
Abstract
Two sediment cores from the Chukchi Sea margin were investigated for the Arctic sea-ice biomarker IP25, along with marine and terrestrial sterols and glycerol dialkyl glycerol tetraethers (GDGTs). This is the first paleoclimatic application of IP25 in the Chukchi-Alaskan region of the Arctic, which is key for understanding Arctic–Pacific interactions and is experiencing rapid sea-ice retreat under present warming. Sea-ice and related circulation conditions were characterized in this study with a multicentury resolution for the long-term Holocene record to multidecadal for the last several centuries. Sea ice was found to be present during the entire record but with considerable spatial and temporal variability. After very low deglacial IP25 values, possibly related to permanent sea ice and/or an iceberg-dominated environment, cores from the upper slope and shelf show IP25 maxima, interpreted as representing a relative proximity to the sea-ice margin, in the early (ca. 8–9 ka) and middle (ca. 5–6 ka) Holocene, respectively. Along with isoprenoid GDGT distribution, this asynchronicity in sea-ice history probably reflects oceanographic evolution of the Chukchi margin affected by the Beaufort Gyre circulation and Pacific water inflow via Bering Strait. Data for the last several centuries, with elevated values of brassicasterol and terrestrial sterols covarying with dinosterol and IP25, are interpreted in terms of long-distance import by currents combined with diagenetic transformations. We infer that high-amplitude variability in the late ‘Little Ice Age’, starting in the late 18th century, is related to the intensity of the Alaskan Coastal Current. This interval is preceded by three centuries of presumably diminished Alaskan Coastal Current but overall increased Bering Strait Inflow resulting in reduced sea-ice cover according to dinocyst-based data.
Introduction
The Arctic is highly sensitive to the changing global climate because of powerful feedbacks collectively known as the Arctic amplification (e.g. Miller et al., 2010). Sea ice plays a major role in these processes by largely controlling the surface-albedo feedback. This setting justifies a high attention of the scientific community to the persistent Arctic sea-ice retreat that has been monitored for almost four decades (e.g. Stroeve et al., 2011). In addition to increases in downwelled longwave radiation and surface air temperatures, intensified advection of oceanic heat and changing atmospheric circulation patterns are identified as the main contributing factors to the ongoing sea-ice loss (Woodgate et al., 2010; Zhang et al., 2008). In particular, recent data indicate a critical role of the Pacific water influx for warming the western Arctic, where sea-ice retreat is most pronounced (Figure 1; Shimada et al., 2006; Woodgate et al., 2010). Understanding the long-term behavior of these processes is complicated by the short duration of the instrumental record and the paucity of observations. For example, the transport of Pacific waters into the Arctic has only been measured directly for the last ~20 years (Woodgate et al., 2010).
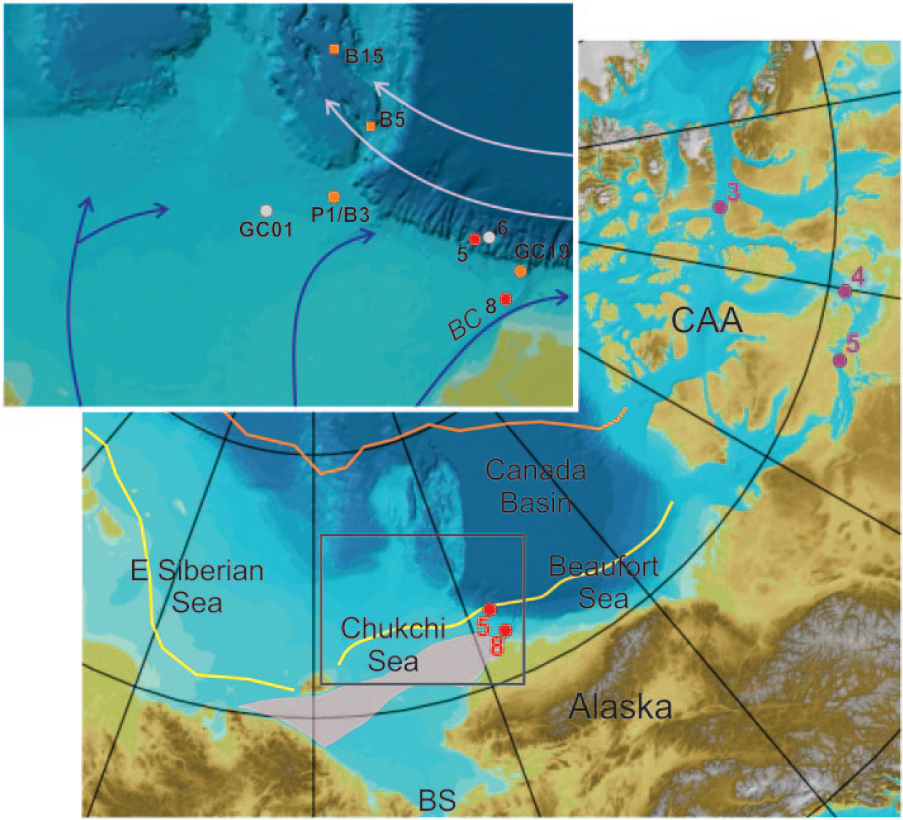
Index map of the western Arctic Ocean and the study area (inset). Cores under study (red circles) and related cores are shown. Purple indicates CAA IP25 records (Belt et al., 2010); orange circles or squares in inset indicate long or box cores with dinocyst-based reconstructions; gray indicates other cores from proxy studies mentioned in this paper; yellow and orange lines show September (yearly minimum) ice extent (15% concentration): late 20th-century mean and 2012 historical minimum; pink field south of the mean ice extent encloses maximal concentrations of IP25 measured in surface sediments (Stoynova et al., 2013); arrows in inset show Bering Strait Inflow branches (blue) and Beaufort Gyre (violet).
Comprehending the development and consequences of the emerging new state of the Arctic with expanding swaths of open water requires a broad paleoclimatic perspective. The present interglacial (Holocene) covering the last approximately 12 ka is a natural object for extended paleoclimatic research, especially considering the widespread availability of Holocene deposits (e.g. Miller et al., 2010; Polyak et al., 2010). Several studies emphasize the significance of the early Holocene (Holocene thermal maximum (HTM)) for insights into the present Arctic warming and sea-ice shrinkage (e.g. Stranne et al., 2014), despite the differences in radiative forcing (insolation vs atmospheric greenhouse composition) between these times. However, Arctic paleoclimate proxy data indicate a considerable geographic heterogeneity for the HTM. While most Arctic regions demonstrate a warming trend and diminished sea ice (e.g. Dyke and Savelle, 2001; Funder et al., 2011), some dinocyst-based reconstructions from the Chukchi margin, an area of pronounced modern sea-ice retreat, suggest the opposite pattern of sea-ice growth during the HTM (de Vernal et al., 2008, 2013). One explanation for this anomalous picture infers atmospheric and oceanic circulation in the western Arctic that favors sea-ice buildup at the Chukchi margin, combined with enhanced ice formation on the adjacent shelves because of stronger seasonality. More studies, employing high-resolution records and various proxies, are, therefore, needed to delineate the Holocene history of the Chukchi region and its applicability for clarifying the long-term consequences of current climate change. In this paper, we investigate Holocene sea-ice conditions in two high-resolution sediment core records from the northeastern (Alaskan) Chukchi Sea margin based primarily on the Arctic sea-ice diatom biomarker IP25 (Belt et al., 2007), along with the concurrent analysis of some other biomarkers (marine and terrestrial sterols and glycerol dialkyl glycerol tetraethers (GDGTs)) for further context. This study represents the first temporal application of IP25 from this region, which plays an important role in Arctic circulation and sea-ice regime.
Study area
The Chukchi Sea, connected to the Bering Sea via the narrow and shallow Bering Strait, acts as a distributor of water between the Arctic and Pacific oceans. The Bering Strait Inflow, an important carrier of heat and freshwater to the Arctic, transports the Pacific water to and across the Chukchi Sea in three major branches, which interact with the wind-driven Beaufort Gyre circulation at the Chukchi shelf margin (Figure 1; e.g. Spall, 2007; Weingartner et al., 2005; Winsor and Chapman, 2004). The eastern branch forms the Alaskan Coastal Current, a buoyancy-driven boundary current along the Alaskan coast (Weingartner et al., 2005). The western branch flows northwestward and can be especially strong, if easterly winds prevent the Alaskan Coastal Current (Winsor and Chapman, 2004). After crossing the Chukchi shelf, this branch, as well as the intermediate central branch, normally turns eastward along the shelf break (Spall, 2007). Under present conditions, thus, both the Alaskan Coastal Current and the recirculated western or central branches can affect the study sites.
Sea-ice conditions in the Chukchi Sea are strongly dependent on the wind patterns and the strength and distribution of the Bering Strait Inflow (Spall, 2007; Woodgate et al., 2010). Average spring–summer sea-ice concentrations in the late 20th century (climatological baseline) varied from ~50% near the Bering Strait to >90% at the northern margin of the Chukchi shelf, with the September ice margin (yearly minimum) well north of Alaska (Figure 1). In recent years, the margin of minimal sea-ice extent in this region has retreated considerably further north (Figure 1), with associated changes in the hydrography, primary production, and ecosystems (e.g. Grebmeier, 2012).
The Bering Strait Inflow and attendant circulation also control sedimentary environments on the Chukchi shelf and slope. Fine sediments are largely resuspended on the shallow, seasonally ice-free Chukchi shelf to be deposited on the northern slope and tributary canyons (Darby et al., 2009). The composition of these sediments may be affected by the circulation pattern and intensity. For example, some minerals associated with the North Pacific provenance, notably chlorite, can be used as a proxy of the Bering Strait Inflow (Nwaodua et al., 2014; Ortiz et al., 2009). Sedimentary organic matter can also bear evidence of the current impact as indicated by the distribution of terrestrial plant biomarkers, potentially related to the Yukon River runoff transported via the Bering Strait to the Chukchi-Alaskan margin (Goňi et al., 2013).
The Holocene history of circulation in the Chukchi Sea, as reconstructed from sediment core records, was initially controlled by the postglacial sea-level rise. The 40- to 50-m-deep Bering Strait was inundated ca. 11–12 ka (Elias et al., 1992; Keigwin et al., 2006), but its complete availability for throughflow required another several kiloannum. Based on the proxy data, maximal Bering Strait Inflow effect on sedimentation at the northeastern Chukchi margin was reached by ca. 5–6 ka (Ortiz et al., 2009), consistent with the time of sea-level stabilization. Further evolution of the Bering Strait Inflow and related Chukchi Sea currents was likely controlled by atmospheric circulation, including the strength and position of the Aleutian Low (Danielson et al., 2014) and/or the interhemispheric wind stress (Ortiz et al., 2012).
IP25 and related biomarker proxy approach
The measurement of the IP25 biomarker in marine sediments has emerged as a powerful approach for paleo sea-ice reconstruction in recent years (Belt et al., 2007; Belt and Müller, 2013). Among its properties, IP25 has been shown to be produced selectively by certain Arctic sea-ice dwelling diatoms during the spring bloom and, on ice melt, deposited in underlying sediments (Belt et al., 2007, 2013; Brown et al., 2011, 2014). As such, its sedimentary occurrence provides a relatively direct measure of the past occurrence of spring sea ice: an attribute not shared with other sea-ice proxies such as planktonic micropaleontological assemblages. Currently, further work is needed to determine whether IP25-based sea-ice reconstruction can be made more quantitative; however, it is noted that previous paleo sea-ice reconstructions based on the presence and directional abundance changes of this biomarker are consistent with outcomes from other sea-ice proxies or other oceanographic and climatic conditions (see Belt and Müller, 2013 for a review). Thus, the presence of IP25 in Arctic marine sediments provides evidence for the past occurrence of seasonal sea ice (Belt et al., 2007; Brown et al., 2011; Cabedo-Sanz et al., 2013), while changes in IP25 abundances track variability in sea-ice cover (e.g. Belt and Müller, 2013).
While being a robust indicator of sea-ice presence, IP25 does not show a straightforward relationship with sea-ice concentration or duration. The peak abundances of IP25 are expected to co-occur with sea-ice margin, from which IP25 values decrease toward both open water and permanent ice cover because of a likely reduced or even absent sea-ice diatom growth (Belt and Müller, 2013; Müller et al., 2011). However, this pattern may be less clear in areas with a more patchy distribution of sea ice, without a well-expressed ice edge (Weckström et al., 2013). Distribution of IP25 in extended sea ice may also be more complex than initially thought. The early results indicated no detectable IP25 in sediments from the Canadian High Arctic with year-round ice cover (Belt et al., 2007). However, since this first report, analytical methods for identifying IP25 have improved, including further procedures that aid its detection at low concentration (Belt et al., 2012). In particular, IP25 has recently been reported in sediments from across the Arctic Ocean with permanent or near-permanent sea-ice cover (Xiao et al., 2015a). A potential solution to the interpretation of absent or very low IP25 is through the measurement of complementary biomarker signatures of open-water conditions. An example of these is brassicasterol, a common lipid component in marine phytoplankton, whose abundance in sediments should also be sensitive to the overlying sea-ice conditions. Thus, for low or absent IP25, accompanying low brassicasterol might be expected under high ice cover, while high brassicasterol would more likely result from low sea-ice or ice-free conditions. Furthermore, Müller et al. (2011) showed that by combining concentrations of IP25 and brassicasterol (or other open-water indicators) into a single phytoplankton-IP25 index (PIP25), it may be possible to obtain more quantitative estimates of sea-ice conditions than from IP25 alone, and several applications of the PIP25 index for paleo sea-ice reconstruction have appeared in recent years (e.g. Cabedo-Sanz et al., 2013; Müller et al., 2012; Stoynova et al., 2013; Xiao et al., 2015b). However, the assumptions and limitations associated with the use of the PIP25 approach have been discussed in detail by Belt and Müller (2013), and proponents of its use continue to emphasize that more work is needed to validate this approach.
The analysis of other biomarkers such as campesterol and β-sitosterol, believed to be derived mainly from terrestrial sources (higher plants), can provide information that is complementary to IP25 regarding oceanographic settings (Fahl and Stein, 2012, and references therein). GDGTs also have the potential to provide relevant information, as marine production of isoprenoid GDGT can be affected by sea-ice cover, while branched GDGT may help identify the provenance of organic matter, especially the input of terrestrial material from soils (Park et al., 2014, and references therein).
A detailed investigation of the distribution of IP25 has been performed in surface sediments of the southern Chukchi Sea, not quite reaching our study area (Stoynova et al., 2013). The IP25 distribution pattern shows a pronounced peak zone extending sublatitudinally across the Chukchi shelf at ~70–72°N (pink field in Figure 1). This zone corresponds to climatological average spring–summer sea-ice concentrations of 70–80% and is likely related to a stable ice edge occurring in early summer. A sharp southward decrease in IP25 values co-occurs with maximal concentrations of dinosterol, a biomarker of open-water conditions often proximal to sea ice. A comparable IP25 decrease north of the peak zone, toward more lasting sea-ice cover, has been documented west of the Chukchi Sea, but no samples were available to characterize this transition at the Alaskan margin. Yet further north, low IP25 values were found in the Canada Basin of the Arctic Ocean along with low concentrations of both brassicasterol and dinosterol, consistent with a lasting to permanent ice cover (Xiao et al., 2015a).
Materials and methods
Sampling and age constraints
Sediment cores HLY0501-05TC/JPC and -08TC/JPC (trigger or jumbo piston cores), hereafter referred to as 5JPC and 8JPC, were raised from the northeastern (Alaskan) margin of the Chukchi shelf in 2005 from the USCGC Healy (Figure 1; Darby et al., 2005). Multicore 8MC raised nearby 8JPC was also used in this study. Core 8JPC is sited in the eastern part of the shelf, on the flank of the Barrow Canyon, a major conduit for water and sediment transport to the Alaska Beaufort margin and Canada Basin. Based on its relatively shallow water depth (90 m), the core site was exposed during the last glaciation and inundated during the postglacial transgression (e.g. Keigwin et al., 2006). In contrast, core 5JPC was raised from the continental slope at 415 m depth, where sediment deposition was not interrupted by sea-level changes.
Various stratigraphic, sedimentological, and geochemical data on these and nearby cores (Figure 1) have been reported in a number of papers (e.g. Darby et al., 2009, 2012; Farmer et al., 2011; Faux et al., 2011; Lisé-Pronovost et al., 2009; McKay et al., 2008). JPC to TC offsets because of overpenetration, which is especially common for JPC, were estimated from the comparison of various proxies, primarily measured continuously, such as bulk density, magnetic susceptibility, and diffuse spectral reflection (Darby et al., 2009). In addition, the 8MC to TC offset was estimated from data in this study. Sediments in most of 8JPC and ~13 m in 5JPC consist of bioturbated clayey silts indicative of marine environments (Darby et al., 2009), with a more sandy composition near the 8JPC bottom, possibly related to shallow-water erosion and redeposition during shelf flooding. In 5JPC, the homogeneous, fine-grained marine unit is underlain by a more complex lithostratigraphy with laminations and coarse ice-rafted debris indicative of glaciomarine environments affected by glacial or deglacial processes (McKay et al., 2008; Polyak et al., 2009). Organic carbon (OC) content shows a steep increase from below 1% in the upper part of the deglacial unit to nearly 1.5% in the lower Holocene and then a slight, gradual increase toward the core top with only minor variability (Figure 2; Curry, 2009; Faux et al., 2011; McKay et al., 2008). A previous study of organic markers in 5JPC indicates predominantly marine sources of organic matter in the Holocene with lower but continual contributions from terrestrial sources (Faux et al., 2011), consistent with data from surficial sediments at the Chukchi margin (Belicka et al., 2004).

Distribution of IP25, sterols, and total organic carbon in cores 5TC/JPC and 8MC/TC/JPC versus composite core depth. OC data are from McKay et al. (2008) and Curry (2009) for 5JPC and 8JPC, respectively. IP25 and sterol data are presented in microgram per cubic centimeter sediment. 14C ages (cal. ka BP) are shown at the core depth axes. Gray vertical line indicates the bottom of the marine unit. Sampling levels in JPCs are shown by dots; no symbols are used for more densely sampled MC/TCs and for OC data. Scale bars for some proxies may slightly differ between the cores for illustrative purposes.
Age constraints were provided by 6 and 10 accelerator mass spectrometry (AMS) 14C ages of mollusk shells from cores 5JPC and 8TC/JPC, respectively (Figure 2; Supplement 1, available online; Darby et al., 2009), with concurrent age controls from paleomagnetic data (Lisé-Pronovost et al., 2009) and 210Pb in the upper part of 5TC (McKay et al., 2008). 14C ages were converted to calendar ages using the CALIB7.0 program and Marine13 dataset (Reimer et al., 2013). Local reservoir corrections (ΔR) were taken as 500 years for 8JPC washed by surface waters with a strong Pacific component and 0 years for 5JPC washed by subsurface Atlantic waters below 200 m (Darby et al., 2012; McNeely et al., 2006). We note that the actual ΔR values in the study area may have varied during the Holocene, especially at shallower sites, because of changes in the sea level and hydrographic structure.
The age model, except for 5TC, was constructed by linear interpolation between the 14C datings, which fall within the interval of ca. 2.4–7.7 cal. ka, as well as the assumed modern age of the 5TC and 8MC tops. The best 14C-based age control covers the interval of 4.1–7.3 ka in 8JPC and 4.7–6.9 ka in 5JPC. Ages below the dated ranges were extrapolated to the bottom of the Holocene marine unit, but in 8JPC, this estimate was complicated by an age inversion in the lower part of the core (Figure 2), reflected in two age models below this level (solid and dashed lines in Figure 3). The resulting age of the marine unit bottom differs between the two cores by 1 to nearly 2 ka (depending on the choice of the lowermost dating used in 8JPC), which could be an artifact of extrapolation and/or a true asynchroneity related to different water depths. Age constraints for glacial or deglacial sediments in 5TC were estimated using lithological tie points from the regional stratigraphic context, such as the bottom of iceberg-rafted deposits and the onset of sediment transport from the Bering Strait (Polyak et al., 2009). The distribution of linear sedimentation rates in the Holocene marine unit shows maximal values in both cores around 5–6 ka, with especially high rates attained in 8JPC. The age model for 5TC was estimated from 210Pb data measured in the upper 15 cm (McKay et al., 2008). Although we cannot guarantee a robust extrapolation of sedimentation rates from the analyzed interval to the entire ~2.5-m-long 5TC, we use this approach as no 14C age has been obtained from this core. Bioturbation in the Arctic Ocean and specifically in the study area has been estimated as modest, for example, 2–3 cm at water depths >150 m and somewhat higher at shallower depths (Clough et al., 1997; Pirtle-Levy et al., 2009), and is, therefore, unlikely to have a strong effect on the age model, especially in 5TC/JPC.
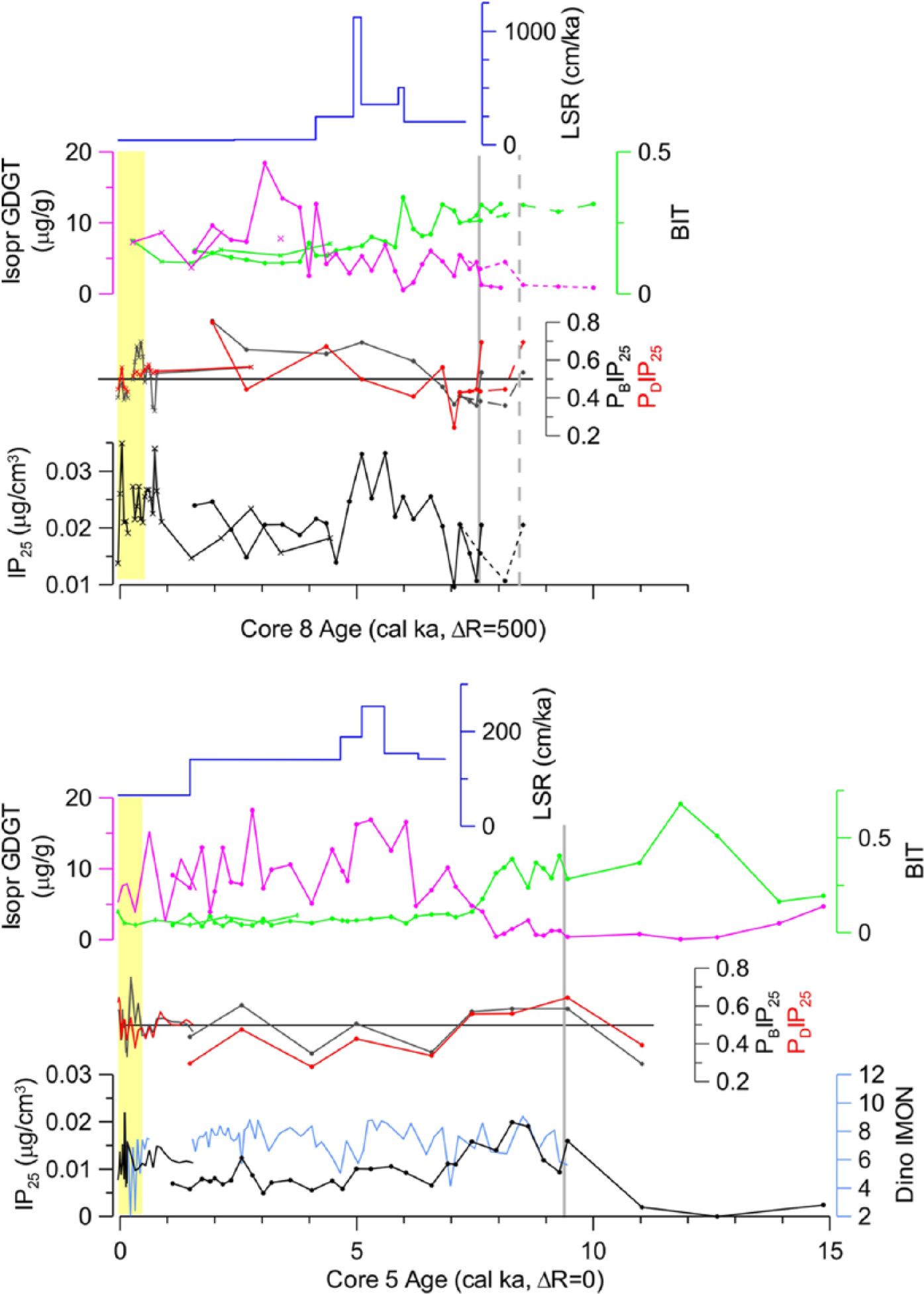
Distribution of IP25 and related proxies in cores 5TC/JPC and 8MC/TC/JPC versus age: IP25, PIP25 indices, isoprenoid GDGT concentrations, and linear sedimentation rates (LSR). Samples are indicated by dots in JPCs and crosses in 8MC/TC; no symbols are used for densely sampled 5TC. Yellow field on the left highlights subsurface interval with elevated sterol concentrations. PIP25 indices are calculated using separate balance factors for <0.4- to 0.5-ka and older intervals. A dinocyst-based reconstruction of sea-ice duration (months/year) for 5TC/JPC (Farmer et al., 2011; ages recalibrated) is also shown. Gray vertical line shows the bottom of marine unit. Dashed lines near the JPC8 bottom shows the alternative age model because of 14C inversion.
A total of 62 and 49 samples were collected for IP25 analysis from cores 5TC/JPC and 8MC/TC/JPC, respectively; 37 and 32 of them were also analyzed for marine or terrestrial sterols. Isoprenoid GDGT data cover 47 and 34 samples from the same cores as part of a broader GDGT study. Samples were mostly taken from the Holocene marine sediments at intervals intended to provide a multicentury-scale resolution (~200–400 years per sample at most of the record length). The uppermost part of the cores (~0.5 m of composite core depth) was sampled for IP25 at higher resolution, up to 10–20 years per sample in 5TC, in order to characterize the most recent record in more detail. In addition, several samples from the lower part of 5JPC span the deglacial sedimentary sequence. Samples were stored in a refrigerator following collection and then subsampled and freeze-dried for further processing.
Biomarker analysis
IP25 and sterols were analyzed following methods described previously (Belt et al., 2012; Brown et al., 2011). Briefly, 9-octylheptadec-8-ene (9-OHD, 10 µL; 10 µg/mL) and 5α-androstan-3β-ol (10 µL; 10 µg/mL) were added to ca. 1–2 g of each freeze-dried sediment sample prior to extraction to permit quantification of IP25 and sterols, respectively. Samples were then extracted using dichloromethane or methanol (3 × 3 mL; 2:1 v/v; ultrasonication; 15 min), centrifuged (2500 r/min; 1 min), and dried (N2). The resulting dried total organic extracts (TOEs) were dissolved in hexane (ca. 1 mL) and purified using column chromatography (silica) with IP25 (hexane; 6 mL) and sterols (20:80 methylacetate–hexane; 6 mL) collected as two single fractions. Analysis of individual fractions was carried out using gas chromatography–mass spectrometry (GC-MS) with operating conditions as described previously (Belt et al., 2012). Sterols were derivatized (N,O-bis(trimethylsilyl)trifluoroacetamide (BSTFA); 50 µL; 70°C; 1 h) prior to analysis by GC-MS. Mass spectrometric analysis was carried out in total ion current (TIC) and single-ion monitoring (SIM) modes. Individual lipids were identified on the basis of their characteristic GC retention indices and mass spectra obtained from standards. Quantification of IP25 was achieved by dividing its integrated GC-MS peak area by that of the internal standard (9-OHD) in SIM mode (both m/z = 350) and normalizing this ratio using an instrumental response factor (obtained from laboratory standards of each analyte) and the mass of sediment (Belt et al., 2012). Analytical reproducibility (5%, n = 4) was monitored using homogenized sediment material with a known concentration of IP25, similar to those found for the sediments under study (Belt et al., 2012). Values were further checked through analysis of this homogenized sediment for every 8–12 sediment samples extracted, as per the recommendation of Belt et al. (2012). Sterol concentrations were obtained by comparison of their respective peak areas in SIM mode (brassicasterol, m/z = 470; campesterol, m/z = 382; dinosterol, m/z = 500; and β-sitosterol, m/z = 396) with those of the internal standard (m/z = 333) and normalized as per IP25. Since we did not have a laboratory standard of dinosterol, we determined its instrumental response factor by analysis of sediment with known concentration (Faux et al., 2011). PBIP25 and PDIP25 values were determined from IP25 and brassicasterol or dinosterol data according to the method of Müller et al. (2011). Biomarker concentrations were also combined with dry bulk densities and expressed in microgram per cubic centimeter to account for changes in sediment density (Belt et al., 2012). Because of a relative sparsity and uneven downcore distribution of age control points, we prefer not to express IP25 data as annual fluxes as done in some studies (Belt et al., 2010; Müller et al., 2009; Vare et al., 2009).
GDGTs were analyzed as described in Park et al. (2014), using the recommended methodical guidelines of Hopmans et al. (2000), Huguet et al. (2006), and Schouten et al. (2007).
Results
IP25 concentrations in core 5TC/JPC range from nearly 0 to 0.025 µg/cm3 of dry sediment (Figures 2 and 3). Despite overall low values, all samples in the deglacial or Holocene sediments have nonzero IP25 concentrations. One sample analyzed from the bottom-most unit, presumably deposited during the glacial maximum (beyond the stratigraphic range in Figure 3), did not show detectable levels of IP25. In most of the deglacial section until estimated 11 ka, IP25 values stay very low, then increase to ~0.02 µg/cm3 toward the bottom of the marine unit, and gradually decrease from ca. 8.5 to 6.5 ka. In the remainder of the middle to late-Holocene, IP25 levels remain low, around 0.01 µg/cm3 but become somewhat higher at ca. 1 ka and, especially, within the last two centuries.
In core 8MC/TC/JPC, IP25 concentrations are similar to those in 5JPC, with overall slightly higher values of ~0.01–0.035 µg/cm3, but the downcore distribution differs considerably (Figures 2 and 3). In the lower to middle Holocene, maximum IP25 values occur in the interval ca. 5–7 ka, with the highest levels attained between 5 and 6 ka, and overall lower values below and above this interval. In the late-Holocene, IP25 values in 8JPC are still higher than in 5JPC and especially high, although variable, in the last millennium. We note that the analyzed 8JPC record does not extend below the marine Holocene unit, where sedimentation was probably predominated by shallow-water processes.
Downcore distribution of dinosterol is generally similar to that of IP25 in both cores (Figure 2). Brassicasterol shows a similar pattern between the cores, with a distribution alike IP25 in 5JPC but not in 8JPC. It is also distributed similar to terrestrial sterols, with a fivefold increase in both cores at the subsurface interval corresponding to the last four or five centuries. To account for this increase in sterol concentrations, the balance factor for the PIP25 indices derived from IP25 and brassicasterol or dinosterol data was calculated for the subsurface interval and the rest of the stratigraphy separately (see more discussion below).
Concentrations of isoprenoid GDGT in both 5JPC and 8JPC have low to moderate values under ~18 µg/g (Figure 3). In the late deglacial interval to the early Holocene (until ca. 9 ka), concentrations are low in both cores and then increase markedly in 5JPC to a maximum around 5–6 ka. Later, in the Holocene, isoprenoid GDGT values are overall high but variable. Similar GDGT distribution characterizes core ARA-03B GC01 further west on the Chukchi shelf (Figure 1; Park, personal communication, 2015). In contrast, in 8JPC, isoprenoid GDGT concentrations exhibit a distinct maximum around 3 ka. The branched and isoprenoid tetraether (BIT) index is high in deglacial sediments in 5JPC and decreases in both cores to very low values after ca. 8 ka in 5JPC and 2 ka later in 8JPC (Figure 3).
Discussion
Background interpretation of the observed biomarker distributions
Concentrations of IP25 in the Holocene record of both cores under study, when normalized to OC, constitute mostly ~1–2 µg/g OC, an order of magnitude lower than peak values identified in surface sediments of the Chukchi shelf further south (Stoynova et al., 2013) but comparable with values found north of the study area (Xiao et al., 2015a). It must be noted, however, that the IP25 data reported by Stoynova et al. (2013) appear to have an offset from the other Arctic datasets, possibly attributed to interlaboratory calibration issues (Xiao et al., 2015a), so the difference of our IP25 values from those of Stoynova et al. (2013) may not be that large. In any case, although low, these values along with attendant concentrations of brassicasterol and dinosterol equivalent mostly to ~5–20 µg/g OC (higher brassicasterol in the youngest sediment) fall within the range deemed useful for characterizing Arctic sea-ice conditions (Müller et al., 2011; Xiao et al., 2015a). Together with PBIP25 and PDIP25 indices averaging between ~0.3 and 0.6 at most of the core length in 5JPC and slightly higher in 8JPC, the observed IP25 values correspond to the conditions of marginal ice zone to extended ice cover common for the northern edge of the Arctic continental margin (Xiao et al., 2015a). We note that even lower values of IP25 have also been used for reconstructing paleo-sea-ice conditions in the Arctic Ocean (Xiao et al., 2015b).
Slightly higher concentrations of IP25 and PIP25 indices at most of the core length in 8JPC might indicate that this site was overall closer to the sea-ice margin than 5JPC. However, biomarker concentrations can have site-specific differences, as exemplified by the CAA cores that have very different average IP25 values despite coherent temporal patterns (Belt et al., 2010). Therefore, we base our interpretation primarily on downcore changes in IP25, which are considerably different between JPC5 and JPC8 at most of the record (Figures 2 and 3). This pattern is shared by dinosterol to some extent, whereas brassicasterol and terrestrial sterols have a more comparable distribution in both cores, with a concerted steep increase followed by a large variability in the upper part of the record. Cross-plotting of brassicasterol versus terrigenous sterol concentrations (Figure 4) confirms their close relationship, especially evident in the upper part of the cores (last four or five centuries), where concentrations are overall higher and vary within a broad range. Dinosterol and IP25 also show a relationship with brassicasterol in this interval but without a corresponding increase in their concentrations, which makes this youngest record notably different from the rest of the stratigraphy.
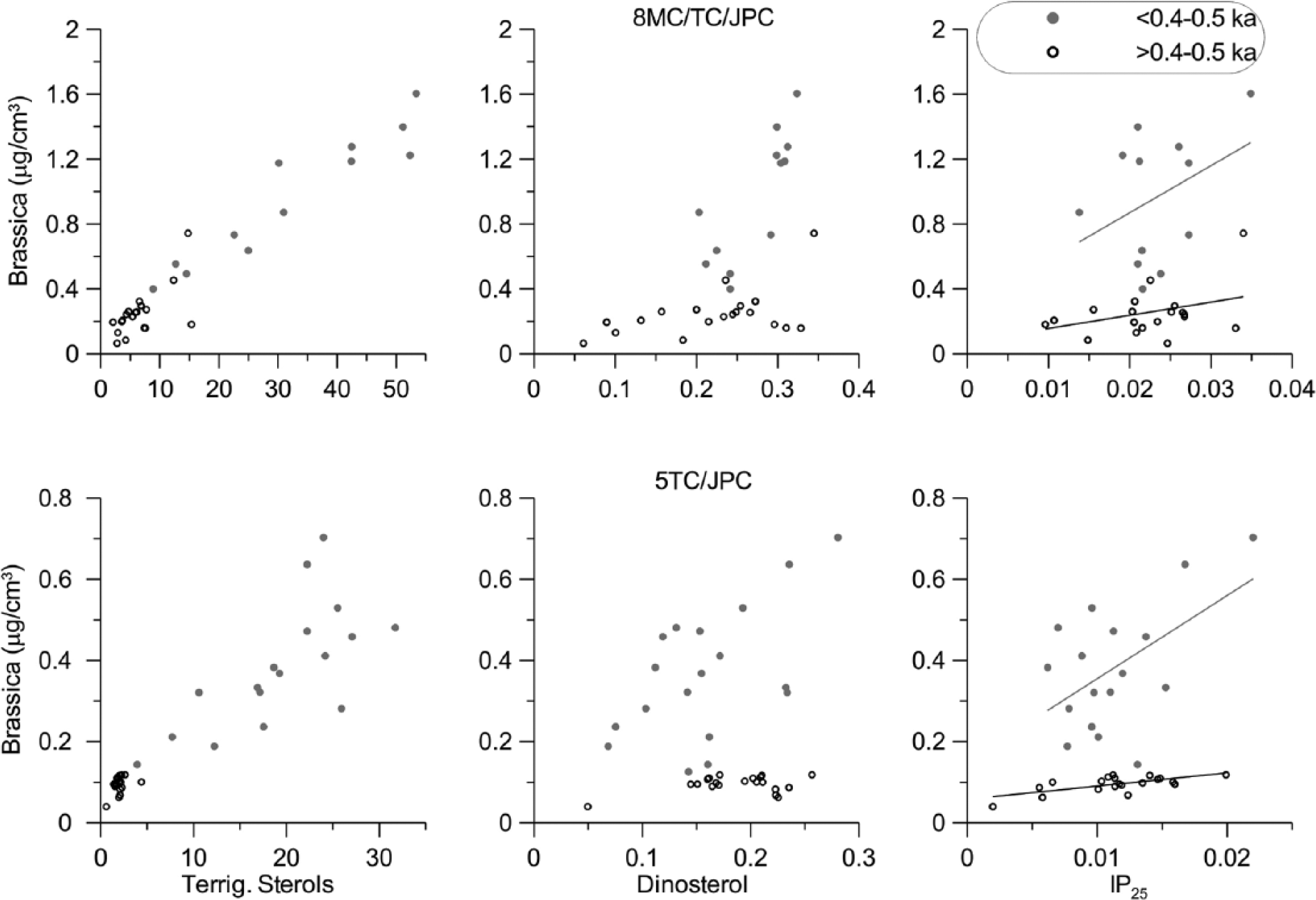
Distribution of brassicasterol versus terrestrial sterols, dinosterol, and IP25 in cores under study. All data are presented in microgram per cubic centimeter sediment. Different symbols indicate two distinctly different patterns: in the subsurface interval (younger than ~0.4–0.5 ka) and in the rest of the record.
The close relationship of markers from different sources is unusual and indicates an impact from an external factor such as a dilution by mineral sediment matrix or a co-delivery to the core sites by the same transportation mechanism. As mineral dilution in the upper part of the cores is unlikely, because of a lack of co-variation of the sterol data with the total OC content (Figure 2), we infer that delivery of both marine and terrestrial components by ocean currents provides a likely explanation. In support of this, we note that the study sites are affected by cross-shelf currents originating from the North Pacific, notably by the Alaskan Coastal Current that has an especially strong influence on 8JPC (Figure 1). The Alaskan Coastal Current carries fines from the coastal areas of the eastern Bering Sea encompassing sites with high marine organic production and the estuarine areas including the large Yukon River and can, therefore, provide an effective transport mechanism for both marine and terrestrial organic markers. Indeed, the relationship between brassicasterol and IP25 in the broad regional context (Figure 5) shows that data from the younger record in cores under study fall between the Chukchi shelf and the North Pacific signature, thus indicating the likelihood of long-distance sediment transport. In comparison, data from the older record show more affinity to the Chukchi shelf sediments, suggesting the prevalence of more local processes during most of the Holocene. Elevated concentrations of brassicasterol and terrestrial sterols in the subsurface record could also be controlled by diagenetic transformation, but this, alone, cannot adequately explain the observed high variability in biomarker values (more discussion in the final section below). Regardless of the exact mechanism, we infer that interpretation of the youngest record (last four or five centuries) is likely to be biased by long-distance sedimentary inputs, possibly in combination with some diagenetic changes. In contrast, the rest of the stratigraphy under study may be more suitable for reconstruction of local sea-ice conditions. We note that the range of our Holocene data could be even closer to the recent Chukchi Sea IP25 values of Stoynova et al. (2013), because of a potential interlaboratory data offset, as suggested by Xiao et al. (2015a).
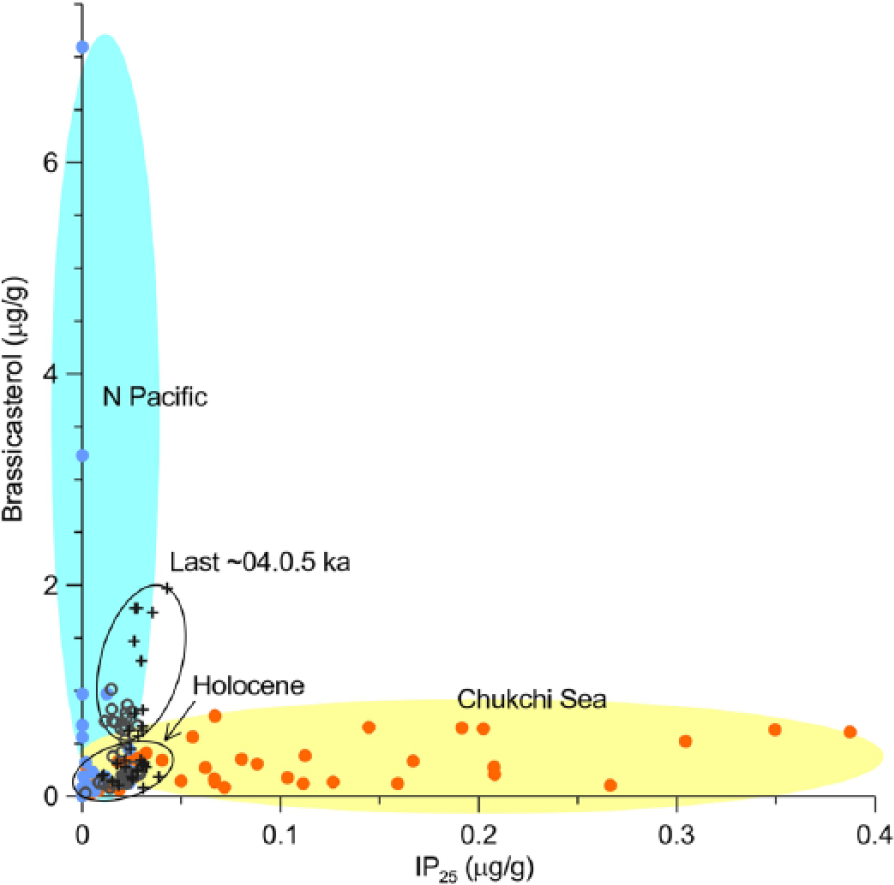
Comparison of the distribution of brassicasterol versus IP25 in cores under study and surface sediments of the Chukchi Sea and North Pacific (data from Stoynova et al., 2013). All data are presented in microgram per gram sediment for compatibility. Gray circles: 5JPC; plus signs: 8JPC; orange and blue dots: surface sediment samples from the Chukchi Sea and North Pacific, respectively.
A dual application of biomarker and transfer functions based on, for example, dinocysts, which consistently occur in Arctic Holocene sediments, offers a potentially promising approach to sea-ice reconstructions. However, a comparison between IP25 and dinocyst-based reconstructions of sea-ice conditions in 5JPC shows a mixed picture of consistent patterns at some intervals but a considerable divergence at others (Figure 3). Overall, the dinocyst data show more variability compared with the biomarker record. There is currently no consensus on why outcomes derived from these two sea-ice proxies should be different (Belt and Müller, 2013). Apparent inconsistencies in temporal sea-ice profiles may better reflect differences between the precise signatory natures of the individual proxies themselves, rather than anomalies. Thus, peaks in IP25 are believed to reflect primarily spring–summer sea-ice edge (Belt and Müller, 2013; Müller et al., 2011), whereas dinocyst reconstructions are related to a long-term, annual perspective (i.e. months/year cover; e.g. de Vernal et al., 2005, 2008). The interrelationships that may exist between the two approaches are, thus, in need of further attention. Nevertheless, what is clear from the application of both approaches to the Alaskan margin sites is the pervasive occurrence of sea-ice cover throughout the Holocene.
Long-term Holocene record
Interpretation of extremely low IP25 concentrations in the deglacial record, exemplified by the lower part of 5JPC (Figures 2 and 3), is not straightforward. One possibility is that low IP25 values along with low brassicasterol and dinosterol concentrations identified for the top of this interval reflect permanent sea-ice cover such as in the Canada Basin in recent conditions (Xiao et al., 2015a). The paleogeographic setting at the Chukchi margin during deglaciation was also potentially amenable for sea-ice buildup because of meltwater inputs from the retreating Laurentide Ice Sheet and a not fully open Bering Strait. On the other hand, the deglacial environment was affected by strong iceberg discharge, as indicated by the high content of coarse debris including rocks with the Laurentide provenance (Darby et al., 2001; McKay et al., 2008; Polyak et al., 2009). The imported nature of the deglacial sediment is further corroborated by a high content of branched GDGT (BIT index; Figure 3) that is indicative of enhanced delivery of terrestrial (soil) organic material (Hopmans et al., 2004). Numerous icebergs likely disrupt the ‘normal’ development of sea-ice cover and the formation of ice-related biotic assemblages, which could result in very low IP25 and spurious PIP25 values. In addition, sedimentation overwhelmed by terrigenous material delivered by icebergs and meltwater further reduces IP25 concentrations. For a comparison, Alonso-García et al. (2013) showed that high content of coarse debris resulting from rapid discharges of icebergs coincided with relatively low IP25 concentrations in sediment from the East Greenland Shelf during the second half of the 19th century despite relatively high ice conditions overall.
Regardless of the interpretation of deglacial sea-ice conditions, a conspicuous increase in IP25, along with dinosterol, with a transition to the marine Holocene unit (Figures 2 and 3) likely indicates an increasing proximity of the sea-ice margin. An accompanying increase in brassicasterol, however, should be considered with caution as a similar pattern in 8JPC is not coupled with changes in IP25. The brassicasterol profile in both cores is comparable with that of terrestrial plant sterols, with a slight offset in the position of peak values in the lower Holocene, and could be thus related to an increasing long-distance advection via the widening Bering Strait. A further decrease in sterol concentrations may indicate a subsided advection to the study sites, possibly because of circulation changes, combined with reduced local phytoplankton production.
A notable feature in the observed IP25 distribution in the marine unit is the difference between the two cores, with a broad maximum in the early Holocene (from estimated 10–11 ka to ca. 6.5–7 ka) in 5JPC versus the delayed maximum at 5–7 ka in 8JPC that corresponds to especially high IP25 fluxes considering high sedimentation rates during this time interval exceeding 1 m/ka (1 mm/a) around 5 ka (Figure 3). Changes in PIP25 indices generally follow this pattern, especially consistent for PDIP25 in 5JPC. Provided these changes represent mostly local conditions, the decrease in IP25 and PIP25 after ca. 8 ka in 5JPC and ca. 5 ka in 8JPC may indicate an overall reduction in the duration of ice cover. A comparison with GDGT distribution shows that concentrations of isoprenoid GDGT in both cores increased after the decline of IP25, peaking at ca. 5–6 ka in 5JPC and ca. 3 ka in 8JPC. This delay of isoprenoid GDGT relative to IP25 is consistent with the inferred negative effect of sea ice on local GDGT production (Park et al., 2014), while the temporal shift between isoprenoid GDGT maxima underscores the asynchronicity in sea-ice conditions across the Chukchi margin as expressed in IP25 records.
Sedimentary factors may have a considerable effect on the distribution of organic matter and its constituents in sediments at the Chukchi margin by controlling transportation, redistribution, and deposition of fines (Darby et al., 2009). Faux et al. (2011) have concluded that Holocene sediments in 5JPC were mostly well mixed prior to deposition. However, sedimentary factors are unlikely to play a significant role in the observed asynchronous pattern of biomarker distribution between 5JPC and 8JPC as maximal sedimentation rates, and thus, maximal resuspension of fines on the Chukchi shelf occurs at about the same time (ca. 5–6 ka) in both cores (Figure 3). A more plausible explanation is related to circulation changes, such as the distribution of Bering Strait Inflow water between different branches and strength of the Beaufort Gyre (Figure 6).
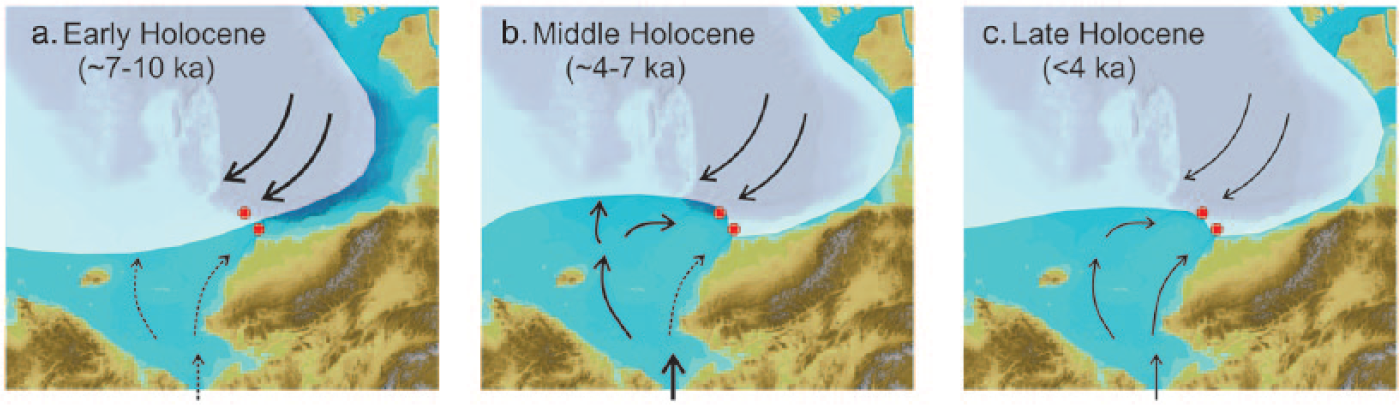
Conceptual scheme of circulation and sea-ice extent at the Chukchi-Alaskan margin during the early and middle Holocene (roughly 7–10 and 4–7 ka, respectively) in comparison with recent conditions (see Figure 1 for more details). Arrows show Bering Strait Inflow; thicker arrows indicate higher current strength; yellow line indicates (a and b) inferred patterns of Holocene summer ice margin and (c) observed late 20th-century ice extent; red dots show location of cores under study.
The early Holocene pattern of biomarker distribution at the Chukchi margin, exemplified primarily by 5JPC, is more difficult to interpret because of nonanalog conditions related to an incompletely open Bering Strait and potentially lingering meltwater. Nevertheless, a pronounced peak of IP25 and relatively high PIP25 values in 5JPC indicate an overall significant presence of sea ice in this area, consistent with an earlier conclusion based on dinocyst assemblages (de Vernal et al., 2008, 2013). This conclusion contrasts an inferred early Holocene ice retreat in other parts of the Arctic Ocean periphery, notably north of Greenland and the Canadian Arctic (Dyke and Savelle, 2001; Funder et al., 2011). This contrast is illustrated further by a comparison of IP25 records from the Chukchi margin and the straits of the Canadian Arctic Archipelago (CAA; Figure 7; Belt et al., 2010; Vare et al., 2009). The early Holocene IP25 maximum in 5JPC has no counterpart in any of the CAA cores, while for the easternmost location (core 3), this interval has minimal IP25 values, interpreted to represent the lowest sea-ice occurrence (Vare et al., 2009). A possible reason for anomalously extended ice cover at the Chukchi-Alaskan margin in the lower Holocene might be related to insufficient advection of warm waters via the Bering Strait (Figure 6a) and intensified import of ice by the Beaufort Gyre. Indeed, mineral provenance records indicate increased transport of sediment by way of the Beaufort Gyre at that time (Yamamoto, personal communication, 2015), possibly because of its mobility provided by melting sea-ice at the margins under overall warmer climatic conditions, similar to the intensified Beaufort Gyre movement in recent decades (Shimada et al., 2006).
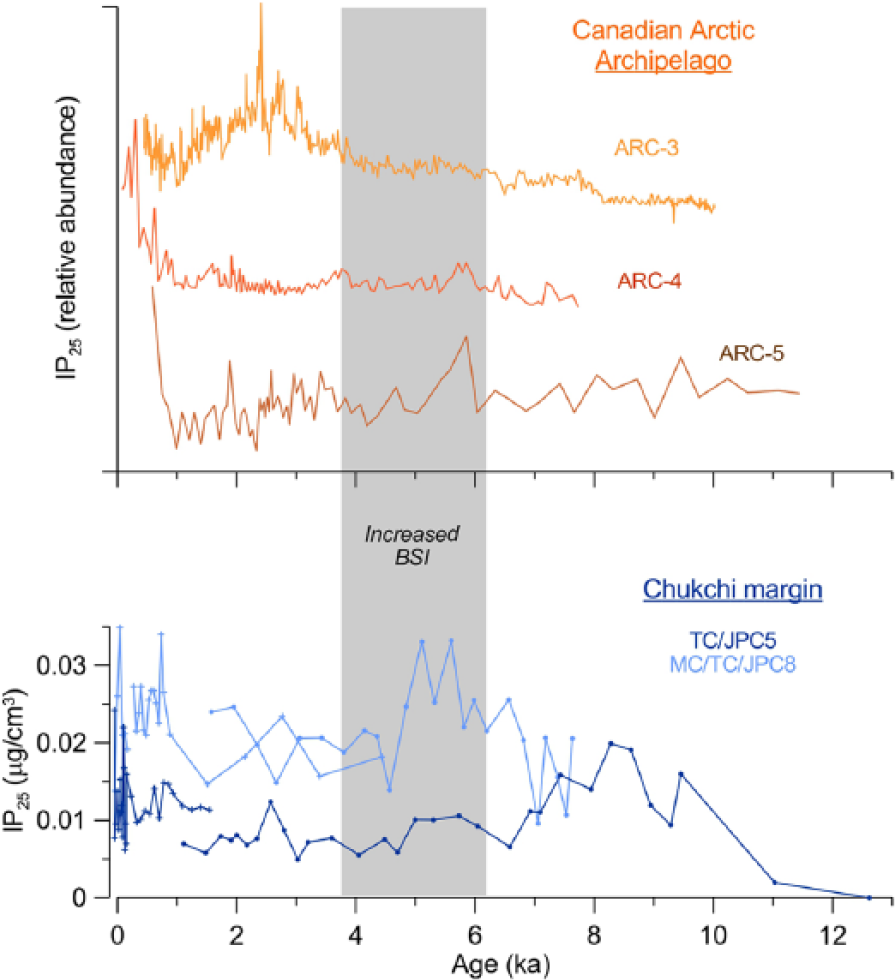
Comparison of IP25 data from the Chukchi-Alaskan margin (this study) and the Canadian Arctic Archipelago (CAA) straits (see Figure 1 for site location; Belt et al., 2010; Vare et al., 2009). The CAA IP25 datasets are shown in relative abundance because of vastly different concentrations, with an offset for illustrative purposes. Gray bar marks the period of increased Bering Strait Inflow inferred from a sedimentary proxy record (Ortiz et al., 2009, 2012).
The delayed IP25 peak in 8JPC occurred at a time when the Bering Strait Inflow reached its maximum in the middle Holocene (between ca. 6 and 4 ka) as indicated by a sediment provenance proxy record proximal to 5JPC (Ortiz et al., 2009) and confirmed, further, by data from a more westward core GC01 (Figure 1; Yamamoto, personal communication). This maximal inflow is also reflected by peak sedimentation rates at the Chukchi margin (Figure 3) because of intense resuspension and deposition of fine sediment (Darby et al., 2009). High Bering Strait Inflow volumes favor westward diversion of more Pacific water at the expense of diminished Alaskan Coastal Current contribution (Winsor and Chapman, 2004), which is consistent with our inference of sea-ice retreat at the 5JPC site, but higher ice coverage in 8JPC further east (Figure 6b). A comparison with the IP25 records from the CAA shows that sea-ice expansion by way of the Alaskan Coastal Current may have affected the proximal part of the CAA straits (i.e. cores 4 and 5) but not the eastern area exemplified by core 3 (Figures 1 and 7). After ca. 4 ka, the strength of the Bering Strait Inflow decreased, probably driven by changes in atmospheric circulation (Ortiz et al., 2009, 2012), resulting in a more equitable distribution of Pacific water between western and eastern branches and, thus, of sea-ice extent (Figure 6c). Based on higher IP25 values in 8JPC, ice conditions were probably more severe, overall, in the eastern area. It is also possible that summer sea-ice distribution in the northern Chukchi Sea was irregular, because of a relatively sluggish circulation: a pattern consistent with historical observations and related modeling (Spall, 2007). A prominent IP25 peak at ca. 2–2.5 ka in the eastern CAA core 3, possibly representing Neoglacial cooling (Figure 7; Vare et al., 2009), is absent both in the Chukchi and western CAA cores.
Last millennium
A notable pattern in IP25 and sterol data in the record corresponding to the last several centuries requires further discussion, especially considering the heightened relevance of this time interval for comparison with modern natural climatic conditions. Overall low and stable sterol values in most of the Holocene record show a pronounced increase in variability, along with concentrations of brassicasterol and terrestrial sterols, in the last four or five centuries, according to the existing age models based on 210Pb data in 5TC (McKay et al., 2008) and 14C interpolation in 8MC/TC (Figures 2 and 8). IP25 values in this interval exhibit a similar variability, without an increase relative to the peak values in the older Holocene record. As discussed above, the co-variation including the concerted rise in both marine (brassicasterol) and terrigenous (higher plants) sterols in the recent sediments may be related to diagenetic processes, long-distance co-delivery by currents, or a combination of these factors.
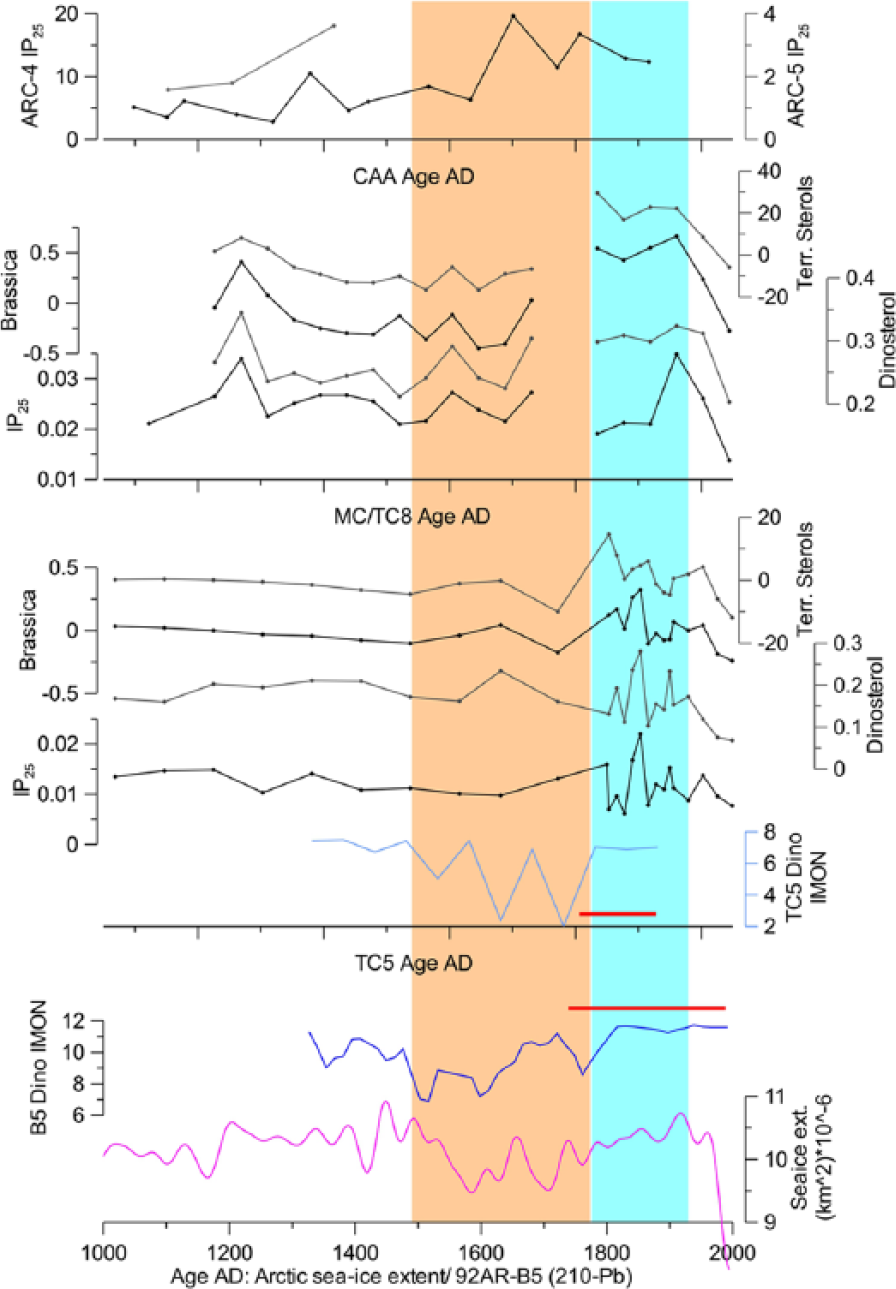
IP25 and sterol data for the last millennium (this study), compared with IP25 records from the CAA (Belt et al., 2010), dinocyst-based sea-ice proxy curves (5TC: Farmer et al., 2011; B5: de Vernal et al., 2008), and Arctic-wide reconstruction of sea-ice extent (Kinnard et al., 2011). Red horizontal bars show the range of 210Pb data for 5TC and B5 (de Vernal et al., 2008; McKay et al., 2008). Vertical pink and light blue fields indicate early and late ‘Little Ice Age’ in the Arctic–Pacific region. IP25 and sterol data are presented in microgram per cubic centimeter sediment. Brassicasterol and terrestrial sterol profiles are quadratically detrended. Scale bars for some proxies may slightly differ between the cores for illustrative purposes.
Diagenetic degradation of organic matter is expected in subsurface marine sediments, including the Arctic seas, and can affect all organic constituents, although likely at different rates (e.g. Belicka et al., 2004; Goňi et al., 2013; Haddad et al., 1992; Harvey et al., 1986). The apparent lower (or absent) diagenetic loss of dinosterol in our data could be related to a lower reactivity compared with other sterols, especially since the former does not contain a ring double bond found in all Δ5 sterols (e.g. brassicasterol), and which plays a key role in abiotic sterol degradation reactions including photo- and autoxidation (Christodoulou et al., 2009; Rontani et al., 2009). This inference is also consistent with data from subsurface records in the western Arctic indicating generally only subtle downcore diagenetic changes in biomarker distributions including dinosterol (Belicka et al., 2004). While diagenetic reactivity of different biomarkers requires further investigation, the high variability in the record under discussion cannot be explained by organic matter degradation alone; neither can it be attributed to the impact of burrowing organisms, which appears to affect only a few upper centimeters in sediments throughout the study region (Clough et al., 1997; Pirtle-Levy et al., 2009). This indicates that the observed variability in sterol and IP25 concentrations is likely related to changes in circulation, while an overall downcore decrease in concentrations of some biomarkers could be controlled by diagenetic losses. In order to obtain a clearer picture of the inferred current-related variability, the profiles of brassicasterol and terrestrial sterols have been quadratically detrended (Figure 8).
Interestingly, the sharp rise in terrestrial sterols is not accompanied by a comparable increase in branched GDGT (BIT index; Figure 3), which indicates a likely different provenance of imported terrestrial material. In the deglacial time and early Holocene, it was mostly represented by soil-derived organic matter, probably originating from the melting of Laurentide ice and/or the Mackenzie River. In contrast, in the last centuries, terrestrial material was related largely to higher plant debris that is not common for high-Arctic river load and is more likely transported from the Yukon River by way of the Alaskan Coastal Current. This provenance interpretation is consistent with biomarker distributions in modern sediments from the Chukchi-Alaskan margin and the Mackenzie area (Goňi et al., 2013).
Reconstructions of sea-ice cover duration based on dinocyst assemblages from 5TC and a box core B5 further north (Figure 1) indicate overall less ice during the late 15th to late 18th century AD, before a subsequent expansion of sea-ice cover in the late ‘Little Ice Age’ (LIA; Figure 8; de Vernal et al., 2008; Farmer et al., 2011). A generally similar pattern can be seen in several proxy records around the Arctic Ocean and in a Pan-Arctic millennial sea-ice synthesis based on these records, including B5 (Kinnard et al., 2011). While the Pan-Arctic record can be affected by multiple factors including the North Pacific and, especially, North Atlantic climatic variability (Kinnard et al., 2011), the Chukchi region has more affinity to the North Pacific atmospheric and oceanic circulation. In particular, the strength and position of the Aleutian Low pressure system largely controls the Bering Strait Inflow and, thus, sea-ice conditions in the Chukchi Sea (Danielson et al., 2014). While no long-term proxy record exists strictly for the Aleutian Low, several paleoclimatic studies from the Northwest Pacific region provide relevant information. A strong westerly Aleutian Low, which enhances Bering Strait Inflow, is consistently indicated by ice cores from southern Alaska (Fisher et al., 2008; Porter, 2013) and lake records from the Alaskan interior (Gonyo et al., 2012, and references therein) for the 16th–18th century, the time of reduced sea ice at the Chukchi margin according to dinocyst-based reconstructions (de Vernal et al., 2008; Farmer et al., 2011). This circulation setting, however, negatively affects the Alaskan Coastal Current that delivers warm water and sediment load to the northeastern Chukchi Sea and further downstream along the northern Alaskan coast (Winsor and Chapman, 2004). In contrast, the more easterly or weaker Aleutian Low in the late LIA favored more ice cover developed in the main Chukchi Sea but stronger Alaskan Coastal Current, which increased the delivery of sediment load to the northeastern area. This circulation history appears consistent with overall anomalously high and variable sterol and co-varying IP25 values in the record under study corresponding to the late LIA and relatively depressed values in the preceding three centuries (Figure 8). It is unclear to what extent IP25 values reflect local sea-ice conditions, but a strong covariation with sterols that were likely imported from a long distance suggests a mostly external source for IP25 as well.
The circulation history inferred for the last millennium, with principally two alternating regimes of the Bering Strait Inflow, appears similar to the interpretation of long-term changes in the lower and middle Holocene, as discussed above (Figure 6). However, many of the conditions earlier in the Holocene were not quite analogous to the more recent situation. These include a different configuration of the Bering Strait, a lingering Laurentide ice sheet that strongly affected the atmospheric circulation (e.g. Kaufman et al., 2004) and potentially other climatic and oceanographic factors. These nonanalog conditions may have caused the differences between patterns in the earlier record and in the last several centuries, such as an explicitly asynchronous distribution of IP25 between the study sites and mostly uniformly low brassicasterol and terrestrial sterol concentrations in the early to middle Holocene.
We speculate further that short-term variability, expressed especially in the uppermost part of a densely sampled 5TC (Figure 8), reflects major fluctuations in the Alaskan Coastal Current linked to a documented multidecadal variability in the Aleutian Low (e.g. Hetzinger et al., 2012), although this inference requires more detailed studies on high-resolution cores from the Chukchi-Alaskan margin. The reason for a contrast between a variable LIA and relatively monotonous earlier biomarker record, especially apparent in fiveTC (Figure 8), is also not clear from the existing data as the sampling points below the late LIA record are not that sparse to miss fluctuations in the data measured altogether. A more pronounced amplitude in sterol and IP25 variations in the late LIA could be accentuated by diagenetic degradation downcore that makes earlier variability less discernable, but the question remains whether this amplitude change also has a paleoclimatic significance. Anomalously high-amplitude variability in the late LIA is not apparent in related proxy records from other Arctic or subarctic regions, except for an IP25 dataset north of Iceland that does not, however, extend far beyond the last millennium (Massé et al., 2008). On the other hand, a millennial tree-ring record of the Pacific Decadal Oscillation, which is linked with the Aleutian Low history, suggests that a strong, persistent decadal to multidecadal variability is only characteristic for the last two centuries (MacDonald and Case, 2005).
A comparison with the CAA data shows a pronounced rise in IP25 in the early LIA in cores 4 and 5 but not in the eastern core 3 (Figures 7 and 8; Belt et al., 2010), indicating an extended sea-ice cover in the western CAA, possibly related to a restricted Alaskan Coastal Current. However, a comparison with core 3 may be inconclusive as it lacks sediment from the last five centuries according to the age model applied (Vare et al., 2009).
Conclusion
This study provides the first paleoclimatic application of IP25 and related biomarkers for evaluating the Holocene sea-ice history in the Chukchi-Alaskan region of the Arctic Ocean, which plays a critical role in Arctic–Pacific interactions and is currently experiencing a dramatic retreat of sea ice under present warming. The long-term record investigated in two sediment cores, mostly at multicentury time scale, indicates an overall persistent presence of sea ice throughout the Holocene, but with considerable spatial and temporal variability. The pre-Holocene (deglacial) record from the Chukchi margin has very low IP25 values, probably related to some combination of sea-ice and iceberg-bearing environments. An IP25 maximum around 8–9 ka in a core from the shelf break suggests expanded sea ice in the western Arctic Ocean despite the early Holocene warming, consistent with some earlier dinocyst-based reconstructions (de Vernal et al., 2008, 2013). In a core further southeast, peak IP25 values, presumably characterizing a relative proximity to sea-ice margin, were reached ~3 ka later. In both cores, isoprenoid GDGTs have maximum concentrations with an ~1- to 2-ka delay relative to IP25, probably signifying sea-ice retreat. The asynchronous development of sea ice at these sites may be related to circulation history such as an undeveloped Bering Strait Inflow and, possibly, a stronger Beaufort Gyre in the early Holocene and a higher, westward deflected Bering Strait Inflow in the middle Holocene.
The biomarker record for the last several centuries demonstrates a strong temporal variability along with overall elevated values of brassicasterol and terrestrial sterols. This anomalous increase in biomarker concentrations is interpreted in terms of long-distance import by currents combined with diagenetic transformations. Although probably not representing local sea-ice conditions, this record may have value for characterizing broader circulation patterns. In particular, we infer that high-amplitude variability in the late LIA, starting in the late 18th century, is related to the intensity of the Alaskan Coastal Current. This interval is preceded by three centuries of presumably diminished Alaskan Coastal Current, but overall increased Bering Strait Inflow resulting in reduced sea-ice cover according to dinocyst-based data (de Vernal et al., 2008; Farmer et al., 2011).
Results of this study offer further opportunities to investigate linkages between atmospheric and oceanic processes in the Arctic–Pacific region on various time scales. The data obtained also highlight a need to resolve differences between individual proxies, such as IP25 and dinocyst assemblages. Emphasis should, therefore, be put on a multiproxy approach to sediment cores, with high temporal resolution from locations representing key circulation and sea-ice features.
Footnotes
Acknowledgements
The authors thank two anonymous reviewers for insightful comments that helped to improve the manuscript.
Funding
This work was partially supported by the US National Science Foundation award PLR-1404370 to LP. STB and PC-S thank the University of Plymouth for financial support and to John Price for carrying out preliminary IP25 measurements. The GDGT study was supported by a grant-in-aid for Scientific Research (B) from the Japan Society for the Promotion of Science, no. 25287136, to MY.