
Abstract
European beech and Norway spruce are late successional tree species that have become rapidly dominant in northern Europe in late-Holocene. The northern distribution limit for natural beech forests is in SE Norway, where beech forests and boreal spruce forests meet. Here we have estimated the size, composition and age of the macroscopic charcoal pool to infer past fire history and the establishment of neighbouring Norwegian beech- and spruce forests. To encompass landscape level scales of variations in the charcoal pool, we have analysed the charcoal record in 100 soil cores that were collected using a restricted random procedure. The sizes of the soil charcoal pools ranged from 2 to 1214 g m−2, and they were significantly more spatially variable in the beech forest landscape than in the spruce forest landscape. We show that today’s beech forests took over the dominance from Norway spruce in the landscape about 300 years ago, and that fire disturbances on the landscape level preceded the establishment of beech in the former spruce forest landscape. Interestingly, large-scale fire disturbances have not occurred ever since beech gained dominance. Conversely, we show that today’s spruce forests took over the dominance from Scots pine in the landscape, and that also the establishment of spruce was preceded by fire disturbances on the landscape level.
Keywords
Introduction
Millions of hectares of boreal forests burn each year (Soja et al., 2007) and recurring fire is, and has always been, a key disturbance factor and driver of tree species composition in the boreal forest (Bonan and Shugart, 1989; De Lafontaine and Payette, 2011; Zackrisson, 1977). Charcoal, which is produced when forests burn, constitutes a vital part in boreal forest soils as it functions as a booster of ecosystem productivity (Zackrisson et al., 1996) and a long-term storage of recalcitrant and non-cycling carbon (Ohlson et al., 2009; Santín et al., 2016). Moreover, as charcoal can stay in boreal forest soils for thousands of years, it also functions as a historical archive for forest fire activity in a long-term Holocene time perspective (De Lafontaine and Asselin, 2011; Gavin et al., 2003; Ohlson et al., 2011). Given the importance of soil charcoal in terms of ecosystem productivity, carbon dynamics and fire history, it has received considerable research interest in recent decades (e.g. Clark et al., 1997; Lehmann et al., 2011; Preston and Schmidt, 2006; Santín et al., 2016; Scott, 2010) and numerous studies have explored the soil charcoal pool to estimate its size and to unravel relationships between past fire activity and contemporary forest structure and composition (see Kuosmanen et al., 2014; Ohlson et al., 2009, 2011; Touflan et al., 2010 and references therein). However, the distribution of charcoal in boreal forest soils appears to be extremely patchy, also across very fine spatial scales (Kasin et al., 2013; Ohlson et al., 2013; Preston, 2009), and there is still considerable uncertainties about the size and age of the boreal forest charcoal pool and if, and how, it relates to the composition of today’s forests.
European beech (Fagus sylvatica L.) and Norway spruce (Picea abies (L.) Karst.) are late successional tree species, which both have become rapidly dominant in northern Europe in late-Holocene (Giesecke et al., 2007; Seppä et al., 2009). Climate is considered to control the continental scale migration and establishment of these two species (Björkman and Bradshaw, 1996), but disturbances prior to their stand establishment success are pointed out as the main reason for their rapid dominance on the local scale (Lindbladh et al., 2000). A range of disturbances are known to be of importance in this context, and examples of controlling disturbance factors are fire and anthropogenic activities such as agriculture (e.g. forest grazing) and selective tree cutting (Bradshaw and Hannon, 1992; Lindbladh et al., 2000, 2008). Even though the spread and establishment success of both beech and spruce have been favoured by disturbances, there appears to be differences between the two species, that is, spruce spread as a migrating front tracking areas of regional climate, to invade and establish in forests on rather poor soils, while the spread and establishment of beech seems to have been more directly linked to anthropogenic activities and disturbance by fire prior to stand establishment on richer forest soils (Bradley et al., 2013; Bradshaw and Lindbladh, 2005). In addition, modern forest management practices have favoured both species during the two last centuries (Brunet et al., 2012; Lindbladh et al., 2014). Today, European beech forests reach their natural northern distribution limit in southeastern Norway (Bjune et al., 2013), where they meet boreal forests dominated by Norway spruce. In the future, on the other hand, beech is expected to expand its northern distribution range on the expanse of spruce because of climate warming (Hickler et al., 2012). This in turn implies severe future competition between beech and spruce in northern Europe.
In this study, we aim at quantifying the size of the soil charcoal pool and estimate its age and composition in neighbouring beech and spruce forest landscapes in southeastern Norway. As fire disturbance is supposed to be more important for the establishment success of beech than for spruce (Bradshaw and Lindbladh, 2005), we hypothesize that the charcoal pool should be larger and younger in beech forest soils than in spruce forest soils. Moreover, we hypothesize that the charcoal pool in today’s beech forest landscape should be characterized by particles from former tree species that dominated the landscape prior to the beech forest. Finally, as the beech forest landscape in our study is characterized by a more rugged terrain than the spruce forest landscape, we expect that the spatial variation in the size of the soil charcoal pool should be largest in the beech forest landscape.
Materials and methods
Study sites
Our study area is located in the southern boreal vegetation zone (Moen, 1999) and belongs to the Oslo Rift geological area in southeastern Norway (Figure 1a). The beech forest landscape, Brånakollane (59.19 N, 10.05 E; c. 200 m a.s.l.), is situated above the marine limit and is a nature reserve with natural and well-developed beech forests covering c. 19 ha (Bjune et al., 2013). The terrain topography is hilly with monozonite quartz rocks and outcrops (Geological Survey of Norway, 2016), and the soil cover, which consists of brown soil, is generally thin. Typical features are that the ground is covered by thick layers of fallen beech leaves, and that the forest ground vegetation is very sparse, consisting mainly of Oxalis acetosella, Anemone nemorosa, and a couple of grass- and fern species. We used old forest maps, produced by the Fritzøe forest property in 1896 (see Lie et al., 2012), to locate a neighbouring spruce forest landscape with long spruce forest continuity and mature tree stands (59.17 N, 10.05 E; c. 100 m a.s.l.). This landscape is located 2 km south of Brånakollane and below the marine limit. The bedrock consists of syenite quartz and the terrain is less rugged as compared with that in the beech landscape (Figure 1b). The soils are weakly podzolized and Norway spruce is clearly dominating in the tree layer, but a few specimens of beech, Scots pine Pinus sylvestris and pedunculate oak Quercus robur are also present in the tree layer. The ground vegetation is lush and feathermosses, in particular Pleurozium schreberi and Hylocomium splendens are abundant. Polytrichum and Sphagnum mosses are also common. Bilberry Vaccinium myrtillus dominates among the vascular plants in the ground vegetation.
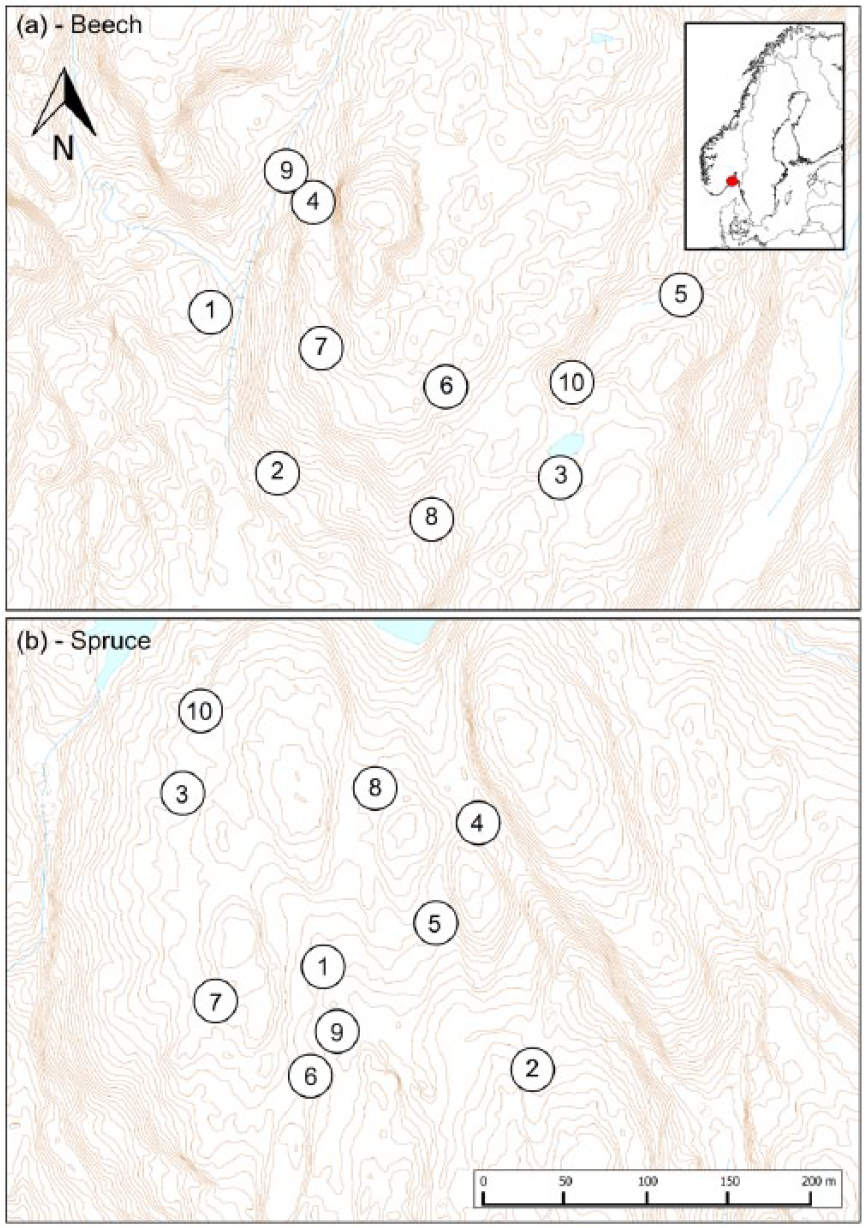
Location of the study area in SE Norway (insert) and positions of sample plots (10 × 5 m2) in (a) the beech and (b) spruce forest landscapes. Elevation contours are 1 m and note the rugged terrain relief in the beech forest.
There has been no significant forestry activity in the Brånakollane beech forest landscape since 1837 (Bjune et al., 2013). However, prior to that, it is very likely that forest structure and composition have been impacted by human land-use, in particular forest grazing and use of fire to improve grazing quality, which both activities are known to have a long history in these parts of Norway (see, for example, Blanck et al., 2013; Storaunet et al., 2013). The same kinds of historic anthropogenic impacts are also likely to have occurred in the spruce forest landscape.
Soil sampling and charcoal analysis
A total of 10 sampling plots of 5 × 10 m2 were placed subjectively in each forest landscape to represent the variation along gradients in aspects, moisture, topography, and nutrient conditions. Each plot was gridded with 5 × 10 lines, 1 m apart, yielding 50 intersection points and 5 of these were selected at random for soil sampling (Figure 2). This is a random restricted sampling approach, which is proven to be efficient in terms of covering the variation in forest landscapes by limited numbers of samples (see, for example, Ohlson et al., 2009; Økland, 1996). Figure 1 shows the positions of the sample plots in the forest landscapes.

Bubble plots showing patchiness and spatial variation in the size of the soil charcoal pool in randomly selected position in gridded 5 × 10 m2 sampling plots located in neighbouring beech- and spruce forest landscapes. Circle size indicates the amount of soil charcoal in a given sampling position. Note the logarithmic scale in which the two scale marks between 0 and 1000 represent 10 and 100 g m−2, respectively. Sample plot locations in the forest landscapes are shown in Figure 1.
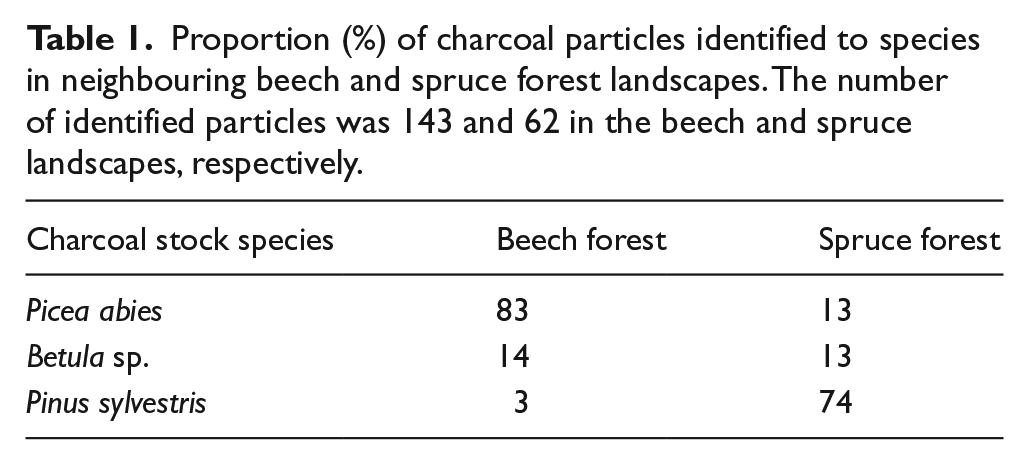
Soil samples for charcoal analysis were collected using a circular soil corer with an inner diameter of 58 mm, giving a total of 100 soil samples (50 in each forest landscape). As it is typical for boreal forest soils that the macroscopic charcoal occur in the transition between the organic top soil and the underlying mineral soil (Ohlson et al., 2009), we ensured that each soil sample consisted of a complete soil profile that contained the entire organic top soil and extended about 4 cm down into the underlying mineral soil (the sample depth into the mineral soil varied among the soil samples because of the occurrence of coarse tree roots, stones, rocks etc.). The soil samples were dried at room temperature and manually searched for macroscopic charcoal particles (approximately >0.5 mm) using forceps, a stereo-microscope and a magnification lamp according to methods described in Ohlson et al. (2009) and Kasin et al. (2013). The charcoal was then dried to constant weight at 105°C and its mass was determined by a high precision scale (Precisa 205 A SCS). Charcoal was recognized according to Hörnberg et al. (1995) and Ohlson and Tryterud (2000), and a subset of charcoal particles c. >1.5 mm were identified to species by wood anatomical characters. This subset of particles was picked out from 20 randomly selected soil samples (10 from each forest landscape). As there were proportionally more large charcoal particles in the beech than in the spruce forests, the number of selected and identified particles from the beech forests exceeded that from the spruce forests, that is, 143 versus 62 (see Table 1).
Proportion (%) of charcoal particles identified to species in neighbouring beech and spruce forest landscapes. The number of identified particles was 143 and 62 in the beech and spruce landscapes, respectively.
Five large and species-identified charcoal particles from each of the two forest landscapes were randomly selected for radiocarbon dating by AMS at Beta Analytic, USA. The mass of the dated particles ranged from 0.066 to 0.154 g, and all particles used for dating were given hot HCl acid washes and alkali (NaOH) washes. The alkali washes were then followed by a final acid rinse to neutralize the samples prior to drying and 14C-dating. This cleaning and washing procedure is established and supposed to remove ingrowth of fine roots, fungi and bacteria from the charcoal. The INTCAL13 database (Reimer et al., 2013) was used to calibrate the 14C age by taking the mean of the calibration data as the intercept point for a calibration range end-limit and the mathematics used for the calibration scenario follows Talma and Vogel (1993).
Statistics
To test for the effect of forest landscape type on charcoal mass, we performed an ANOVA with sample plot as random factor. Charcoal mass data were log transformed before statistical analysis to better meet assumptions of normality and equal variances. To estimate the degree of spatial variation in the size of the soil charcoal pool, that is, its patchiness, we standardized the variation among the 5 × 10 m2 sample plots by calculating the coefficient of variation (CV) for each sample plot (n = 5), for then to compare the amount of variation (CV) between the beech and spruce forest landscapes using Student’s t-test on the sample plot CV values (n = 20). All statistical tests were performed in R (version 3.2.0).
Results
Macroscopic charcoal was present in 89% of the soil samples, and the overall size of the soil charcoal pool averaged 162 ± 44 g m−2 (± 1 SEM). There was seemingly more charcoal in the beech forest than in the spruce forest, that is, on average 230 ± 86 versus 93 ± 16 g m−2. However, the charcoal pool in the beech forest was characterized by a profound variation (Figure 3a) and the average size of the soil charcoal pool did not therefore differ significantly between the beech and the spruce forests (F = 1.29; df = 1, 18; p = 0.271; ANOVA).
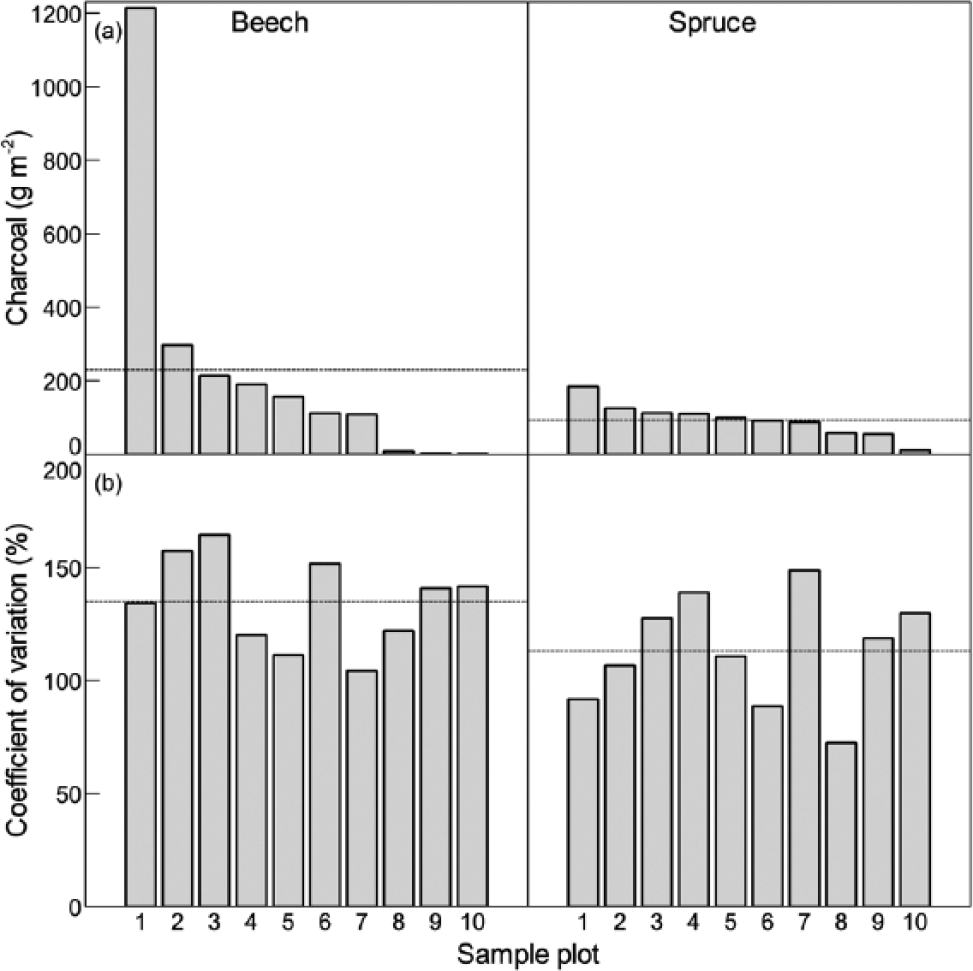
(a) Average amount of soil charcoal in 10 × 5 m2 sample plots in neighbouring beech and spruce forest landscapes. Five soil samples for charcoal analysis were collected at random position in each sample plot. Plots are arranged according to their average amount of soil charcoal, and their locations in the forest landscapes are shown in Figure 1. (b) Within sample plot variation in soil charcoal amounts. The coefficient of variation (CV) is significantly larger in the beech- than in the spruce forest (t = −2.162; df = 18; p = 0.044). Landscape level averages are shown by stippled lines.
Interestingly and importantly, all species-identified charcoal particles in the beech forest originated from either spruce, birch or pine, and none originated from beech (Table 1). This in turn indicates that large-scale fire disturbance has been absent in the beech forest landscape ever since beech gained dominance, and as most of the identified charcoal particles in today’s beech forest originated from spruce, this is evidence of fire disturbance in the spruce forest prior to beech establishment. Moreover, as pine charcoal dominated in today’s spruce forest (Table 1), this is a corresponding evidence of fire disturbance in the pine forest landscape prior to the establishment of the spruce forest.
As regards spatial variation in the charcoal pool, the amount of soil charcoal showed a significantly larger variation in the beech forest than in the spruce forest (Figure 3b), and this was the case for the variation among the five individual soil samples that were collected in a given 5 × 10 m2 sample plot, as well as for the variation among sample plot averages. For example, the largest within sample plot variation among individual soil samples ranged from 0 to 3112 g m−2 in the beech forest, while the corresponding variation in the spruce forest ranged only from 48 to 396 g m−2. Here, it is worth noting that this variation occurred within sample plot positions as close as 4 m to each other in the beech forest, and the high degree of patchiness and spatial variation is clearly shown in Figure 2. The larger variability in the beech forest was also reflected by the large variation among sample plot average values, which ranged from 2 to 1214 g m−2 in the beech forest, and from 11 to 184 g m−2 in the spruce forest.
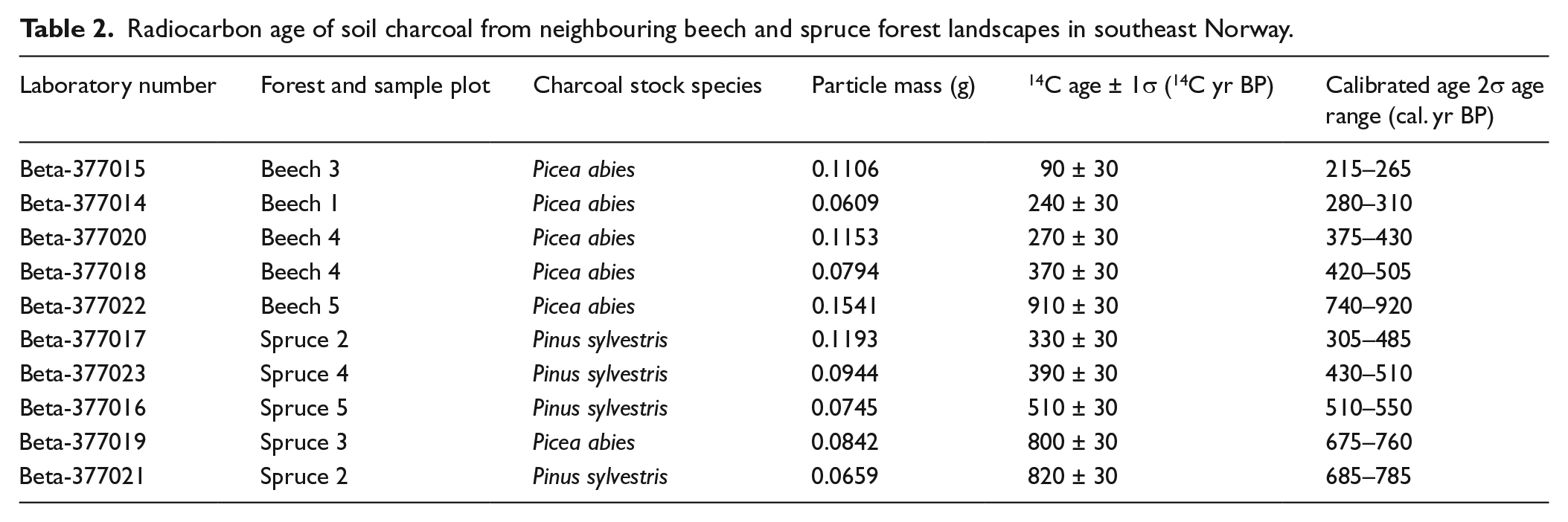
The overall average radiocarbon age of the soil charcoal was 473 ± 88 years, and the average age of the charcoal was higher in the spruce forest than that in the beech forest, that is, 570 ± 102 versus 376 ± 141 years (Table 2). However, the age difference of the charcoal between the two forest types was not statistically confirmed (t = 1.115; df = 8; p = 0.297), and as was the case for the size of the charcoal pool, also the radiocarbon ages showed a larger variation in the beech forest as compared with the spruce forest as both the oldest and the youngest charcoal particles originated from the beech forest (Table 2).
Radiocarbon age of soil charcoal from neighbouring beech and spruce forest landscapes in southeast Norway.
Discussion
As macroscopic charcoal was found in as much as 89% of the soil samples, we conclude that historic fires have been ubiquitously present in both forest landscapes in our study area. Actually, this figure on macroscopic charcoal occurrence in soil samples is among the highest proportions that have been recorded in northwestern Europe so far, and as comparison, the average probability for the presence of macroscopic charcoal in boreal soil samples from this area is about 50% (Ohlson et al., 2009). Also, the size of the charcoal pools is among the largest recorded, and this was in particular the case for three of the sample plots in the beech forest landscape with averages >200 g m−2, which parallel the largest pool sizes reported from pyrogenic forests elsewhere in northwestern Europe (Kasin et al., 2013; Ohlson et al., 2009, 2013). Presupposing that our results are representative for the 19-ha beech forest landscape at Brånakollane as a whole, which is fairly reasonable because of our sampling approach, a figure of 43,700 kg for the total landscape pool of charcoal is indicated. This in turn corresponds to a figure of c. 23,000 kg C as the average C concentration in fossil soil charcoal is estimated to 53% (see Ohlson et al., 2009). As brown soils characterize the beech forest landscape, and as earthworms are known to mix these soils, it is possible that we have slightly underestimated the size of the charcoal pool as the mixing may have moved charcoal down in deeper soil layers than those we have sampled. However, our most important finding is that the beech forest established after landscape level fire disturbances, and that large-scale fires have been absent in the beech forest landscape ever since beech took over the dominance (this because there were no charcoal from beech in the soil charcoal pool – see Table 1). The fire disturbances that paved the way for beech were initiated c. 900 years ago and seem to have culminated during a period about 400 to 200 years ago (i.e. from c. AD 1600 to 1800 – see Table 2). That the establishment of today’s beech forest started c. 1000 years ago, and that beech gained dominance in the same time-period as the fire disturbances culminated, that is, c. 400 to 200 years ago, is supported by Bjune et al. (2013), whose pollen analysis from a small forest hollow in our study area further suggests that spruce was abundant in the forest landscape in about 400 years prior to the establishment of the beech forest. That spruce was present, and likely dominant, before beech is corroborated and confirmed by our results as the majority of the charcoal that we found in today’s beech forest landscape turned out to originate from spruce (Table 1). Thus, our results corroborate Bradshaw and Lindbladh (2005) as well, who suggest that the local stand scale establishment of beech should be favoured by fire disturbance prior to establishment. It was, however, somewhat unexpected that fires were important in the pre-beech spruce forest landscape as wildfire activity is known to have been generally low in Norwegian spruce forests in late-Holocene (Ohlson et al., 2006, 2011; Tryterud, 2003). We do not know why fire occurred in the pre-beech spruce forests, but two possible explanations are ignition by lightning as the study landscape represents geographically high positions, and/or anthropogenic use of fire to improve grazing (see Blanck et al., 2013; Storaunet et al., 2013).
We also show that it is likely that fire has been absent in today’s spruce forest landscape ever since spruce took over the dominance. The main reason for this is that the majority of the species-identified charcoal in the spruce forest originated from Scots pine, which suggests that Scots pine was much more abundant than spruce in the pre-fire forest (Table 1). This in turn indicates that fire disturbances in the pre-spruce pine forests opened the door for spruce establishment, and that the spruce has been capable to fully outcompete Scots pine as the fire disturbances ceased, which is also well documented from dendrohistorical studies in contemporary boreal forest stands (Linder et al., 1997). In this context, it is also worth noting that beech has not been able to take over the dominance in the former pine forest, and one reason for this could be unfavourable soil conditions because of out-wash of fine mineral soil fractions as this landscape is located well below the marine limit (see Geological Survey of Norway, 2016).
Taken together, the results discussed above give support for the importance of fire disturbance as a key determinant of local beech forest establishment. However, as fire disturbance turned out to be equally important for the establishment of the spruce forest as for the beech forest, our results do not support our first hypothesis, that is, that beech should be more dependent on fire disturbance than spruce for establishment success. The reason for the rejection of the hypothesis is that neither the size nor the age of the soil charcoal pool differed significantly between the forest landscapes. In contrast, our results support our third hypothesis that the charcoal pool in today’s beech forest landscape should be dominated by particles originating from former tree species that characterized the landscape prior to beech.
Charcoal age
Charcoal is recalcitrant and several studies have radiocarbon dated boreal soil charcoal that is several thousand years old (e.g. De Lafontaine and Asselin, 2011; Gavin, 2003; Gavin et al., 2003; Ohlson et al., 2009). In the light of this, it was unexpected that none of our dated charcoal particles were older than 910 years. In particular, as Bjune et al. (2013) report that significant amounts of charcoal were produced by recurring fires from c. 12,000 to 2500 yr BP. A possible reason for the lack of old charcoal in the mineral soil that we have sampled is that charcoal survival may be significantly shorter in shallow mineral soils as compared with the wet and deep peat soil that was sampled by Bjune et al. (2013). One reason for this is that it is likely that a fire event may consume a proportion of the charcoal produced by previous fires on mineral soils with shallow organic top soil layer (Kane et al., 2010). However, the influence of fire on the size of the soil charcoal pool remains poorly understood, although recent research indicates that only a small proportion of the pool may be consumed by contemporary fires (Santín et al., 2013; Tinkham et al., 2016). A further reason for the low age is that abiotic processes such as gelifluction and cryoturbation degrade charcoal (Preston and Schmidt, 2006). Furthermore, soil charcoal is an attractive habitat for microorganisms (Hockaday et al., 2007; Zackrisson et al., 1996), and this may contribute further to charcoal degradation as soil charcoal microbial communities are characterized by high biological activity (Pietikäinen et al., 2000).
As regards charcoal age, our results only give marginal support for the second hypothesis that charcoal should be younger in the beech forest as compared with the spruce forest. This is because there was no significant statistical difference in charcoal age between the forests, although the charcoal on average was almost 200 years younger in the beech forest. Here, a larger number of radiocarbon dated particles would have been helpful. A further support for our hypothesis is that the youngest individually dated charcoal particle was found in the beech forest. However, also the oldest individually dated particle was found in the beech forest (Table 2), which challenges the same hypothesis. Moreover, ‘the old wood problem’ sensu Bowman (1990) may have contributed to discrepancy in charcoal age between the two forest landscapes – Scots pine was the dominating charcoal stock species in the spruce forest landscape and it has a longer lifespan than Norway spruce, which was the dominating charcoal stock species in the beech forest landscape; see Table 1.
Spatial variation and patchiness in the soil charcoal pool
Experimental burns have shown that macroscopic charcoal is produced and deposited locally (Clark et al., 1998; Ohlson and Tryterud, 2000), and therefore we suggest that the spatial distribution of the pre-fire charcoal stock (i.e. the fuel load) may be a key explanation to the high degree of patchiness. Coarse woody debris such as large branches, stumps and tree trunks on the ground are of particular importance in this context as they are discrete units and make up a substantial proportion of the fuel load in the forest. An incomplete fire consumption of these structures will result in large amounts of charcoal with highly local and patchy distributions. The explanation for the patchier distribution in the beech forest may be the rugged terrain in the beech forest landscape (see Figure 1a). The rugged terrain in turn gives rise to downhill movement of charcoal stock material (e.g. logs and branches) as well as charcoal, which in turn will emphasize the patchiness of the soil charcoal pool. During periods of heavy rainfall, there will also be a downhill wash of water and soil erosion, which may also have potential to increase the patchiness of the soil charcoal.
Fire driven ecosystem change
According to our results, both forest landscapes have gone through fundamental transformations during the last c. 1000 years as pine forests were transformed to spruce forests, and spruce forests were transformed to beech forests. These transformations were fundamental, as ecosystem structure, function and biodiversity were radically changed because of shifts of forest key species. Our results further indicate that fire disturbances have triggered these events, which started about 1000 years ago. It is probable that these ecosystem changes are among the most rapid and most radical in a late-Holocene time perspective as Lindbladh et al. (2000) show that south Scandinavian forests have gone through their most rapid changes during the last c. 1000 years.
Footnotes
Acknowledgements
The authors are thankful to Kjell Lie for his positive attitude and for giving them access to a network of closed forest roads.
Funding
This study was supported by the Research Council of Norway (project 225018; From spruce to beech forests – fundamental ecosystem transformation driven by climate change). Marta Vázquez del Olmo’s stay in Norway was supported by an EU Erasmus exchange grant.