
Abstract
Morphological analysis of plant remains from archaeological sites provides evidence regarding the domestication process of crop species, and the changing economic behaviors of humans during the foundation and intensification of agriculture. In contexts from the Neolithic and early state periods of Henan, China, morphometry of genus Setaria millet seeds is shown to provide data on production versus consumption contexts of archaeological deposits, in connection with site function and settlement hierarchy. Comparative morphometry of modern Setaria seeds sheds light on larger archaeobotanical issues, including problematizing the distinction between domesticated and wild/weedy seeds. Statistical analysis suggests that subdivision of archaeological millets below the genus level is less useful in some cases than consideration and comparison of genus-level populations.
Keywords
Introduction
In the emerging and expanding societies of Neolithic North China, agriculture played a vital role in consolidating power, and supporting increased sedentism and social complexity (Barton et al., 2008; Cohen, 2011; Lee et al., 2007; Liu et al., 2002–2004). These human–plant interactions are key to understanding the changing social behaviors that led to the emergence of state-level societies worldwide (D’Altroy and Hastorf, 2001; Gumerman, 1997; Nesbitt, 1995; Smith, 2006). How best to use crop remains to draw conclusions about economic behaviors, however, has been an ongoing challenge. A key element of such interpretations is the differentiation of features associated with production- or consumption-oriented activities, as patterns of production and consumption are expected to change during Neolithic and state formative periods. This study suggests morphological and statistical methods for determining whether crop remains from a single feature represent agricultural production (particularly threshing), or food consumption (in this case, storage).
The use of seed morphology and statistics allow us to address several pressing archaeobotanical issues, including the pitfalls of over-identification, the potential for seed remains in interpreting site function, and the genetic and phenotypical plasticity of crop seeds. Morphological changes have been shown to occur relatively late in the domestication process (Lee, 2012a; Lee et al., 2011; Zeder, 2015), and therefore we investigate the usefulness of generalization, regarding all seeds of a genus as a single statistical population. We demonstrate that this is mathematically sound and prevents the exclusion of seeds which do not appear domesticated, but may well have been cultivated. Using the genus Setaria, of which domesticated foxtail millet (Setaria italica ssp. italica) was a primary staple crop in North China throughout the target periods, we carry out extensive morphological measurement and statistical analysis on both modern and archaeological seeds to suggest protocols for differentiation between production- and consumption-oriented features within sites. Rather than obscure trends, we find that inclusion of all Setaria seeds in our statistical tests reveals crucial information for behavioral inference, and may be vital in overcoming barriers to collaboration among researchers using various identification schemas.
Archaeological background
Domestication is long and slow process, often requiring several millennia to fix alleles in plant and animal species in order to suit them to human exploitation (Allaby et al., 2008; Harris, 2007; Jones and Liu, 2009; Larson, 2011). East Asia is best known as the domestication center for rice, with sites in the Yangzi and Huai River regions showing exploitation of wild or early cultivated rice as part of a broad-spectrum diet in the early Neolithic (Yang et al., 2015), millennia of low-level cultivation (Chi and Hung, 2013; Fuller et al., 2009), and eventual domestication (Deng et al., 2015). This recent research on rice demonstrates that seed size did not increase in the early stages of domestication, but that selection by humans is visible in non-shattering spikelet bases, making the plant easier to harvest. While extensive non-seed plant remains are not always available from archaeological sites, these findings serve as an important reminder that genetic and morphological changes in target species may be observed hundreds or thousands of years after the advent of human intervention.
In North China, two species of millet were likely domesticated relatively quickly by semi-nomadic peoples in the early Holocene (Yang et al., 2012), and domesticated forms became widespread in around 8000 BP (Bettinger et al., 2010; Crawford, 2009; Schontz and Rether, 1999; Underhill, 1997). Cultivation spread quickly, and even the Yangzi valley provides evidence of millet being grown alongside rice as early as 5800 cal. BP (Nasu et al., 2007). Broomcorn (Panicum miliaceum) and foxtail millet and other dry-field crops, such as (Triticum aestivum) and soybean (Glycine max ssp. max), are more suited to the temperate northern climate, and archaeology in Henan and other regions has shown that Neolithic cultures in North China relied heavily on dry crops rather than rice (Barton et al., 2008; Crawford et al., 2005; Lee et al., 2007; Zhao, 2011). Millets are adapted to the arid environment of the North, and are fast-growing and easily cultivated (Baker, 2003; Fogg, 1983; Lu, 1998). While long thought to prefer warmer and wetter environments (Liu et al., 2009), foxtail millet has recently been shown to be the more cold-tolerant of the two, making it better suited to marginal agricultural areas such as the Tibetan plateau (Guedes, 2015). Foxtail millet’s use as a staple crop in central China throughout the Neolithic makes it an ideal lens on the nature of social changes occurring in the Yellow River valley at this time.
Genetic distinction among Setaria sp. is subtle. Populations of S. italica and S. viridis from the same region sometimes show less genetic distance than is observed between populations of S. italica from different regions (Doust et al., 2009). Only one allele of domesticated S. italica differs from wild S. viridis among 28 loci on Setaria genomes analyzed by Wang et al. (1995). Also, S. italica still retains a high proportion of genetic diversity (45%) from S. viridis, compared with other crops, notably rice (20%) (Shi et al., 2008; Wang et al., 2010). This leads to the continuing agricultural problem of preventing widespread S. viridis populations from interfering with S. italica crops (Rao et al., 1987), an issue we can assume also faced Neolithic and historic farmers.
The distinction between wild and domesticated seeds in archaeobotany is an indispensible tool for detecting trends of cultivation, domestication, and agriculture, but the genetic similarity and frequent admixture between S. viridis and S. italica complicate this issue. Additionally, as pointed out by Song et al. (2013), immature seeds of S. italica can be mistaken for wild species. Based on ethnography and experimentation, Song et al. note the potential of immature grains in differentiating depositional contexts associated with threshing and winnowing, in which many smaller seeds are removed from crop yields, and provide guidelines for identifying such seeds. The researchers call for stricter maintenance of distinct identification categories for the purpose of comparison among scholars. The identification of as many seeds as possible to the species level is the goal of any archaeobotanist; however, many archaeological seeds which display clear Setaria traits (particularly a distinctively shaped embryo running 2/3 to 5/6 of the seed length) simply do not strongly resemble modern S. italica ssp. italica or S. italica ssp. viridis comparative specimens, mature or otherwise. Given our understanding of Setaria genetics and frequent co-occurrence of domestic and wild plants in agricultural settings, these seeds constitute evidence of the same behaviors as their more clearly defined counterparts, and should be studied in the same way. We investigate this issue by testing the usefulness of genus-level analysis in Setaria, addressing the need for comparison and cooperation while eliminating much of the concern over identification bias between researchers.
Archaeological seeds in this study come from the valley of the Yiluo River, a tributary of the Yellow River, which contains sites that provide the earliest evidence of state-level societies in China (Figure 1). Researchers involved with the Yiluo Project have provided multiple perspectives on phases from the early Neolithic through the beginning of the historical period (Lee, 2004; Liu et al., 2002–2004; Rosen, 2007), and an agricultural trajectory with a detailed sequence from direct dating on crops is provided by Lee et al. (2007). The major Neolithic phases in this region are the Peiligang or early Neolithic (8000–7000 cal. BP), the Yangshao or middle Neolithic (7000–5000 cal. BP), and the Longshan or late Neolithic (5000–4000 cal. BP). The state period begins with the Erlitou period (3900–3500 cal. BP), named after the site of Erlitou which functioned as an urban center, and which coincides with the beginning of bronze production in the region (Lee, 2002). The early phase of the Shang dynastic period is known as the Erligang (3600–3300 cal. BP), and the late Shang begins ca. 3300 cal. BP, and by convention ends at the historical date of 1046 BC.
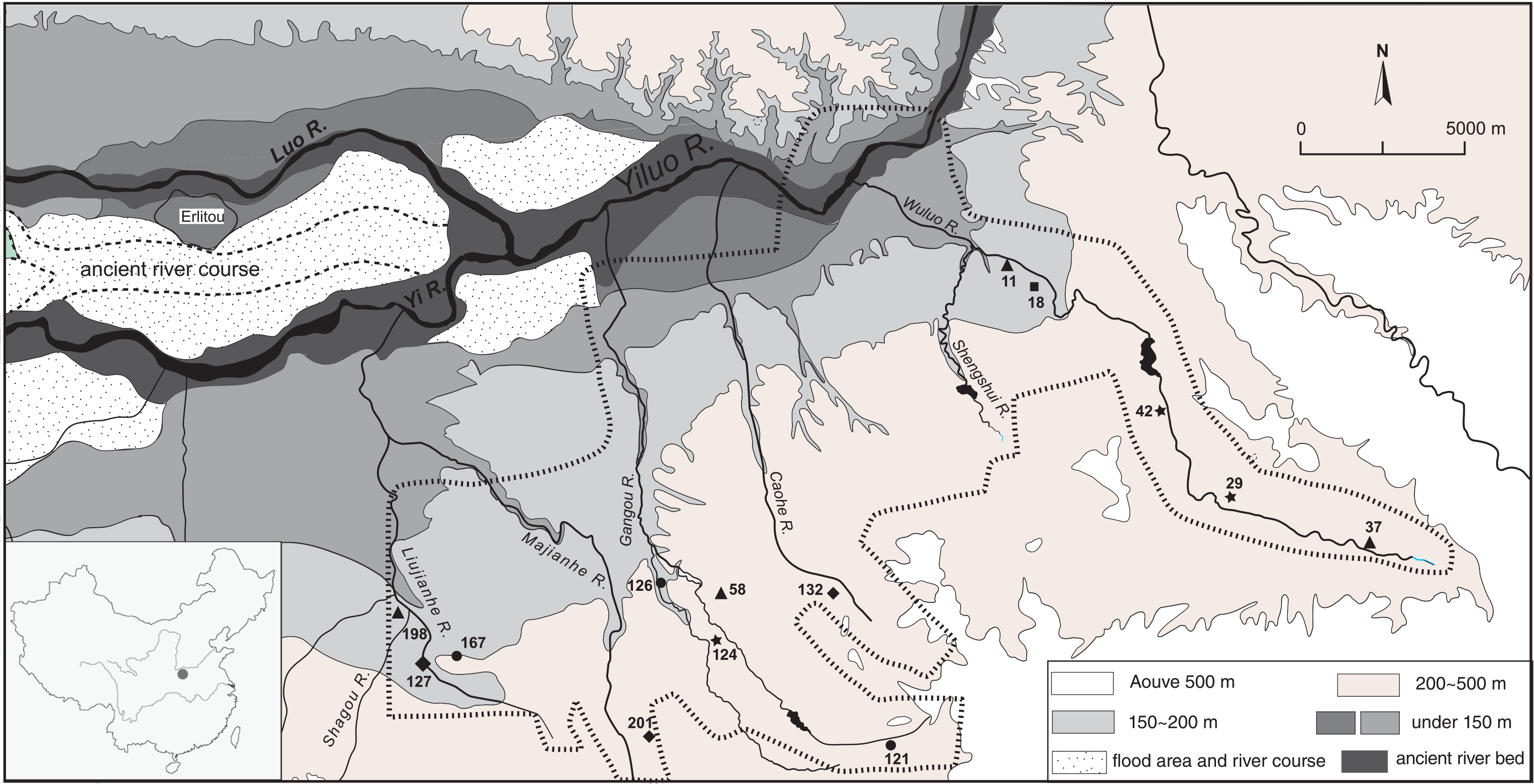
Map of the Yiluo valley. Sites inside the hashed line (| | | |) are those surveyed by the Yiluo Project team. Peiligang sites – ★; Yangshao sites – ■; Longshan sites – •; Erlitou sites – ♦; Erligang sites – ▲. Site codes and names are listed in Table 1.
This study includes analysis of 12 post-flotation samples taken from the soil matrix of Yiluo Project survey sites (Table 1), and a single feature from the Yangshao occupation of Huizui, a multi-component town site. Huizui would later develop into a craft production center, and it is possible that it served as a regional center even in the Yangshao (Lee and Bestel, 2007; Liu et al., 2007). The sample from Huizui was collected during the 2006 excavation, and all other samples came from pits revealed in terrace cuts during survey in 2007. The vast majority of seeds recovered overall were millet taxa (Panicoid). As expected among domesticates in the Yiluo region, foxtail millet was more common than broomcorn millet in all samples. Wheat was recovered from two out of three Upper Erligang sites. No rice was present in any sample discussed here, but is known from other Yiluo sites (Lee et al., 2007).
Number of Setaria-genus seeds recovered and sampled, relative frequency of Setaria (% of total number of seeds per site), and total seed density (seed number per liter of soil floated) in Yiluo basin sites.
Seeds from survey sites were recovered from middens filled with ash and other artifactual debris, whereas the Huizui sample was taken from a grain storage pit (see Supplementary Figure 1, available online). This in situ granary provides important contrast to the remains from survey sites, which were likely the domain of full-time farmers, and therefore may show patterns indicating specific behaviors related to agricultural production. Because of the relative abundance of seeds in the granary sample, the Huizui pit is the only feature used from the Yangshao period. For the purposes of site-to-site comparison, we have chosen seed samples from the Erligang phase at the survey site of Tunzhai, based on its particularly high seed density and preservation. Earlier studies show small sites like Tunzhai were probably agricultural villages, as opposed to political or economic centers (Lee et al., 2007). While in the following discussion of analyses and results, the feature from Huizui will be discussed as consumption-oriented and the feature from Tunzhai as production-oriented, it is important not to conflate the features with the sites themselves. Individual archaeological features may record specific behaviors, such as threshing or cooking, that are exclusively in the realm of food production or consumption, but sites are expected to be dedicated to either production or consumption only in a rarity of cases. Rather, the inhabitants of an agricultural village would have both farmed crops and eaten them, and some residents in a town or city may well have engaged in agricultural activities. The pattern of production and consumption features, however, is an important line of evidence in interpreting site function.
Materials and methods
In this study, we not only collected and measured millet from modern consumption contexts (i.e. commercially processed grains), but also obtained whole stalks of S. italica, which is commonly sold in the United States as pet food. This is not directly comparable with archaeological remains, as millet tends to ‘pop’ during charring, extending its lateral dimension (Markle and Rosch, 2008), but modern millet still provides essential data on morphological variation in S. italica crops.
When gathering archaeological samples for this study, seeds were pooled into a minimal number of categories (Figure 2). Seeds that appeared to belong to the genus Setaria were grouped into two categories: those that definitely represented domesticated foxtail millet (here referred to as S. italica), and those that were small and indeterminate in subspecies (Setaria sp.), which might include wild, weedy, and immature domesticated seeds. The purpose here was to have a subsample of seeds that were definitively domesticated, easily identified by overall size and plumpness that does not occur in wild seeds, so that the relationship of these to other seeds in the same genus could be investigated. Smaller Setaria seeds are not necessarily non-cultigen, but the vast majority could not be conclusively identified as S. viridis, immature S. italica (in sensu Song et al., 2013), or any other particular Setaria population.

Archaeological (lower) and modern (upper) Setaria, all shown with 500 micron scale bar. Modern specimens contrast a normal-sized (upper left) and a seed of intermediate maturity (upper right). Archaeological specimens show S. italica (far left) from the Erlitou-period Shang Zhuang site and three wild-sized Setaria from the Erligang-period Tunzhai (second from left) and the Longshan-period Nanshi sites (two on the right). A continuous black line outlining modern specimen (upper left) illustrates the area dimension measured digitally.
Once analyzed and counted, certain seeds were selected for detailed morphometry. Seeds were selected from their groupings primarily on the basis of intactness; the effort was made to choose only well-preserved, complete seeds, as to prevent biased size data from distorted shapes. A second consideration was variation. The full range of size and shape were pulled from samples where it was not possible to measure all fully intact seeds. Seeds were collected from each taxonomic group and time period. Because of the low seed density in Peiligang and Eastern Zhou samples, focus was limited to the four periods of Yangshao, Longshan, Erlitou, and Erligang.
Modern seeds were obtained from several contexts. To sample a human consumption context, seeds were obtained from a grocery store in Korea, representing domestically grown crops. One variety of foxtail millet intended for human consumption was sampled, as well as a second variety of foxtail millet that is instead intended as animal fodder. Both had undergone processing to remove unwanted items such as small seeds, chaff, and other waste. In the portion sampled, no significant difference in size was detected between the two types of foxtail, and they were subsequently grouped together as grocery seeds (see Supplementary Table 1, available online). Seeds intended for the purposes of planting were also obtained from this context (labeled as ‘seeds for sowing’), and these are somewhat larger on average, though also grown in Korea. Neither of these procurement methods controls for the location of the seed on the stalk or spike, and moreover, both represent a selection for preferred seeds, discarding smaller ones. In order to recreate a production context, it was necessary to obtain whole millet stalks, and the most expedient way of doing this was to purchase US-produced S. italica stalks intended as food for pet birds. While not produced in East Asia, and not intended for human consumption, these whole stalks still provide the opportunity to measure the morphological variation possible in a production context, and the morphological distribution of seeds along a single stalk.
Modern seeds intended for planting or pet food retained their paleae and lemmae, and these were removed in laboratory so as to be comparable to both archaeological and more processed modern seeds. Each modern and archaeological seed was measured in five dimensions: length, width, thickness, area of seeds, and embryo length (Figure 2). Thickness is measured with the seed in lateral view; the other measurements are all taken in dorsal view, with the indentation of the embryo visible. Length measures the point where the seed attaches to the rachis to the apex of the seed. Width is the maximum transverse dimension, perpendicular to length. The embryo (in modern specimens) or embryo scar (in archaeological specimens) is a distinctive feature on many grass family species, and its length is taken from the point where the seed attaches to the rachis, to the apex of the embryo. Thickness is the maximum transverse dimension with the seed in lateral view, measuring the distance between the dorsal and ventral planes. Area is perhaps a misleading label, as it measures not the actual three-dimensional surface area of the seed, but the two-dimensional area of the seed when in dorsal view, as if silhouetted (Figure 2). From the five measurements taken on each seed, we limit the following statistical analysis to thickness and area, as the most representative metrics. Area provides a precise two-dimensional measurement of the seed in cross-section, essentially encompassing both length and width measurements. Thickness measures the lateral axis of the seed, perpendicular to the plane of area. Thus, thickness and area provide a three-dimensional perspective on each seed while using only two variables. In addition, each seed was photographed as the measurements were taken. Measurements and photographs were taken with Nikon’s NIS Elements software, using camera feed from a Nikon AZ100 binocular microscope.
Results
Modern seeds
In this section, we will discuss analyses related to determining the range of variation found in modern seeds, in an attempt to establish a base level before moving on to archaeological seeds. As noted above, we use two variables, thickness and area, that together provide three-dimensional morphological data. These measurements are not independent variables, as all proportions of seeds increase as they become mature. This tendency makes all seeds fall along a single diagonal line in a bivariate analysis of area and thickness, when the seeds from a whole spike are measured (Figure 3). In agreement with Song et al. (2013), observations made during dissection and measuring noted that seeds were generally either round and plump, or extremely small, thin, and apparently undeveloped, with relatively few seeds of intermediate size. This provides the first major distinction between seeds from different contexts: millet from the grocery (consumption) context forms a single cluster, and seeds from the whole-stalk (production) context show bimodal variation (Figure 4). The distribution of morphological data has important ramifications for the types of statistical tests appropriate for determining distinctions between populations. For samples with normal distributions, where the average size is common and extremes are infrequent, a Student’s t test can determine whether two groups likely represent different populations. When the distribution is not normal, as with the whole-stalk S. italica, a non-parametric Wilcoxon test is more appropriate. Because grocery seeds show a distribution close to normal, and whole-stalk seeds do not, we will deploy both types of tests in our analyses below.

Thickness by area plot of modern S. italica seeds from a whole spike (n = 98, triangle), archaeological S. italica (F, n = 132), and wild-sized Setaria (s, n = 74). Specimens from the Yangshao storage pit were marked as a circle (•). Histograms illustrate thickness (vertical) and area (horizontal) distributions of archaeological specimens. Both histograms highlight a tight distribution of Yangshao seeds (light gray) in contrast to all archaeological seeds (black).
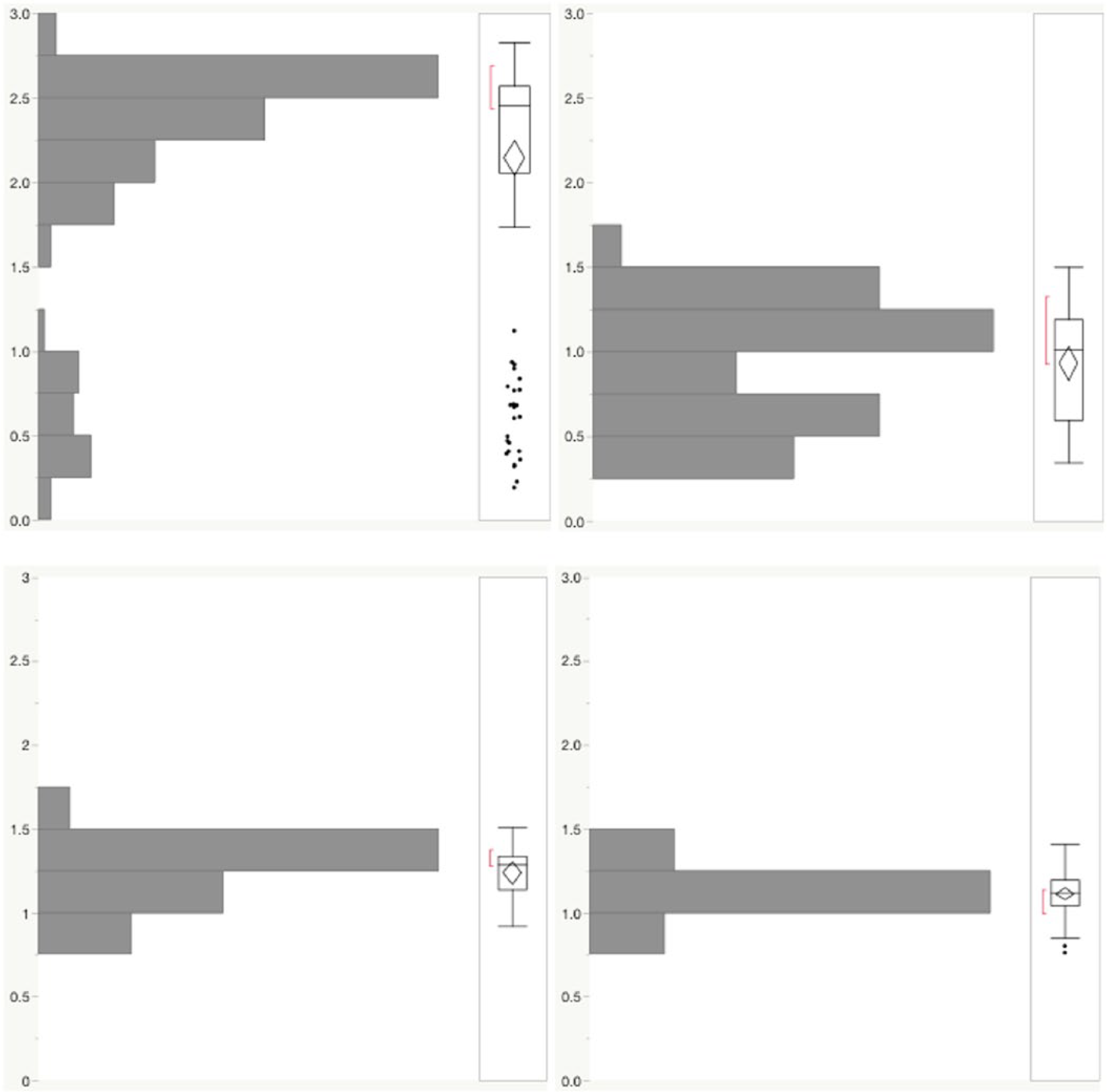
Histograms of area measurements for select Setaria populations. Modern seeds (left) show distinctly different distribution patterns dependent on context. Archaeological seeds (right) reflect a similar division between a village site (Tunzhai) and a larger town (Huizui).
Archaeological seeds
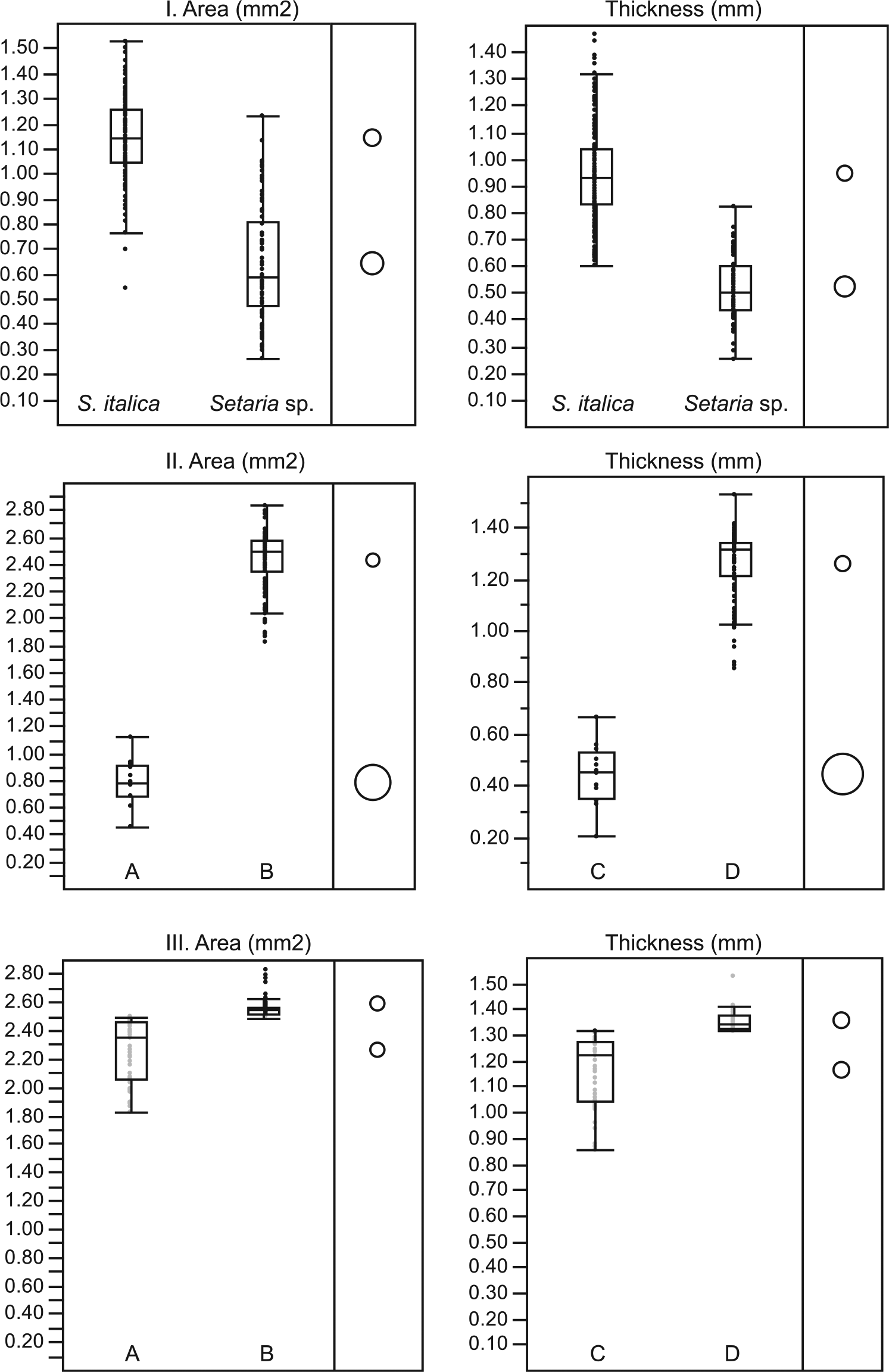
Archaeobotanical remains represent a fraction of the living plant populations of the past, and may or may not possess a similar range of morphological variation to the life assemblage from which they derived (Lee, 2012b). For this reason, we use the above examination of modern seeds to define the parameters for statistical tests on archaeological seeds. The ancient Setaria seeds used in this study do not show consistent size distributions, and we know from modern S. italica that a normal distribution should not be assumed, even if it is observed. The first pressing question for archaeological Setaria is if seeds identified as S. italica (domestic foxtail) can be distinguished from those labeled as Setaria sp. (small foxtail), and for this we used both Student’s t and Wilcoxon tests, as suggested by the modern Setaria analysis. Both tests show a significant difference between the populations of archaeological S. italica (n = 132) and Setaria sp. (n = 74), more than enough to justify their separation (Figure 5, Table 2). When plotted by thickness and area, the S. italica and Setaria sp. form a pattern that should be treated with suspicion. The two types of seeds overlap somewhat, with no obvious gaps or groupings along the diagonal line of data points, but very few seeds (<3%) over 1.1 mm2 in area or 0.8 mm in thickness are labeled Setaria sp. (Figure 3). This could be evidence not of a real division, but more of a cutoff point enforced on what is actually a continuum. This idea will be tested with the modern seeds below.
Student t tests run on seed thickness and area dimensions of archaeological S. italica and wild-sized Setaria (
Non-parametric difference tests run on seed thickness and area of archaeological Setaria italica and wild-sized Setaria (

The other main question raised by the archaeological seeds is that of context, whether seeds from the Yangshao storage pit show different morphological trends than those from survey sites. Given the above results, maintaining the distinction between domestic and small foxtail seeds at this level of analysis would be onerous, as S. italica data would phase out at a certain threshold, with Setaria sp. data picking up without an obvious gap, splitting the data between graphs for no apparent reason. To ease interpretation, we considered all Setaria seeds as a single group, rather than separated into domestic and small foxtail, with strong results. As was the case with the modern seeds, the distinction between production and consumption contexts can be seen clearly with the simple plot of thickness and area (Figure 3). The seeds from the Yangshao storage pit show the least variation, clustering in a manner (and, in fact, a morphological range) reminiscent of modern grocery seeds.
Comparison
Pooling data on all Setaria-genus seeds allows for more expedient statistical analysis, and now we will investigate whether this is statistically appropriate. The fact remains that both parametric and non-parametric tests identified the seeds in S. italica and Setaria sp. categories as significantly different. In this section, we question whether this reflects a genuine biological distinction or merely a self-fulfilling effect of the partition. We attempted to treat the modern S. italica obtained from whole stalks as if they were archaeological seeds, and we were not aware they represented a single species. This experiment was limited to a single stalk, to keep a single genotype in play. The first attempt was to create groupings based on the natural break in morphological data. To reflect the split in binomial distribution of these seeds (Figure 4), seeds with areas less than 1.2 mm2 were placed in group A (n = 12), and those above 1.8 mm2 in group B (n = 86). This essentially separates the plump seeds from the undersized ones. The same was done for thickness, with seeds below 0.7 mm in thickness placed in group C (n = 12), and above 0.8 mm in group D (n = 86), again reflecting the binomial split. Unintentionally but by no means coincidentally, the same 12 specimens comprise group A as group C, namely, the undersized seeds. Unsurprisingly, both Student’s t (Figure 5) and non-parametric tests (Table 2) identify the plump seeds and undersized seeds from the same stalk as significantly distinct populations, despite their being from not just the same species, but the same individual plant. Differences in seed size therefore cannot necessarily be expected to correspond to difference in taxon. Admittedly, this is proof by straw-man argument, but it provides a basis to try the experiment somewhat more randomly.
Using the data from the same S. italica stalk, we first eliminated the outlying undersized seeds, leaving only plump seeds (n = 86). We then used the median area measurement (2.5 mm2) as a cutoff point, placing half of the seeds with smaller areas into group A (n = 43), and half with larger areas into group B (n = 43). Using the median thickness (1.3 mm), the 43 smaller seeds were labeled group C, and the 43 larger seeds group D. This time groups A and C did not represent the same 43 specimens. Rather, some seeds had higher that median thickness but lower than median area, and vice versa.
Groups A and B, seeds from the same stalk divided in half only on the basis of their measured area, tested as different populations in every dimension but embryo length (which is distinctive at the genus level). The same held true for groups C and D, the same seeds divided by relative thickness. This suggests that statistical tests such as Student’s t and Wilcoxon used to compare archaeobotanical materials below the genus level may only be telling us what we already think we know. Any morphological cutoff, when reliably and consistently enforced, can create statistically distinct populations out of a continuum.
When archaeological Setaria seeds are treated as a single category, important comparisons can be made with the data we gathered from modern seeds. We have already shown how the Yangshao seeds in this study, all of which were recovered from the single, well-preserved context at Huizui, show less morphological variation than the other samples of archaeological seeds (Figure 3). For comparison, we selected Setaria seeds from the small, Erligang-period settlement (2 ha) at Tunzhai in the Yiluo region (Figure 1, Supplementary Figure 2, available online). The Tunzhai seeds therefore all come from a single, well-preserved context, but present an excellent counterpoint to the Huizui seeds as they represent the activities of full-time farmers, and therefore have a higher probability of preserving crop-production behaviors. As we learned with modern whole-stalk seeds and grocery seeds, the morphological distribution provides the clearest distinction between production and consumption contexts. In fact, the Tunzhai seeds and Huizui seeds present a remarkably similar scenario. The Erligang seeds show a discontinuous distribution with multiple peaks in the area dimension, similar to those of the modern seeds from a whole spike (Figure 4). The Yangshao seeds show a normal distribution, similar to those from a modern consumption context, represented by seeds from the grocery store.
In addition to the above statistical tests, all seed data, including at least three types of non-Setaria panicoid millets were submitted to principal component analysis (Supplementary Figure 3, available online) and correspondence analysis (Supplementary Figure 4, available online) alongside the Setaria seeds. The results were inconclusive, and can be found in our supplementary materials (available online).
Discussion
Distinguishing depositional contexts
Ethnographic studies on modern millet farmers have produced information on specific activities that may be traceable in the archaeological record (Reddy, 1997; Song et al., 2013). The most promising of these from the standpoint of a morphological study is winnowing, the primary purpose of which is the removal of smaller and/or incompletely filled seeds from crop yields (though this may not be the only point at which this occurs). Any archaeological assemblage of foxtail millets containing small and immature seeds should therefore represent deposition before or immediately after the winnowing process, and in a Neolithic or later context implies that only full-time farmers had so far interacted with the seeds. The feature at Tunzhai fits this model well, containing both large and small seeds and so likely representing a pre-winnowing deposition. As shown with comparison to modern seeds, a hallmark of this is not only the presence of a wide morphological range but also a discontinuous variation of seed size, indicating a break in ripening. Here again, Tunzhai is easily categorized as a production-oriented feature.
Consumption-oriented features, then, should have little evidence of extremely small seeds, and as seen in the sample of modern grocery seeds, morphological variation close to normal. The seeds recovered from the millet storage pit at Huizui align with these expectations (Figure 3). Consumption contexts become more common in sites as they are divorced from the work of food production, and focus instead on craft production and administration. This makes features such as the Huizui granary an important line of evidence for investigating specialization in Neolithic and early state societies.
Domestication status and agriculture
In analyzing the morphological variation of seeds to provide a lens toward depositional context, including all seeds identified to the genus Setaria proves useful. Perhaps the most intriguing difference between archaeological and modern Setaria is this issue of intermediate-sized seeds, largely absent from modern samples, but frequent archaeologically. Although the Erligang Setaria’s discontinuous distribution echoes that of modern whole-stalk seeds (Figure 4), the gulf between small and large seeds is not as pronounced. If the intermediate-sized seeds are S. italica, this could be because of the protracted nature of domestication, as a wild grass such as S. viridis produces seeds on a single spike that mature over the course of days or weeks (Lu, 1998). Simultaneous ripening may not have been entirely fixed in domesticated millet populations even into the historic period (in sensu Zeder et al., 2006). Delays in the consistent expression of domestic-type traits have been documented in the major crops of Southwest Asia such as wheat (Brown et al., 2009; Weiss et al., 2006) and Eastern North America (Gremillion and Piperno, 2009; Smith, 2011). In the Neolithic and at present, small seeds cannot be ruled out as resulting from agricultural processes and cannot be reliably identified as non-domesticated, as seen in East Asian legumes (Lee, 2012a; Lee et al., 2011).
Additionally, S. italica is not assumed to be the only species or variety represented in the small Setaria category. While very few seeds strongly resembled modern comparative samples of S. viridis, we must remember that S. italica and S. viridis are in fact conspecific. Smaller archaeological seeds could be weeds harvested along with the crop, foraged wild grass, or the result of admixture between wild and domestic species. However they were grown, they were processed in the same way as crop seeds and included in consumption storage, even after the extremely small seeds had been discarded. If ancient humans chose to keep these seeds together, it may be appropriate for archaeologists to consider them as a single category.
Conclusion
The statistical analyses of millet morphology in this study have provided several unique insights. Perhaps the most striking relates to methods of identification in archaeobotany, as distinctions below the genus level in Setaria were more a hindrance than a help to useful analysis in the cases outlined here. While we can be certain that obviously large and plump seeds are domesticated, the reverse is not true of smaller seeds. Over-identification is tempting but dangerous since, as shown by comparisons between modern and archaeological samples, the variation of seed morphology cannot be expected to look the same in comparative materials as it does in the archaeobotanical record. Enforcing a distinction between clearly domesticated foxtail millet and smaller seeds of the same genus remains prudent, and of course seeds clearly identifiable to species should be labeled as such, but these categories should be considered together in analyses of size and distribution patterns. In addition to proving statistically sound and helpful for behavioral inference, analyzing all Setaria-genus seeds together will better allow for comparison between researchers.
Second, morphometry of Setaria seeds in production and consumption contexts served to strengthen interpretations of feature type and settlement function. As predicted by Song et al. (2013), and borne out here by direct evidence from both modern and archaeological seeds, different economic activities lead to different morphological patterns in Setaria seeds recovered. This can be applied broadly, as it amounts to taking relatively few measurements of any staple crop in question. Third, no single statistical test applied here is appropriate for determining morphological distinction in plant species, but rather each question must take into consideration the level of distinction desired, the amount and distribution of data, and the implications of tests on comparative materials.
Although focused on seeds measured in fractions of millimeters, morphometry in archaeobotany provides insights on some of archaeology’s most intriguing questions. By thoroughly examining the morphological variation of millet in features over the millennia spanning the Neolithic, the early state period, and early dynastic China, we can address issues of food production, settlement patterning, and social complexity. The ability to distinguish feature types based on seed morphology, such as with the Yangshao storage pit at Huizui, can help track economic specialization and settlement hierarchy. In addition to allowing continually larger settlements throughout the Neolithic, agriculture was a necessary precursor to the establishment of urbanized states. The provisioning of cities with food is an immediate and crucial step in consolidating centralized power and authority (Gallagher and McIntosh, 2015; Smith, 2003; Zeder, 1991). As agricultural systems intensify and settlements become increasingly economically differentiated, patterns of production and consumption shift to meet these changing needs. By identifying key variables that distinguish between the remains of production- and consumption-oriented activities, this study provides a new method for analyzing archaeological seed remains with a view to the political economy of urbanization.
This study included only one feature associated with consumption, and yet was able to detect strong morphological patterns differentiating it from production-oriented features. More data from different archaeological contexts will likely reveal further discernable trends. We intend to refine this technique in the future, especially with the addition of modern data from other millet taxa, both wild and domesticated. A better understanding of the morphological variation in weedy Setaria and other Panicoid seeds will expand the possibilities for drawing inferences from archaeobotanical remains. We believe morphometric analysis used in this way has great potential, and would like to see it tested beyond north China, and beyond the millet tribe, with other staple foods in domestication centers around the world.
Footnotes
Acknowledgements
The data used in this article were collected in support of the Australian Research Council (DP 0450025, Liu), the University of Oregon Startup Fund (Lee), and the University of Oregon Foreign Language and Area Studies Fellowship (Walsh), and the Laboratory Program for Korean Studies through the Ministry of Education of Republic of Korea and Korean Studies Promotion Service of the Academy of Korean Studies (Lee and Walsh). We appreciate Frances White and Madonna Moss (University of Oregon), Gary W Crawford (University of Toronto), and the Huizui Archaeology Team for their support and insight. We are grateful for the comments of two anonymous reviewers, whose suggestions have greatly improved this paper.
Funding
This research was supported by the University of Oregon (Startup Fund, Foreign Language and Area Studies Fellowship), the Australian Research Council Department of Industry and Science (Grant 0450025), and the Laboratory Program for Korean Studies through the Ministry of Education of Republic of Korea and Korean Studies Promotion Service of the Academy of Korean Studies (AKS-2015-LAB).