
Abstract
The Holocene–Pleistocene transition in the upland loess-mantled regions of the central Great Plains is punctuated by the Brady Soil, which separates the late-Pleistocene Peoria Loess and the Holocene Bignell Loess. Previous research on the Brady Soil at the Old Wauneta Roadcut site in Southwestern Nebraska has produced paleoenvironmental information based on well-constrained luminescence and radiocarbon ages, stable carbon isotope data, and chemical and physical data. While the research indicated high effective moisture during formation of the Brady Soil and a shift to warm-season C4 vegetation from the cool-season C3-dominated vegetation of the Peoria Loess, those data do not provide any detail as to plant community composition and significant underlying climatic inferences. Assemblages of phytoliths and other biosilicates extracted from the Brady Soil provide specific information on vegetation communities and indicate shifts of plant taxa comprising these assemblages. Short-cell phytolith count data reveal a shift from dominance of Pooideae (C3) grasses, with relatively large numbers of arboreal dicot spheres and a few Cyperaceae (sedge) present in a savannah or open woodland in the Bølling-Allerød, to a mixed, open Chloridoideae (C4) and Pooideae (C3) grassland in the early-Holocene. Stipa-type Pooideae, a cool-season grass preferring drier soil conditions, marks the onset of the Younger Dryas. Large-cell phytoliths such as long cells, bulliforms, and trichomes, provided further definition of the climate history. This comprehensive biosilicate study of the Brady Soil has provided a more detailed paleoclimatic reconstruction than that generated with bulk sediment-derived δ13C data, or even with short-cell phytolith data alone.
Keywords
Introduction
The Pleistocene–Holocene Transition (PHT) was a time of rapidly changing climatic conditions associated with the final stages of deglaciation in the Northern Hemisphere (e.g. Alley, 2000; Zielinski and Mershon, 1997). Ice cores, for example, have yielded a great deal of high-resolution, paleoenvironmental information characterizing the PHT in the North Atlantic, leading to a better understanding of these abrupt climate changes (e.g. Alley, 2000; Orombelli et al., 2010; Zielinski and Mershon, 1997). North Atlantic climate of the late Pleistocene has been characterized by a warm Bølling–Allerød (B-A; ~14.7–12.65 k cal. yr BP) and a cold, arid Younger Dryas (YD; ~12.65–11.5 k cal. yr BP) nearing full-glacial conditions, transitioning back to warm conditions in the early-Holocene (Björck et al., 1998; Zielinski and Mershon, 1997). YD cooling has been linked to a temporally and spatially expanded polar vortex, which also increased the strength of the westerly wind flow, whereas warming in the early-Holocene is likely the result of expanded subtropical high circulation systems because of greater seasonality of temperatures (Alley, 2000; Zielinski and Mershon, 1997). These large and rapid climate fluctuations in the North Atlantic resulted from complex system interconnections, all of which are plausibly related to oceanic heat transfer variability (Alley, 2000). Complex climate interactions, however, can create diverse environmental conditions throughout the Northern Hemisphere and local expressions of the PHT climatic changes in continental interiors are often lacking in detail.
A sizable repository of environmental data for the late Quaternary exists within loess–paleosol stratigraphy of the loess-mantled, central Great Plains, a region providing a window into climatic patterns of the interior North American continent. Late-Pleistocene and Holocene loess of this region is thickest in south-central Nebraska, just south of and underlying the Nebraska Sand Hills (Figure 1), decreasing in thickness into Kansas and northeastern Colorado (Bettis et al., 2003; Kollmorgen, 1963; Mason, 1998, 2001; Thorp and Smith, 1952). Much of this loess has a strong provenance signature linking it to Oligocene and Miocene rocks of the White River and Arikaree Groups exposed northwest of the major loess deposits, in South Dakota, Nebraska, and Wyoming (Aleinikoff et al., 2008; Muhs et al., 2008). Based on those provenance data, sediment comprising the late-Pleistocene loess deposits is primarily from non-glacial dryland dust sources, much of it probably stored temporarily in dune fields immediately northwest of the thickest loess. Some loess was derived from the erosion of pre-existing late Quaternary loess deposits as well as fluvial and glaciofluvial systems (Mason, 2001; Mason et al., 2008; Muhs et al., 2008).
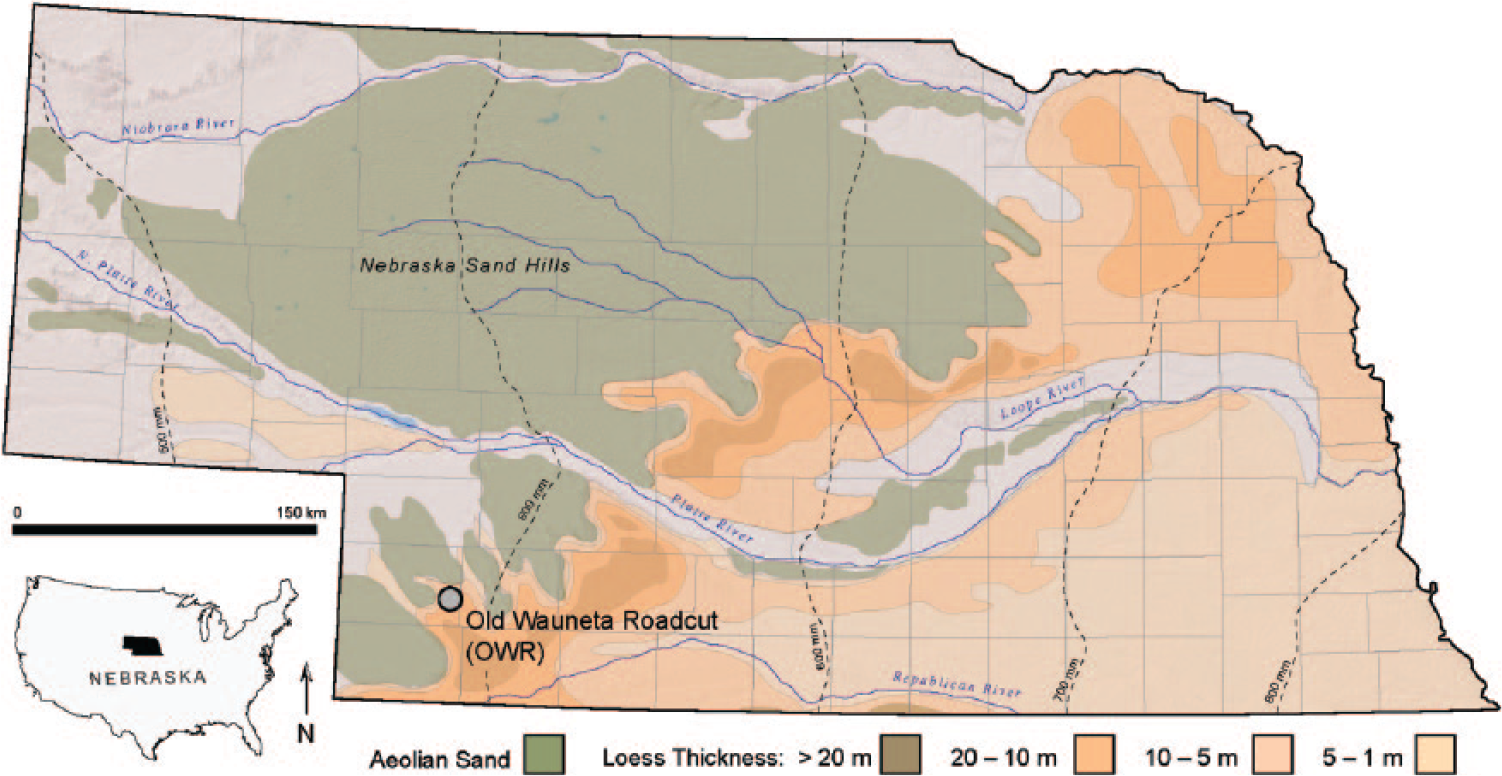
Old Wauneta Roadcut location in southwestern Nebraska. Aeolian sand and loess distribution and thickness (modified from Bettis et al., 2003).
Undifferentiated pre-Illinoian loess, Illinoian Loveland Loess, middle-Wisconsinan Gilman Canyon Formation, late-Wisconsinan Peoria Loess, and Holocene Bignell Loess collectively form the recognized sequence of loess packages within the central Great Plains (Figure 2; Baars and Maples, 1998; Johnson and Willey, 2000). This sequence contains intercalated paleosols, including the Brady Soil (Schultz and Stout, 1948), which was formed during an interval of well-defined upland pedogenesis spanning the PHT, and separates the Peoria and Bignell Loess units.

Regional late Quaternary loess/paleosol stratigraphy (modified from Baars and Maples (1998) and Johnson and Willey (2000)).
Mason et al. (2008) have constrained the beginning of Brady pedogenesis to ~14.7–13.5 k cal. yr BP and identified the termination ~9 k cal. yr BP. Pedogenesis was therefore initiated near the end of the late Pleistocene (through both the B-A and YD) and extended over 2000 years into the early-Holocene. Reconstructions of environmental conditions during the PHT, based primarily on soil organic matter–derived stable carbon isotope (δ13C) shifts from C3- to C4-dominant vegetation (Feggestad et al., 2004; Jacobs and Mason, 2004; Johnson and Willey, 2000; Mason et al., 2008; Miao et al., 2007) derived from the Brady Soil, indicate a general trend of warming and drying from the late Pleistocene into the early-Holocene. Cordova et al. (2011) linked the dominance of C3 plants in the late Pleistocene to decreased summer precipitation due to diminished subtropical air flow and associated the warming trend in the early-Holocene with increased seasonality and higher summer temperatures as a result of orbital forcing. Marin-Spiotta et al. (2014) found high concentrations of black carbon derived from biomass burning in the Brady Soil, which they attributed to the increase of C4 grasses and a warming climate during Brady pedogenesis, as well as to drier conditions as the burial of the Brady Soil began because of a return of high rates of loess flux.
Few locations have, however, provided definitive information on the YD interval for the Great Plains because the interval is not always expressed as a specific litho- or biostratigraphic unit, though available data have suggested a short-lived increase in aeolian activity during the YD (Holliday et al., 2011; Mason et al., 2008). This inferred YD signal within the central Great Plains loess record differs greatly from the cold, near full-glacial conditions of the North Atlantic region, but records from both regions have indicated similar drying trends. Notably, a coeval increase in dust concentration appears in the ice-core record from Greenland (Zielinski and Mershon, 1997).
Soil organic matter–derived δ13C data have been employed for environmental reconstruction globally because of the relative ease of analysis and low per-sample cost (e.g. Cerling, 1984; Wynn, 2000; and Roberts et al., 2013 in Africa; and Frakes and Jianzhong, 1994; Wang et al., 2008; and Lu et al., 2013 in China). In the central Great Plains, application of δ13C data derived from loess and other aeolian deposits has become common, for example, Muhs et al. (1999) in eastern Colorado, Feggestad et al. (2004), Miao et al. (2007), and Mason et al. (2008) in Nebraska, and Arbogast and Johnson (1998), Johnson and Willey (2000), Johnson et al. (2007), and Cordova et al. (2011) in Kansas.
A limitation of the δ13C climatic proxy is the generalized designation of vegetation as C3-dominated, C4-dominated, or mixed flora types without discriminating between woody vegetation and cool-season grasses. Global studies have been attempting to move beyond δ13C-based vegetation reconstructions by using the more time-consumptive pollen or phytolith analyses to produce distinct vegetation assemblages with family- or genus-level information (e.g. Alexandre et al., 1997; Barboni et al., 2007; Bremond et al., 2005; Garnier et al., 2013; Lu et al., 2006; Magill et al., 2013).
Extraction of proxy records of vegetation change beyond that of δ13C has been challenging in the central Great Plains due specifically to the lack of natural lake catchments and bogs for quality fossil pollen preservation. However, biogenic soil microfossils, such as phytoliths and other biosilicates, present an optimal resource for paleoenvironmental reconstructions in the Great Plains, an environment in which grasses dominate and fossil pollen grains are inconsistently preserved. A few biosilicate-based paleoenvironmental reconstructions have been reported for this region (e.g. Bozarth, 1992; Cordova et al., 2011; Fredlund and Tieszen, 1994, 1997; Fredlund et al., 1985; Kurmann, 1985; Twiss, 1983), and only one study has attempted to reconstruct the PHT through phytoliths from the Brady Soil (Bozarth, 1998).
Opal phytoliths are microscopic silica bodies formed by the uptake of water and dissolved materials from the soil and subsequent precipitation of silica within or between plant cells (Mulholland and Rapp, 1992b; Piperno, 1988, 2006). Because short-cell phytoliths in grasses can be identified to the subfamily level (Piperno, 2006), opal phytolith analysis is used to quantify paleovegetation assemblages based on distinguishing grass subfamilies (Pooideae, Chlorideae, and Panicoideae) and phytolith types for trees and shrubs. With phytolith frequency data permitting identification of specific changes in vegetation assemblages through time, they serve to enhance the knowledge of paleoenvironments. Large-cell phytoliths, such as long cells, bulliforms, and trichomes, have often been overlooked in previous phytolith studies in that they are primarily non-diagnostic phytolith forms, but they have been found useful in determining drought episodes and variations in evapotranspiration rates (Bremond et al., 2005; Delhon, 2007; Sangster and Parry, 1969).
The objective of this paper is to refine the environmental reconstruction of the PHT for the central Great Plains through the application of biosilicate analyses to a relatively high-resolution reconstruction of the paleovegetation record for the Brady Soil. Biosilicate classification and inventories permit a more quantitative reconstruction of vegetation assemblages than that provided by δ13C data alone. The goals of employing phytolith analysis herein were to (1) distinguish C3 plant types of woody plants and cool-season grasses to determine the extent to which woody plants grew at a given location, (2) ascertain whether or not a YD climate response can be detected with this proxy, and (3) determine relative progression of temperature and moisture conditions through the PHT and the relative magnitude of vegetation response.
Study site
Selection of the Old Wauneta Roadcut site (40.500561° N, 101.417872° W; ~1017 m a.m.s.l.) was based on its location along the exposed edge of a thick loess deposit that exhibits a clearly expressed Brady Soil (Figures 1 and 3) and on availability of extensive data from previous investigations, including detailed AMS 14C- and OSL-derived age profiles of the Brady Soil (Feggestad et al., 2004; Jacobs and Mason, 2004; Marin-Spiotta et al., 2014; Mason et al., 2008; Miao et al., 2005, 2007). The site is the roadcut exposure associated with an abandoned county road located ~10 km northwest of the town of Wauneta in Chase County, Nebraska, and ~75 km south of the North and South Platte River valleys (Figure 1). Modern climate characteristics, as recorded at Wauneta, Nebraska, include a January mean temperature of −1.3°C, a July mean temperature of 25.7°C, and mean annual precipitation of 493 mm, with the majority of the precipitation occurring in summer months (High Plains Regional Climate Center (HPRCC), 2013). Vegetative cover consists of a mixed-grass prairie with a variety of prairie-community forbs. The abandoned roadcut sits on a topographical high at the northern edge of a loess-mantled upland (regionally referred to as ‘tables’), forming the south bluff of the broad valley of Spring Creek. A west-facing, vertical cut exposes late Quaternary stratigraphy consisting of the late-Wisconsinan Peoria Loess and Holocene Bignell Loess, separated by the Brady Soil, formed within the uppermost Peoria Loess during the PHT (Figures 2 and 3).

Old Wauneta Roadcut exposure (a) with a pedostratigraphic profile of the Brady Soil (b) characterized by thick, cumulic, organic-rich horizons spanning the Pleistocene–Holocene transition.
Methods
A profile of the roadcut exposure was cleaned back to a fresh surface, measured, described and otherwise documented, and then sampled for δ13C and phytolith analysis. Discrete 5-cm samples were collected extending from the uppermost Peoria Loess, through the Brady Soil, and into the lowermost Bignell Loess. Although δ13C data for the Brady Soil at the Old Wauneta Roadcut were reported by Miao et al. (2007) and Mason et al. (2008), δ13C measurements were derived from the samples in this study for direct comparison to phytolith frequencies. Preparation for δ13C and percent organic carbon measurements included removal of roots and other organic debris by hand picking and removal of carbonates by treating samples with 1 M HCl followed by rinsing until a pH of 5 was obtained. Isotopic analyses were conducted at the University of Kansas WM Keck Paleoenvironmental and Environmental Stable Isotope Laboratory (KPESIL) using a Costech Elemental Analyzer to derive organic carbon content and a ThermoFinnigan MAT 253 isotope ratio mass spectrometer to determine raw δ13C values. A calibration curve was produced using soil, peach-leaf, and yeast standards to provide corrected δ13CVPDB values.
Phytoliths and other biosilicates were extracted from 5 g of sediment after dissolution of carbonates using 10% HCl, clay removal, oxidation of organic matter using 30% H2O2 (warmed in a water bath until reaction subsided), and separation of biosilicates from most other mineral matter using a ZnBr2 (s.g. = 2.3) heavy-liquid floatation (Bozarth, 2008). Sample isolates were mounted on microscope slides in immersion oil to allow for three-dimensional observation. Transects were scanned until ⩾200 short-cell phytoliths were identified and tallied. Diagnostic grass short cells consisting of shapes described as bilobate, cross, saddle, and trapezoid, each with many variations, were identified according to Twiss et al. (1969), Brown (1984), and Mulholland and Rapp (1992a). Distinguishable dicotyledon (tree and shrub) phytolith morphologies include spheres, polyhedrons, jigsaw-puzzle type, honeycomb assemblages, branched tracheids, segmented hairs, scalloped phytoliths, and platelets, and were classified based on descriptions by Bozarth (1992) and Wilding and Drees (1973). Additional biosilicates, such as large-cell phytoliths, sponge spicules, and diatoms, were tallied during microscopic analysis as supplementary proxy data. Frequency records were rendered in graphical format using Tilia software (Grimm, 2011), with assemblages displayed as percentages of the sum of all biosilicates.
Paleoenvironmental conditions have been expressed as three biosilicate-derived indices: (1) a C3:C4 grassland ratio derived from frequencies of short-cell grass phytoliths as described by Twiss (1987); (2) a Bulliform Index (BI) showing water stress because of evapotranspiration as described by Delhon (2007), and (3) a proposed Soil Moisture Index (SMI) derived from large-cell (bulliforms, trichomes, and long cells) grass phytolith frequencies in addition to the short-cell phytoliths. All indices were calculated from raw phytolith counts and converted to a scale of 0–1. The C3:C4 grassland ratio compares Pooideae to a sum of all short-cell phytoliths to express temperature characteristics. Values approaching 1 correlate to Pooideae-dominated (C3) cool-season grasslands, while values closer to zero are associated with warm-season Chloridoideae-dominated (C4) grasslands. The BI is the ratio of bulliform phytoliths to long-cell (elongated) phytoliths and is based on the concept that silicification of larger cells requires greater soil moisture availability to facilitate an increase in dissolved silica uptake (Delhon, 2007). An increase in temperature, in combination with greater soil moisture availability, will create an environment with increased evapotranspiration of water through the bulliform cells of a plant, leading to higher counts of bulliform phytoliths (Delhon, 2007; Sangster and Parry, 1969). Greater evapotranspiration, or water stress is therefore indicated by higher BI values approaching 1. The premise that relative high soil moisture allows for large-cell phytolith development was also exploited to create an SMI, which uses both short- and large-cell phytoliths to account for moisture availability from the atmosphere as well as from soil storage. The SMI is a modification of the aridity index originally introduced by Twiss (1987), who interpreted moisture levels based on Chloridoideae frequencies divided by the sum of short-cell phytoliths. More recent work on the distribution, phylogeny, and climate space of C4 grasses indicates, however, that the Twiss (1987) ratio is more reflective of temperature conditions (Edwards and Still, 2008), similar to the C3:C4 grassland ratio. The SMI calculation uses Panicoideae (C4 grass) frequencies to indicate the humid climate and/or high available soil moisture content required for growth (Bremond et al., 2005; Twiss, 1987), instead of representing warm temperatures. The SMI takes also into consideration genus-level grasses that indicate moisture availability. At the study site, Stipa-type Pooideae were observed, which are indicative of a cool-season (C3) grass that grows in arid conditions (Pohl, 1954). The formulation of SMI used herein is the following ratio: (Chloridoideae + Stipa-type)/(Chloridoideae + Stipa-type + Panicoideae + Pooideae + long cells + trichomes + bulliforms). SMI values range from 0 to 1, with the higher values indicating low-moisture availability.
Results and discussion
Stratigraphy
The Brady Soil at the study site is a well-developed, cumulic soil (cf. fine, smectitic, mesic Typic Argiustolls) with a solum of >1 m thick, which began to develop as the rate of Peoria Loess deposition diminished. Morphology of the Old Wauneta Roadcut has been described in previous research (Jacobs and Mason, 2004; Mason et al., 2008) and was sampled according to those findings. For the profile of this study, the surface of the Ab horizon begins ~580 cm below the modern surface, though above this is an ACb horizon, which presumably reflects pedogenesis dissipating because of initiation of Bignell Loess deposition. The dark colored (~10YR 3/1 moist) Ab horizon transitions into an ABb horizon that remains similarly dark in color (~10YR 4/1 moist), but displays slight carbonate content. Dark A-horizon colors correlate to increased soil carbon percentages and perhaps somewhat to increased concentrations of charred material (Marin-Spiotta et al., 2014) (Figure 4). A continued pickup in illuvial clay and carbonate content occurs at the base of the solum, within the Bkb horizon, which displays a slight shift to a hue yellower (~0.3Y 4/2 moist) than that of the overlying A horizons and decreased soil carbon content, to concentrations similar to those of the Peoria Loess (Figure 4).
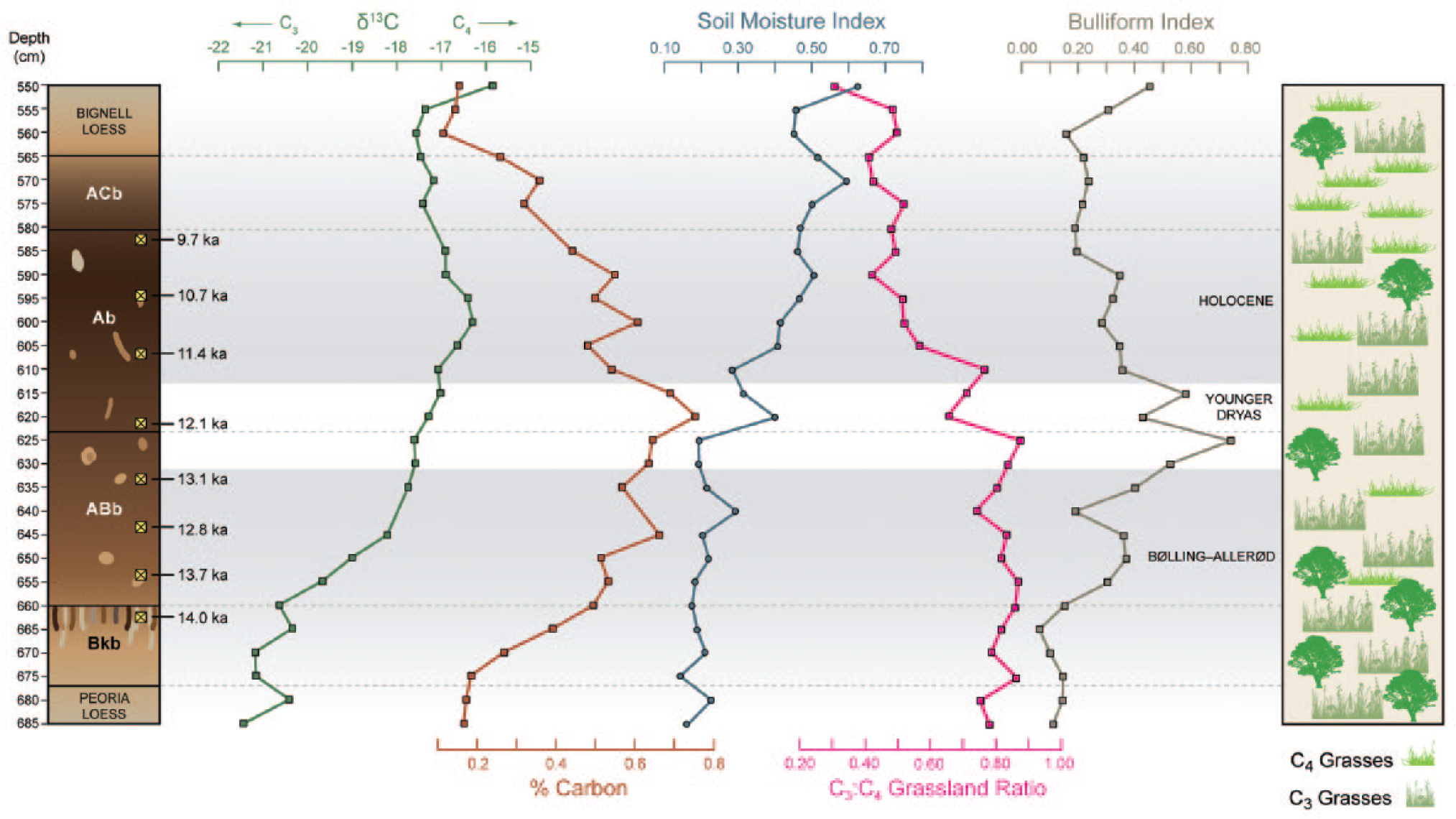
Stable carbon isotope (δ13C), percent organic carbon (%C), and climate index trends for the Brady Soil. Values are shown with respect to the Brady Soil stratigraphy and calibrated radiocarbon ages (Mason et al., 2008). C3:C4 Grassland Ratio values nearing zero are C4-dominant grasslands indicating warmer temperatures, while values nearing 1 are C3-dominant grasslands indicating cooler temperatures. Soil Moisture Index values nearing zero indicate more mesic conditions with values nearing 1 indicating increasing aridity. Bulliform Index values nearing zero equates to low water stress environments with increasing water stress and evapotranspiration occurring nearing a value of 1. The right column displays the general distribution of deciduous trees, C3 (tall) grasses, and C4 (short) grasses from the open woodland/savannah conditions in the late Pleistocene to the grass-dominated early-Holocene.
Considerations for proxy interpretation
With any proxy-based approach, there are issues to be considered before paleoenvironmental reconstructions can be confidently established. For the Brady Soil at the Old Wauneta Roadcut, the mixing of sediment by bioturbation requires consideration. Moderate (2–5 cm) to large (5–10 cm) burrow systems are visible within the soil profile and bounding Peoria and Bignell Loess units, and consequently can be avoided through careful sampling. A bulk, 5-cm sample size was chosen for this site because of the visible evidence of an ~5-cm mixing depth of high-density, small burrows (e.g. cicada nymph-like) that are difficult to avoid during sampling. Through observation and experimentation, this sampling approach provides a relatively high-resolution account of environmental properties without distortion by bioturbation signals. Mixing within soil horizons is not always easily detectable, however, and the approach to site assessment must be adjusted accordingly in heavily bioturbated soils. Pairing phytolith analysis with more generalized climate proxies, such as δ13C, and with the chronostratigraphy of Mason et al. (2008) provides substantiation of stratigraphic integrity. Age values are in stratigraphic sequence with the exception of the anomalous age of 12.8 ka within the B-A. However, given the consistency of results from sampling multiple vertical profiles at this site (Mason et al., 2008; Miao et al., 2007), the placement of B-A, YD, and early-Holocene age classifications are appropriate. A thorough examination of bioturbation at this site is presented in a separate study by Woodburn et al. (unpublished data).
Phytolith and isotope analyses
A total of 28 samples were examined to produce a relatively high-resolution vegetation reconstruction through the PHT (Figure 5). A stratigraphically constrained cluster analysis of phytolith frequencies (CONISS) (Grimm, 1987) performed in Tilia defined three major zones of vegetation assemblages. These zones correspond with the timing of North Atlantic climate episode chronostratigraphy and include the B-A, YD, and the early-Holocene based on the distribution of calibrated radiocarbon ages obtained by Mason et al. (2008) (Figures 4 and 5). δ13C analysis for this study shows a similar range of values (though with less variability) as those provided by Miao et al. (2007). Both δ13C and phytolith counts indicate a mixed C3/C4 signal through the PHT, with C3 dominance in the late Pleistocene and C4 grasses increasing in the early-Holocene, though phytolith analysis established that late-Pleistocene B-A vegetation has a greater C3 dominance than indicated by δ13C.
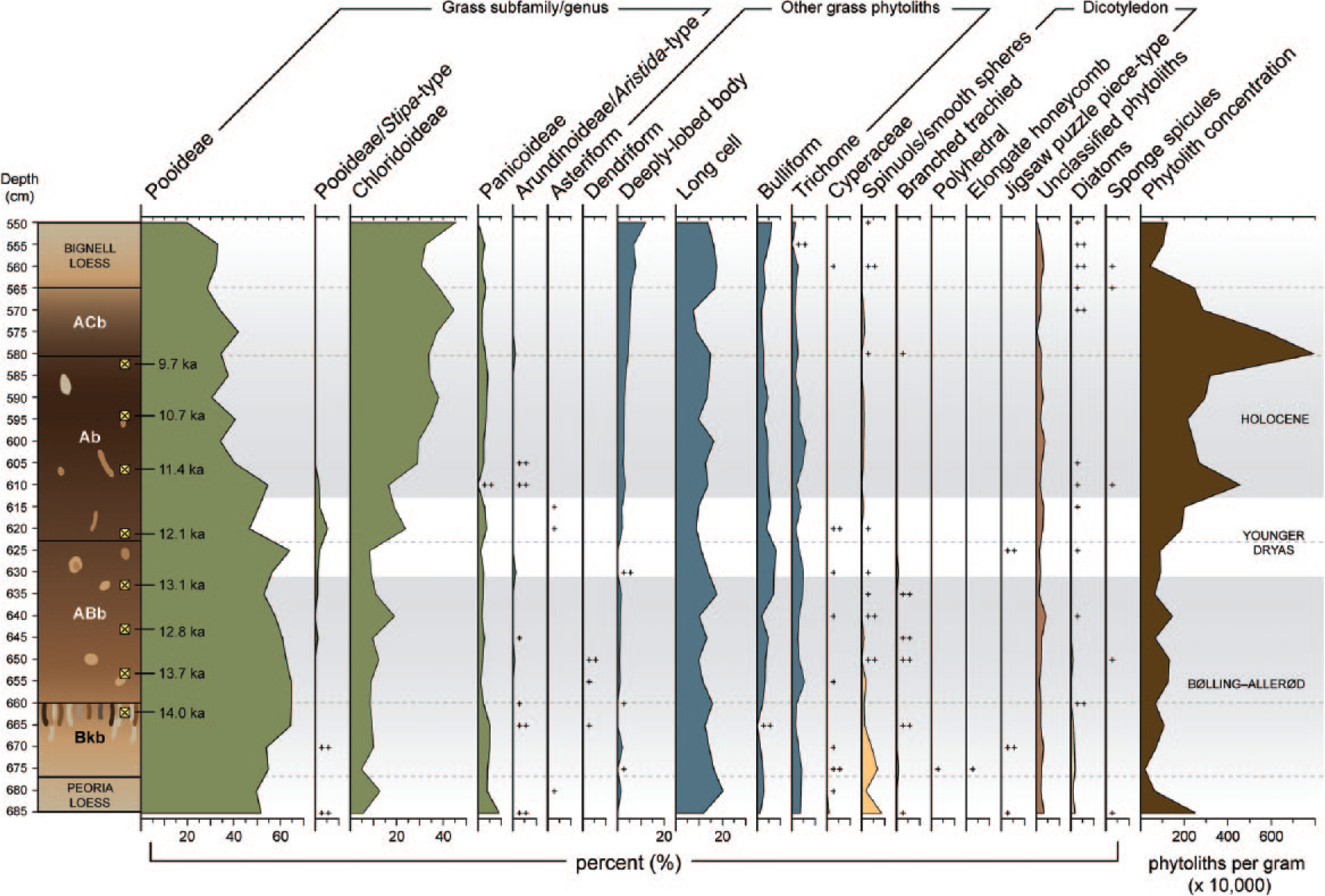
Biosilicate frequencies and phytolith concentrations shown with respect to the Brady Soil stratigraphy. Shaded zones indicate distinct vegetation assemblages that correlate to the North Atlantic chronosequence.
Vegetation dynamics
Stable carbon isotope data correlate well to the general trends of C3 and C4 phytolith frequencies, though the use of phytolith analysis provided the capacity to distinguish C3 plant types of cool-season grasses and woody plants, as well as quantification of shifts in vegetation assemblages. Paleoenvironmental indices constructed using phytolith information added the ability to detect subtle climatic adjustments, thereby further articulating the nature of the environmental change.
Phytolith analysis shows that the C3 signal in the B-A is due largely to cool-season Pooideae grasses, which produce trapezoidal short cells, as well as to the presence of trees and shrubs as indicated by spheres, branched trachieds, and other dicotyledon phytolith types (Figure 5). Phytoliths from woody plants are typically rare types because of low production and are consequently underrepresented in the phytolith record (Piperno, 2006). Dicotyledons also produce low quantities of silica bodies, and some types such as the thin, fragile polyhedral-type phytoliths are poorly preserved (Piperno, 2006). Accordingly, the appearance or increased frequency of woody plant phytoliths denotes a change from grassland to a more wooded vegetation structure. Moreover, within the Bkb horizon (at the depth of 675 cm), elongated honeycomb and polyhedral-type phytoliths were identified, thereby representing the presence of deciduous trees (Figure 5), an indication of greater moisture availability during the B-A.
Pooideae (C3) grass dominance continues through the YD, which signifies cool temperatures, but an increase in Chloridoideae (C4) grasses occurs for a short interval, as recorded from the sample taken at 620 cm (Figure 5). This is in agreement with findings by Bozarth (1998), though he reported data from only four samples dispersed throughout the Brady Soil. An associated peak in Stipa-type (a cool-season Pooideae grass growing in arid environments) bilobate phytoliths suggests, however, a decrease in moisture availability. The peak in Stipa is also coeval with slight decreases in long-cell phytoliths within the YD, thereby corroborating lower moisture availability. In the early-Holocene, a further increase in warm-season (C4) Chloridoideae grasses, revealed by greater counts of squat, saddle-shaped phytoliths and reduced concentrations of dicotyledon phytoliths, characterize a warming and drying trend.
Low counts of Cyperaceae (sedge) cone phytoliths and diatom biosilicates (found only as small, broken pieces) co-occur at a few depths within the profile (Figure 5). Their highest frequencies appear in two intervals of differing environmental conditions: the cool, moist Pleistocene B-A (when Brady Soil formation was commencing) and the warm, dry early-Holocene (when soil formation was ending). Occurrences correspond to the highest rates of sediment input during the end of the Peoria Loess deposition within the Bkb and ABb horizons of the Brady Soil, and to the onset of Bignell Loess deposition (Figure 5). In this topographic setting, Cyperaceae and diatoms were very likely transported with the loess clastic component and are markers for increased aeolian sediment influx. Potential sources include the adjacent Spring Creek valley, playas on the uplands northwest of the site, the Platte River valley, and perhaps the Miocene Ogallala Formation, which includes fluvial and lacustrine facies.
High-moisture availability during the late Pleistocene, indicated by the low values of the SMI (Figure 4), denotes mesic conditions at this time, as do the Pooideae-dominated grasses and woody vegetation. Evapotranspiration rates signified by the BI (Figure 4) are lowest in the Bkb soil horizon during the late Pleistocene when woody vegetation was at its peak. The SMI shifts to exhibit moderate and drier conditions as pedogenesis continues into the Holocene, with a rapid drying trend in the middle of the YD. Reconstructed evapotranspiration exhibits two maxima during the beginning and near the end of the YD, yet the vegetation assemblage was still dominated by C3 types because of sufficient levels of moisture still available during these periods. In that C3 grasses retain their dominance during the early and late YD, an increase in temperature must have occurred to force the increase in evapotranspiration. During the B-A and YD, there are also periodic decreases in evapotranspiration (BI) that correspond to relatively short periods of increased aridity as indicated by the SMI (specifically at 620 and 640 cm; Figure 4). A lowered rate of evapotranspiration during times of drought seems counterintuitive, but these shifts in the indices can be explained by considering the specific short-term changes in associated vegetation, including the peak in Chloridoideae in the B-A and YD (at 640 and 620 cm), and in drought-tolerant Stipa-type Pooideae in the YD (at 620 cm) (Figures 4 and 5). The increase in these vegetation types strongly influences SMI, indicating greater aridity; however, because the Chloridoideae and Stipa use moisture more efficiently, the BI values are lowered in times when cooler temperatures are not the controlling factor. Similarly, although the overall decrease in the BI evapotranspiration seen with warming temperatures in the early-Holocene also seems counterintuitive (less evapotranspiration with warmer temperatures), it is partially controlled by the increase in more water-efficient C4 Chloridoideae vegetation.
The nearly equivalent concentrations of cool-season Pooideae and warm-season Chloridoideae during the Holocene imply an increased seasonality with increased summer temperatures. The formation of a rainy season is also suggested by these concentrations. It is possible that soil moisture retention during and after the rainy season was greater because of an increase in soil-water-storage capacity in the Ab horizon, as supported by the similar levels of Panicoideae during both the late Pleistocene and Holocene. These conditions would allow for the high percentage of Pooideae grasses to continue thriving in a warmer climate. During the termination of Brady Soil pedogenesis, evapotranspiration rates increase once again with the increases in temperature and corresponding aridity, and with the acceleration of Bignell Loess deposition (Figure 4).
This investigation provides another level of support to the changing environmental conditions through the PHT, and findings correlate well with other studies in the central Great Plains (e.g. Cordova et al., 2011; Marin-Spiotta et al., 2014; Mason et al., 2008; Miao et al., 2007). The change to warmer and drier conditions occurs gradually in the early-Holocene based on similar frequencies of Pooideae and Chloridoideae followed by continued increase of Chloridoideae at the beginning of Bignell Loess sedimentation. Enhanced seasonality and the increase of C4 grasses in the early-Holocene are similar to the trend reported by Cordova et al. (2011); however, this shift occurred earlier in southwestern Nebraska than the 10.2 k cal. yr BP reported at Kansas sites. The onset of this climate transition occurs more than 2000 years prior to the end of Brady Soil formation, yet stable surface conditions and pedogenesis continued during this time of increased aridity. Mason et al. (2008) attributed the mature nature of the Brady Soil to an extended period of high effective moisture across the PHT. Climate indices produced from phytolith data have shown that the B-A had high moisture availability paired with relatively low evapotranspiration rates. Lower precipitation characterizes the period of early-Holocene pedogenesis, yet available soil moisture slightly offsets decreased precipitation. Low evapotranspiration rates persisted in the early-Holocene because of an increase in C4 grass concentrations. Both of these situations describe environments with effective moisture sufficient for pedogenesis. Increased water stress on plants and a spike in aridity during the YD are environmental conditions that support the possibility for a brief rejuvenation of loess sedimentation postulated by Mason et al. (2008). Based on phytolith concentrations, surface conditions are most stable in the early-Holocene immediately following the YD, continuing to the top of the Ab horizon at 580 cm when loess influx rates increase once more. During Bignell Loess deposition, the dramatic increase in aridity, coupled with increased evapotranspiration rates, leads to an environment of low effective moisture, and surface instability ensues.
Conclusion
Data and interpretations provided herein represent the first known paleovegetation reconstruction across the PHT, as recorded by phytoliths, in the Brady Soil of the central Great Plains. The use of phytolith frequencies in concert with multiple paleoenvironmental indices provides detailed vegetation characteristics in loess–paleosol settings in the absence of fossil pollen records. At the Old Wauneta Roadcut site, the general trend of a cool, mesic Pleistocene transitioning to a warm, dryer Holocene was previously documented with δ13C data (Miao et al., 2007). Phytolith analysis confirms this trend, but revealed a significant level of additional information. Phytolith data indicate that during the onset of Brady Soil formation in the late-Pleistocene B-A, the region was in savannah to open woodland, subsequently transitioning to an open grassland environment by the early-Holocene. Another significant contribution of phytolith analysis to the reconstruction of the environment during formation of the Brady Soil is the documentation of the short-term environmental change during the YD, characterized by the distinctive Stipa-type Pooideae signal, a decrease in long-cell phytoliths, and dramatic changes in water stress. Hence, phytolith analysis displayed the warming and drying sequences not evident in the δ13C data.
Supplementary moisture information, obtained through large-cell phytolith counts and derived SMI and BI, reveals shifts in seasonality and occurrence of short or limited intensity drought episodes within larger climate events. As expressed in these indices, the early-Holocene mixed-grass prairie environment indicates a change to greater seasonality, with warmer summers and a rainy season, whereas the onset of Bignell Loess deposition was coeval with both aridity and temperature increases. Values from the proposed SMI calculation correlates well with general climate trends shown by other proxies, yet provides data for relatively short-term (few ka) drought periods. SMI and BI values provide useful information when determining the controlling factor (temperature and/or water availability) for changes in vegetation communities.
Footnotes
Acknowledgements
The authors are grateful to the landowners for permitting them to make repeated visits to the site.
Funding
Funding for this work was provided by awards from the University of Kansas General Research Fund and a Kollmorgen Fellowship from the University of Kansas Department of Geography and Atmospheric Science.