
Abstract
The late-Holocene bird remains retrieved during archaeological excavations on pelagic island of Palagruža, Croatia, were analysed. The results of the analysis revealed presence of representatives of nine families: Procellariidae, Sulidae, Ardeidae, Accipitridae, Phasianidae, Rallidae, Laridae, Strigidae and Corvidae. Among the bird remains, the most numerous were remains of the Northern Gannet, Morus bassanus, which were widely distributed in the Mediterranean during the late Pleistocene. The finding of the Northern Gannet bones on Palagruža represents the first fossil record of this species in the Adriatic Sea and indicates a possible breeding site on the island. The direct Accelerator Mass Spectrometry (AMS) radiocarbon dating on two gannet bones revealed the age of 2878 ± 34 and 2694 ± 45 yr BP. Gannet bones from Palagruža were some 6000 years younger than other fossil records of that species from the Mediterranean. It is possible that during the gannet’s northward distributional shift, probably caused by centennial climatic variability and human pressure, gannet colony on that remote island in the Adriatic lasted longer than in the rest of the Mediterranean and existed until the late-Holocene. Lower sea-surface temperatures compared with the eastern Mediterranean, higher nutritive productivity and remoteness of Palagruža provided adequate breeding opportunities during that period.
Introduction
Studies of fossil bird remains on islands, especially isolated ones, can provide valuable insights into past bird assemblages and improve our current understanding of the changes in bird fauna over long periods of time (Alcover et al., 1992; Steadman and Takano, 2013; Weesie, 1988). During the Holocene, large numbers of insular bird species underwent full extinction or local extirpation. Around the world, numerous seabird species became extinct, while many other faced local extinction or large distributional shifts (Ramirez et al., 2010; Steadman, 1995, 2006). Two main causes were identified: environmental changes (climate, sea level, sea temperature, sea currents) and anthropogenic influence through habitat destruction, disturbance, introduction of non-native predators (such as dogs, cats, rats and pigs), hunting and egg collecting (Milberg and Tyrberg, 1993; Ramirez et al., 2010; Rando and Alcover, 2010; Zotier et al., 1999). The effects of impoverishment mechanisms on Mediterranean seabirds were previously discussed (Zotier et al., 1999). Late Pleistocene Mediterranean marine avifauna suggests cool, biologically productive sea that might have been an important refugium for boreal seabirds during the coldest periods of glaciation (Tyrberg, 1999). The Pleistocene records of seabirds in the Mediterranean comprise at least 20 marine bird species (Zotier et al., 1999), including extant Mediterranean breeding species (such as shearwaters, gulls and Shag Phalacrocorax aristotelis), with modern range in the northern Atlantic (Northern Gannet Morus bassanus, Atlantic Puffin Fratercula arctica) and globally extinct species (Great Auk Pinguinus impennis) (Tyrberg, 1998). The succession of glaciations during Pleistocene changed coastlines and islands, while humans colonized islands with dramatic consequences on the marine bird communities (Martin et al., 2000; Rando and Alcover, 2010). During Pleistocene and early-Holocene, breeding colonies of marine birds shifted from some islands to others and several species disappeared (Zotier et al., 1999). As a result, the Holocene provided fossil remains of only 11 marine bird species (Zotier et al., 1999).
Among species that showed difference in distribution between late Pleistocene and modern range is the Northern Gannet. The first record of genus Morus in the northern hemisphere was derived from the middle Miocene deposits of Romania (Mlíkovský, 2009). In late Pleistocene, when the Mediterranean Sea was an important refugium for boreal seabirds, the Northern Gannet was widely distributed in the Mediterranean, reaching Crete to the east (Tyrberg, 1998, 1999, 2008; Weesie, 1988; Zotier et al., 1999), but it shifted its range towards north and is today distributed in the northern Atlantic (Figure 1). It inhabits productive seas with plenty of fish and breeds on uninhabited islands or inaccessible cliffs. In autumn and winter, the Northern Gannet is present in the western Mediterranean, with small numbers wintering in the eastern Mediterranean (BWPi 2.0.1, 2008; Del Hoyo et al., 1992).
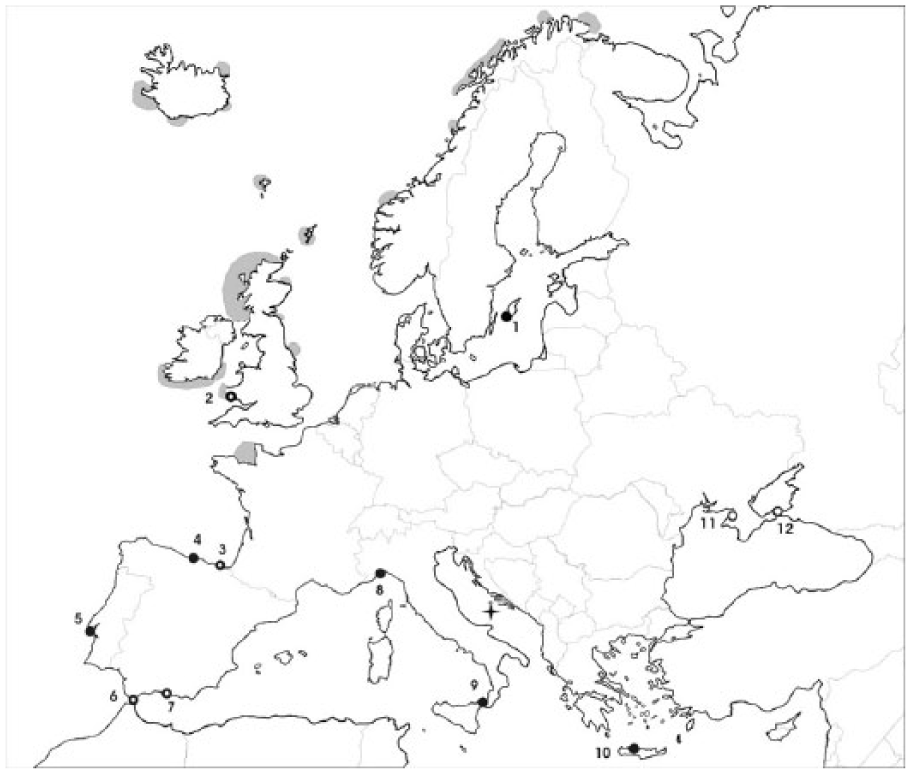
Location of Palagruža (black star) with localities of fossil remains of the Northern Gannet: full black dots indicate late Pleistocene sites, dots with thick line sites with both late Pleistocene and Holocene remains, and empty dots Holocene sites). Grey areas indicate current Northern Gannet’s breeding area. Localities are numbered as follows: 1 – Ajvide; 2 – Paviland cave, Port Eyon; 3 – Santa Catalina; 4 – Cueva de Torre; 5 – Gruta di Figueria Brava; 6 – Sevelles and Ibex Cave; 7 – Cueva de Nerja; 8 – Arene Candida; 9 – Archi; 10 – Simonelli Cave; 11 – Mirmekia; 12 – Pantikapaion and Nymphaion.
Fossil records of seabirds are relatively rare in Croatia. Palaeontological studies were mostly conducted on coastal or island sites (e.g. Malez, 1988, 2001; Malez-Bačić, 1979; Oros Sršen et al., 2014), which were a part of mainland during the last glaciation while the coastal sites of glacial age are now mostly submerged as a result of a sea level rise during postglacial period (Surić and Juračić, 2010; Tyrberg, 1999). Contrary to other Adriatic islands formed when Holocene marine transgression separated the westernmost ranges of the Dinaric Mountains from the mainland by drowning the valleys between them, Palagruža and two other off-shore islands, Jabuka and Sušac, were probably islands during the entire Pleistocene (Forenbaher and Kaiser, 2005; Sikora et al., 2014). Eventually, they might have been a part of the mainland as sea cliffs only during the Last Glacial Maximum (ca. 30–19 ka BP; e.g. Antonioli and Vai, 2004; Ridente and Trincardi, 2005).
The fossil avifauna of Palagruža has not been studied yet. The interest for avifaunal remains from Palagruža was inspired by the latest archaeological discovery of a Greek sanctuary dedicated to the Trojan hero Diomedes, mentioned in ancient literary sources (Kirigin et al., 2009). In these texts, ancient writers regularly describe birds that lived on that island (Kirigin and Čače, 1998). These intriguing descriptions encouraged us to closely examine remains of bird bones in order to gain insight into species assemblages of Palagruža during different time periods. This paper thus presents the first list of avifaunal remains from Velika Palagruža and the first find of the Northern Gannet in the Adriatic Sea. The second aim of this study was to identify the status of the Northern Gannet in the Adriatic Sea during the late-Holocene, to explore the chronology and to identify probable causes of its range changes.
The study site
Velika Palagruža (42°23′N, 16°15′E) is a small island in the central Adriatic Sea and the greatest island of the Palagruža archipelago (Figure 1). It is situated 57 km from the Gargano promontory to the south, and 60 km from the Island of Lastovo to the northeast (Korbar et al., 2009). Velika Palagruža is a narrow, rocky ridge, 1390 m long and 270 m wide with the highest point of 103 m a.s.l., situated on the mid-Adriatic (or Palagruža) Sill (Bognar, 1996). Palagruža Sill acts as a natural barrier to incoming and outgoing water masses from the Adriatic, which causes intensive mixing of waters of different origin (Vukadin and Stojanoski, 1996) and upwelling of nutrients-rich bottom waters in euphotic zone resulting in enhanced productivity (Sinovčić, 1996). This area is one of the richest fishing areas in the Adriatic Sea, especially for small, pelagic fish (sardine, anchovy and mackerel; Sinovčić, 1996). The climate of Palagruža is Mediterranean with hot summer (Csa according to Köppen’s climate classification system; Šegota and Filipčić, 2003). The mean annual air temperature ranges from 12.5°C to 20.5°C (Lukšić, 1996), while mean annual sea temperature range is 12.0–22.0°C (Vukadin and Stojanoski, 1996). There are no sources of freshwater other than modest amounts of rainfall throughout the year, and the mean annual amount of precipitation is ca. 300 mm (Milković, 1996). It can be assumed that 2500 years ago, Palagruža occupied a very similar area as it does today because of its steep coastline (from 12° to more than 55°; Bognar, 1996) and the rise of the Adriatic sea level by only 2 m over the last 2000 years (Surić, 2009).
Archaeological fieldwork recovered the earliest evidence of human presence on the eastern part of Palagruža, on site Jonkova njiva, dated ca. 6000 BC to the early Neolithic period (Forenbaher and Kaiser, 2005). The central flat area of the island, Salamandrija (Figure 2), where the bird bone remains were discovered, is covered by humic soil. This is the largest flat area on Palagruža and the main archaeological site (Figure 3a). The earliest evidence of human activity here dates to late Copper/early Bronze Age. The recovered pottery fragments are attributed to Cetina culture, which belong to period between ca. 2500 and 1800 BC (Forenbaher and Kaiser, 1997). After this period, there is no archaeological evidence of human presence on the island over the next 1200 years. By the end of the 6th ca. BC, Greeks founded sanctuary on Salamandrija plateau and seafarers regularly visited it for centuries (Kirigin and Čače, 1998; Kirigin et al., 2010). Material traces of their presence are dated to the end of the first century AD (Miše and Šešelj, 2008). The sanctuary itself, as well as earlier layers, was destroyed by intensive human activity of the later period: construction of the late Roman fortification, the mid-19th century church of St Michael, several cisterns, water collecting pavements and two Italian military installations, as well as by naval bombardment during the First World War (Kirigin et al., 2010; Figure 3b).

Island of Velika Palagruža, Croatia. The white arrow indicates archaeological site Salamandrija.
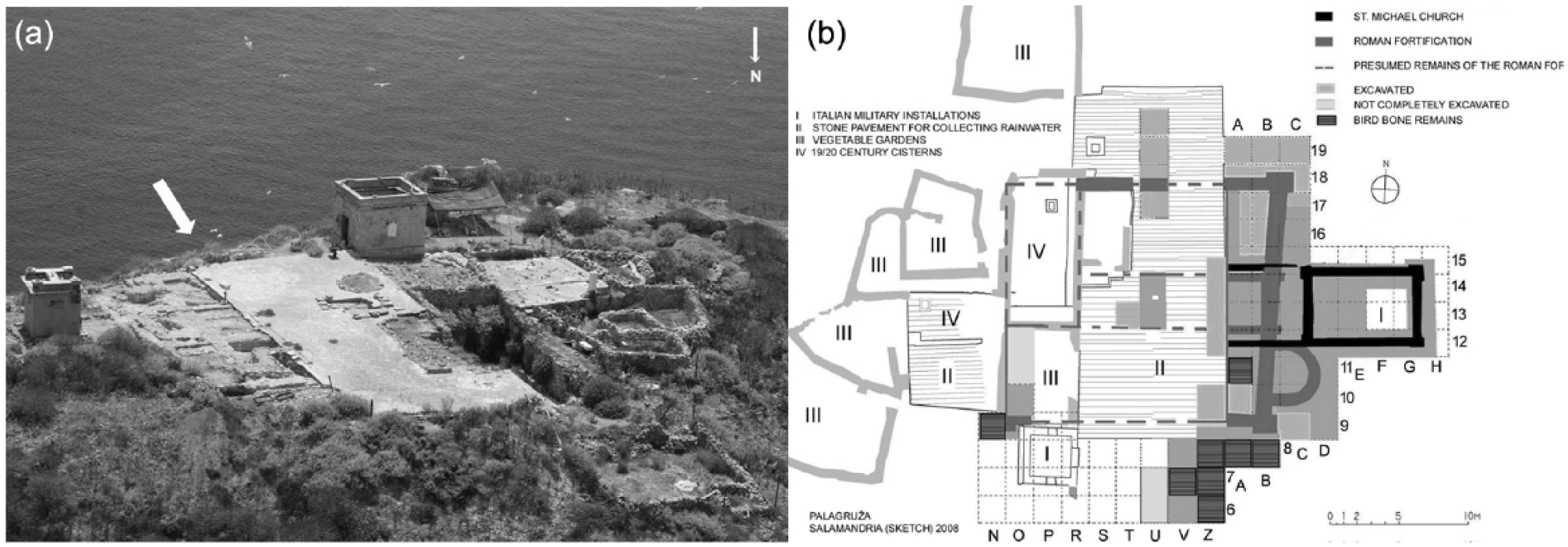
Island of Velika Palagruža, the Salamandrija site (modified after Kirigin et al., 2010): (a) the white arrow indicates southern slope, where the bird bones were found; (b) sketch plan of the excavation area, with marked quadrants in which the bird bones were found.
Materials and methods
The present paper investigates avifaunal remains collected during archaeological excavations of Salamandrija area on Velika Palagruža in 1996, and from 2002 until 2008 (Kirigin and Čače, 1998; Kirigin et al., 2010). The material comes from various archaeological layers excavated from the southern slope of Salamandrija in trenches A8, B8, N9, Vi6, Vi7, V7, Z6, Z7 and Z8 (Figure 3b), where the sediment was preserved. Layers were mixed and hard to interpret because of destruction of the site over different periods of human activity and/or erosion. Layers 4050 and 8150 emerged after the destruction of Greek sanctuary (established in the second half of the first millennium BC) and are mixed with earlier Bronze Age material (the third millennium BC); 4041 is a layer of late Roman building construction preparation, which can be dated to the 3rd to 4th century AD. Thus, it can be presumed the sediments were deposited between ca. 4500 and 1600 years before present.
The bird remains include 132 complete and fragmented bones in total, of which 63 (48%) were anatomically identifiable. The bones were anatomically, taxonomically and taphonomically analysed. Identifications were made using modern comparative osteological collections at the Institute for Quaternary Paleontology and Geology of the Croatian Academy of Sciences and Arts and Natural History Museum in Vienna (NHMV), as well as relevant literature (Cohen and Serjeantson, 1986; Gilbert et al., 1985; Stucchi, 2003; Tomek and Bocheński, 2000; Van Tets et al., 1988). Taxonomically indeterminate samples were classified into four size categories: ‘Small’ (song-bird size and order Passeriformes except fam. Corvidae), ‘Medium’ (dove size), ‘Large’ (mallard size) and ‘Very Large’ (goose size and larger). Corvidae was the only taxonomically analysed family from the order Passeriformes. The bones were coded with site initials and year of excavation (e.g. Pg 02) followed by quadrant (e.g. Z8) and archaeological layer (e.g. 4050), while the last number is bone code number, for example, Pg 02 Z8 4050, 5.
Remains are quantitatively presented as a number of identified specimens (NISP) that represents the number of all skeletal elements and fragments (Lyman, 2008). The minimum number of individuals (MNI) of each species was counted per each layer according to the most abundant skeletal element (rights or lefts). Bone modifications were examined under 10× magnification hand lens and compared with the literature data (Andrews, 1990; Lyman, 1994; Serjeantson, 2009). Measurements were made using digital calliper and rounded to 0.1 mm. They were taken after Tomek and Bocheński (2000) for Corvids and Von den Driesch (1976) for other species.
The collagen was extracted from two bones belonging to Northern Gannet derived from layer 4050 (Figure 4) and was dated via Accelerator Mass Spectrometry (AMS) radiocarbon method at the 14CHRONO Centre Queens University Belfast. The dates were calibrated by radiocarbon CALIB 7.1.0 (Stuiver and Reimer, 1993) calibration program and MARINE13 (Reimer et al., 2013) radiocarbon age calibration curve when marine reservoir correction ΔR was applied. The marine reservoir effect for the Adriatic Sea is 424 ± 57 14C years and reservoir correction ΔR is 77 ± 57 14C years (Faivre et al., 2015).
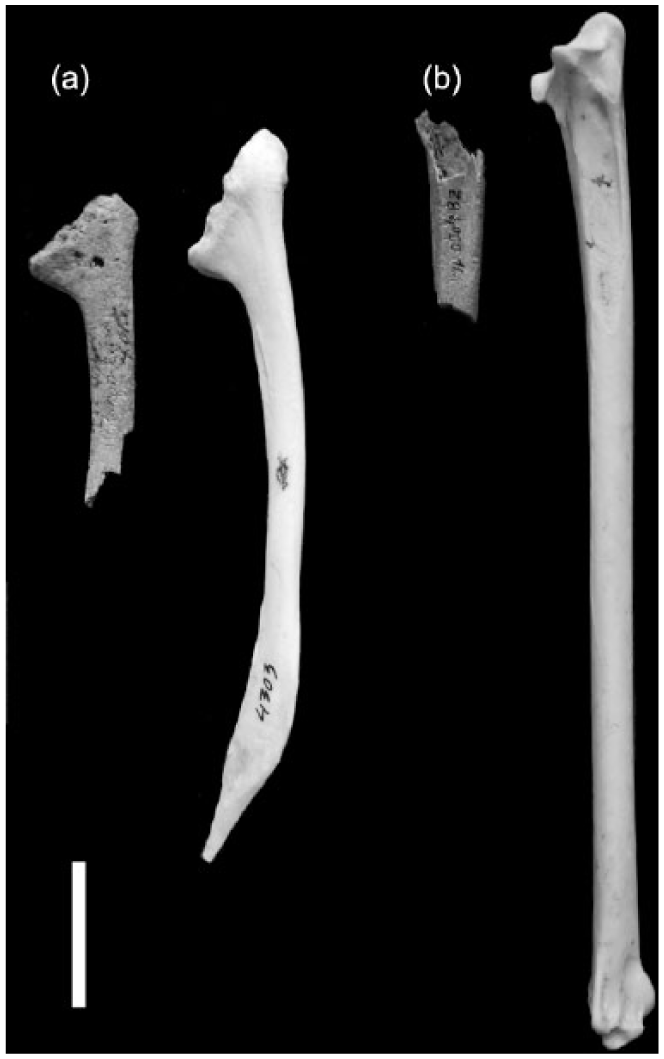
AMS radiocarbon dated bones of the gannet (Morus bassanus) compared with modern complete specimens from Natural History Museum in Vienna (on the right side): (a) medial view of the right proximal scapula (Pg 02 Z8 4050, 6) and (b) ventral view of the right proximal ulna (Pg 96/02 Z8 4050, 12). White line represents 2 cm.
The data on extant avifauna of Palagruža were taken from three sources: the surveillance of autumn migration in late September and October 1897 and 1898 (Godez, 1899, 1901), short study of breeding fauna and spring migration in May 1911 (Babić and Rössler, 1912) and the project ‘Fauna of Croatian islands’ of the Croatian Natural History Museum conducted in August and September 1997 (CNHM, unpublished data).
Results
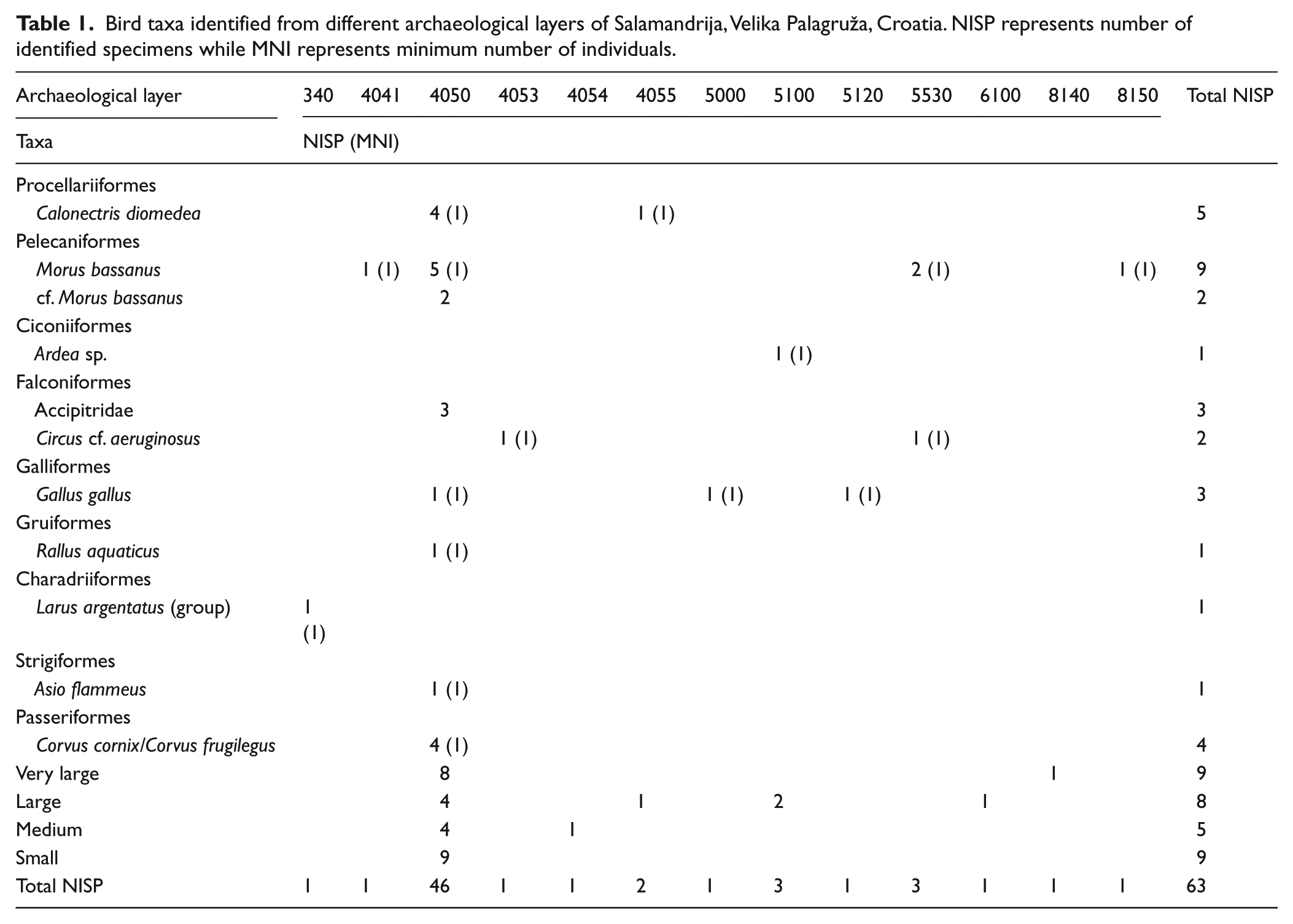
Avifaunal assemblage of Velika Palagruža includes 11 different taxa determined from 63 fragments, of which seven could be determined to the species level (Table 1). The 63 remains belong to nine families: Procellariidae, Sulidae, Ardeidae, Accipitridae, Phasianidae, Rallidae, Laridae, Strigidae and Corvidae (metrical data for these remains are provided in the supplementary material). According to the NISP, the most numerous were remains of the Northern Gannet (Table 1). The MNI of the Northern Gannet was one per each layer and four in total. All remains belonged to adult birds. The most abundant bones were wing bones (Table 2, Figure 5). Since the gannet’s presence was recorded for the first time in osteological material from the Adriatic Sea region, these bones are discussed below in more detail:
Bird taxa identified from different archaeological layers of Salamandrija, Velika Palagruža, Croatia. NISP represents number of identified specimens while MNI represents minimum number of individuals.
Bird skeletal element representation by taxon from island of Velika Palagruža, Croatia.
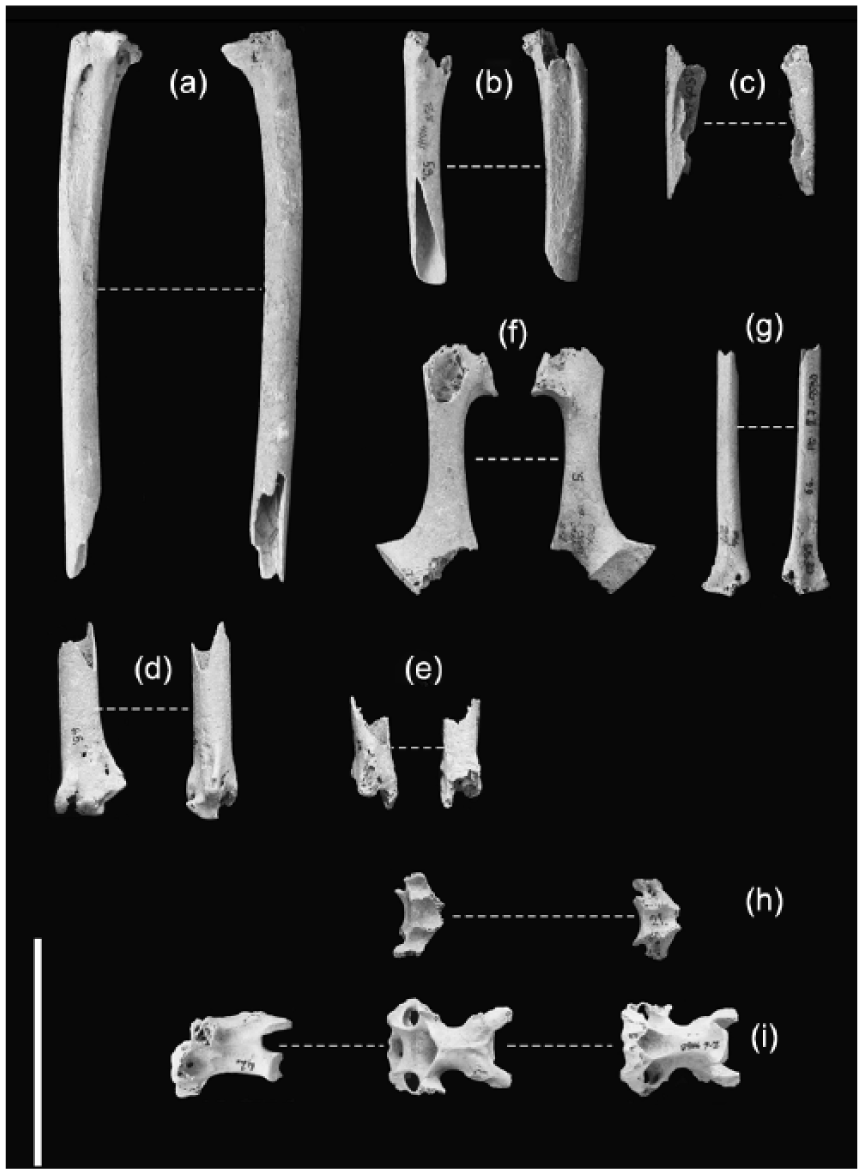
The bones of Morus bassanus (inv. no.: 5, 9, 27, 53, 65, 66, 70) and cf. Morus bassanus (inv. no.: 21, 42) from Salamandrija, island of Velika Palagruža: (a) proximal left ulna (Pg 04 A8 8150, 70), ventral and dorsal aspect; (b) proximal right ulna (Pg 96 Z8 4041, 53), dorsal and ventral aspect; (c) proximal left ulna (Pg 02 Z7 4050, 27), ventral and dorsal aspect; (d) distal left ulna (Pg 02 Z7 5530, 65), ventral and cranial aspect; (e) distal left ulna (Pg 02 Z7 4050, 19), cranial and caudal aspect; (f) left coracoid (Pg 02 Z8 4050, 5), ventral and dorsal aspect; (g) distal left radius (Pg 03 Z7 5530, 66), dorsal and ventral aspect; (h) centrum epiphysis anterior cervical vertebra (Pg 02 Z7 4050, 21), dorsal and ventral aspect; (i) cervical vertebra (Pg 03 Z6 4050, 42), lateral, dorsal and ventral aspect. White line represents 5 cm.
Order: Pelecaniformes (SHARPE, 1891)
Family: Sulidae (REICHENBACH, 1849)
Morus bassanus (LINNAEUS, 1758)
Material: Damaged left coracoid (Pg 02 Z8 4050, 5)
Remarks. The coracoid shows the characteristics of a large Sulid (Stucchi, 2003; Van Tets et al., 1988). The shaft is stout and the tip of the glenoid is pointed. The parts of the bone characteristic for distinguishing between genera Morus and Sula were not preserved (after Van Tets et al., 1988), but size and shape of the bone match the modern comparative specimen of Morus bassanus (Figure 5). Northern Gannet is taxonomically isolated in Europe in Holocene and is unlikely to be mistaken for some other Sulid.
Material: Proximal right scapula (Pg 02 Z8 4050, 6)
Remarks. The bone had the same size and shape of a modern comparative specimen of Morus bassanus (Figure 4a). However, the part of olecranon characteristic for distinguishing between genera Morus and Sula was not preserved (after Van Tets et al., 1988).
Material: Proximal right ulna (Pg 96 Z8 4041, 53; Pg 96/02 Z8 4050, 12), proximal left ulna (Pg 02 Z7 4050, 27; Pg 04 A8 8150, 70), distal left ulna (Pg 02 Z7 4050, 19; Pg 02 Z7 5530, 65)
Remarks. In general characteristics, the specimens resemble ulna of the genus Morus (Van Tets et al., 1988) and modern comparative specimen of Morus bassanus. The proximal part in cross section is triangular at the epiphysis and more squared in lower part, brachial muscle impression is oval and elongated with pneumatic foramen on its proximal end, shaft is straight, distal epiphysis in cross section is triangular, carpal tuber is located more proximally, as described in Van Tets et al. (1988). Although some specimens were not very well preserved, the size and shape matched modern comparative specimens of Morus bassanus (Figures 4b and 5).
Measurements (Pg 02 Z7 5530, 65): Did = 15.4 mm
Material: Distal left radius (Pg 03 Z7 5530, 66)
Remarks. The radius shows the morphological characteristics of the genus Morus. The shaft is straight and distal end has one prominent foramen, more proximally located than in Sula (Van Tets et al., 1988). It resembles radius found in modern comparative specimen of Morus bassanus (Figure 5).
cf. Morus bassanus
Material: Cervical vertebra (Pg 03 Z6 4050, 42), centrum epiphysis anterior cervical vertebra (Pg 02 Z7 4050, 21)
Remarks. The size and shape of the vertebrae match the gannet’s (Figure 5), but since they are fragmented and the vertebrae are among the least reliable parts of skeleton for the identification, the exact determination was not possible, and the bones were assigned to cf. Morus bassanus.
Taphonomy
The majority of all bones (90.5% NISP) were etched by chemicals secreted by roots and show sinuous etching marks with smooth U-shaped cross-channels (as it is described in Lyman, 1994). The rest of the bones showed signs of erosion (3.2% NISP), spalling (1.6% NISP), chemical etching of unknown origin (3.2% NISP) or were not weathered at all (1.6%). Northern Gannet’s bones showed similar pattern: 100% (N = 9) were etched by root’s acids, and eroded on the ends. No sign of human or animal predation was recorded.
The most numerous element in Palagruža assemblage (Table 2) is ulna (NISP = 16), followed by phalanges (NISPanterior + posterior = 8), carpometacarpus (NISP = 7), femur and tarsometatarsus (NISP = 5), than vertebrae (NISP = 4), coracoid (NISP = 3), and tibiotarsus (NISP = 2). The least numerous are maxilla, scapula, humerus and radius with only one identified specimen (NISP = 1).
Absolute age
The radiocarbon dates obtained from the gannets’ bones are presented in Table 3. The widest range of calibrated ages is from 2705 to 2095 cal. yr BP, when 2σ calibration interval with reservoir correction is included.
Radiocarbon dates of Northern Gannet Morus bassanus from the island of Velika Palagruža.

C/N: carbon–nitrogen ratio.
Radiocarbon age and σ calibration intervals are given. Calibration intervals have been calculated using MARINE13 radiocarbon age calibration curve (CALIB 7.1.0) and reservoir correction ΔR = 77 ± 57 for the Adriatic Sea.
Discussion
The bird assemblage from Palagruža, although small in number of identified taxa, provided valuable data about the late-Holocene palaeoenvironment and species occurrence in that locality.
Apart from the Northern Gannet, it includes two other seabird species: Cory’s Shearwater Calonectris diomedea and a gull belonging to Larus argentatus complex, most probably the Yellow-legged Gull Larus michahellis. Fossil records of both species indicate their wide breeding range in the Mediterranean during Pleistocene and Holocene (Zotier et al., 1999). Today, both species breed on Palagruža (CNHM, unpublished data).
Other identified species – Marsh Harrier Circus aeruginosus, Short-eared Owl Asio flammeus, Water Rail Rallus aquaticus, heron Ardea sp. and crow Corvus sp. – for which mostly the presence of single individuals was recorded were most probably only passage birds, while bone remains of the domestic chicken Gallus gallus originated from birds brought to the island for human consumption. The same could be concluded for the majority of determined mammalian remains analysed by Jane Sanford (2015, personal communication), which mostly comprised domesticated animals. Rocky habitats of Palagruža do not provide breeding or feeding opportunities for waterbirds, such as rails and herons. However, because of its geographical location, Palagruža presents an important stopover site in the mid-Adriatic. Studies of bird fauna and migration over Palagruža during the last 100 years resulted in 116 species recorded, among which were also species the remains of which were identified in this study: purple heron Ardea purpurea, grey heron Ardea cinerea, water rail, marsh harrier, short-eared owl and crows (Godez, 1899, 1901; CNHM, unpublished data). These species were identified in the sediments of late Pleistocene and Holocene sites along the Adriatic Sea (Cassoli and Tagliacozzo, 1997; Malez, 1980, 2001, 2009; Malez-Bačić, 1979; Oros Sršen et al., 2014).
Taphonomic analysis corroborated our presumptions about factors of accumulation of bird bones. Functional morphology is an important factor in bone survival (Bovy, 2002; Higgins, 1999) and the differential survival of elements because of bone density should be considered first, before the other factors such as human or animal predators. In general, better survival of wing elements compared with other parts of skeleton can be expected in powerful flyers such as albatrosses, dabblers and gulls, since their wing bones have the highest density among skeletal elements (Higgins, 1999). More terrestrial or diving birds have stronger legs, thus their leg elements are expected to predominate in the fossil assemblage, while wing-to-leg ratio is approximately equal in swimming birds (Cruz, 2007). According to the skeletal element representation in the Palagruža assemblage, preservation was favourable for bones with highest density (wing elements of strong flyers such as gannet) and compact bones (tarsometatarsus, phalanges, coracoid) while fragile elements such as cranium, furcula, sternum, pelvis and fibula were not identified at all (Table 2). This indicates that structural density was an important factor in preservation of bones. The majority of avian bones were etched by root’s acids. Predominant vegetation on the upper ridge and southern slopes of Palagruža is evergreen scrub and herbaceous geophytes (Lovrić and Rac, 2003), and we can presume that similar vegetation existed on the island 2500 years ago. As the roots of these plants do not penetrate deep into the ground (Canadell et al., 1996), it is likely that root etching occurred in the preburial period (Fernández-Jalvo et al., 2010). This indicates the bones were naturally deposited on the ground after the birds have died, were shallow buried and were eventually covered by plant roots over a period of time. Heavy root etching, which in most cases covers more than a half of bone surface, possibly camouflaged bone surface modifications that could indicate subareal weathering, since spalling and erosion were present on only three bones (4.8% NISP). Hence, it can be assumed that the remains of all species (except domestic chicken) were accumulated by natural death ‘in situ’, and not transported by humans.
Northern Gannet
The record of Northern Gannet bones in the late-Holocene sediments of Palagruža dates to ca. 2550–2300 cal. yr BP, and represents the species’ first finding in the Adriatic Sea. Numerous late Pleistocene gannet’s remains found along Spanish, Italian and Greek coasts and on islands point to its wide Pleistocene and early-Holocene distribution in the Mediterranean and the Black Sea (Boev, 2009; Sánchez Marco, 2004; Tyrberg, 1998, 1999, 2008; Weesie, 1988; Zotier et al., 1999) (Figure 1). The glaciation is considered to result in southward shift of European seabirds, with the Northern Gannet being among breeding species in the Mediterranean during that period (Blondel, 2009; Moreau, 1954). However, there are no records from the Adriatic Sea. The closest localities with gannet remains of the late Pleistocene age are Sicily and Crete, (Weesie, 1988). The latest remains found in the Mediterranean originated from the Late Glacial to early-Holocene (13,330 ± 270 to 8770 ± 140 yr BP) and were found in Cueva de Nerja in Spain (Tyrberg, 1998), while there are no records of this species in the Mediterranean in the late-Holocene (e.g. Sánchez Marco, 2004). Gannet bones from Crimea (the Black Sea) were dated to 4th century BC to 2nd century AD (Boev, 2009). Present records from the Adriatic are some 6000 years younger than other Mediterranean remains. This could indicate that Northern Gannets had inhabited the Adriatic Sea during a more recent period and that Holocene climate change caused the shifting of the species’ range towards the north.
Around 7000 yr BP, gannets occurred in Norway and Denmark; their occurrence was consistent with warm climatological conditions (Montevecchi and Myers, 1997). The gannet is recorded in Britain only in the Late Glacial deposits of Paviland Cave in southern Wales (Yalden and Albarella, 2008) and later in the Holocene sediments in Port Eyon Cave (Mesolithic period, 9000–6000 yr BP, Harrison, 1987), while it is a common species in Neolithic horizons (Yalden and Albarella, 2008). It is recorded in the Neolithic horizons on Gotland in the Baltic (ca. 5000 yr BP, Mannermaa and Stora, 2006). This could point to the species’ wider distribution in early and middle-Holocene before establishing present-day colonies.
The amount of gannet bones found on Palagruža and at least a 100 years span between two absolutely dated bones indicates the species’ regular presence on the island, since the probability of a vagrant species occurring in a fossil avifauna is considered to be very small (Tyrberg, 1998). Migratory behaviour and dispersion represent a significant palaeoecological problem, as an occurrence of migrant bird may potentially represent its summer breeding grounds, its wintering grounds or its route of passage (Cooper, 2005). The habit of migration is not a recent phenomenon, and during the Pleistocene glacial cycles, the bird populations must have increased and decreased migratory habits repeatedly (Bruderer and Salewski, 2008). Therefore, it is necessary to consider the potential seasonal status of the Northern Gannet from Palagruža. While assessing the seasonal status of birds from fossil records, breeding may be indicated by remains of juveniles, medullary bone in adult remains or eggshells (Cooper, 2005), but such remains were not found on Palagruža. Even if it is not possible to assess with certainty the status of the Northern Gannet on Palagruža, its breeding in that locality may be presumed. For many seabirds, the occurrence of bones outside of breeding areas is extremely rare (Serjeantson, 2001). Gannet’s breeding habitat requirements include coastal cliffs or inhabited islands for nesting and productive sea rich with fish, mostly pelagic. Steep cliffs and remoteness of Palagruža present suitable breeding habitat. It is completely devoid of mammalian predators and the presence of avian predators large enough to predate on gannets is also unlikely. The only possible raptor capable of catching gannets would be the white-tailed sea-eagle Haliaeetus albicilla, but it could only feed on pulli or nesting individuals. Still, Palagruža is not suitable for breeding of sea-eagles because of its small size and remoteness. The earliest evidence of human presence on Palagruža dates to the early Neolithic (Forenbaher and Kaiser, 2005). However, Palagruža is too small, remote and waterless to support continuous human settlements, and was therefore only a visiting point for sailors in the Adriatic (Forenbaher and Kaiser, 2005). According to the Serjeantson (2001), the presence of gannet bones on prehistoric sites suggests the proximity of a breeding site. From the first millennium AD, with development of commercial fishing and trade, gannets might have been traded and brought back from far breeding colonies. However, this applies to Britain, where known colonies existed on off-shore islands. The taking of the Northern Gannet by humans is restricted to breeding colonies, so even if the bones and bodies were brought to Palagruža by humans, it would suggest the existence of breeding colonies in that area. Until now, there were no findings of gannet presence from that period anywhere in the Mediterranean. In the Adriatic Sea, Palagruža, together with a few small islands in fairly close proximity, represents the most suitable breeding site for that species. The presence of gannet bones on Crete was interpreted previously also as an indicator of its breeding (Weesie, 1988). Contrary to this interpretation, Panteleyev (2001) considers Northern Gannet fossils from Crimea as an indication of the species’ presence during non-breeding season only. The remains were found in the settlements and no indication of nearby cliffs or islands suitable for breeding was mentioned.
Today, large colonies of Northern Gannet are often homogeneous. However, small groups share breeding cliffs with different seabirds, including common guillemot Uria aalge, black-legged kittiwakes Rissa tridactyla, cormorants Phalacrocorax carbo and Shag Phalacrocorax aristotelis but not gulls Larus sp. (Barrett, 2008; BWPi 2.0.1, 2008). The Yellow-legged Gull is nowadays the most numerous seabird species in the Adriatic and Mediterranean as well as on Palagruža. However, its remains in studied samples were restricted to a single specimen. This might suggest the Yellow-legged Gull was not numerous on the island, possibly excluded by competition with the Northern Gannet.
Today, the Northern Gannet is present in the Mediterranean mostly during autumn and winter, and is much more common in its western part. It is a rare vagrant in the Adriatic Sea, with only three confirmed recoveries along Croatian and four along Slovenian coast during the 20th and 21st centuries (Genov and Malek, 2014; Hanžel, 2013; Hanžel and Šere, 2011; Kralj, 1997; Kralj and Radović, 2002). It is more frequent, although not a regular visitor, of the Italian Adriatic coast, with up to seven birds recorded annually (MORUS-Progetto Uccelli Marini, 2009–2012). Since 1990s, several breeding attempts in the west Mediterranean, along the southern coast of Provence (France), were recorded. Birds attempted nesting on man-made structures: floating docks, boats and yacht decks. Few pairs started to breed regularly at two sites in Provence and in Porto Venere (Italy) (Giagnoni et al., 2015).
Radiocarbon dates obtained for two bird bones suggest presence of Northern Gannet during periods for which it is not possible to confirm human presence on the island, at least not through archaeology. However, dates are given for only two bones and cannot be considered to cover the whole period of gannet’s presence on the island, while the lack of archaeological material cannot prove that the humans have not visited Palagruža during that period. Even though we cannot exclude the possibility that human activities, through direct persecution, disturbance and/or fishing activities, could have had a negative impact on gannets, changes in the environmental conditions, such as sea-surface temperatures, oxygen accessibility, salinities and temperature differences, between the surface and the bottom water, as well as distributional changes of prey could lead to extinction of the Northern Gannet as a breeding species in the Mediterranean.
The global breeding distribution of seabirds is generally considered to result from world’s latitudinal zonation of temperature, which is most clearly seen in the southern oceans, where seabirds breeding distribution appears to be directly related to availability of breeding sites and water temperature, as a determinant of marine food resources (Lufbery and Mougin, 1985; Zotier et al., 1999). It was shown that the productivity in the eastern Mediterranean was higher during the Pleistocene than during the Holocene. The oxygen depletion during the Holocene resulted from climate oscillations (Gallego-Torres et al., 2007). At some 3000 yr BP, the sea-surface temperature of the southern Adriatic Sea was about 6°C lower than the Ionian Sea (Drake, 2012), thus possibly providing better breeding opportunities for seabirds. The overall sea-surface temperatures in the southern Adriatic Sea during the Holocene were lower than current ones and ranged from 13.8°C to 16.8°C, with the lowest temperatures during two cooling events 6000 and 3000 yr BP (Piva et al., 2008; Sangiorgi et al., 2003). Furthermore, two short-lived low salinity events, attributed to major influences of freshwater from the Po River and other coastal rivers, occurred around 3000 and 2000 cal. yr BP (Siani et al., 2013). Such short-term hydrological change could have affected the sea-surface hydrology and consequently reduced the formation of deep bottom waters in the south Adriatic Sea affecting the thermohaline circulation in the Mediterranean Sea, resulting in diminished productivity. A combination of these environmental conditions can affect the distributional patterns and aggregation areas of fish (Maravelias and Reid, 1997), which, in turn, might determine seabirds distribution (Montevecchi and Myers, 1997; Weimerskirch et al., 1994). Distributional changes of prey were identified as possible causes of the reduction of range in other seabirds, such as albatrosses (Eda and Higuchi, 2004).
Conclusion
From the late-Holocene bird assemblage from Palagruža, it is obvious that two groups of species were present, as they are present today: small number of breeding seabird species and several passage birds that use Palagruža as a stopover site during flight across the Adriatic Sea.
Although the Northern Gannet’s breeding on Vela Palagruža was not confirmed, its breeding on the island is presumable. The remains from the Adriatic are some 6000 years younger than other findings of that species in the Mediterranean, confirming its presence in late-Holocene. Further excavations on the other off-shore Adriatic islands are necessary to determine whether Gannet was present in the wider area. Eventual additional remains might help to confirm breeding status and the period of its presence in the Adriatic.
Footnotes
Acknowledgements
We would like to thank Dr Branko Kirigin from Archaeological Museum in Split for providing us with bird bones for the analysis, Dr Anita Gamauf from Natural History Museum of Vienna for reference modern bird bone material and Dr Jane Sanford Gaastra for information about Palagruža mammal assemblage. We would like to express our gratitude to Dr Nicola Baccetti for information about Northern Gannet breeding in Italy and to Dr Igor Boršić for information on floristic characteristics of Palagruža. We would also like to thank Dr Stašo Forenbaher who generously provided us with a photo of Palagruža and Dr Tomislava Bošnjak Botica who translated the Panteleyev’s article. We would like to thank Dr Marco Pavia for useful comments on the first version of this manuscript. We express our gratitude to Dr Davorka Radovčić and an anonymous reviewer for helpful comments that improve our text. We thank Dr Dinko Relković for his contribution in text editing. Authors Ankica Oros Sršen and Jelena Kralj contributed equally to this work.
Funding
This work was supported by the Foundation of Croatian Academy of Sciences and Arts (Grant/Award no. 219), the Institute of Ornithology and the Institute for Quaternary Paleontology and Geology of Croatian Academy of Sciences and Arts.