P3 (450–410 cm). The samples (1–2 g) were treated with tetra-Na-pyrophosphate, HCl 10%, acetolysis, floatation with Na-metatungstate hydrate, HF 40% and ethanol (Florenzano et al., 2012; Van der Kaars et al., 2001). Lycopodium spores were added to calculate pollen concentration. Pollen and Non-Pollen Palynomorphs (NPPs) were identified under a 1000× magnification light microscope based on keys and atlas photographs (e.g. Van Geel et al., 2003) and the reference pollen collection of the Laboratory of Modena. An average of 470 pollen grains in the P1 and P3 cores, and 180 in the low pollen content P2 core, were counted. The pollen sum (PS) accounts for the total number of pollen grains. The pollen zonation relied on core visual examination, litho-chronological correlation among the three cores and cluster analysis (Orloci’s chord distance) run with the TGView, the same software that was used to draw the pollen diagrams (developed and distributed by Dr E. Grimm at the Illinois State Museum; Grimm, 2004).
Results
Geomorphology
Two strand plains, trending NW-SE and referred to as the Eutyrrhenian (Pennetta et al., 2011) and the Holocene (hereafter ESP and HSP, respectively), border two distinct, wet and depressed zones to the north (NDZ) and to the south (SDZ) of the present Garigliano River channel (Figure 1). The former strand plain, which is triangular in shape, is locally up to 1.5 m b.s.l. Its shorter side parallels the river through some 800 m, and in turn, the longer one stretches some 1.4 km along the HSP. The trapezoidally shaped SDZ is up to 1.2 km wide and extends some 4 km parallel to the coast. This depression locally deepens to 2.5 m b.s.l. From the analysis of historical maps, it was observed that up to the 18th century, both depressions were partially submerged; at present, they are kept dry by ongoing land reclamation works.
Based on the aerial photographs, palaeochannel traces up to 50–70 m wide have been recognized in the NDZ, both in between the present river channel and the SDZ and along the eastern side of the ESP. They are of some concern because they record the past wandering of Garigliano River.
In addition, the numerous Roman sites along the river and the remnants of the Marica sanctuary soundly prove that the river course has not changed at least since Roman Times. Some additional and very subordinate palaeochannels have also been identified; their origins, however, can be referred to as ephemeral events. It is noteworthy that a straight trace that is less than 10 m wide, N-S oriented and crossing part of HSP has been recognized between the southern apex of the NDZ and the area closer to the river mouth. Its location, width and almost perfect rectilinearity are suggestive of an ancient artificial channel. However, although a channel connecting the NDZ to the river is reported on a map issued in 1714, any reliable evidence that matches the identified trace is lacking.
Currently, the HSP width maximizes at 600 m. The beach ridges to the north of the river channel are only 2–3 m high and more spaced than their southern analogues; in contrast, the latter are from 3 to 9 m high. The beach ridges assessment rules out a significant lateral shift of the river mouth and indicates two progradation phases separated by a severe erosional event. The most ancient phase is better recorded in the southern delta wing where the ‘c’ accreting pattern (Kukavicic and Pranzini, 2003) of the ridges suggests a possible 350 m seaward shift of the river mouth relative to its 1943 location. Subsequently, the beach ridges were truncated (‘g’ eroding pattern after Kukavicic and Pranzini, 2003), which triggered a landward shift of the river mouth of approximately 700 m. The subsequent progradation phase, witnessed by the ‘c’ and ‘d’ accreting patterns in the southern wing and the ‘c’ and ‘b’ patterns in the northern one, again resulted in some 500 m of progradation compared with the 1943 river mouth location. The Roman pottery in the sediments of the last progradation is consistent for assigning a probable pre-Roman age to the erosion event.
Lithology
The detailed lithology from the mechanical cores drilled in distinct areas and the main features from the manual drillings are discussed in this section (Figure 2).
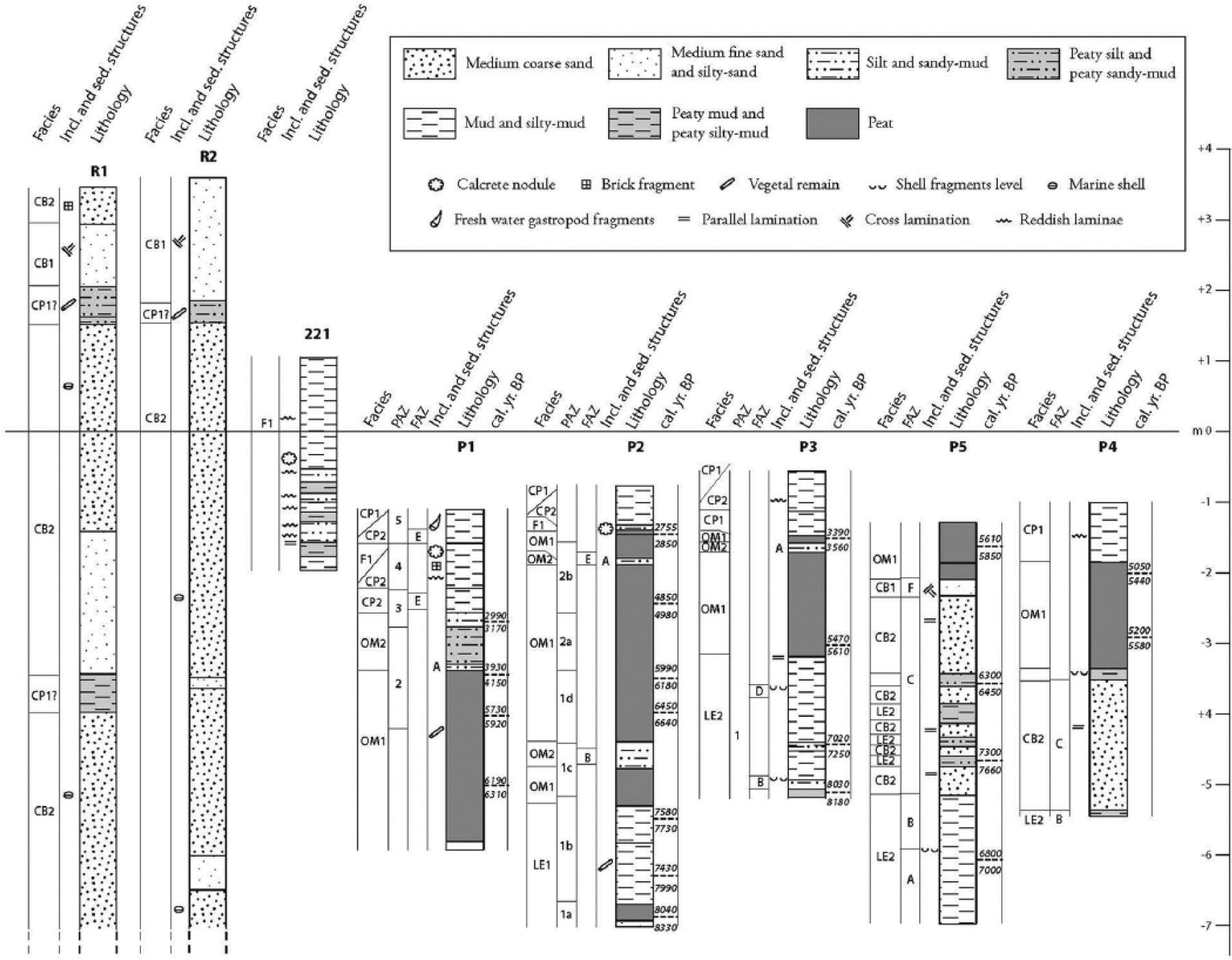
A summary of the features shown by the mechanical cores from the depressed zones (P1 through P5) and in HSP (R1 and R2). A manual core is also shown, and it is characterized by fluvial phases from between the twin depressed zones. The stratigraphic position of the pumice level marked with an ‘A’ in cores P1, P2 and P3 is consistent with the timing of the Avellino eruption event.
Holocene strand plain
Mechanical core R1 shows a basal bed (1300–715 cm deep) of coarse sand with poligenic pebbles and marine bivalves. Upwards, an organic, dark-grey mud level (715–680 cm deep) abruptly appears and is overlain by a 190-cm-thick level of fine sand containing polygenic pebbles. The next 490–180 cm exhibits medium–coarse sand with marine bivalves, carbonates and volcanic pebbles. From 180 to 140 cm in depth, a sandy-mud level with altered plant remains and rare carbonate pebbles occurs. The succession is capped by well-sorted, ochreous sand containing bladed pebbles and brick fragments.
Core R2 shows coarse sand, polygenic bladed pebbles and marine bivalves at the bottom (1200–1000 cm deep). Upwards (1000–200 cm), a medium–coarse sand bed with subrounded or bladed fine pebbles and scarce marine bivalves occurs. Its base displays two fine sand levels with mud laminae. Then, at depths from 200 to 170 cm, a muddy-sand level with vegetal remnants is recorded. The sequence is capped by well-sorted ochreous fine sand with carbonate clasts.
The seven manual cores (from depths of 1.45 to 1.80 m) drilled along the trace of the straight channel joining the NDZ southern apex and the river channel consist of fine ochreous sand resting on medium grey sand. The transition between these two sediments lies at approximately 100 cm and at 50 cm in depth inside and outside the rectilinear trace channel, respectively.
Depressed areas
Mechanical cores P1 and P2 are from the NDZ, and P3, P4 and P5 are from the SDZ. The stratigraphy is as follows:
P1 – Three main layers occur: peat with vegetal debris (465–230 cm deep), peaty-silt (230–170 cm deep) gradually changing into grey-green mud (170 cm deep/top) with rare mollusc and brick fragments and thin silt intercalations.
P2 – Three main layers are exhibited: dark-grey mud levels (625–450 cm deep) with wood fragments (up to 5 cm long), peat and grey silt intercalations. Then, at depths from 460 to 65 cm, a peat layer under the topmost 65 cm thick grey mud layer is shown. We note that the latter contains two pumice levels that, based on geochemical analyses, refer to Roccamonfina volcanic activity. The peat layer interbeds two silty-sand levels. The deeper (440–360 cm deep), ochre-coloured level shows medium sorting, a sharp bottom and top transition along with cross lamination and mollusc fragments. The upper level (110–100 cm deep) is confined by sharp contacts, grey coloured and poorly sorted, and carries gastropod fragments and some pumice grains.
P3 – Grey and locally peaty-mud levels intercalated with levels of mollusc fragments comprise the bottom of the sediment suite (460–260 cm deep), which is separated by a sharp contact from the peat (260–120 cm). Upwards, a thin silty level with some pumice grain and muddy-peat intercalations (120–60 cm) occur, transitioning to grey-brown mud (60 cm–top).
P4 – Four main layers are shown. The basal organic mud thin level (445–439 cm deep) sharply changes into well-sorted grey sand with rare bivalves and parallel laminations (439–253 cm). Upwards, an organic mud level with mollusc fragments (253–243 cm deep) underlying peat (243–80 cm) occurs. The topmost 80 cm of the sequence comprises alternating grey and brown mud levels.
P5 – Six main layers occur. At the bottom, there is a dark-grey mud with shells and a centimetric bivalve fragment level (571–383 cm deep); then, through a sharp contact, there are at first parallel laminated grey sand and dark-grey silty levels (385–281). Upwards, a peaty-mud level occurs (281–256 cm deep) that is covered by grey sand (256–108 cm deep) and ochreous sand (108–81 cm deep) with parallel and cross-laminations and reworked bivalves at the top. A sharp transition marks the capping 80-cm-thick peat level.
The 19 manual cores from the depressed areas yielded a stratigraphy consistent with that observed in the uppermost 3 m of the nearby mechanical cores.
Area confined between the twin depressions
Here, close to river channel, no mechanical cores were drilled; therefore, the stratigraphy is that provided by the nine manual cores from 3 to 3.5 m in depth. The sediment suite includes grey-greenish mud with reddish thin silt intercalations, pumice grains, local calcrete and small round-shaped pebbles generally overlying peat and/or peaty-silt. Close to HSP, these latter organic sediments are intercalated with fine sands containing sporadic mollusc fragments. Figure 2 shows the log of one of the manual cores.
Radiocarbon dates (see Table 1)
F aunas
Molluscs: The 28 species (bivalves and gastropods) that were identified match those in modern marine, non-marine and lagoonal environmental settings in the Mediterranean area (see Table 2). The species compositions of the mollusc assemblages in the analysed cores were fairly similar.
Foraminifera: Cores P1 and P3 were completely barren of foraminifera, whereas cores P2, P4 and P5 showed appreciable foraminiferal content, particularly in the lower part of the succession. In total, 14 benthic species belonging to 11 genera were identified. The miliolid group included species related to the Adelosina, Massilina, Miliolinella, Pseudotriloculina, Quinqueloculina, Siphonaperta and Triloculina genera and only occurs in core P5, particularly in the lowermost portion (575–400 cm deep). Ammonia tepida and Haynesina germanica, ranging from 33% to 90% and 6% to 72%, respectively, are constantly present in all of the foraminifera-bearing samples. Further taxa account for less than 6%. Despite being very thin, the foraminifera tests generally are well preserved; only some miliods were fragmented or altered. In core P5, the foraminifera content was at a maximum and showed a decreasing trend from the bottom to the top. Poor faunal diversity was observed, apart from core P5, where the number of species first rises towards the middle of the core; upwards, it declines and disappears.
Results of the radiocarbon (14C) dating. The calibration was performed with the software after Ramsey (2005).
Foraminifera and mollusc assemblages for each FAZ and mechanical core.
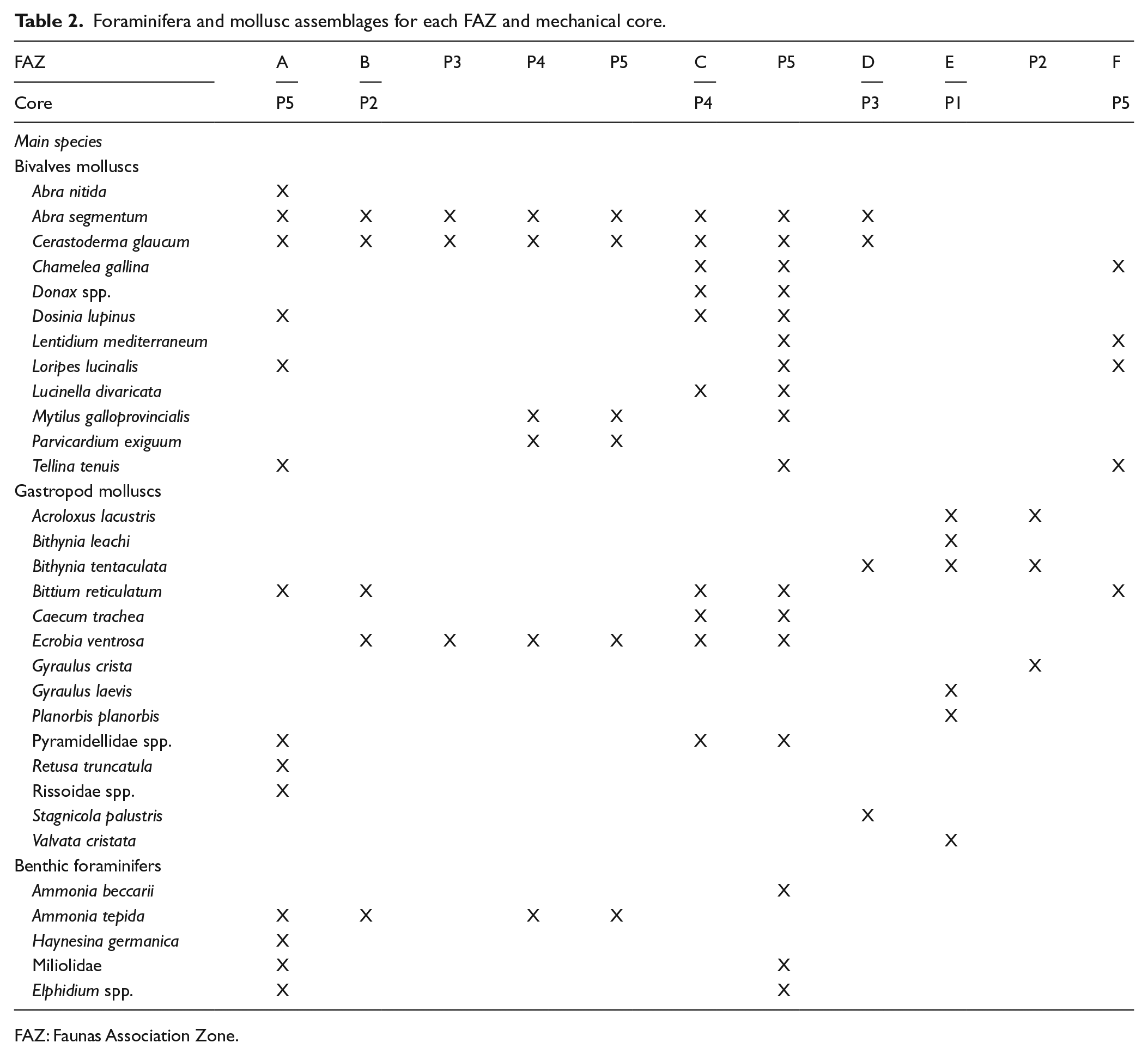
FAZ: Faunas Association Zone.
The recognized distinct foraminifera and mollusc assemblages (Table 2) were effective for distinguishing different palaeoecological environments. For the stratigraphic positions of each FAZ, see Figure 2.
FAZ A (core P5, 575–460 cm, older than 7500 yr BP)
Here, the marine euryhaline taxa dominate. Bittium reticulatum (12–35%) and Loripes lucinalis (12–27%) prevail, and Abra segmentum, Cerastoderma glaucum, Parvicardium exiguum, Tellina tenuis and Rissoidae spp. account for 0–8.5%, 0–17%, 0–4%, 0–4%, 0–6% and 1–14%, respectively. These species tolerate changes in salinity and commonly populate the modern paralic environments of the Mediterranean area. However, the presence of marine species such as Abra nitida, Dosinia lupinus, Retusa truncatula and Pyramidellidae spp. indicates the input of marine waters, a feature matching the benthic foraminifera assemblage, which shows enhanced diversity because of a mixture of marine shallow water taxa (miliolids, Elphidium spp., Ammonia beccarii, Lobatula lobatula, Neoconorbina terquemi and Rosalina bradyi) and euryhaline species (A. tepida, Ammonia parkinsoniana and H. germanica) (Langer, 1993; Murray, 1991; Sgarrella and Moncharmont-Zei, 1993). At the top of the zone, the occurrence of the brackish species Ecrobia ventrosa, with a low percentage (7%), and the decrease in foraminifera diversity (mainly typical marine taxa) pinpoint the transition to a more restricted environment.
FAZ B (core P2, 395–380 cm; P3, 495–430 cm; P4, 450–435 cm; P5, 460–385 cm; ranging in age from ca. 8000 to ca. 7000 yr BP)
A common feature is that brackish and marine euryhaline species (E. ventrosa, C. glaucum, A. segmentum, Mytilus galloprovincialis, P. exiguum) are well represented; in contrast, typical marine species (e.g. D. lupinus, Lucinella divaricata, Caecum trachea, R. truncatula and Pyramidellidae spp.) as well as freshwater species (Pisidium sp., Acroloxus lacustris, Bithynia tentaculata, Lymnaea peregra and Theodoxus sp.) are very rare. Accordingly, the foraminifera assemblage is dominated by euryhaline taxa, such as A. tepida, and by a reduction in plain marine species; thus, it exhibits decreased diversity. Because it is tolerant to a wide range of salinities and temperatures, A. tepida points to paralic environments such as estuaries and lagoons (Debenay et al., 2000; Di Bella et al., 2011; Frezza and Carboni, 2009; Jorissen, 1988; Mendes et al., 2004). The association of this species with H. germanica, Haynesina depressula and Porosononion granosum, which frequently are found in lagoonal settings (Albani et al., 1998; Jorissen, 1988), suggests a tolerance to increased nutrient supply (Jorissen, 1988; Murray, 1991). In core P5, this zone trends upwards towards more constrained conditions, as highlighted by the rise of the brackish E. ventrosa (from 13 to 61%) and the parallel decrease in euryhaline A. segmentum (from 33 to 2%), C. glaucum (from 50% to 6%) and the foraminiferal content.
The most restricted conditions are highlighted at a depth of 410 cm, where E. ventrosa has a maximum of 61%, and rare specimens of the freshwater gastropod Theodoxus sp. and only two foraminiferal species, A. tepida and H. germanica, were found. As a result, the assemblage mirrors a brackish environment, such as a lagoon or a moderately confined infralittoral environment, that is affected to some extent by freshwater input.
In contrast, in the sample from a depth of 395 cm, that is, at the top of the zone, an environmental change towards more open conditions is recorded by the decrease in E. ventrosa to 40% and the rise in P. exiguum (24%), C. glaucum (11%) and Pyramidellidae spp. (7%), along with the occurrence of customary marine foraminiferal taxa (mainly miliolids).
A similar environment can be inferred at the bottom of core P4 (at 445 cm), where C. glaucum (30%) prevails over E. ventrosa (15%), and some marine species, such as L. divaricata, C. trachea and Pyramidellidae spp., are poorly represented.
More restricted environmental conditions, which compare with those recorded in core P5 (at 410 cm), are seen in core P2 (at 380 cm). Here, E. ventrosa strongly prevails (81%); marine euryhaline species like A. segmentum (8%), B. reticulatum (7%) and C. glaucum (2%) occur; and rare specimens of freshwater gastropods (Pisidium sp., A. lacustris, B. tentaculata, L. peregra and Theodoxus sp.) were recovered. In summary, the foraminiferal assemblage in this sample, which was dominated by A. tepida and H. germanica, denotes typical holigotypic characters. Finally, in the sample at 440 cm from core P3, the most represented species are A. segmentum (55%), E. ventrosa (16%) and C. glaucum (15%).
FAZ C (core P4, 435–250 cm; core P5, 385–105 cm, between ca. 7700 and ca. 6000 yr BP)
The brackish E. ventrosa persists, although it is less abundant than in FAZ B. The euryhaline molluscs A. segmentum, B. reticulatum, C. glaucum, L. lucinalis and T. tenuis, along with foraminifera such as A. tepida, also occur. Notably, the typical marine shoreface taxa Lentidium mediterraneum, Chamelea gallina and Donax spp. appear for the first time. Accordingly, the foraminifera assemblage exhibits a significant frequency of shallow marine taxa (e.g. A. beccarii and miliolids, core P5). In this zone, the observed richer faunal assemblage and diversity relative to those of the two above zones suggest that the influence of marine water grew in importance.
FAZ D (330–310 cm; P3, ca. 6000 yr BP)
FAZ D is characterized by the co-occurrence of marine euryhaline (A. segmentum and C. glaucum) and freshwater species (Bithynia spp., Stagnicola palustris and Valvata piscinalis).
FAZ E (core P1, 140–120 and 50–25 cm; core P2, 120–100 cm, younger than 3000 yr BP)
Only freshwater species living in standing, vegetated waters (i.e. A. lacustris, B. tentaculata, Gyraulus laevis, Planorbis planorbis and Valvata cristata) occur.
FAZ F (core P5, 105–80 cm, ca. 6000 yr BP)
The marine shells and shell fragments uncovered here are significantly weathered, thus providing evidence for subaerial exposure.
P alynology
Pollen preservation and diversity were good in most of the samples (Figure 3). Seven samples from core P2, which were too poor in pollen, were not reported in the diagrams. As a rule, arboreal pollen (mean: ca. 60%) dominates.
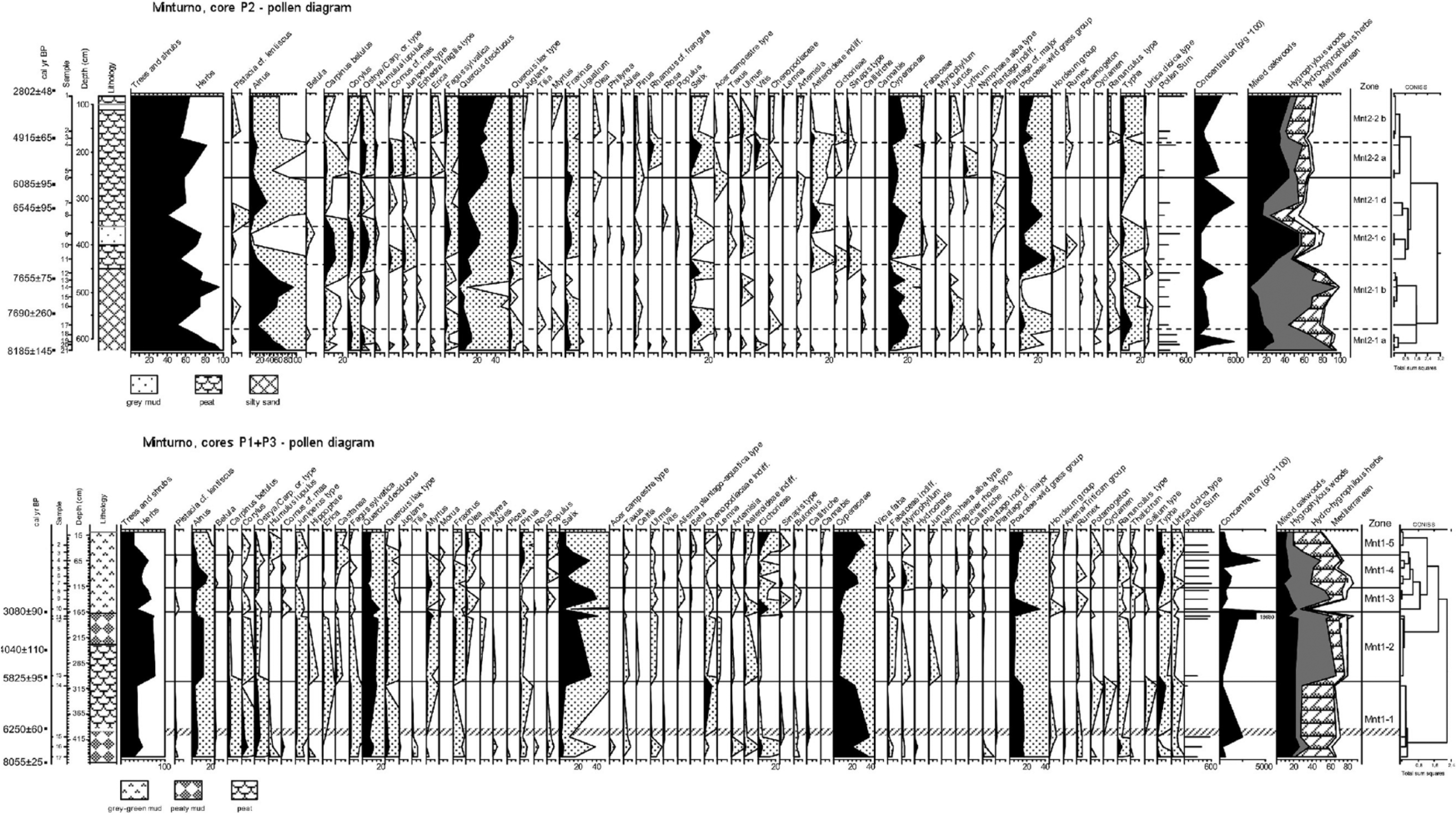
Percentage pollen diagrams of the Minturno (Mnt), Garigliano Delta Plain. The black curves of selected taxa (with enhancement curves × 10) are calculated on a Pollen Sum that included all pollen grains. Bottom: cores P3 and P1 (3 and 14 samples, respectively); the screen line separates the cores, whereas the dotted line separates the Mnt1 pollen zones. Top: core P2 (21 samples); the dotted lines separate sub-zones (a-d) within the Mnt2 pollen zones. Pollen counts, pollen concentration per gram of sediment, selected totals, zone names and cluster analyses are reported. The pollen taxa nomenclature follows Moore et al. (1991), as well as the Northwest European Pollen Flora (Janssen et al., 1974 and following). Chronology refers to the calibrated ages (see the text), whereas the pollen curves follow the alphabetical order of the botanical families.
Alnus is the dominant tree (27% in P2; 7% in P1 + P3), together with Salix (2% in P2; 18% in P1 + P3) and the deciduous Quercus (7% in P2; 11% in P1 + P3). Mixed oak woods are also represented by Carpinus betulus, Corylus and Ostrya carpinifolia/Carpinus orientalis type and by traces of Fraxinus, Ulmus, Tilia and Acer campestre type. The Mediterranean trees and shrubs are Pistacia cf. lentiscus, Quercus ilex type, Myrtus and Phillyrea. Olea may be included, but its significance as a cultivated tree is very important. Actually, its pollen, with those of Juglans and Castanea, which were both recorded in the cores, form the ‘OJC group’ that is the basis of the understanding of the development of cultural landscapes on the Italian peninsula (Mercuri, 2014; Mercuri et al., 2013). Hygrophilous Cyperaceae-sedges (6% in P2, 20% in P1 + P3) and the Poaceae wild grass group (3% in P2; 12% in P1 + P3) prevail among the herbs. Local freshwater habitats are consistently marked by limno-telmatophytes. Chenopods, pointing to brackish water habitats, are common but not abundant; hints of low water salinity are provided by the scant halophilous plants. Among the herbs, only Asteraceae, including Cichorieae, are common (0.4% in P2; 4% in P1 + P3).
Given that the spectra likely result from both wind and water transported pollen, they yield a picture of the plant associations at different altitudes and distances from the study site; thus, the pollen spectra denote the co-presence of plants from local wet areas and dry regional environments. The Local Pollen Assemblage (LPAZ) and the sub-zones identified in the three studied cores are as follows.
Sequence P1 + P3 (450 cm, 17 samples; five zones; last 8000 years)
Plants from wet environments, mainly Salix, Alnus and Cyperaceae, dominate, as well as oak (Quercus deciduous) and grasses (Poaceae wild grasses) are well represented (Figure 3, bottom). The forest decreasing trend depends on the gradual dropping of oak wood paralleled by the increase in Poaceae; at the final phase, the fall of hygrophilous woods matches the increase in Cyperaceae. Algae such as Botryococcaceae, Zygnemataceae (including Mougeotia) and Coleochete (an epiphytic alga on macrophytes) mark the wet habitats (Poulíčková et al., 2006).
LPAZ Mnt1-1 (450–302 cm; P3, samples 17, 16, 15; P1, sample 14; from ca. 8000 to ca. 5800 cal. yr BP)
The forest cover, with 11% of Quercus deciduous, just attains 38% on average in these spectra. The hygrophilous trees are Salix (3%), Alnus (5%) and Populus (0.4%). The broadleaved C. betulus (2%) and O. carpinifolia/C. orientalis type (3%) are well represented, and Corylus (3%) and Juniperus type (1.4%) are the best-represented shrubs. Q. ilex type and Myrtus represent Mediterranean taxa (2.6%). Among the OJC group, only Juglans is recorded (0.1%). There are high values of Cyperaceae (30%) and Poaceae (15%), and 2% of Cichorieae. Chenopodiaceae (2%) maximizes atop the zone (9% in sample 14), where Typha is also high (3%).
Although the samples originated from cores at different distance from the coast, the spectra show that a mosaic of freshwater and brackish water environments coexisted during a phase with open environments and mixed woods located far from the site.
LPAZ Mnt1-2 (301–166 cm; P1, samples 13, 12, 11; from ca. 5800 to ca. 3100 cal. yr BP)
Forest cover doubles (74%), especially because of hygrophilous trees (Alnus 11% and Salix 31%), whereas C. betulus and other broadleaved trees/shrubs decrease; Ulmus starts a continuous curve. Quercus deciduous (13%) and Juniperus type (1.8%) remain steady. It is noteworthy that the Mediterranean taxa increase (8%), and Myrtus (5%) is well represented atop of the sub-zone; myrtle, an entomophilous species, could point to the locally enhanced expansion of evergreen vegetation, possibly because of rising temperatures. Olea and Castanea occur (OJC = 0.6%), and the chestnut shows an almost continuous curve. Cyperaceae drastically decrease (9%) together with Poaceae (6%), Chenopodiaceae (1%) and Cichorieae (0.2%). As for the wet environments, Typha (2%) drops, whereas Juncus (0.5%) and Myriophyllum (0.3%) steadily remain significantly low. Finally, hygrophilous woods spread along the riverbed, probably in response to enhanced wetness.
LPAZ Mnt1-3 (165–117 cm deep; P1, samples 10, 9, 8; from ca. 3100 to ca. 2700 cal. yr BP)
Although the curve of forest cover (54%) fluctuates, overall significant droppings are noted for Alnus (4%) and, although less enhanced, for Salix (26%), as well as for many telmatophytes (Typha). In contrast, Populus (0.5%), as well as several freshwater aquatics (Callithriche, Nymphaea alba type and Potamogeton), increases and spreads. Quercus deciduous decreases gradually (from 19% to 9%), which is analogous to Juniperus type (0.8%) and the Mediterranean taxa (3%). The OJC group includes all the three trees (1%) and provides evidence of human activity in the area. Cyperaceae start to increase again (from 5% to 13%); Poaceae wild grasses (15%) peak at 33% in sample 10, matching the maxima in the cereals (2% in the same sample, both Hordeum and Avena/Triticum groups), and then they decrease again. Cichorieae (5%) become more abundant, reflecting the spreading of pastures (Florenzano et al., 2015; Mercuri et al., 2010) and, together with cereals, the development of agro-pastoral systems.
LPAZ Mnt1-4 (116–55 cm; P1, samples 7, 6, 5, 4; from Roman to Middle ages?)
Forest cover remains fairly steady (52%), although the hygrophilous tree assemblage changes: Alnus increases (11%), and Salix decreases (19%) again. Deciduous Quercus decreases to 9%. Myrtus again gains importance (mean 4%), and in sample 7, it peaks at 8%. At Lago d’Averno, the Roman landscape witnessed enhanced forest cover with abundant deciduous vegetation (mostly oaks, hornbeams and hazels), and the evergreen vegetation included increased shrubs, mainly Pistacia and Myrtus. The OJC group (Juglans is absent) accounts for 1.2%, and Castanea increases. Poaceae start to gradually rise (from 5% to 10%), and Typha (5%) and Cyperaceae (21%) again increase significantly. The notable Myriophyllum occurrence (mean: 3%) includes both Myriophyllum verticillatum (3% in sample 7) and Myriophyllum alterniflorum (4% in sample 6). Both species are known to require adequate lighting and soft water (Chatenet et al., 2006). Chenopodiaceae remain insignificant (0.6%), Cichorieae decreases (2%), and the cereals total 0.3%.
The zone records a further expansion phase in evergreen vegetation and the spreading of freshwater environments. However, here the hygrophilous woods developed less than in LPAZ Mnt2. Furthermore, the dramatic fluctuations in Salix abundance could suggest that willows were exploited by humans.
LPAZ Mnt1-5 (54–10 cm; P1, samples 3, 2, 1; modern age)
Forest cover drops notably (34%), owing to the decline of riparian trees (Salix 8%; Alnus 3% and Populus 2%); thus, it again approaches what was observed in LPAZ Mnt1. Quercus deciduous (8%) and the other broadleaved trees remain steady. Humulus lupulus (2%) rises significantly, and the Mediterranean taxa minimize at 1%. On the other hand, the OJC group, including pollen from the three trees, maximizes at 4%. Typha (6%), Cyperaceae (28%), Cichorieae (7%) and the Poaceae wild grass group (13%) expand, in addition to the cereals (0.8%), thus providing evidence of agrarian systems with pastures and fields in the area.
Sequence P2 (625 cm, 21 samples; two zones and six sub-zones; last ca. 8200 years)
The spectra, which point to a more regional pollen rain, show a general decrease in Alnus that matches the gradual increase in Quercus deciduous, Poaceae and Cyperaceae (Figure 3, top).The pollen was deposited over a short time under high sedimentation rates, and therefore, the improved resolution provides detailed data for the two zones corresponding to the earlier phases (LPAZ Mnt1-1 and Mnt1-2 of P1 + P3).
LPAZ Mnt2-1 (625–248 cm; P2, samples 21 to 6; from ca. 8200 to ca. 5800 cal. yr BP)
The initial widespread forest cover changed into open and freshwater environments:
Mnt2-1a (625–580 cm; samples 21–18): The impressive expansion of the forest cover (mean: 85%; peaking at 97% in sample 21 at 625 cm) was largely because of Alnus (57% on average). Alder abundance and the lithostratigraphy suggest that Alnus grew locally in a riparian wood. Deciduous Quercus (11%), Fraxinus and Corylus (4% each) pollen were recruited from oak woods (22%). The OJC group (0.1%) is only represented by Juglans. Cyperaceae (8%) and Typha (2%) are the only herbs reaching significant values.
Mnt2-1b (581–440 cm; samples 17–12): The forest cover decreases (mean: 75%), deciduous Quercus drops to 7% and then (sample 14, 490 cm) the woody pollen peaks at 97%. Myrtus (0.3%) is common, and the OJC group is completely missing. Wet habitats spread, with Alnus (54%) and Cyperaceae (11%). Typha first peaks (6% in sample 17) and then declines.
Mnt2-1c (439–360 cm; samples 11, 10, 9): Important changes are recorded here. Oak woods expand (47%; compared with the 12% of the previous sub-zone), mostly because of C. betulus (11%), Corylus and O. carpinifolia/C. orientalis (6% each). The Mediterranean taxa increase to 6%, including Q. ilex type (5% vs 1%). Meanwhile, hygrophilous woods and herbs contract (17% vs 74%) where habitats with aquatics prevail following the withdrawal of the riparian woods. Cyperaceae decrease (7%), and the Poaceae curve (14%) shows a steady increase shared in part with Asteraceae. A significant 3% of cereals are found in sample 11 (430 cm).
Mnt2-1d (359–254 cm; samples 8, 7, 6): Forest cover decreases to 53%, and pollen from wet environments increases again (29%), with Alnus blooming at 18% (sample 7, 310 cm). Oak woods reduce to 26%, and Q. ilex type (still 5%) is high at the bottom and drops upwards. Cyperaceae (10%) and Poaceae (18%) expand, thus pointing to spreading grasslands. Some record of Salicornia, a halophilous species among Chenopodiaceae (0.9%), is found in sample 6 (255 cm). A mere 0.2% of cereals were found.
LPAZ Mnt2-2 (253–80 cm; P2, samples 5 to 1; from ca. 5800 to ca. 3100 cal. yr BP)
An increasing trend in the mixed oak wood, which was synchronous with the increasingly widespread freshwater environments, is noticed:
Mnt2-2a (253–180 cm; samples 5, 4): The forest cover reaches 73%, with an increase in oak wood (39%, including deciduous Quercus 28%). Salix is high (7%), and Alnus accounts for 10%. Rhamnus cf. frangula and Vitis are also high (5% each) in sample 5 (185 cm). In addition, Cornus mas (4%) is significant. Cyperaceae drop to 5%, and Typha (3%) and Lythrum (2%) dominate the wet environments.
Mnt2-2b (179–80 cm; samples 3, 2, 1): Forest cover decreases to 58%, especially because of the definitive fall of Alnus (7%). The increasing trend in deciduous Quercus (32%) in the uppermost part of the diagram could correspond to a regional retrieval of riparian woods and a spreading of oak woods. A comparison with the LPAZ Mtn1-2 from the P1 + P3 diagram suggests that such changes occurred on a regional scale. Finally, Myrtus (0.4%) is present again, and Cyperaceae again increases to 15%.
Discussion
Architecture of the sedimentary succession
The results, particularly those from the mechanical cores, denote for the first time the lateral–vertical relationships among the following depositional elements (Figure 4).
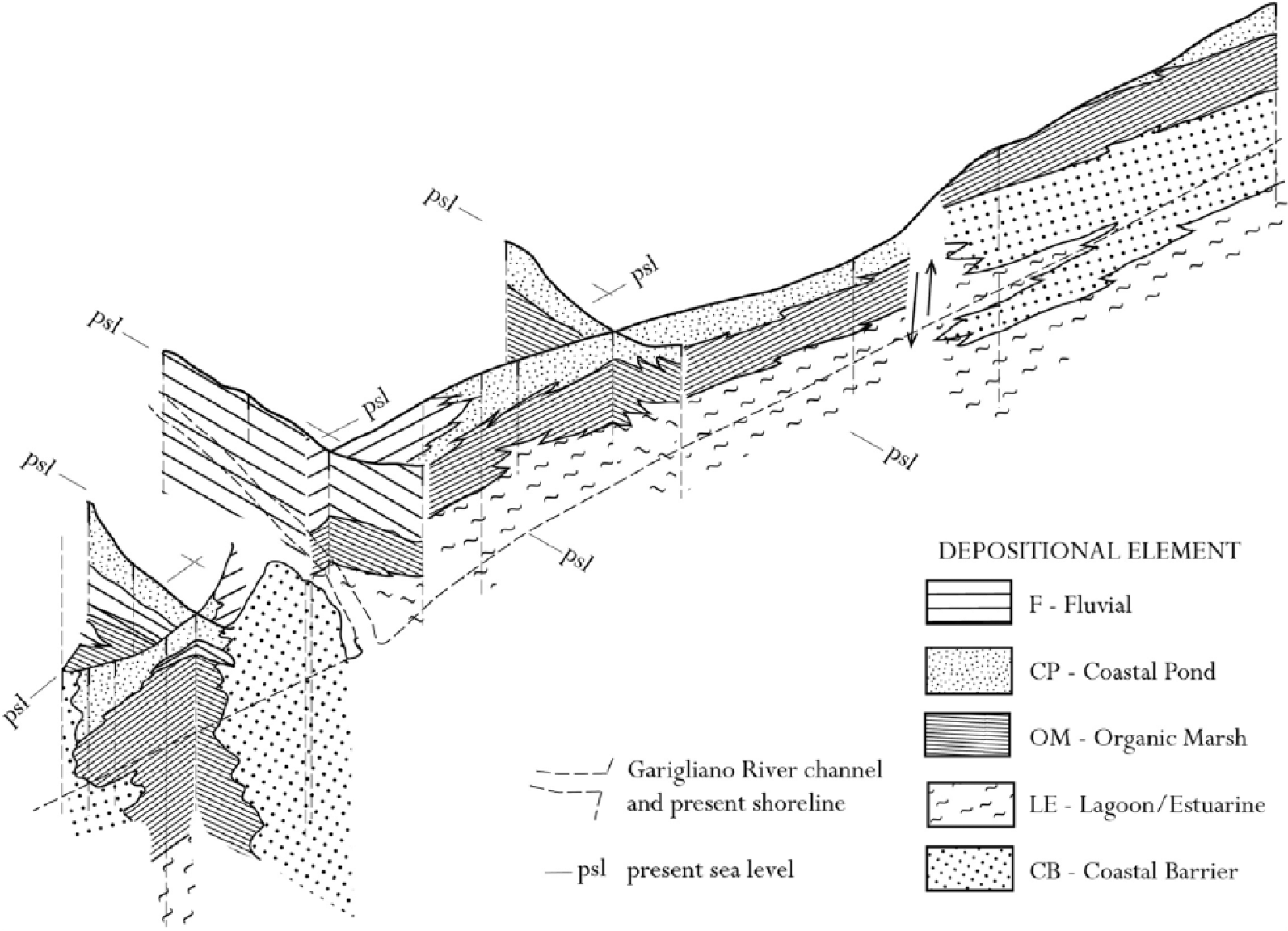
Fence diagram showing the stratigraphic correlation among the depositional elements.
Lagoon-estuary
The lagoon-estuary (LE) element occurs at base of the sediment suite and shows two facies.
LE1 (lower P2 core portion) comprises dark-grey mud levels that include wood debris, peat and grey silt intercalations. The sediments (mean depositional rate: 3 mm/yr) occurred from 8200 to 7500 yr BP. Fauna remnants are lacking, but freshwater wet environments are mirrored by the pollen LPAZ Mnt2-1a and 1b.
LE2 (bottomward cores P3, P4 and P5) includes mud and silt with local levels of peat or mollusc fragments along with intercalated sandy bed with parallel lamination (comparable with coastal barrier/beach, hereafter CB, and discussed later). The sequence is accumulated from 8000 to 5500 yr BP at rates from 0.7 to 1.1 mm/yr. The FAZ (A, B and C) and LPAZ Mnt1-1 consistently suggest a mosaic of wet environments that included fresh, brackish and marine habitats. As a whole, the above facies record a lagoonal/estuarine environment that developed between the end of the postglacial sea level rise and the early stage of the subsequent quasi-still stand.
In particular, the LE1 facies reflect a humid environment that probably had some fluvial inputs and had wetlands rich in hygrophilous woody vegetation. In turn, LE2 partly reflects an ongoing lagoon infilling under variable marine and fluvial inputs.
Organic marsh
Organic marsh (OM) overlies LE in both the NDZ and SDZ, whereas in the southern area, it rests on CB. Two facies are represented.
OM1 comprises a homogeneous peat suite with well-preserved vegetal remnants and is devoid of fauna, corresponding to LPAZ Mnt1-1 and Mnt2-1. It shows an increasing spreading trend in hygrophilous woods along the riverbeds. To the north of the Garigliano River, sedimentation occurred from 7500 to 2800 yr BP. In the opposite direction, the sedimentation began at approximately 5500 yr BP (cores P3 and P4). It lasted up to ca. 5200 yr BP in core P4 and up to 3500 yr BP in the P3 core, which is closer to the river.
OM2 includes grey-greenish silts that are sometimes peaty (with occasional FAZ E), and it locally contains pumice grains and grey medium–fine sand with parallel laminations (showing the FAZ B).
Both facies reflect a marshy environment with almost continuous sedimentation of organic litter locally interrupted by levels of clasts that, according to the fauna remnants, can be regarded as washover fans or crevasse splays. The stratigraphic position of level A (Figure 2) suggests that the pumice likely issued from the Avellino eruption (3945 ± 10 cal. yr BP, according to Sevink et al., 2011). The sedimentation rate ranges from 0.6 to 0.8 mm/yr, apart from the huge value >6 mm/yr observed in core P4.
Coastal ponds
CP lies on OM and is common in the upper part of the sequence, both in the NDZ and SDZ. It only lacks core P5 where OM outcrops. It includes two distinct lithofacies.
CP1 comprises dark-grey or brown mud with local peat levels. The fauna only include rare freshwater gastropods, whereas pollen analysis denotes LPAZ Mnt1-2, Mnt1-3, Mnt1-4 and Mnt1-5. The facies deposition occurred from 4000 to 2800 yr BP, subsequent to the decline of the OM sedimentation. Obviously, any further accumulation ceased following the reclamation of land in the 19th century.
CP2 is only represented by an intercalation of grey-ochre silt levels with rare laminations and pumice grains within CP1.
The facies mirror a freshwater CP in which CP1 records a quiet deposition governed by decantation and CP2 inputs left by flooding events. The facies history shows that drying phases alternated with shallow limpid ephemeral ponds prior to the progressive drying up. A sedimentation rate from 0.3 to 0.5 mm/yr has been observed.
Fluvial
Fluvial (F) includes just one facies (F1) and is spread between the NDZ and SDZ near the present Garigliano River. It mainly includes grey or grey-greenish mud levels with thin reddish silt laminae interbedded. Locally, calcrete nodules, small rounded pebbles, pumice and parallel lamination can be observed, along with subfossil tree roots and rare brick fragments. The facies, which is locally interdigitated in CP and OM, refers to an alluvial deposit mostly derived from overbank events.
Coastal barrier
CB outcrops throughout the seaward area and is more than 10 m thick. It is buried only locally under OM. It includes two facies.
CB1 comprises ochre-coloured, well-sorted fine-medium sand locally exhibiting cross-laminations. Rare lamellar or subrounded carbonate and volcanic pebbles and brick fragments are observed, along with scant marine shells. The facies, which is less than 3 m in thickness, outcrops between the sea and the depressed areas; it has also been found, barren in brick fragments, at bottom in core P4.
CB2 never outcrops and as a rule occurs below CB1. The suite, which ranges in thickness from 2 (cores P4 and P5) to 7 m (cores R1 and R2), comprises grey medium–fine to medium–coarse sand with subrounded or bladed carbonate and volcanic clasts. Locally, marine shells and low-angle parallel lamination are observed.
Both lithofacies refer to the CB that were built during the final phase of the postglacial transgression and the subsequent strand plain formation. CB1 records both beach and dune ridges; CB2 records the next foreshore and shoreface.
We would like to note an intriguing puzzle. It is well known that in coastal environments, peat deposits approximately occur at sea level (Vella et al., 1999). However, according to previous data on sea level changes (Alessio et al., 1992; Lambeck et al., 2010), at 8000 yr BP, the sea level was some 10 m below the present one. This conflicts with the ca. 8000 yr BP peat layers found at 7 and 5 m b.s.l. in the P2 and P3 cores, respectively. A similar discrepancy is also seen in the 5500 yr BP peat at depths of 1.5 and 2.5 m in cores P5 and P4, respectively, given that in those times the sea level was 3 m below the present one. As a tentative explanation for these systematic discrepancies, we can invoke a sediment suite uplifting prior to 3000 yr BP, which would be contemporary to the emersion of the marsh bottom (as shown in core P5). The timing of the uplifting event is constrained by the 3000 yr BP old peat level at a depth of 1.5–2.0 m and is fairly compatible with the contemporary sea level (from 1 to 2 m below the present sea level). This hypothesis receives some support by the huge sedimentation rate of the marshy sediments (>6 mm/yr, core P4), and it is conceivable that the vertical dislocation, which would have been responsible for a significant sliding of sediments towards the most depressed areas, determined the observed striking thickness.
The hypothesized uplift implies the existence of a recent vertical tectonic until now excluded in this area during the Holocene.
Diachronic environmental changes
The architecture of the sedimentary succession shows the evolution of a coastal/deltaic depositional system from the last phase of the postglacial sea level rise up to the present high stand (Figure 5).
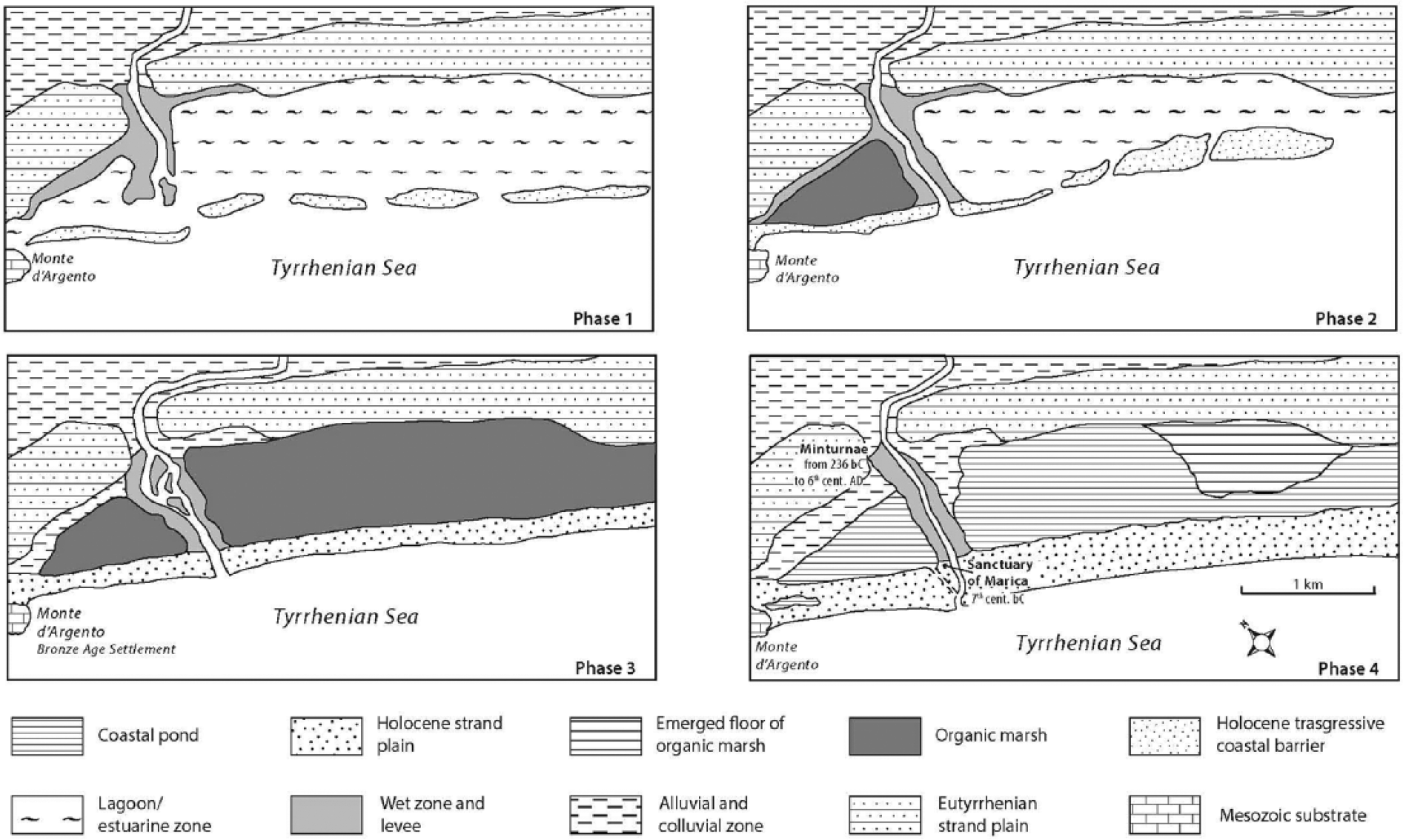
Sketch of the evolutionary phases of the Garigliano delta plain.
Phase 1 (8200–7500 yr BP)
A wet zone without marine influence, partially emerged and covered by hygrophilous woods, was present in the northern zone. On the contrary, in the southern part, a restricted lagoon developed. The higher sedimentation rate in the northern zone likely depended upon the river channel proximity. Accounts after Mazzini et al. (1999), Bellotti et al. (2007), Milli et al. (2013) and Sacchi et al. (2014) report contemporary morphological features in several Tyrrhenian river mouths, where lagoons or wave-dominated estuaries developed.
Phase 2 (7500–5500 yr BP)
During the final transgression stage and the beginning of the sea level still stand, the CB migrated landward and set apart, in the northern zone, a depressed area where a freshwater marsh with organic sedimentation developed. In the southern zone, the width of the lagoon diminishes. During the following quasi-still stand, the CB progressively extends, producing a mosaic of fresh and brackish water environments bordered landward by mixed woods.
Phase 3 (5500–3000 yr BP)
At the beginning, a delta cusp developed similar to several Italian wave-dominated deltas (Amorosi et al., 2013; Bellotti, 2000; Bellotti et al., 2004, 2011; Bravetti and Pranzini, 1987). Because of the CB changing into a strand plain, a freshwater marsh also developed in the southern part. Brackish environments progressively disappeared, and the river wandered between the twin marshes. Woods were widespread with an abundance of hygrophilous trees. After 4000 yr BP, because of the input of alluvial sediments, the marshes progressively changed into CP with prevailing clastic sedimentation lasting up to the uplifting of the southernmost area. Woodlands declined, and the early anthropic impact is revealed by the archaeological and pollen records.
Phase 4 (3000 yr BP to present)
Human impact on the territory increases because of farming development. The early landscape is characterized by two CP (separated by the river course) and by a partially eroded delta cusp. Close to the mouth, between the right riverbank and the northern pond, the Marica sanctuary was built. This was also the landscape on which the Roman colony of Minturnae flourished since the 3rd century BC. The town, with its main road, was founded on a drained area of the Eutyrrhenian strand plain overlooking the northern pond. The sea, just 1 km away, could be reached by the river, which lazily brushed the town, or, alternatively, the sea could be reached by the riverbanks between the twin ponds. Seawards, the ponds were bordered by a some hundred metres wide strand plain. Based on the sea level in Roman times, which was −0.5/−0.8 m according to Alessio et al. (1992) and Lambeck et al. (2010), as well as the sedimentation rate in the CP, the estimated maximum depth was approximately 1.5 and 2.5 m for the northern and southern ponds, respectively. It is noteworthy that these figures are consistent with the ancient Latin accounts, which do not mention any use of the ponds as landing facilities in Roman times, which is contrary to that claimed by Remmelzwaal (1978). Therefore, the Roman harbour could only have been located in the river channel. The pollen and fauna data rule out the use of the ponds as saltworks. The northern pond probably was considered to be a holy zone because of the Marica sanctuary. The significant presence of M. alterniflorum suggests that clear and oligotrophic water fill the ponds, probably because of the proper water management in Roman times. The subsequent lack of maintenance of the drainage system in the Middle Age resulted in enhanced solid inputs to the ponds, with consequential phases of partial drying. The ponds were never completely filled in, and the areas now must be kept dry by ongoing reclamation. Forest cover decreased contemporary to the reduction in the wetlands and the expansion of agrarian systems. Based on the historical map and the similarity with other wave-dominated deltas on the Tyrrhenian coast (Tarragoni et al., 2011, 2015), it is argued that the origin of the most recent cusp, although triggered during Roman times, developed mainly during the 15th–19th centuries, that is, at the beginning of the new erosional phase.
Conclusion
The results of the interdisciplinary research on the Garigliano delta plain highlight the following conclusions.
The Garigliano wave-dominated delta developed after 6000 yr BP. At the time, the river wandered through the delta plain, and lateral shifts in its mouth were negligible. The evolution of the delta involved two main progradational phases that were broken, probably in the pre-Roman age, by a severe cusp retreat. Between 5500 and 3000 yr BP, two marshes with organic sediments developed on the coastal plain. They then progressively changed into freshwater CP that submitted to dominant clastic sedimentation.
In contrast with previous understandings, the sedimentary succession apparently was affected by a vertical dislocation prior to 3000 yr BP.
The pollen data suggest that a fairly open environment characterized the basin, which was dominated by freshwater wetlands with scarce human impact in the middle Holocene. The pollen data show that oak and riparian woods spread regionally during the earlier phases, and the wet habitats alternately expanded and contracted. In both the dry and wet areas, forests opened and lost richness, probably because of anthropic impacts (Malavasi et al., 2014). Grasslands, probably wet pastures, spread.
During the Roman Period, the CP were unsuitable for harbour facilities or salt production. The right bank river became a port, and the zone near the river mouth was used as a sacred area, which probably included the northern pond. The human impact on the Garigliano delta landscape in the Roman Period seems to be limited compared with that in similar Mediterranean deltas (e.g. the Tiber, Arno and Rhône).
Footnotes
Acknowledgements
Thanks to Paolo Censi Neri and Maurizio Cacopardo (ISPRA) for their skilled help during the drilling, and to Maurizio Barbieri who provided details on some volcanoclastic sediments.
Funding
This work was carried out with the financial support of the Sapienza – Università di Roma and IGAG (CNR).