
Abstract
An archaeological dimension expands the concept of biodiversity by generating long-term perspectives, combining multiple approaches and methodologies to enhance understanding of environmental changes. Aiming to investigate the patterns of biodiversity of marine mollusks over time, a comparison was made between three sets of archaeological sites (shellmounds) located in three geographic areas of the south-southeastern Brazilian coast. Sites dated between 1000–2000 and 4000–5000 yr BP were analyzed. A total of 37 bivalves and 26 gastropods were recorded. To describe the malacological community two indices of taxonomic diversity were used: average taxonomic distinctness and variation in taxonomic distinctness. The results showed a tendency of reduction in biodiversity patterns in the last 5000 years measured as average taxonomic distinctness. The observed reduction in biodiversity does not seem to be directly related to climate change over the period and there is a lack of data which could demonstrate any association between reduction in biodiversity and cultural causes such as preferences, technical level or food taboos of the prehistoric human populations. On the other hand, changes in sea level during this period are well documented and have been responsible for dramatic changes along the coast of this region. Therefore, it is likely that it could have affected local biodiversity and caused the observed pattern.
Introduction
A growing number of marine scientists are now including historical analyses that incorporate archaeological and other data sets into the development of better management plans, ecosystems restoration, and a sustainable oceans policy (Erlandson and Rick, 2008). Adding an archaeological dimension expands the concept of biodiversity by generating long-term perspectives, combining multiple approaches and methodologies to enhance understanding of environmental changes that are critical locally and globally (Jennings and Polunin, 1996; Kronen and Bender, 2007). Thus, based on archaeozoological research, it is possible to recover data about the past and build biodiversity scenarios over time. To that end, it is important to be familiarized with the taxonomic classification, the behavior, and the ecology of organisms, especially concepts related to biogeography, ecosystems, population ecology, and organism habits and habitats (Tchernov, 1992).
Many coastal sites contain accumulations of marine mollusks’ shells and other remains of organisms like crustacean and sea urchin test, fish, bird, and mammal bones (Figuti, 1993; Gaspar, 2000; Lima, 1991, 2000; Stein, 1992) left behind by fishing and foraging peoples. In optimal locations, populations sometimes lived continuously for centuries or returned repeatedly over the millennia, leaving stratified records of changes in local ecosystems through time (Erlandson and Rick, 2010). In such cases, shellmounds can provide long-term information on biodiversity, ecology, and impacts on coastal ecosystems.
Shellmounds are archaeological sites found in almost all coastal areas around the world. In Brazil, the term for it is sambaquis and they are particularly common on the central-southern coast, where they were built by prehistoric human populations that lived between 8000 and 2000 yr BP (Lima et al., 2002, 2003). According to Souza (1981), shellmounds are thanatocoenosis par excellence, that is, ensembles of dead organisms representative of the fauna existing in the area at the time in which they were formed. Most of these sites consist of a round, cone-shaped hill of varied size. The base usually measures from 2 to 10 m in diameter, while the height generally exceeds 2 m in height, although sometimes exceeding 25 m height. The amount of time that a population remained at a particular shellmound site varied depending on availability of food and other resources (Gaspar, 2000). Stratigraphic variation in the relative abundance of major food species can indicate changes in their economic importance or natural abundance over time and may provide important clues about the effects of humans on local ecosystems (Erlandson and Rick, 2010).
Shells left in archaeological sites attest to the use of the flesh for human food and for bait, as well as shells for building material, burial layers, and containers. Furthermore, this material shows that many of such gastropods and bivalves species have been modified into ornaments and artifacts. Through analyzing shells, it is possible to determine when the shells were harvested, how the environment developed over time, timing and effect of past cataclysmic environmental events, the geographical location along the coast where traded shells originated, the degree that shellfish contributed to the diet, and human competition with other predators (Claassen, 1998). In addition, because these sites contain sets of organisms representative of part of the mollusk fauna existing at the time they were built, they give insights into paleoenvironmental aspects related to these species and to their biodiversity and biogeography (Froyd and Willis, 2008; Fürsich, 1995; Lindbladh et al., 2007; Scheel-Ybert et al., 2006). However, several researchers (Amesbury, 1999, 2007; Amesbury et al., 1996; Jones, 2009; Keegan et al., 2003; Souza et al., 2012) analyzing the number of species, families, and functional diversity of alimentary guilds have recorded a shift in the diversity of bivalves and gastropods over time which can be related to environmental and/or cultural causes.
Aiming to investigate the patterns of biodiversity of marine mollusks over time, a comparison was made between three sets of archaeological sites located in three geographic areas of the Brazilian coast and describing two different intervals in time. These archaeozoological analyses contribute to the understanding of biodiversity through four perspectives: first, they can provide long-term data on marine mollusk diversity; second, they allow comparison of archaeological data between different localities along the Brazilian coast; third, their information can be used to understand and characterize the patterns of biodiversity of mollusks; and finally, the hypothesis of changes in pattern of biodiversity can be tested for a tropical marine context over the past 5000 years. Studies of this nature may prove valuable tools for a better understanding of the biodiversity and marine biogeography of the Brazilian coast.
Materials and methods
Changes in patterns of biodiversity over time were inferred by comparison between archaeological sites contiguous in space but with an age difference of 2000 years. A conservative procedure was assumed for comparison. For this study, only sites which had been 14C radiocarbon dated were used, leaving 156 usable sites remaining out of the 578 originally studied in Souza et al. (2012). From these 156 sites, only those that had no dating overlap were selected. To prevent any doubt regarding overlap, only those sites dated to between 1000–2000 and 4000–5000 yr BP were chosen. Thus, a safety margin of about 2000 years difference between sites was maintained. Of this group, only sites with more detailed malacological citations were used (‘detailed’ was defined based on citation of species which are not very common, therefore, showing both systematic and methodological care). From here, only sites that were very close geographically (maximum distance 10 km) were included, preventing regional ecological variation among them. Additionally, only locations which represent the same exploitation area for the shellmound’s people were used. Exploitation areas are defined by archaeologists as the distance radius walked by people of sambaquis to gather their resources and, generally, include different environments such as mangroves, lagoons, rock shores, sandy beaches, and so on (Kipfer, 2000).
Following all those procedures, only eight shellmounds remained subject to comparison: (A) – Sambaqui Pontinha (22°55′28″S, 42°31′01″W), Manitiba I (22°55′50.01″S, 42°34′58.81″W), and Beirada (22°55′32.04″S, 42°32′33.46″W), at Rio de Janeiro state; (B) – Sambaqui COSIPA 4 (22°55′28″S, 42°31′01″W), COSIPA 1 (23°52′29″S, 46°22′21″W), and Piaçaguera (23°52′S, 46°22′W), at São Paulo state; and (C) – Sambaqui Espinheiros II (26°16′59.9″S, 48°47′35.3″W) and Morro do Ouro (26°18′51.5″S, 48°49′37.9″W), at Santa Catarina state (Figure 1).
Study area: (A) Rio de Janeiro state, (B) São Paulo state, and (C) Santa Catarina state.
The remains from the archaeological sites of Rio de Janeiro state were part of the Museu Nacional/Universidade Federal do Rio de Janeiro’s collection, and the material from the sites of Santa Catarina are in the Museu Arqueológico de Sambaqui de Joinville. All of the malacological material pertaining to these particular sites was analyzed; however, it was not possible to screen the material from the shellmounds of São Paulo, and as such, data from these sites were obtained only from the literature.
The whole of the archaeological material was sieved through a mesh of 0.5 mm and the sea shells were cleaned from remaining sediment with a brush. The collection of malacological material was sorted by the following procedure: (1) selection of bivalves and gastropods, (2) cleaning with the help of a brush, in those cases where there was much sediment adhering to the shells. and (3) washing the shells so as to facilitate observation. When unopened bivalves were discovered, the sediment in their interior was preserved for other studies.
Shells were identified at the lowest possible taxonomic level. The following shell characteristics were analyzed: (1) bivalves – shape, type of hinge and sculpture, presence and shape of muscle scars, palial sinus, and palial line; (2) gastropods – type of spire (high, low, tapering), type and depth of suture, and the relation between the total height and the height of the body whorl, shape of spire whorls (straight sided, straight going to convex, straight going to concave) and sculpture, outer lip, columellar folds, siphonal, and posterior canals. After identification, the shells were re-bagged, labeled, and placed in the original bags and boxes. In addition, a survey of all publications related to the studied sites was carried out in the libraries of the institutions where the collections were inspected. This survey aimed to obtain data on the sites, such as location, dating, description, conservation state, dimensions, as well as verification of malacological citations.
For each archaeological site, an inventory was built of the malacological fauna from bibliographic analysis and sorting of archaeological material deposited on the technical reserves of the museums. Based on these inventories, a master list was constructed containing data on the presence (1) and absence (0) of the bivalve and gastropod encountered at each site analysis and frequency (f) obtained by dividing the number of sites where the species found by the total number (eight) of sites examined.
To describe the malacological community, two indices of taxonomic diversity were used. The first one, average taxonomic distinctness (AvTD), or Δ+, is defined as the average taxonomic path length between any two randomly chosen species, traced through a Linnaean or phylogenetic classification of the full set of species involved (Clarke and Warwick, 1998). Five taxonomic levels (species, genus, family, order, and class) were used with step lengths between them used as units. The second taxonomic diversity index used, variation in taxonomic distinctness (VarTD), or Λ+, reveals the evenness of the distribution of taxa across the hierarchical taxonomic tree. More precisely, this index provides additional information regarding the extent to which particular taxa are over- or under-represented in samples. Funnel plots with 95% confidence limits for Δ+ and Λ+ were constructed according to Clarke and Warwick (1998, 2001). In this approach, a randomization test was used to detect the expected Δ+ and Λ+ values derived from a pool of species from sambaquis of each of the region (master list). It is useful to examine both AvTD and VarTD because they capture independent aspects of the taxonomic diversity in a data set. These indices were calculated using PRIMER v6 (Clarke and Gorley, 2006).
Changes in patterns of biodiversity over time can be because of natural causes (such as changes in sea level during the studied period) or human impact (overexploitation of resources). Some of the possible causes were analyzed. To test the hypothesis related with sea-level changes, habitats of the molluskan species were defined. A chi-square test was performed to verify whether the number of lagoon and mangrove-associated species was different among samples (these species would be the first to disappear as sea level fells as their environment would be the most affected by such phenomenon). In the same way, Wilcoxon tests were used to verify tendencies of reduction in the number of species for each environment and also reduction in the number of feeding guilds among recent and older times. Both effects would be expected if changes in patterns of biodiversity were because of natural causes. In a similar way, the hypothesis of human impact was analyzed using Wilcoxon test to verify whether there was a tendency of reduction in species with known anthropological use in recent times compared with older times. Used species are supposed to be more intensively exploited than those not used.
Results
Table 1 presents a list of species of marine mollusks found at each shellmound that was analyzed with their habitat, depth, and frequency (f) obtained. A total of 37 bivalves has been identified, 35 being at the species level and 2 at the genus level comprised in 17 families were recorded and Veneridae Rafinesque, 1815 had the greatest diversity with 12 species, followed by Arcidae Lamarck, 1809 with four species and Cardiidae Lamarck, 1809 with three species. These three families together account for 51.4% of the total bivalves identified. The bivalves Anomalocardia brasiliana (Gmelin, 1791) and Lucina pectinata (Gmelin, 1791) occurred in 100% of the sites.
Presence (1) and absence (0) of the bivalve and gastropod species encountered at each site analyzed and frequency of each species relative to the eight studied sites (f). Data on sediment and habitat of living and depth range of occurrence are given.

PO: Pontinha; MA: Manitiba I; BE: Beirada; C4: COSIPA 4; C1: COSIPA 1; PI: Piaçaguera; ES: Espinheiros II; MO: Morro do Ouro; C: Coral; CA: calcareous algae; G: gravel; GA: green algae; M: mud; Mr: mangle roots; P: piling; R: Rock; S: sand; SG: sandy grass; SM: sand-mud.
A total of 26 gastropods have been identified, 24 being at the species level and 2 at the genus level comprised in 18 families were recorded. Naticidae Forbes 1838 and Olividae Latreille 1825 had the greatest diversity, both with two genera and four species. Four species occurred in the three studied areas: Stramonita haemastoma (Linnaeus, 1767) presents in 87.5% of the sites, Nassarius vibex (Say, 1822) in 75%, Olivancillaria vesica vesica (Gmelin, 1791), and Bulla striata Bruguière 1792 in 50%.
Among the sites of Rio de Janeiro state, Beirada showed the highest number of species (14 bivalves and 9 gastropods), followed by Pontinha with (11 bivalves and 3 gastropods), and finally Manitiba I (9 bivalves and 4 gastropods). In the state of São Paulo, Piaçaguera was the site with the greatest richness of species with 12 bivalves and 9 gastropods, followed by COSIPA 4 (10 bivalves and 7 gastropods) and COSIPA 1 (7 bivalves and 4 gastropods). In Santa Catarina state, Morro do Ouro (17 bivalves and 17 gastropods) presented a greater number of species than Espinheiros II (14 bivalves and 6 gastropods).
Comparative analysis between sites from different time periods within Rio de Janeiro state (Figure 2a) revealed that average taxonomic distinctness is significantly lower (p = 0.004) for recent (Pontinha) than eldest (Beirada and Manitiba) sites. Shellmounds of São Paulo (Figure 2b) presented one of the eldest sites (Piaçaguera) showing values of average taxonomic distinctness significantly higher than the expected (p = 0.018) in relation to the younger COSIPA 4 and from the also eldest COSIPA 1. Concerning sites from Santa Catarina state (Figure 2c), the observed pattern of decline of biodiversity over time was not statistically significant (Morro do Ouro, p = 0.152; Espinheiros II, p = 0.106). If Bivalves and Gastropods are analyzed separately for sites of all localities, the same pattern is recovered except for Gastropods from Rio de Janeiro sites. Pontinha (Rio de Janeiro) and Piaçaguera (São Paulo) are the only sites which present a statistically significant variation in taxonomic distinctness, meaning a certain level of bias on the averages of taxonomic distinctness obtained for these sites. This bias could be easily explained by the nature sambaqui’s remains which are accumulations subject to human selectivity. Also, differences in experimental procedures can’t be discarded as having some effect on these results. However, both of these effects are expected for all the studied sambaquis. The possible effect of natural and human impacts on the patterns of taxonomic distinctness obtained is discussed ahead.
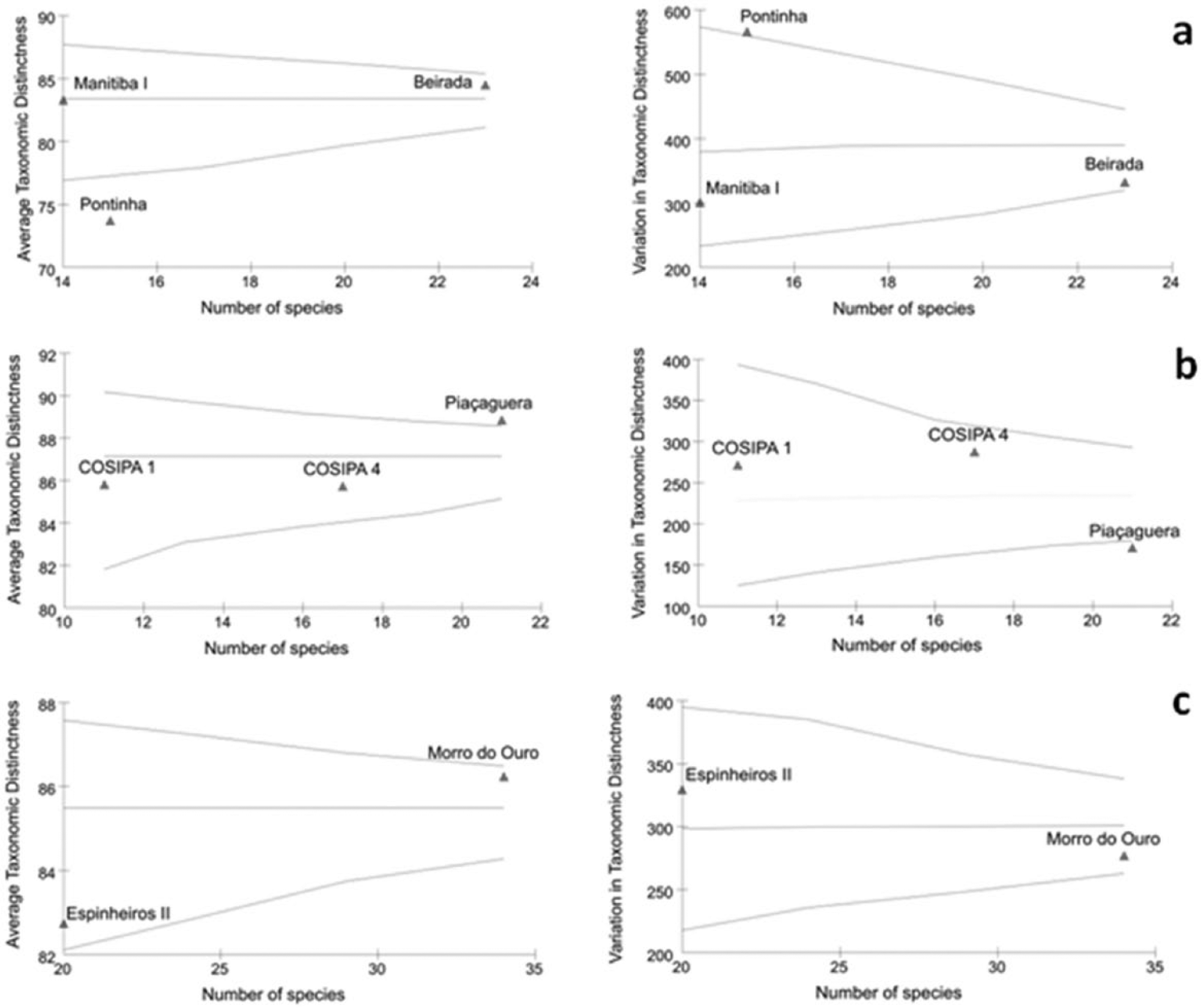
Average taxonomic distinctness and variation in taxonomic distinctness of marine mollusks species from (a) Rio de Janeiro, (b) São Paulo, and (c) Santa Catarina. The triangles represent the values found for the shellmounds and lines (funnel) represent the limit of 95%.
The chi-square performed between proportions of lagoon and mangrove-associated species among early and late sambaquis showed no statistical significant difference for any of the three different study areas (Rio de Janeiro, χ2 = 0.156, p = 0.925; São Paulo, χ2 = 0.034, p = 0.983; Santa Catarina, χ2 = 0.442, p = 0.506). The same is true for the results of Wilcoxon test performed for number of species of known anthropological use among recent and older times (W = 6; Z = 1.604; p = 0.109). However, the Wilcoxon test showed significant reduction in the number of species for each environment (W = 66; Z = 2.966; p = 0.003) and also for the number of feeding guilds (W = 21; Z = 2.251; p = 0.024) among recent and older times.
Discussion
The main result of this paper is the evidence that for three sets of archaeological sites on the southern and southeastern Brazilian coast, there seems to be a slight decrease in biodiversity in the more recent times (2000 yr BP) as compared with older time periods (4000 yr BP). The nature of vestiges accumulations in shellmounds as well as how they were studied can produce this effect of apparent reduction in the biodiversity. However, this observed pattern repeats in three independent sets of data, coming from three geographically distant regions. Thus, even though the hypothesis of an artifact generating these results cannot be discarded, it is unlikely since it would be expected by chance alone that at least one discordant result would be found. If it is assumed that gathering practices (human ‘sampling’) stayed constant in their modalities through the archaeological times, therefore, the explanation must be related to natural causes (e.g. climate, sea-level change) or cultural reasons (e.g. increased size of human populations, overexploitation of some resources), although they are not mutually exclusive.
Gathering practices of sambaqui people was restricted to manual collection carried out during low tide or by the use of simple tools manufactured of shells, bones, or animal teeth and lithic artifacts (Waselkov, 1987). The activity was also limited to relatively nearby areas of the camp (the exploitation areas). Figuti (1993, 1995), in studying the sambaquis of São Paulo (Cosipa 1 and Cosipa 4), was not able to find any difference in the fisheries techniques, which showed no improvement for the period of 4210 ± 90 to 1180 ± 60 BP. In contrast, Guimarães (2013) found, for sambaquis of Rio de Janeiro (Pontinha-recent; Manitiba and Beirada-older times), cultural differences associated with the type of caught fish (Micropogonias furniere was the preferred catch for older times which change for fishes from the Ariidae family in recent times). However, no differences were found regarding the mollusk gathering, which kept the same patterns of abundance over time. Studies concerned with changes in fish and gathering practices over time were not found for sambaquis from Santa Catarina. In fact, Nishida et al. (2004) investigating the traditional human communities of mollusks gatherers in Northeast Brazil concluded that gathering practices nowadays remains the same as those used by sambaqui people. Therefore, the assumption of no important changes in mollusk gathering practices over the period of the studied sambaquis seems a reasonable idea.
Anthracological and palynological analyses indicate little change for the period studied (Scheel-Ybert et al., 2009). Data obtained from the reconstruction of the paleoenvironment of the Rio de Janeiro and Santa Catarina coastal zones showed that the vegetation in the Upper Holocene (saltmarshes, mangroves, sandbank etc.) was virtually the same between 6000 and 1200 BP. In this sector of the coast, marked changes in the vegetation composition only occurred after the colonial and the post-colonial periods, and these changes were associated to wood extraction, settling on the coastal zone and tourism impacts (Scheel-Ybert, 2000, 2001). In a similar way, palynological records obtained in a bog flooded forest on the São Paulo coastal area did not show any variation of the vegetation associated to climate between 4300 and 1000 yr BP (Bissa et al., 2000; Ybert et al., 2003).
Regarding the cultural changes that could explain the observed pattern, certainly diverse factors, such as culture, preferences, technical level, food taboos, and the way the shells were discarded and/or utilized as building material, played a relevant role on the composition of the fauna found in shellmounds (Prummel and Heinrich, 2005). A peculiar and quite evident feature of archaeological sites is that the presence of organisms is related to the selectivity of the populations that built them. Furthermore, differences in species preservation potential and the researcher’s choices, objectives, excavated area, type of mesh used, and so on may also play an important role on shellmounds fauna composition determination (Scheel-Ybert et al., 2006). Although there is a lack of information on the demography of sambaqui human populations, the failure to demonstrate that the observed slight reduction in biodiversity was because of the reduction in the number of species of known anthropological use indicates that human impact can’t be taken as a strong cause for the observed pattern. Regarding the effects of excavation methodology on the determination of mollusk fauna recovered from the studied sambaquis, detailed information on the taken procedures is not available, what precludes further speculation on this.
Amesbury (2007) analyzing archaeological remains in the Mariana Islands also observed a change over time on the biodiversity of bivalves and gastropods compared with number of species, families, and functional diversity of alimentary guilds. Initially, this suggested the possibility of overexploitation of the resources. However, recent anthracological and palynological evidence supported another explanation, one of non-anthropogenic environmental change seems the more likely. In this case, the relative sea-level decline, which took place in the Mariana within the last 4000 years, caused the demise of mangrove habitats and of mollusks species.
According to Lima (2000), the rise of sea levels altered the Brazilian coastal contours bringing significant changes in the vegetation and fauna, making them more exuberant and providing more marine food resources to fisher-gatherer populations. Similarly, our data showed that changes occurred in the mollusk assemblages in a period after the optimum climate, when the Earth was 2–3° warmer and sea level was 3.5–4 m higher than nowadays (Angulo et al., 1999; Angulo and Lessa, 1997; Martin et al., 1979/1980). Taking into account the relatively high sea level during this period, the hypothesis that the observed pattern of higher mollusk diversity during the earlier times can be associated with the establishment of more flooding habitats along the coast can be put forward.
Generally, coastal dynamics are not the same during the ascent or descent of the relative level of the sea. Historical fluctuations in sea level were responsible for several changes in the coastal region, including for the formation of several lakes and lagoons (Paulay, 1989; Suguio et al., 2005). Indeed, the lagoon environment is considered to be particularly rich in fish and mollusks (Oyuela-Caycedo, 1996), for example, certain resources which in this habitat are much more accessible than in the open sea. Additionally, mangrove ecosystems are critical not only for sustaining biodiversity but also because of their direct and indirect benefits to human activities (Feller et al., 2010; Koch et al., 2009; Walters et al., 2008).
If sea-level changes are responsible for changes in patterns of biodiversity over time, a reduction in one (or some) of the biodiversity levels (namely, general number of species, community, taxonomic distinctness, functional diversity, and genetic diversity) would be expected, some of which were tested here. It was not possible to find any statistical significant differences in the number of lagoonal and mangrove-associated species between samples from any of the localities examined. These species should be most sensitive and those which should have disappeared from the samples as sea levels fell. However, a statistical significant reduction was observed for both: (1) number of mollusk species for the environments represented in the archaeozoological material and also (2) a loss of functional diversity inferred as a reduction in the number of feeding guilds.
In summary, the changes in patterns of biodiversity observed in this study was observed for taxonomic distinctness, functional diversity, and reduction in the number of species for all different environments represented in the archaeozoological material. This pattern of loss of biodiversity coincides with a decrease in sea level, which after reaching a peak (3–5 m above the present level) at around 5500 yr BP would have descended gradually until reaching the current level, without major fluctuations (Angulo and Lessa, 1997). This dynamic may have affected the patterns of biodiversity in the time range studied in the sense of reducing biodiversity in recent times.
Conclusion
The results for thanatocoenosis of mollusks from shellmounds on the coast of south-southeastern Brazil show a reduction in biodiversity patterns in the last 5000 years measured as average taxonomic distinctness, functional diversity, and reduction in the number of species in the different environments. It was not possible to demonstrate that this loss of biodiversity was because of resource depletion caused by a highly predatory and wasteful system of collection in which both juvenile forms as well as other species unsuitable for consumption were inexorably collected together with adult animals, what has been demonstrated by zooarchaeological studies. However, the observed reduction in biodiversity coincides with changes in sea level during this period, which are well documented and have been responsible for dramatic changes along the coast of this region. Therefore, it is likely that such sea-level changes have affected local biodiversity and causing the observed pattern.
Footnotes
Acknowledgements
We would like to acknowledge the following institutions which allowed analysis of their archaeological samples deposits at Museu Arqueológico de Sambaqui de Joinville and Museu Nacional/UFRJ. We also acknowledge the help of Rachel Harrington-Abrams who with her suggestions improved greatly the quality of this paper.
Funding
This work was supported by Brazilian Scientific Council-CNPq (grant number 477818/2006-4) and Coordination of Improvement of Higher Education Personnel – CAPES-PNPD (post-doctoral scholarship for R. Souza) (grant number 007340/2011-09).