
Abstract
The aims of this article are, first, to investigate the middle- and upper-Holocene woodland history along the altitudinal gradient between the lowlands and uplands of Central Europe (190–550 m a.s.l.) and, second, to outline possible biases inherent in the charcoal record based on a comparison with the pollen record and its known biases. Our anthracological data set contains 42,547 determinations made in 120 charcoal samples taken at 69 sites. The lowest elevated part of the study region (below 200 m a.s.l.) is characterized by the long-term presence of a species-rich hardwood forest (mixed oak–elm–ash forest). Quercus charcoals dominated in the rest of the altitude zones during the Neolithic and Aeneolithic; however, shrub charcoals appearing in samples from areas with chernozem soils (generally up to 230 m a.s.l.) indicate open-canopy oak woodlands. The species composition differed along the altitudinal gradient during the Bronze Age period, when Carpinus, Fagus and Abies expanded to altitudes above 230 m a.s.l., while Fagus was more abundant above 290 m a.s.l. Broadleaved trees (Quercus, Fraxinus, Ulmus, Acer and Carpinus) and shrubs are generally more represented in charcoals than pollen. Since broadleaved trees are usually nutrient demanding and able to re-grow easily after being felled, we suppose that their charcoal record is influenced by two main factors: bias of the initial location of the archaeological site and bias caused by long-term human influence on forest vegetation in the vicinity of settlements. These results underline that combining charcoal and pollen analysis has great potential for studying phenomena in cultural landscapes, as each of the methods approaches nature from the opposite side of the human–nature gradient.
Introduction
Postglacial tree succession in Central Europe is characterized by gradual changes in tree composition, which are associated with expansions and declines of woody species (Firbas, 1949). The vegetation history of woodlands has long been influenced by human activities.
Traditionally, reconstructions of vegetation history have been based on interpretations of pollen data. The most advanced techniques deal with the complexity of the pollen–vegetation relationship numerically. Quantitative vegetation reconstructions can filter out the biases of pollen production and pollen dispersal (e.g. Hellman et al., 2008; Sugita, 2007a). Pollen sites, by contrast, especially in the lowlands of Central Europe, host an abundant cover of trees and herbs (e.g. Alnus, Salix and Poaceae), which are usually the target taxa of vegetation reconstructions. The quantitative approach to pollen analysis nevertheless also has to deal with the effect of local taxa (Abraham et al., 2014) because its assumption about the sedimentation basin as an area free of target taxa is rarely fulfilled.
Analysis of soil charcoals or charcoals from archaeological sites offers one alternative approach to reconstructing past vegetation at terrestrial sites over centuries and millennia (e.g. Asouti and Austin, 2005; Clark et al., 1998; Nelle et al., 2013; Robin et al., 2012). Especially since the matrix of Central European vegetation is largely composed of terrestrial habitats, where pollen grains are usually not conserved. The charcoal assemblage obtained from an archaeological site depends on several factors that have to be taken into account when interpreting anthracological data (Marston, 2009; Théry-Parisot et al., 2010). It is assumed that the composition of firewood is primarily a function of its availability, as people probably obtained firewood from woody species growing in the close vicinity of their settlements – today’s archaeological sites (Jansen et al., 2013; Shackleton and Prins, 1992). The collection of common firewood is mostly considered non-selective, but collecting wood for specialized production (e.g. metallurgy, funerary customs or house building) is highly selective (Marston, 2009; Rubiales et al., 2011; Théry-Parisot, et al. 2010). Charcoal spectra of woods serving special purposes are frequently connected to specific archaeological objects, and the record generally shows reduced species composition (e.g. Novák et al., 2012). Although charcoal data from archaeological sites are widely used for reconstructing woodland vegetation in Central Europe, the combination of pollen and anthracological data has been used only sporadically in the landscape context (e.g. Kozáková et al., 2011; Nelle et al., 2010).
Central and northern Moravia (the Moravian Gate and the Upper Moravian Vale) is a phytogeographically important corridor between the Pannonian and Polish Lowlands, but their Holocene vegetation history has not been sufficiently studied. The northern part of the Pannonian Basin and the Western Carpathians play an important role in the florogenesis of Central Europe in terms of the existence of cryptic glacial refugia and as a migration route of thermophilous species (e.g. Jamrichová et al., 2014; Provan and Bennett, 2008; Stewart and Lister, 2001). The study region is influenced by long-term agricultural management (Čižmářová et al., 1996), so a relatively rich anthracological record from archaeological sites is available. The pollen record is limited partly due to the lack of sediments suitable for pollen analysis and due to inconsistent palynological exploitation of this area. Natural archives are restricted to wetland habitats scattered around the river Morava. Only two of them (Černovír and Hrdibořice) have been investigated palynologically (Jankovská, 2003; Puchmajerová, 1947; Salaschek, 1936).
We aimed to reconstruct the woodland history of the middle- and upper-Holocene (7500–1450 BP) using charcoal data from archaeological sites and pollen data from natural profiles. Based on our experience with present-day vegetation and taphonomic process, we expected differences in species composition along the altitudinal gradient and also differences between the pollen and charcoal records and the biases inherent in them. Our main questions were as follows: (1) Do anthracological data show any pattern along the altitudinal gradient? (2) For which species is the anthracological record biased compared with the pollen one and its biases?
Materials and methods
Study area and present-day vegetation
The investigation was carried out in central and northern Moravia (NE Czech Republic; Figure 1). The lowland along the rivers Morava and Odra is framed by uplands to the west, east and north. Since the Neolithic, natural vegetation has been influenced by long-term agricultural activities (Čižmář, 2004); however, the presence of humans as early as the Palaeolithic has been documented (e.g. Neruda and Nerudová, 2013; Svoboda, 2008). Within this area, we distinguished two altitudinal regions: the lowland (between 180 and 230 m a.s.l.) and the hilly region (between 231 and 550 m a.s.l.).
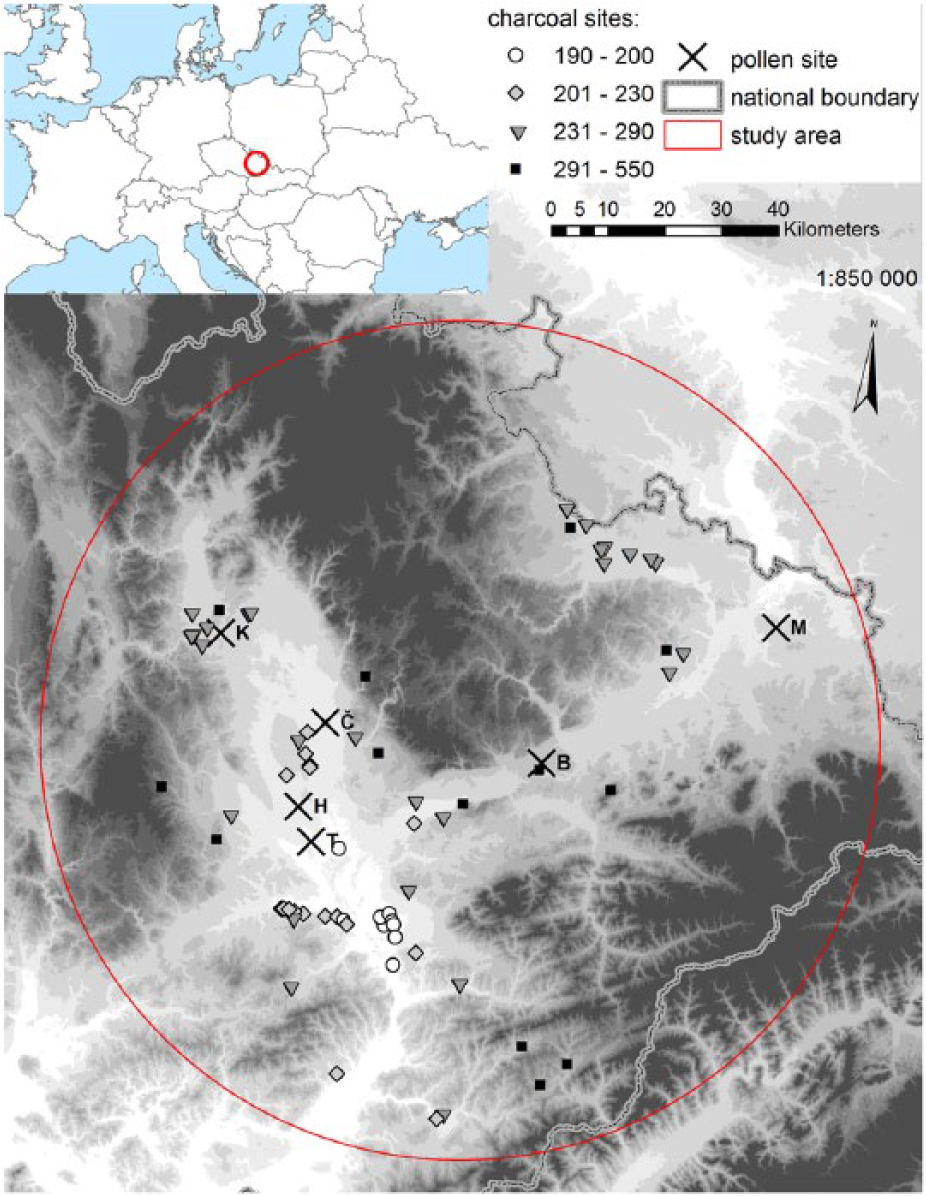
Map of the study area. Anthracological sites are marked by symbols corresponding to Figure 4. Letters by pollen sites mean: T: Tovačov; H: Hrdibořice; Č: Černovír; K: Kačení louka; B: Bělotín; M: Michálkovice; the grey scale follows altitudinal breakpoints: 200, 230, 290, 350, 400, 450, 500, 550 and 600 m a.s.l.
The lowland is characterized by the presence of quaternary sediments such as loess, gravels and alluvial sediments (Czudek, 1972). The most common soils are chernozem, luvisols and fluvisols (AOPK, 2005). The lowland area is part of the Thermophyticum phytogeographic region with a slightly dry and continental Central European climate. The mean annual sum of precipitation is 550–600 mm, and the mean annual temperature is 8.5–9°C (meteorological sites: Kroměříž, Olomouc, Prostějov and Převov; Quitt, 1971). Intense agricultural activity has been affecting the lowland region since mediaeval times, especially after World War II. At present, the lowland region is predominantly used for agriculture, and forests cover only a small percentage of the landscape. Present-day riverine forests belong to the Ulmenion alliance (Chytrý, 2013), and the occurrence of Salix and Alnus is associated with river banks and wetlands. Oak–hornbeam forests (un. Carpinion) are typical of the drier part of the lowland, which does not belong to the Morava river catchment. Plantations of hybrid poplar (e.g. Populus × canadensis) are widespread in the region. As potential natural vegetation, hardwood forests of lowland rivers (un. Ulmenion) and oak–hornbeam forests (un. Carpinion) have been reconstructed in the area (Neuhäuslová, 2001).
Most of the hilly territory lies between 231 and 350 m a.s.l., but several hills rise above 550 m a.s.l. Cambisols and luvisols are the most common soil types in the hilly region. The area is part of the Mesophyticum phytogeographic region. The mean annual sum of precipitation is 600–800 mm, and the mean annual air temperature is 7–8.5°C (meteorological sites: Bělotín, Mošnov and Opava; Quitt, 1971). An oak–hornbeam (un. Carpinion) and beech forest (un. Fagion) has been reconstructed as potential natural vegetation in the area (Neuhäuslová, 2001). The proportion of forested areas is larger than in the lowland region, but forests still cover a smaller part of the landscape. Spruce plantations are commonplace and have been planted in the region since the 19th century.
Anthracological analysis
We assembled a charcoal data set containing our own unpublished charcoal data supplemented with already published older data from the study area (Beneš and Komárková, 2007; Berkovec et al., 2005; Novák, 2008; Novák and Krasnokutská, 2009; Opravil, 1960, 1962, 1965, 1967, 1968, 1972, 1980, 1991, 1993, 1994, 1996, 1997). We included 120 charcoal samples from 69 archaeological sites. Some archaeological sites are polycultural, so several samples in our data set have the same coordinates. If material from a certain archaeological period and site comprised several subsamples, the data were merged into a single charcoal sample. For our study, we chose only samples with at least 30 charcoal records.
A total of 62% of the charcoal samples were separated by flotation (73 charcoal samples), but some older charcoal records were separated by direct picking during archaeological excavations (45 charcoal samples, 38% of samples). Charcoal analysis was performed only on fragments belonging to the largest fraction (>2 mm); smaller pieces that were taxonomically unidentifiable were excluded (e.g. Robin et al., 2014). Our charcoal samples were identified with the help of a reference collection and standard identification keys (Greguss, 1972; Schweingruber, 1978) using an episcopic interference microscope with 200–500× magnification. The charcoals were quantified as the number of analysed fragments. Shrubs were visualized in macro-charcoal diagrams summarizing the following shrub species: Berberis, Cornus, Clematis, Euonymus, Frangula, Juniperus communis, Ligustrum, Lonicera, Prunus, Rhamnus, Rosa and Viburnum. For easier visualization, Salix, Populus and Salix/Populus were merged into one group (Salix/Populus).
For easier interpretation, we divided the anthracological samples according to the archaeological period. The age of most charcoal samples was determined based on an archaeological chronology (e.g. pottery fragments of typical vessels). The 12 periods of recorded archaeological cultures follow Jiráň and Venclová (2013): Linear Pottery Culture (7550–6850 cal. BP), Late Neolithic (6850–6150 cal. BP), Ancient Aeneolithic (6150–5850 cal. BP), Funnelbeaker culture (5850–5350 cal BP), Middle Aeneolithic (5350–4850 cal. BP), Late Aeneolithic (4850–4450 cal. BP), Bell-Beaker culture (4450–4150 cal. BP), Early and Middle Bronze Age (4150–3250 cal. BP), Late Bronze Age (3250–2750 cal. BP), Hallstatt (2750–2350 cal. BP), La Tène (2350–2000 cal BP) and Roman and Migration period (2000–1400 cal. BP). Some charcoal samples were also subjected to 14C accelerator mass spectrometry (AMS) radiocarbon dating in the Poznan Radiocarbon Laboratory, Poland. The dating was calibrated using the default setting of the Clam script (Blaauw, 2010).
Pollen analysis
Pollen analysis was performed on continuous peat sections sampled using piston corers. Six pollen profiles were evenly distributed between the lowland and the hilly region (Figure 1); however, the profiles Černovír and Kačení louka are situated on the upper terrace of the river Morava in the border area between the lowland and the hilly region.
Pollen samples of peat were prepared by standard acetolysis (Moore et al., 1991). From all counted palynomorphs, we considered here only pollen types of trees and shrubs because only these are relevant for our comparison with anthracological results. The sequences were dated using radiocarbon (AMS) measurements performed on macro-fossils of terrestrial plants and charcoals. The radiocarbon dating was performed in the Poznan Radiocarbon Laboratory, Poland (pollen profiles Hrdibořice, Kačení louka, Michálkovice and Tovačov), Center for Applied Isotope Studies, University of Georgia, Athens, US (pollen profile Černovír) and AMS-14C Labor Erlangen, University of Erlangen, Germany (pollen profile Bělotín; Appendix 1). The calibration and chronologies dating all pollen samples were established using the software Bacon (Blaauw and Christen, 2011).
Statistical methods
First, in order to compare pollen and anthracological signal visually, we calculated mean percentage abundance of each taxon per archaeological period in both the charcoal and pollen record. We plotted pollen percentage curves from all sites over each other (R Development Core Team, 2013) to visualize the variability and density of sampling at each site. Similarly, we used separate boxplots for anthracological records from sites in the lowland and in the hilly region.
Second, we tested the quality of anthracological sampling at different altitudinal zones. The area of each altitudinal zone was obtained from the SRTM Digital Elevation Model (Jarvis et al., 2008) with a mean resolution of 6573 m2 per grid cell. Pearson’s correlation coefficient was calculated between the area of each 50-m wide altitudinal zone and the number of anthracological samples in different periods.
Third, we used three multivariate statistical analyses implemented in CANOCO v. 5 (ter Braak and Šmilauer, 2002) to explore the two main factors in the anthracological data sets: altitude and age of samples. Logarithmic transformation of the percentage data and centring by species was used for all ordinations. A preliminary detrended correspondence analysis (DCA) showed the length of the gradient to be 2.03, so a principal component analysis (PCA) was performed on the whole data set. For the purposes of the direct technique, we excluded samples from the altitudinal zone whose number did not correspond to its area and subsequently tested how much of the variability in the charcoal data set could be explained by altitude and age of the sample using redundancy analysis (RDA).
Results
Anthracological results and woodland history
The anthracological analysis revealed 31 charcoal taxa: Abies, Acer, Alnus, Berberis, Betula, Carpinus, Cornus, Corylus, Clematis, Euonymus, Fagus, Fraxinus, Frangula, Juniperus, Ligustrum, Lonicera, Pinus, Picea, Pomoideae, Populus, Prunus, Rhamnus, Rosa, Salix, Sorbus, Taxus, Tilia, Vaccinium, Viburnum and Viscum. Our study analysed 42,547 charcoal records in 120 charcoal samples from 69 sites. The number of anthracological samples per altitudinal zone area significantly correlated with altitudinal zone area only if we exclude the lowest zone (Figure 2, Table 1). We obtained a higher density of charcoal samples from the lowest altitudinal zone 150–200 m a.s.l. (1 sample/20 km2) compared with the general density of charcoal samples in the study area 150–550 m a.s.l. (1 sample/111 km2; Table 1).

Sampling of the study area in the selected altitudinal range: (a) each plus symbol represents a pollen sample, (b) the size of each circle represents the number of determinations per anthracological sample and (c) areas of different altitudinal zones.
Coefficients of correlation between the area of altitudinal zones and the number of charcoal samples from each archaeological period.
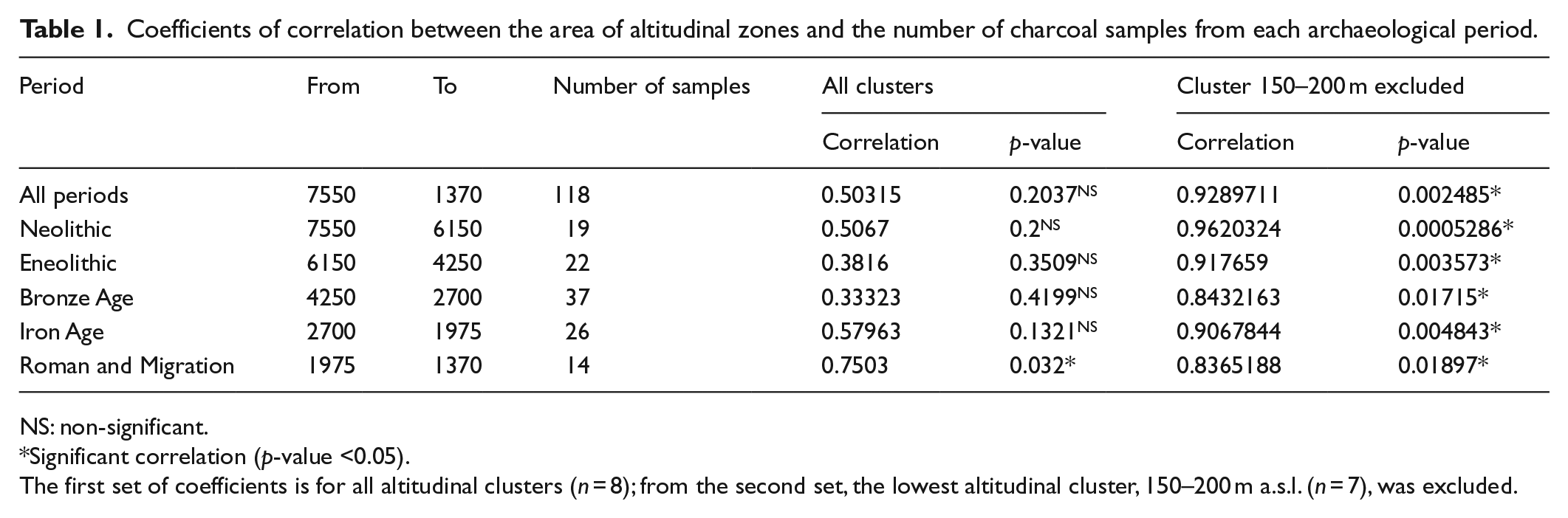
NS: non-significant.
Significant correlation (p-value <0.05).
The first set of coefficients is for all altitudinal clusters (n = 8); from the second set, the lowest altitudinal cluster, 150–200 m a.s.l. (n = 7), was excluded.
The lowland region was characterized by a stable continuity of a species-rich deciduous forest with a dominance of Quercus, a very common presence of Fraxinus and Ulmus and a less common presence of Corylus, Acer and Pomoideae during the whole period under study (Figure 3). The occurrence of Fagus and Abies was recorded since the Linear Pottery culture (7500–6800 BP), but their abundance was low throughout the period under study. Carpinus charcoals were recorded since the Neolithic period, and their first higher occurrences were documented in the southern part of the region. The presence of Tilia charcoals was generally low during the study period. Pinus charcoals were documented sporadically, and their slightly higher abundance was also recorded during the Bronze Age period. Alnus and Populus/Salix charcoal was commonly recorded throughout the period under study, but their abundance was lower. Picea charcoals were very rare and related only to the Neolithic period. The species composition changed relatively little in the lowland region during agricultural prehistory (7500–1500 BP). Since the Neolithic period, Fraxinus excelsior and partly also Ulmus show a gradual decrease. However, Carpinus betulus and Betula increased since the Bronze Age period. The common presence of the shrubs was recorded since the Aeneolithic period (5600 BP).

Percentages of taxa in anthracological samples (boxplots) and mean anthracological curves (polygons) for the lowland and the hilly region. Ticks on the age BP axis indicate the middle year of each archaeological period.
The charcoal record from the hilly region (231–550 m a.s.l.) is outstanding due to the presence of a species-rich forest with a high dominance of oak during the Neolithic–Aeneolithic period (7500–4000 BP). The significant abundance of Corylus charcoal in this period are probably connected with local vegetation history in the vicinity of archaeological sites. Since the Bronze Age period (4000 BP), the species-rich oak forest was gradually replaced by fir–beech and oak–hornbeam forests. The abundance of Fagus and Abies increased mainly in the upper part (290–550 m a.s.l.), whereas Carpinus increased in the lower part (230–290 m a.s.l.) of the hilly region (Figure 4). In this period, the presence of Betula and Acer charcoal slowly increased. The presence of light-demanding Corylus increased during the Iron Age and the Roman period. Charcoals Pinus, Picea and Tilia occurred very rarely over the period under study. The occurrence of Alnus, Populus/Salix, Pomoideae and shrub charcoals was common, but their abundance was low.
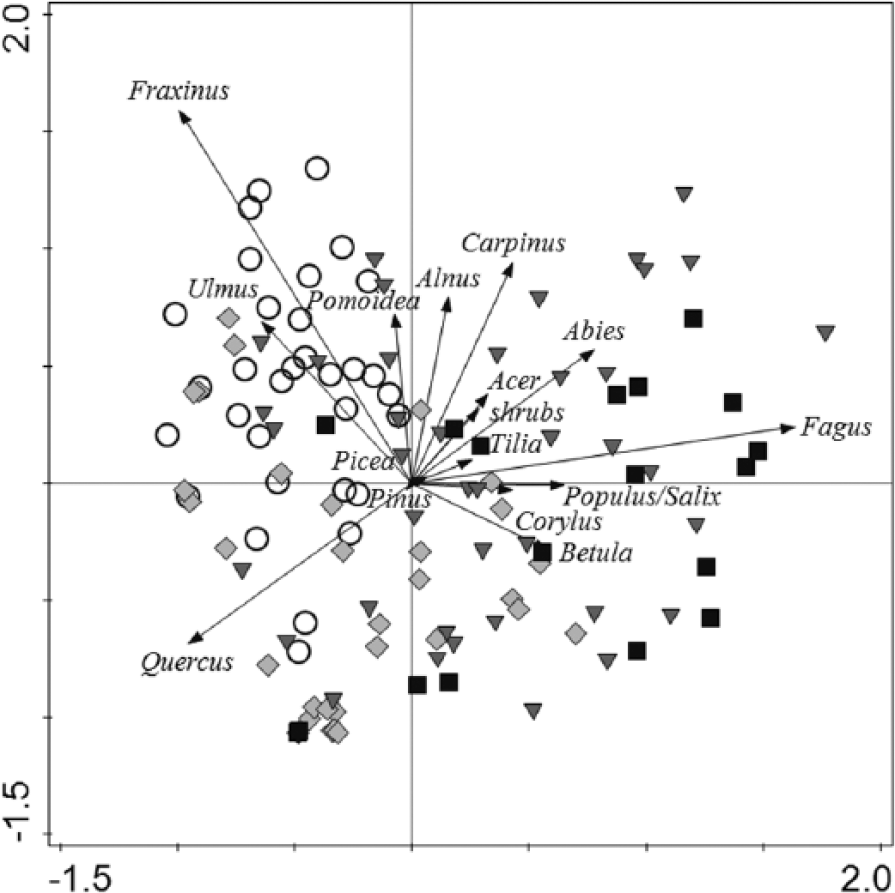
Relationship between charcoal taxa, samples and altitude.
Relationship between charcoal samples and regions
We visualized the relationship between charcoal samples and species in the lowland and in the hilly region in an indirect ordination (PCA) diagram (Figure 4). The lowland region was characterized by the presence of Quercus, Fraxinus and Ulmus, whereas the hilly region was characterized by a connection with Fagus, Carpinus and Abies.
A comparison of samples from within the lowland region revealed differences in species composition between the lower and upper parts of the region. The charcoal records from the lower part of the lowland (up to 200 m a.s.l.) showed a correlation with Fraxinus and Ulmus, whereas sites from the upper part of the lowland (200–230 m a.s.l.) correlated with the high abundance of Quercus. A key characteristic of the upper part of the hilly region (290–530 m a.s.l.) was a higher abundance of Fagus, but an increase in Abies abundance was recorded in the lower part of the hilly region.
Samples from the lower hilly region showed the highest variability. The lower lowland region, by contrast, was relatively uniform and specific compared with the rest of the altitudinal zones. To test the effect of altitude and time in direct ordination (Figure 5), we decided to exclude samples from the lowest altitudinal zone.
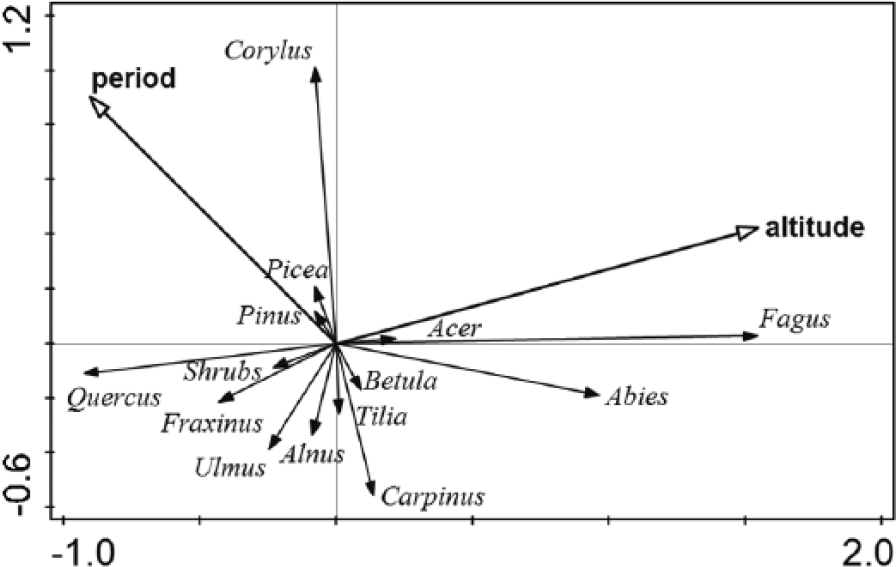
Redundancy analysis (RDA) without samples from the lowest altitudinal zone 190–200 m a.s.l. (p = 0.001, F = 4, 5).
Considering the even altitudinal sampling, the general variability in our data set reflected the altitudinal rather than the temporal gradient because the altitudinal arrow was longer and closer to the main axis. Since the arrows representing both environmental variables pointed in a perpendicular or slightly obtuse angle, altitude and time were not correlated or perhaps slightly negatively correlated. With increasing altitude, the abundance of Fagus and Abies increased whereas that of Quercus decreased. Older samples tended to contain more Corylus charcoals, and younger samples tended to contain more Carpinus, Fagus and Abies charcoals. Both dominants of the lowest altitudes, Ulmus and Fraxinus, were negatively correlated with altitude.
Pollen record
The sampling of the pollen record was relatively sparse, especially in the middle-Holocene; however, we still obtained almost one sample per each 500-time window in the lowland and in the hilly region. Variability of pollen percentages was high when considering adjacent time intervals within sites and also between different sites within one time interval (Figure 6). Records from more than one site for both data sets were available only for four time intervals from 4000 to 2000 BP. In this period, mean pollen percentage curves for most of the taxa were fuzzy, so it was impossible to decide in which altitudinal zone given tree species prevailed. Abies, Tilia and Alnus reach higher values in the hilly region, but one of the lowland sequences also fell among sequences from higher altitudes. Only Picea showed a clear difference between both mean pollen curves, which could be attributed to the altitudinal gradient rather than high variability. Mean percentages of Picea pollen were higher (around 20%) in the lowland than in the hilly region (around 5%).

Percentages of taxa in pollen samples (lines) and mean pollen curves (polygons) for the lowland and the hilly region. Ticks on the age BP axis indicate the middle year of each archaeological period.
Comparison of the anthracological and pollen record
Based on their mean values in the pollen and anthracological records, we divided the taxa into three groups. The first included trees, which were represented more in pollen than in charcoals: Pinus, Picea, Tilia and Alnus. Taxa in the second group were represented by similar values of both proxies: Betula, Corylus and Abies. The third group comprised Quercus, Fraxinus, Ulmus, Acer, Fagus, Carpinus and shrubs, taxa which were represented more in charcoals than as pollen (Figure 7).
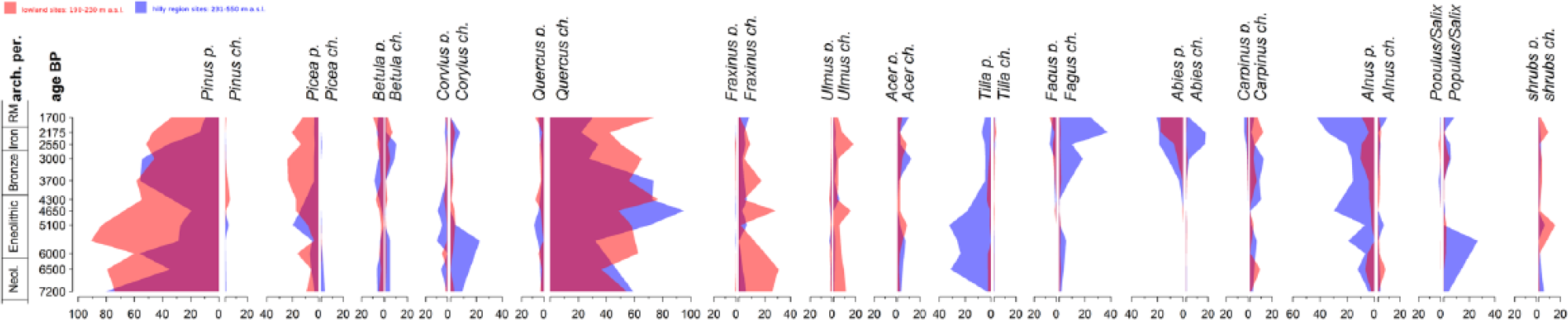
Comparison between the mean pollen (left) and the mean anthracological curve (right) for the lowland and the hilly region. Ticks on the age BP axis indicate the middle year of each archaeological period.
This mismatching representation hampered any comparison of the pattern between the two proxies over time. The slightly decreasing trend of pine pollen, for example, was not visible in the charcoal record because the presence of pine charcoals was very low. The only exceptions were Fagus, Abies and Carpinus, which appeared in the pollen record from the Older Bronze Age. This increase was visible also in the charcoal record in the cases of Fagus and Abies, whereas Carpinus showed a relatively stable presence during the period under study. Regarding the first appearance, both Fagus and Abies were detected much earlier by anthracological analysis, even though their abundance was documented only in low quantities during the Neolithic and Aeneolithic periods. The magnitude of the pollen signal of Abies was slightly higher than in the charcoal record; however, charcoal analysis seems to be more sensitive in detecting the first appearance. Abies was detected in the lowland by similar pollen signal as in the hilly region, but the charcoal analysis did not prove any presence of Abies at lowland sites.
Discussion
Anthracological analysis and reconstruction of a woodland vegetation
Many ethnoanthracological studies (e.g. Biran et al., 2004; Henry and Théry-Parisot, 2014; Türker and Kaygusuz, 1995; Zapata-Peña et al., 2003) show that domestic firewood gathering was an everyday activity and that the majority of charcoals found at archaeological site originated from common firewood. According to the principle of least effort (Prior and Price Williams, 1985; Shackleton and Prins, 1992), firewood gathering took place in the vicinity of archaeological sites, all woody species were collected in a non-selective manner, and the proportion of species was analogical to that occurring in the environment. Some studies assume that the area of wood collection around an archaeological site was equal to the area that could be reached in 1 h on foot (e.g. Beneš, 2004; Petrlíková and Beneš, 2008). Other studies, however, assume the area of firewood collection depended on the geomorphological situation, the size and importance of the archaeological site (e.g. Asouti and Austin, 2005; Novák et al., 2012; Théry-Parisot et al., 2010) and also reflected the social organization of communities and their way of life (Biran et al., 2004; Zapata-Peña et al., 2003). The effects of post-depositional processes and different methods of sampling and quantification on charcoal assemblages have been studied by anthracologists and sedimentologists (e.g. Carcaillet and Talon, 1996; Figueiral and Mosbrugger, 2000; Fréjaville et al., 2013). Although charcoal records from archaeological sites are the results of a very complex processes, every anthracological assemblage is inevitably affected by the kind of forest that surrounded the archaeological site. First, farmers throughout agricultural prehistory situated their settlements close to the most fertile soils. Second, forests in the surroundings of settlements were influenced by humans. We assume that the high signal of Quercus and Carpinus possibly reflects only a certain part of the forest mosaic that was settled. The density of settlements and the anthropogenic impact nevertheless could have affected a significant part of the prehistoric lowland landscape (Čižmár et al., 2000; Čižmár and Geislerová, 2006; Čižmářová et al., 1996). A model of human activity in the lowland area by the southern border of our study region suggests that 20% of the landscape was affected by humans during the Neolithic, which increased abruptly to 40% during the Late Eneolithic and thereafter fluctuated slowly until the end of the Roman period (Kuneš et al., 2015). From the anthracological perspective, the composition of vegetation in the rest of the landscape might remain unknown. The impact of permanent pastoralism or forest management practices possibly eliminated trees that do not re-grow easily from stumps. This interpretation fits with our abundant charcoal evidence of nutrient-demanding broadleaved trees and, especially, shrubs, which are resistant to prehistoric human impact. Detailed archaeological data and vegetation estimates for the past of Southern Moravia, moreover, indicate that the spread of Quercus was triggered by consequences of human activity (Kuneš et al., 2015).
Comparison between charcoal and pollen analysis
Considering the bias of human impact and selective wood collection in the anthracological record, we can use the charcoal record to interpret the pollen record because the charcoal record provides spatially more precise site-related information on species occurrence. Pollen assemblages are influenced by pollen productivity, pollen dispersal, pollen taphonomy and spatial structure of source plants around the pollen site. Regarding the pollen taphonomy and spatial structure of source plants, pollen spectra from any natural archive are theoretically composed of a local and a regional components. At small sites surrounded by a closed canopy, signal from local vegetation in the vicinity prevails; at large bogs, lakes or even small bogs in completely open landscapes, by contrast, the local component is negligible (Bjune et al., 2009; Nielsen and Odgaard, 2005; Sugita, 2007a, 2007b).
The high pollen signal of Pinus is caused by high pollen production and easy dispersal of bisaccate pollen. However, Picea pollen productivity, according to direct (Pohl, 1937) and indirect measurements (Soepboer et al., 2007; Abraham et al., 2014), is 2.5–4 times lower than that of pine, so the high pollen percentage of Picea is quite unexpected, especially since lowland sites show a clearly higher mean than the hilly region. The high pollen signal of both conifers contrasts with their very sparse occurrence in the charcoal record; spruce is present only at three sites. This disagreement can be explained by large populations of both trees at higher altitudes, that is, above 550 m a.s.l., which are relatively close to the pollen sites and surround the elongated lowland. This interpretation that Pinus and Picea were rare in the Moravian lowland and hilly region (190–550 m a.s.l.) is supported by frequent and abundant Pinus charcoal from archaeological sites in adjacent Bohemia (e.g. Kočár et al., 2014; Novák et al., 2015). This shows that Pinus is not under-represented due to firewood preferences and that it could be abundant due to human disturbances of soils in Bohemia.
Tilia and Alnus are over-represented in the pollen record due to their growth in the vicinity of pollen profiles. Alnus is a typical wetland tree growing within the site. A strong signal of Tilia was detected in the pollen record at the small wetland locality Kačení louka. The high abundance of Tilia pollen is probably related to the character of local vegetation on groundwater-influenced soils at margins of a small wetland.
Abies was recorded in the hilly region by charcoals and pollen, but Abies in the lowland is abundant in the pollen record, but its charcoals are almost lacking (Figure 7). This difference in the lowland is given by the high signal of Abies at the Hrdibořice site (Figure 6), which we attribute to limited pollen dispersal from a local population close to the site. Since Abies has the heaviest pollen grains out of the set of taxa, its source area of pollen is small and thus the most comparable with the source area of charcoals. The appearance of Abies pollen at the Hrdibořice site is analogous to sporadic non-zero occurrences of Abies in the lowland charcoal record (Figure 3).
To sum up the main differences between pollen and charcoal analysis, the pollen record reflects taxa on a regional scale if pollen productivity, dispersion or vegetation abundances are high, and at the same time, it also reflects local taxa growing at a site or in its surroundings. However, the charcoal record from archaeological sites can document the first appearance of a species much earlier, especially if it can tolerate human influence. For example, a common presence of Carpinus charcoals has been recorded at archaeological sites of the Linear Pottery culture (7600–6800 BP). The pollen record in our study area, by contrast, does not document hornbeam until the Bronze Age period (4000 BP). Scarce findings of Carpinus charcoals have been documented from the Mesolithic age in southern Moravia (Opravil, 1983a). Macro-fossils such as nutlets and wood have been found in layers corresponding to the Atlantic climatic optimum (Opravil, 1983b, 1984). Carpinus pollen (6%) has nevertheless been detected in sediments dated to the Early Atlantic (8240 cal BP) in the neighbouring region of South Moravia (Doláková et al., 2010). The first appearance of Carpinus in the SE Carpathians and in the North Hungarian Middle Mountains is recorded around 8500 cal BP, and their role as an important woodland element is recorded from about 7500 cal BP (Magyari, 2002). The expansion of Carpinus in Central-Eastern Europe is often connected with anthropogenic activity (e.g. Gardner, 2002; Küster, 1997; Ralska-Jasiewiczova et al., 2002). Important changes in the landscape during the Bronze Age period induced an expansion of Carpinus, as evidenced by our pollen analysis.
The further potential and limitations of pollen and charcoal analysis will depend on the availability of data for both proxies. The advantage of the pollen record that it is continuous is limited in our study by the presence of hiatuses and low sediment accumulation during the middle-Holocene. According to the Macrophysical Climate Model, this period was characterized by a drier and warmer climate (Kuneš et al., 2015). In contrast to pollen profiles, archaeological sites in our study area are plentiful, and charcoals are common in archaeological layers.
Woodland history across the altitudinal gradient
The results of our study draw attention to the noticeable differences in anthracological composition along the altitudinal gradient, and there is a significant pattern. Differences between the altitude zones are relatively small, but the vegetation of the regions is influenced by different environmental conditions (e.g. climate, soil and geomorphology).
Indirect ordination analysis of samples from the lowland region (Figure 4) shows important differences in species composition between the lower and upper parts of the lowland region. Alluvial sediments are widely distributed in the Morava river valley (mainly up to 200 m a.s.l.), so charcoal samples from the lower part of the lowland are characterized, besides the presence of Quercus, by a higher abundance of nutrient-demanding deciduous trees such as Fraxinus, Ulmus and Acer, and hygrophilous tree species such as Alnus. The localities of these charcoal samples are usually situated in the vicinity of the Morava river floodplain. The character of charcoal species composition is analogous to present-day riverine forests (un. Ulmenion). Our anthracological results support the hypothesis of a long-term presence of diverse woodland vegetation with high abundance of resource-demanding broadleaved trees (Fraxinus, Ulmus and Acer). From this perspective, recent hardwood forests along lowland rivers can be considered the successors of Atlantic species-rich oak forests (Quercetum mixtum). It must be remembered that woodlands in the Atlantic period did not have to have the same pattern as recent woodlands in Central Europe, but the species composition is almost analogical. This contradicts the generally accepted opinion that alluvial forests are modern plant communities formed by floodplain evolution in connection with mediaeval and postmediaeval human activities (e.g. Kadlec et al., 2009; Pokorný, 2011). Similar woodland vegetation was also recorded during the early and high mediaeval period at archaeological sites in the vicinity of river Morava (Opravil, 1983b). A high abundance of Fraxinus and Ulmus charcoals was recorded at riverine Neolithic and Aeneolithic archaeological sites in the northern and north-eastern Pannonian basin (Lityńska-Zajac et al., 2008; Moskal-del Hoyo, 2013). We assume that in prehistory, as in the present, hardwood forests (mixed oak–elm–ash forests) were the main type of forest vegetation in the surroundings of lowland rivers in Central Europe. We assume that the presence of willow-poplar forests was restricted to the vicinity of oxbow wetlands, river banks and riverbeds.
Upper parts of the lowland (200–230 m a.s.l.) are covered by loess and chernozem soil. The charcoal spectra are dominated by Quercus, whereas other species are represented less frequently throughout agricultural prehistory. Some studies (e.g. Eckmeier et al., 2007; Vysloužilová et al., 2014) suggest that chernozem soils are able to retain their characteristics under open deciduous woodlands. It is difficult to reconstruct woodland openness from the anthracological record. The frequent occurrence of shrubs (e.g. Cornus sp.) suggests the existence of open-canopy woodlands (Moskal-del Hoyo, 2013). A higher abundance of shrubs can be explained by anthropogenic influence on forest vegetation (Marinova and Thiébault, 2008). We interpret our results with regard to general ecological characteristics of oak. The growth of oak seedlings under a closed canopy is limited (Vera, 2000), so oak woodlands are generally semi-open, and Quercus is characterized as a light-demanding tree. Moreover, the vicinity of the archaeological context indicates human impact that might maintain a certain degree of anthropogenic openness. We interpret the charcoal assemblage in the chernozem area as an open oak woodland, parkland or shrubland with low canopy density. Similarly, a dominance of oak charcoals is documented at archaeological sites in NE Hungary (Moskal-del Hoyo, 2013), and surrounding vegetation is also reconstructed as forests with a more open canopy.
Our study documents the scarce presence of Fagus and Abies charcoals in the lowland region since the Linear Pottery culture (7500–6800 BP), but their abundance was low throughout the period under study. Slovenia and the eastern Alps, and possibly southern Moravia, should be considered the main source areas for the colonization of Central and Northern Europe by beech (Magri, 2008). Poschlod (2015) supposed that the spread of Fagus and Abies in the Central European lowlands was limited due to human impact. Neither of the trees can re-grow easily from the stump if cut down, so their low abundance in the lowland can be connected with long-term woodland management (e.g. coppicing). However, Carpinus is very tolerant to such forestry practices. In the lowland of central Moravia, we can find Carpinus charcoals from the time of the Linear Pottery culture (7500–6800 BP) more frequently than charcoals of Fagus and Abies, but their abundance sharply increased in the Bronze Age period (4000 BP). In Europe, the Bronze Age marked major changes in the organization of society and saw the rise of decentralized societies with supraregional economies based on metals (e.g. Kristiansen and Larsson, 2005). The increasing human impact during the Bronze Age is visible in the growing rate of erosion in regions covered by loess (e.g. Petřík et al., 2015). Many palaeoecological studies from Central and Northern Europe identified human activities as the dominant driver of vegetation change between 4000 and 3500 BP (e.g. Odgaard and Rasmussen, 2000; Pokorný et al., 2015). Similarly, the Bronze Age archaeological sites in our study area extend into higher altitudes (Čižmářová et al., 1996).
The charcoal record in the hilly region during the Neolithic–Aeneolithic period (7500–4000 BP) originates mainly from its lower part 230–290 m a.s.l. (Figure 2). Thus, the high presence of Quercus and common spread of Corylus are similar to the upper part of lowland region. During the Bronze Age, changes in species composition were accompanied by an increase in the abundance of Carpinus, Fagus and Abies and a decrease in the abundance of Quercus. Carpinus was more abundant in the lower part of the hilly region, and Fagus was more abundant in the upper part of the hilly region. The expansion of Fagus has frequently been connected with farming and other human activities, which influenced the forest canopy and increased the rates of forest disturbance (Andersen, 1974; Björkman, 1997; Küster, 1997; Latałowa, 1992). The first more pronounced pollen evidence of human impact on European mountains usually comes from the Bronze Age (e.g. Kozáková et al., 2015; Sköld et al., 2010). Bronze Age human activities have also been recorded in the highest parts of the Jeseníky Mountains (Novák et al., 2010), which are adjacent to the study area. However, the expansion of Fagus was also associated with its Holocene spread from glacial refugia, natural processes (e.g. the Iversen cycle, climatic changes or wildfire events) or some combination of these factors. The Bronze Age expansion of Fagus was also recorded in numerous pollen profiles from surrounding mountain ranges (e.g. Dudová et al., 2010, 2013) and in slightly later profiles from the South Moravian Lowland (Kuneš et al., 2015). The Fagus expansion at the interface between the Carpathians and northern Pannonia started at a similar time, between 5500 and 4000 cal. yr BP (e.g. Bodnariuc et al., 2002; Feurdean, 2005; Hájková et al., 2013; Petr et al., 2013; Rybníček and Rybníčková, 2008).
Conclusion
Pollen analysis alone revealed vegetation changes on an unknown spatial scale. However, with the help of our charcoal record, we could gain some idea about the spatial distribution of selected species, even though the sampled woodland was influenced by previous human impact and the selection of settlement sites. Additionally, we found that abundant charcoal samples from many sites and periods can rectify the generally discontinuous character of the charcoal method.
The results of our study underline that combining pollen and charcoal analysis has great potential for studying phenomena in cultural landscapes since each of the methods approaches nature from the opposite side of the human–nature gradient.
We reconstructed the middle- and upper-Holocene woodland history of central Moravia and revealed important differences along an altitudinal gradient. Lower parts of the lowland region (mainly up to 200 m a.s.l.) are characterized by a higher abundance of nutrient-demanding deciduous trees. Our anthracological results support the hypothesis of the long-term presence of diverse woodland vegetation. A hardwood forest (mixed oak–elm–ash forest) was the main type of forest vegetation in the surroundings of lowland rivers during middle- and upper-Holocene history.
Striking dominance of Quercus charcoal was recorded in the upper part of the lowland region (200–230 m a.s.l.) throughout agricultural prehistory. Oak woodlands are generally semi-open, and Quercus is characterized as a light-demanding tree. We reconstructed woodland vegetation in the vicinity of archaeological sites in the chernozem soil region as being a woodland with a more open canopy.
We traced an analogous charcoal record in the lower part of hilly region (230–290 m a.s.l.) during the Neolithic–Aeneolithic period (7500–4000 BP). Nevertheless, during the Bronze Age period, we can trace an increase of Carpinus, Fagus and Abies accompanied by a decrease in Quercus abundance. The abundance of Fagus was noticeably higher in the upper part of the hilly region (290–540 m a.s.l.). Numerous palaeoecological studies from Central and Northern Europe document a similar increasing human impact during the Bronze Age. Human activities have become a strong driver of vegetation change in lowlands and hilly regions of Central Europe.
Footnotes
Appendix
Result of 14C dating (AMS method) from the pollen profiles studied.

| Laboratory code | Pollen profile | Depth (cm) | 14C age (BP) |
|---|---|---|---|
| Erl-7652 | Bělotín | 60 | 857 ± 47 |
| Erl-7627 | Bělotín | 100 | 2683 ± 78 |
| Erl-7653 | Bělotín | 150 | 3476 ± 47 |
| UG-13600 | Černovír | 61 | 7900 ± 30 |
| UG-13601 | Černovír | 189 | 9220 ± 25 |
| UG-13602 | Černovír | 205 | 10200 ± 170 |
| Poz-62219 | Hrdibořice | 95 | 2745 ± 30 |
| Poz-65370 | Hrdibořice | 147 | 8040 ± 50 |
| Poz-65371 | Hrdibořice | 211 | 9170 ± 50 |
| Poz-62221 | Hrdibořice | 240 | 9650 ± 60 |
| Poz-62220 | Hrdibořice | 307 | 10070 ± 60 |
| Poz-65463 | Kačenní louka | 30 | 205 ± 30 |
| Poz-62207 | Kačenní louka | 63 | 1605 ± 30 |
| Poz-62206 | Kačenní louka | 100 | 3365 ± 35 |
| Poz-65461 | Kačenní louka | 129 | 7310 ± 50 |
| Poz-62205 | Kačenní louka | 145 | 9230 ± 50 |
| Poz-20323 | Michálkovice | 100 | 740 ± 30 |
| Poz-65459 | Michálkovice | 180 | 2140 ± 35 |
| Poz-20491 | Tovačov | 20 | 2415 ± 30 |
| Poz-20492 | Tovačov | 35 | 2700 ± 30 |
| Poz-20509 | Tovačov | 55 | 2530 ± 35 |
| Poz-20493 | Tovačov | 75 | 3220 ± 35 |
| Poz-20494 | Tovačov | 115 | 8720 ± 50 |
| Poz-20495 | Tovačov | 180 | 7320 ± 40 |
| Poz-20572 | Tovačov | 270 | 12220 ± 70 |
Acknowledgements
The authors are thankful to Tomáš Bešta for his assistance with profile sampling and Fred Rooks for language corrections.
Funding
Our research was supported by grant GAČR 13-11193S from the Czech Science Foundation. V. Jankovská thanks the support received as part of project RVO 67985939.