
Abstract
In the early-Holocene, animal domestication processes entailed important changes to the subsistence strategies of Neolithic populations. Among the first domestic species, pigs played a key role as they soon came to be one of the main sources of meat. Several methodological approaches have been followed in archaeology to differentiate between wild and domestic forms in the faunal remains found at early Neolithic sites. Among these, biometry is essential. The biometric analysis applied to a significant sample of Sus domesticus and Sus scrofa remains from 53 sites in the Iberian Peninsula dated between 10,000 and 800 cal. BC reveals differential dynamics between the wild and domestic forms resulting from changes in the climate during that time and the increasingly systematic selective pressure of husbandry. Whereas the wild animals increased in size, the inverse tendency is documented in the domestic population, which gradually decreased in size after the early Neolithic. The point of greatest divergence is seen in the Bronze Age. Significant differences are also documented in different geographic areas, which corroborates the influence of climate on the physical characteristics of wild populations. The range of variability in each population also differs chronologically as it is relatively greater in the Neolithic, which may be connected with the existence of different ways of adopting and breeding domestic pig among the first Neolithic communities, some of which may have involved continuous cross-breeding between the two populations. The results are an initial point of reference for the classification of archaeological remains of prehistoric pig in the Iberian Peninsula, a key area for the study of the dynamics of neolithisation.
Introduction
The dynamics of animal domestication processes in the Iberian Peninsula are being studied profusely at the present time particularly because of the contribution this topic can potentially make towards understanding the neolithisation phenomenon in the western Mediterranean. Several millennia separate the first material manifestations of neolithisation in the Near East and in Iberia as the presence of the first domestic animals is documented millennia later in Iberia than in the former region. An eastern origin has traditionally been claimed for these first domesticates. However, recent publications stress the possibility of the domestication of autochthonous species in the case of bovids (Wright et al., 2014) and suids (Larson et al., 2005; Vai et al., 2015). For the latter species, palaeogenetic studies have identified several independent areas of domestication, which suggests the hypothesis of repeated domestication actions, spatially and temporally. Studies based on geometric morphometrics correlated with palaeomolecular analysis of a large collection of pig molars found at Neolithic sites in the Balkan Peninsula equally indicate that the domestication of this species may have included numerous modalities, with a continuous flow between wild and domestic populations (Evin et al., 2015). The possibility also of access, through exchange or hunting feral animals, to domestic animals by Mesolithic hunter-gatherers in southern Scandinavia and northern Germany, suggested by mutations detected in the MC1R gene associated with the colour of the coat (Krause-Kyora et al., 2013), is further evidence that animal domestication or the inclusion of domestic animals in subsistence was the result of diverse pig management strategies, some of which may have led to their domestication.
This historical issue of the first experiences in animal domestication has been studied through different perspectives (Saña, 1999, 2005; Vigne, 2015; Zeder, 2012). Whereas in zoological or environmental approaches, the emphasis is placed on the animal per se; in historical approaches, the weight of the analysis falls on the social formation responsible for the process, without whom the transition from wild to domestic condition would have been impossible. In the framework of the latter approach, several views can be differentiated (Saña, 1999, 2005). From the economic viewpoint, the conceptualisation of the animal as a resource has led to the domestication process being understood as a continuous series of stages each defined by the specific type of relationship existing between society and the animal resources. However, it is still difficult to determine in which stage the animal can be regarded as domestic. Although integral analytical approaches (biometry, stable isotopes, palaeogenetics and geometric morphometrics; Vigne et al., 2005) are now able to identify stages or situations that had been archaeologically invisible, by themselves their resolution is not sufficient to determine the initial animal domestication. This is only possible through a social approach embracing all the changing relationships that would allow continued control over reproduction and restricted access to animal resources.
The data currently available about suids allow the proposal of several trajectories through which the control of their reproduction and therefore their domestication would be feasible (Dwyer, 1996; Nakai, 2012; Rowley-Conwy, 1995, 2001; Sillitoe, 2007). Some of these models were designed bearing in mind the ethnographic studies that have documented a wide range of strategies in the management of the reproduction of pigs in the initial phases of their domestication (Baldwin, 1979; Nakai, 2012; Yen, 1991). Zeder (2012) also suggests joint commensalism as a scenario leading to the domestication of this species. In this case, the greater sedentarism of populations in the early-Holocene may have encouraged wild boar to approach human settlements. However, the key issue lies in the criteria followed in the selection involved in reproduction, widely dependent on the strategies of adoption and maintenance of animals that are applied. According to the degree of interdependence between the wild population and the one managed directly by human groups, the results obtained in the ethnographic studies cited above can be summarised in three main scenarios. First, human action is limited to capturing infant or juvenile specimens that were born in the wild. In the second, reproductive females are monitored continually, and in the third, both males and females depend entirely on humans. In some situations, therefore, the flow between wild and domestic populations would be common (Marshall et al., 2014). This is especially true taking into account that because of their feeding habits and behaviour, the breeding and maintenance of this species can be based on extensive practices in which the animals do not live in confined or enclosed spaces (Albarella and Trentacoste, 2011; Hadjikoumis, 2012; Redding and Rosenberg, 1998).
The possibility of hybridisation between populations is therefore continual (Ramírez et al., 2015). This adds certain complexity to the study of initial animal domestication, as its archaeological representation based on faunal remains may be highly variable and difficult to identify. Several studies have underscored the difficulty in separating wild and domestic forms in the case of pigs (Mayer et al., 1998; Payne and Bull, 1988). Additionally, such aspects as the different degrees of inter-relation and mixing between the two and the potential presence of feral specimens should be considered. One of the criteria most often used in this situation is based on biometry (Albarella et al., 2006b; Evin et al., 2014; Mayer et al., 1998; Payne and Bull, 1988; Rowley-Conwy et al., 2012), although recently numerous analytical methods such as palaeogenetics (Krause-Kyora et al., 2013; Larson and Burger, 2013), stable isotopes and geometric morphometrics (Balasse et al., 2016; Evin et al., 2015) have been proposed.
In the case of the Iberian Peninsula, no integrated study has examined the variability in pig populations in the early- and middle-Holocene. Only the studies of Albarella et al. (2005, 2009), Davis and Mataloto (2012) and Davis and Detry (2013) integrate data from Portugal in order to examine the dynamics in the changes in size of wild and domestic pigs over a long period of time. Hadjikoumis (2010) also carried out a full study of pig farming in the post-Neolithic period in his doctoral thesis. More precisely, Altuna and Mariezkurrena (2011) have published a summary for northern Iberia based on the biometric differentiation between Sus scrofa and Sus domesticus, from the Upper Palaeolithic to the Middle Ages. The results prove the existence of size differences between the two populations that can be identified in archaeological remains. In the Iberian Peninsula, the criteria most often used to discriminate between the wild and domestic species have been biometrics, slaughtering patterns, morphological traits and the archaeological context. Some authors stress the implicit limitations when the assemblages belong to the early Neolithic (Altuna, 1980; Altuna and Mariezkurrena, 2011; Castaños, 1984, 1991).
In order to reconstruct the history of suid populations (Sus scrofa/Sus domesticus) and their management in Iberia during prehistory, this paper presents an exhaustive analysis of the biometric data currently available for different time periods. The goal is to infer the forms of management of these species between 10,000 and 800 cal. BC. In first place, the variability of Sus scrofa from the Epipalaeolithic to the Bronze Age is assessed and later compared diachronically with the characteristics of the domestic populations. The explanatory models formulated in Iberia in relation to the pig domestication process envisage a complex and varied situation in the early-Holocene, in accordance with the large and contrasting environmental variability in the peninsula. The empirical record reveals the existence of a few sites in the north of Iberia specialising in hunting wild boar in the early Neolithic, whereas at other sites domestic pig breeding was well established and hunting was very limited (Saña, 2013). Published biometric studies for particular sites suggest discontinuity between wild and domestic populations, which might be linked to the widespread adoption of the domestic form at most human settlements. However, given the diversity of scenarios that have recently been proposed and identified in other geographic areas and the methodological difficulties in differentiating between Sus scrofa and the first domestic forms, it is necessary to study this phenomenon diachronically from a territorial perspective to be able to infer the diversity of situations in which the domestication and adoption of domestic pigs took place.
Materials and methods
This study is based on the exhaustive statistical analysis of the biometric data in the eastern part of the Iberian Peninsula, as most of the sites with published archaeozoological studies are located in that area. The sample consists of 53 archaeological sites in the north and east of the peninsula (Figure 1), covering a time from 10,000 to 800 cal. BC (Table 1). The inclusion of sites in each period has respected the criteria followed by different researchers, using the general chronocultural systematisation for the Iberian Peninsula. Only data for adult individuals at well-dated sites with no taphonomic problems have been used. The systematisation of all the measurements enables a continuous interpretation of the data temporally, without any significant gaps of periods with no data. For the measurements, the standardised criteria published by von den Driesch (1976) have been followed, including cases in which Payne and Bull’s (1988) proposal has been possible. The attributions of the remains to Sus scrofa or Sus domesticus made by the different researchers have been followed, while remains with no specific attribution have also been included in the analysis for comparison, noting that this category is only represented at some Neolithic and Chalcolithic sites and that the sample of measurements is small and circumscribed to a few sites.
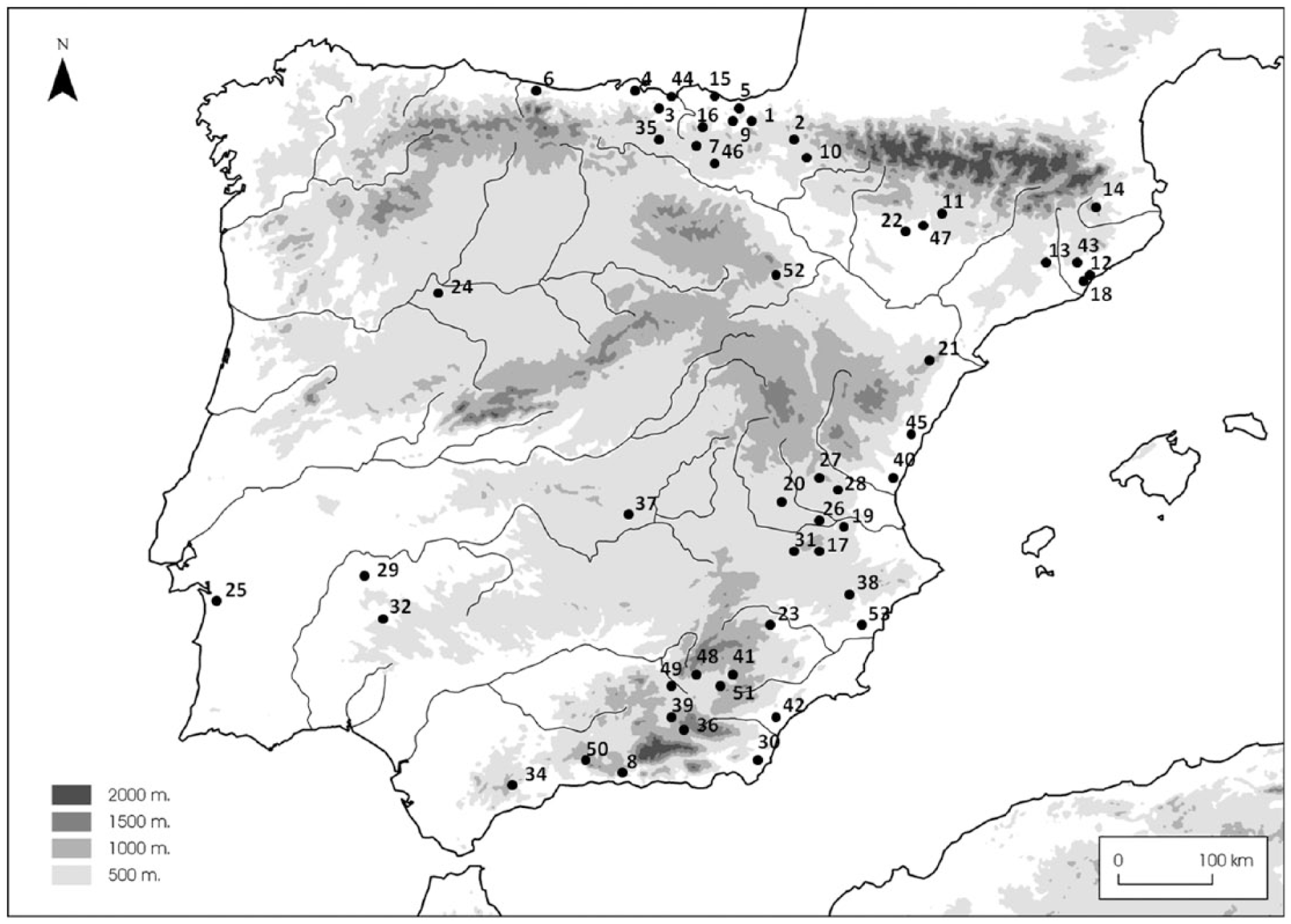
Map with the location of the archaeological sites included in this study.
Information about the sites, chronology and characteristics of the sample.

SC: Sus scrofa; SD: Sus domesticus; Sus sp.; n: number of measurements.
The data have been compared with the log ratio technique (Meadow, 1999), using the data published by Albarella and Payne (2005) as the theoretical population. The comparison used the following logarithmic formula: (log x − log m) = log (x/m), where x is the measurement of the archaeological sample and m is the measurement of the standard population. Based on the recommendations from several studies, wherever possible, dental remains have been studied separately from the post-cranial skeleton (Albarella et al., 2005; Rowley-Conwy et al., 2012). Based on the log size index (LSI) values, in order to compare the data according to chronology and geographic area, the Kruskal–Wallis test has been used, with a statistical significance probability threshold of α = 0.05 (Hammer et al., 2001). The distribution of the measurements in box plot diagrams has enabled the assessment of continuity and discontinuity between wild and domestic populations bearing in mind that the interquartile range and median are represented in the box. Finally, multivariate statistics have been applied to assess the data integrally. The procedure has been cluster analysis, with which the degree of similarity between the data has been evaluated to determine whether or not the population can be divided into different size sub-groups. To apply this latter procedure, the databases have been formed by counting the LSI values according to chronology for each of the samples.
Results
The results of the Sus scrofa sample are given first, comparing the variability obtained diachronically. Then, the characteristics of the Sus domesticus populations are described, and finally, the data are compared and analysed together.
Variability in Sus scrofa in the Iberian Peninsula from 10,000 to 800 cal. BC
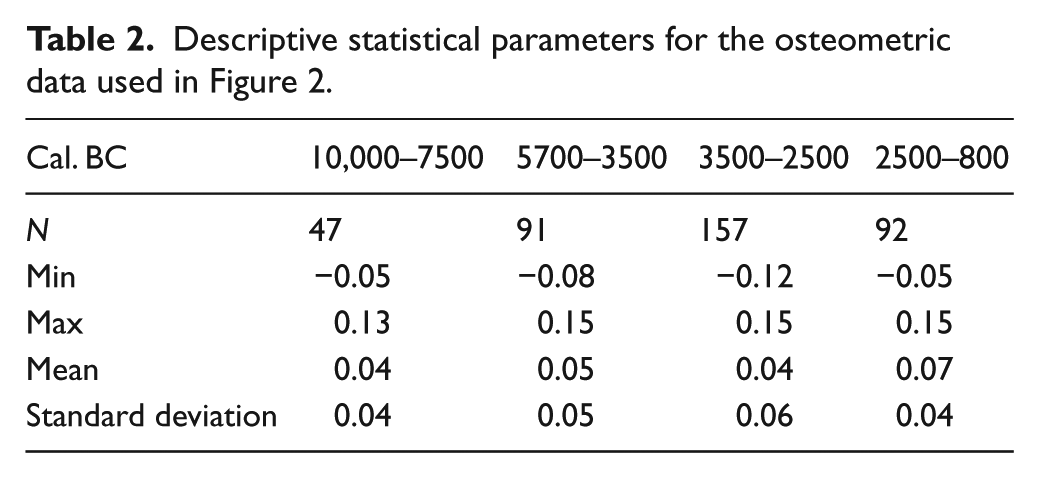
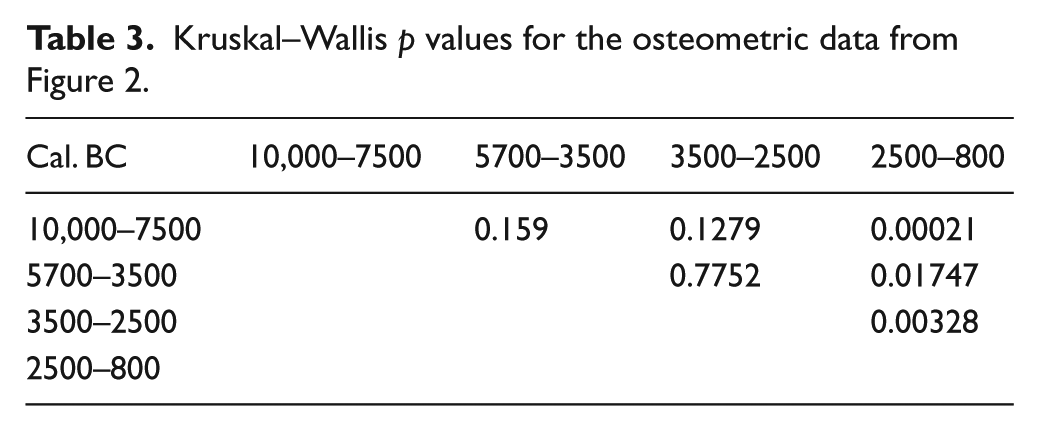
First, the variability in Sus scrofa populations has been compared according to chronology and based on the log ratio values obtained, using box plot charts (Figure 2). The results reveal differential dynamics in wild boar size during prehistory, in agreement with the values of the descriptive statistical parameters obtained for each sample (Table 2). Between 10,000 and 5700 cal. BC, Sus scrofa displays greater homogeneity and a smaller size than in the Neolithic (5700–3500 cal. BC). After that time, the tendency is towards a gradual increase in wild boar size until the Bronze Age (2500–800 cal. BC). This trend is clearly reflected in the mean values of each population, ranging from 0.04 in the Epipalaeolithic to 0.07 in the late Bronze Age. The variability in each time interval also differs, with a maximum in the Chalcolithic (SD = 0.06). Kruskal–Wallis p values (Table 3) attest statistically significant differences between Bronze Age populations and those in other chronological intervals. They also indicate continuity in size dynamics from the Epipalaeolithic to the Neolithic. Based on these results, a statistically significant turning point may be proposed, located temporally between 2500 and 800 cal. BC, when this species reaches its largest size. In order to assess the role that geography and climate may have played in this variability, the log ratio values for the time intervals corresponding to the Neolithic and Bronze Age have also been compared, differentiating between the osteometric data in northern Iberia and in the Mediterranean region. The data for the rest of periods have not been represented as the number of measurements is too small for some areas. The results (Figure 3) show that the Mediterranean specimens were relatively smaller than the ones in the north. However, in both the Mediterranean and northern regions, the size of the specimens increases between the early Neolithic and the Bronze Age. Kruskal–Wallis p values (Table 4) show that differences in the northern population between the Neolithic and Bronze Age are not significant.
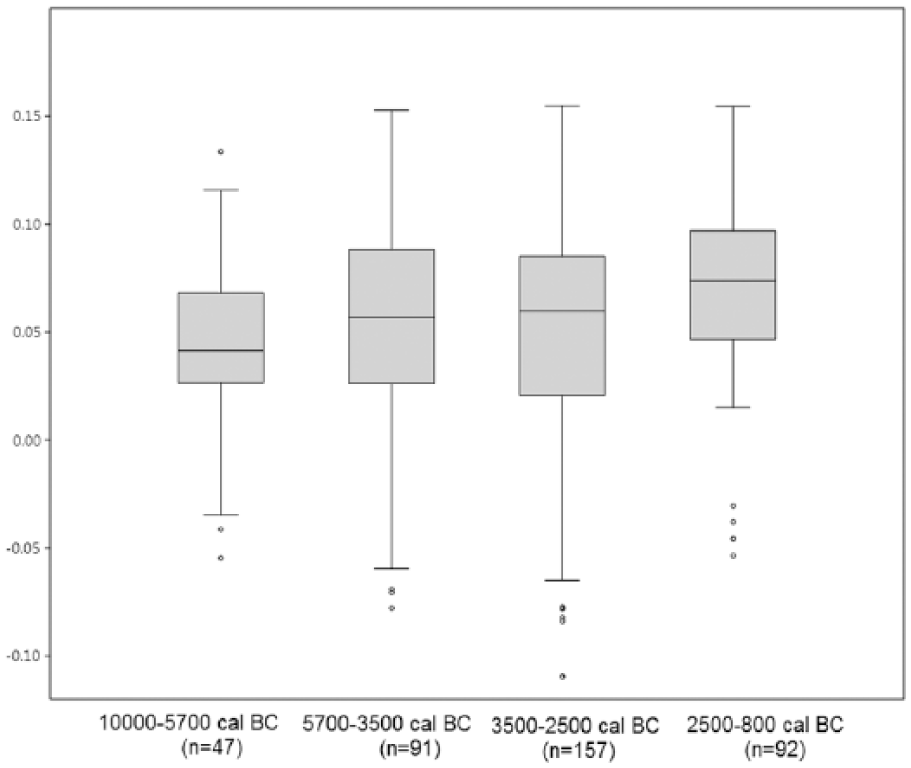
Box plot diagram with the Sus scrofa LSI values in chronological order.
Descriptive statistical parameters for the osteometric data used in Figure 2.
Kruskal–Wallis p values for the osteometric data from Figure 2.
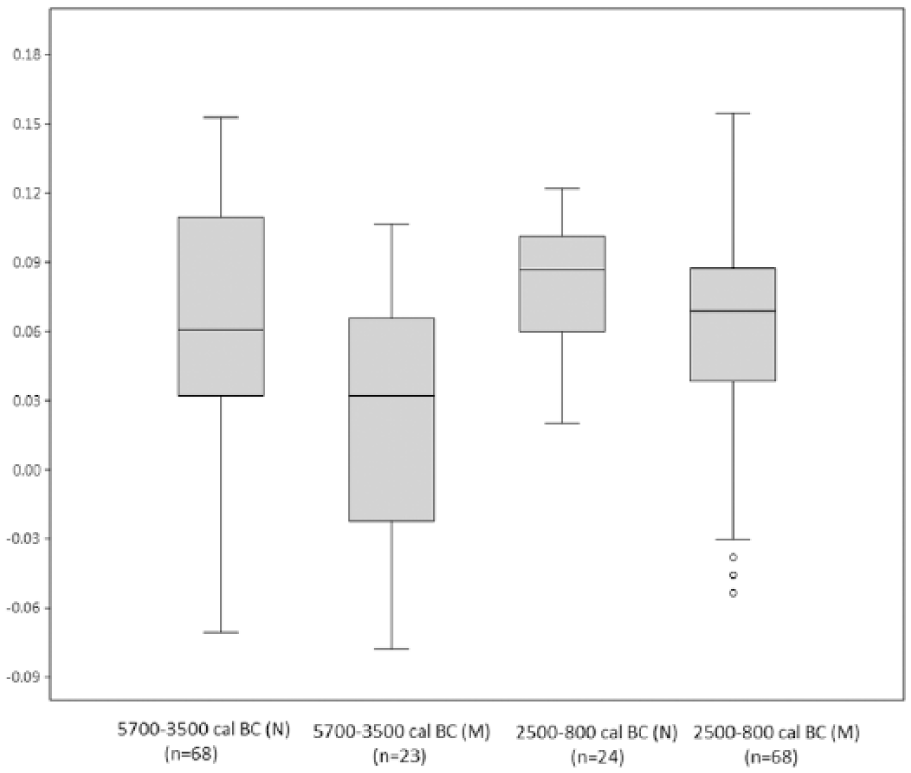
Box plot diagram with the Sus scrofa LSI values, differentiating between the north-east zone (N) and the Mediterranean area (M) for the Neolithic and Bronze Age.
Kruskal–Wallis p values for the osteometric data from Figure 3.

Finally, as differences in the dynamics of growth and plastic modification in elements of the post-cranial skeleton and teeth can affect the size and form of the animal differentially, as a consequence of different nature of the pressures, the data have been itemised according to that criterion (Figure 4). The results show greater relative variability in the post-cranial elements, where the Neolithic populations display the minimum and maximum values. It should equally be noted that between the Epipalaeolithic and early Neolithic, the size of teeth decreases relatively, while the other skeletal elements display the inverse trend.
Box plot diagram with the Sus scrofa LSI values, differentiating between the values obtained for post-cranial skeletal elements (B) and teeth (T) in chronological order.
Variability in Sus domesticus in the Iberian Peninsula from 10,000 to 800 cal. BC
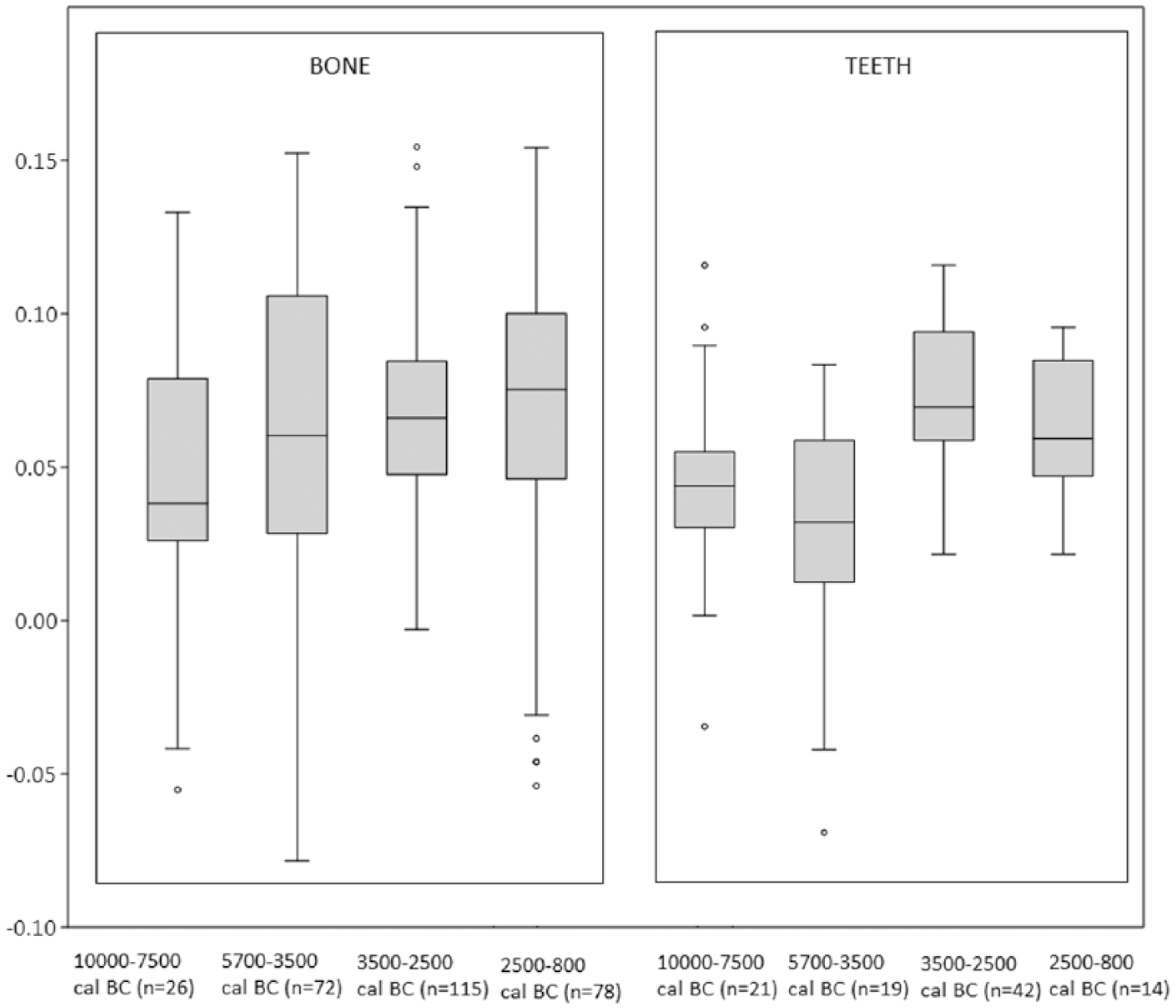
The number of measurements obtained for Sus domesticus in each time interval is larger than for Sus scrofa. The distribution of the log ratio values in box plot charts (Figure 5) indicates differential dynamics for the populations in each time interval. However, significant similarity is seen in the mean size of domestic pigs between 5700 and 2500 cal. BC. After this time, pig sizes tend to decrease, although with a greater degree of variability, similar to the situation in the early and middle Neolithic (Table 5). Kruskal–Wallis p values (Table 6) attest significant differences in the size of domestic pigs between the Neolithic-Chalcolithic and the Bronze Age. Specimens from the north and east of the peninsula have also been differentiated for Sus domesticus (Figure 6). In this case, differences in animal size in the two areas are not so great and are totally imperceptible during Bronze Age (Table 7). In all time intervals, greater variability is documented in the Mediterranean region. The distribution of the values according to skeletal element (Figure 7) displays great homogeneity in the measurements obtained for teeth and the post-cranial skeleton, although the post-cranial elements exhibit relatively higher means in all time periods (Table 8). However, Kruskal–Wallis p values (Table 9) reveal significant differences in the dynamics of bones and teeth in the various periods. The significant decrease in size documented after 2500 cal. BC affects both the post-cranial skeleton and teeth.

Box plot diagram with the Sus domesticus LSI values in chronological order.
Descriptive statistical parameters for the osteometric data used in Figure 5.

Kruskal–Wallis p values for the osteometric data from Figure 5.


Box plot diagram with the Sus domesticus LSI values, differentiating between the north-east zone (N) and the Mediterranean area (M) for the Neolithic and Bronze Age.
Kruskal–Wallis p values for the osteometric data from Figure 6.


Box plot diagram with the Sus domesticus LSI values, differentiating between the values obtained for post-cranial skeletal elements (B) and teeth (T) in chronological order.
Descriptive statistical parameters for the osteometric data used in Figure 6.
Kruskal–Wallis p values for the osteometric data from Figure 7.

Integrated analysis of pig population in the Iberian Peninsula, 11,500–800 cal. BC
Following the analytical procedure applied individually for Sus scrofa and Sus domesticus, the log ratio values have been represented jointly according to time intervals in order to make comparisons (Figure 8a), also including the category Sus sp. for comparative purposes (Figure 8b). The chart shows a tendency towards a separation between the two populations, especially after 2500 cal. BC, when the distributions of values of wild and domestic populations barely overlap. The most homogeneous populations would be those between 3500 and 2500 cal. BC, for both Sus scrofa and Sus domesticus. In contrast, the greatest variability in wild animals is seen in Neolithic populations, and in the Bronze Age for domestic pigs. The specimens corresponding to the category Sus sp. (Figure 8b) occupy an intermediate position, with values that are relatively closer to the domestic population in both the Neolithic and Chalcolithic. Kruskal–Wallis p values display similarities between this population and the Sus domesticus population in the Neolithic (p = 0.1) and Chalcolithic (p = 0.16). The general pattern obtained in Figure 8a is also represented in the results obtained in the cluster analysis of Euclidean distances (Figure 9), differentiating the wild species and the domestic pig based on two groupings. In the case of Sus scrofa, as the Kruskal–Wallis p values indicated, the most significant turning point was in the Chalcolithic. For Sus domesticus, the dynamic follows the same trend, with animal sizes relatively more similar between 7500 and 2500 cal. BC than in the Bronze Age.

(a) Box plot diagram comparing the Sus scrofa and Sus domesticus LSI values in chronological order. (b) Comparison of the variability of the category of Sus sp. in relation to Sus scrofa and Sus domesticus.
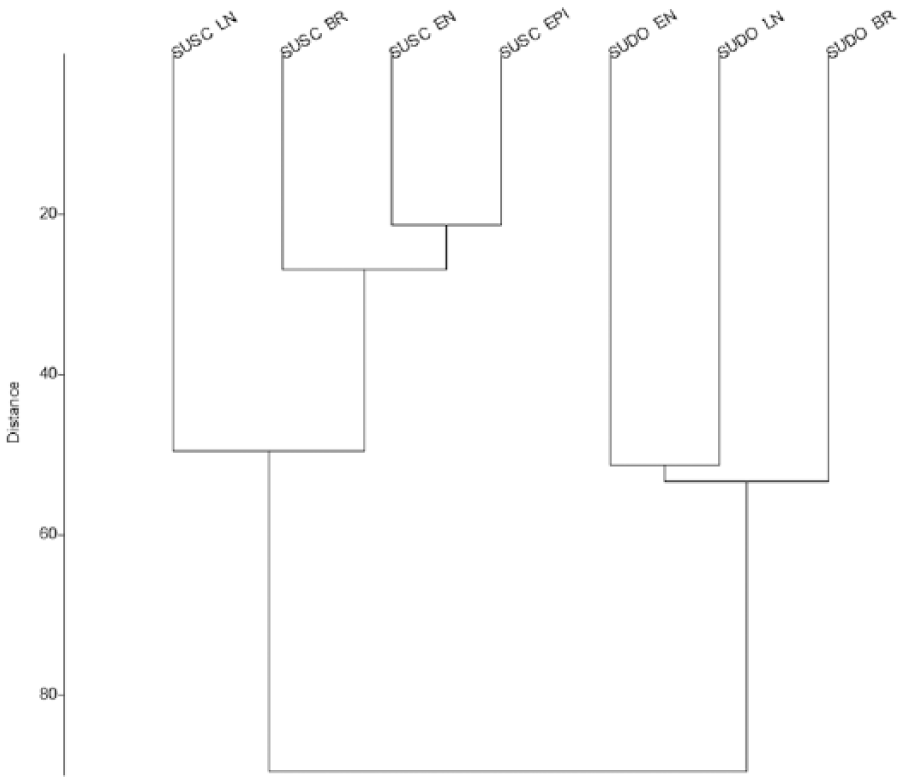
Cluster diagram with the distances between the LSI values according to species and chronology (SUDO: Sus domesticus; SUSC: Sus scrofa).
Discussion
The size variability of pig populations in prehistory reflects the different selective pressures they were subjected to, either environmental or social. The documentation of different dynamics for Sus scrofa and Sus domesticus during the period studied here clearly demonstrates this. Whereas in the Holocene wild populations tended to increase in size, domestic populations decreased and the maximum difference between the two was reached after 2500 cal. BC.
The increase in size of Sus scrofa in the Holocene has been documented in other parts of Europe (Albarella et al., 2006a, 2006c; Rowley-Conwy, 1995), and particularly in Portugal during the Chalcolithic (Albarella et al., 2005; Davis and Mataloto, 2012). Several reasons have been put forward for this, such as the temperature and climate, and relaxation in selective pressure on this species related to hunting activity (Albarella et al., 2005, 2006b; Davis and Detry, 2013; Davis and Mataloto, 2012). According to these authors, the greater pressure exercised on the species in the Mesolithic would be a consequence of demographic growth, and later the stress caused by the intensive hunting of the species decreased because of the availability of domestic animals, from which most of the necessary proteins would be obtained. The inverse correlation between climate and size has been attested by a number of authors in the case of pig (Davis, 1981; Davis and Simões, 2016; Magnell, 2004; Mayer et al., 1998; Rosell et al., 2001; Rowley-Conwy et al., 2012). This is also evidenced in this study on a small scale by comparing the samples from northern Iberia with those from the Mediterranean area. The mean size in the latter area is slightly smaller, especially for the older wild boar populations (Epipalaeolithic/Mesolithic). The difference decreases from 2500 to 800 cal. BC, possibly as a result of the species adapting to the anthropic environment.
The greater relative variability in both Sus scrofa and Sus domesticus populations in 5700–3500 cal. BC coincides with the initial domestication or adoption of the species in the Iberian Peninsula. The greater difficulty in discerning between the remains of the two species is a factor that may have influenced this result. Aspects derived from the different forms of domestication or integration of the domestic species in subsistence strategies of the last hunter-gatherers and first Neolithic communities should also be considered. The exhaustive analysis of the data on the management of pigs in Iberia in the early Neolithic indicates that different forms of husbandry were practised (Saña, 2013). At the same time, as hunting Sus scrofa intensified at some Neolithic cave sites in the north, such as Zatoya (5743–4590 cal. BC) and Marizulo (5214–4718 cal. BC), in other areas intensification in pig farming has been documented. One of the clearest examples is the open air settlement of La Draga (5200–4720 cal. BC) in north-east Iberia, where a pattern of the selective slaughtering of males at the time of their meat optimum, in order to maximise meat production, has been recorded (Saña, 2011, 2015). This practice has also been proposed, based on demographic patterns, at the Neolithic site of Cueva de Chaves in Aragon (5740–5490 cal. BC; Castaños, 2004). While at La Draga biometry has proven the presence of two groups of different sizes, one of them coinciding with a domestic population (Saña, 2011), the size of the pigs at the early Neolithic site of Cueva de Chaves is relatively larger, similar to a wild population, and according to Castaños (2004), it may represent an example of mixed breeding.
Several factors may have contributed to this greater variability. During the initial phases of domestication and husbandry, crossing domestic animals with wild specimens may have been common, whether intentional or not. Ethnographic studies have described extensive strategies in pig farming, and such strategies favour cross-breeding (Albarella et al., 2006a; Marshall et al., 2014). Palaeogenomic studies on this species (Larson and Burger, 2013) stress that in the different stages of domestication, phenomena of admixture and introgression would be common. As genome data are available for more animal species, cross-breeding is seen to have been a common phenomenon (Orozco-Terwengel and Bruford, 2014). The greater degree of relative variability seen in Sus scrofa between 7500 and 3500 cal. BC compared with older (11,500–5700 cal. BC) and more recent chronologies (3500–800 cal. BC) may also be indicative of this situation, as the phenotype variability increased as a consequence of hybridisation (Šprem et al., 2011). An increasing number of studies have shown that after domestication, pigs can again acquire the characteristics of the wild population through the phenomenon of feralisation (Giuffra et al., 2000). Although it is now possible to follow and identify these situations with geometric morphometrics and palaeogenetics (Evin et al., 2015), in the present case, biometry is able to approach the matter only in terms of variability. In this respect, considering the variability and overlapping seen in the early Neolithic, the potential contribution of Iberian Sus scrofa populations to the formation of domestic herds cannot be ignored.
Other factors often cited in relation to the decrease in size of specimens during the initial phases of domestication include changes in mobility patterns due to constraints on movement (Lasota-Moskalewska et al., 1987) and a poorer diet and increase in stressful situations (Davis and Detry, 2013; Dobney and Ervynck, 2000). These situations would all arise with more frequent direct contact between pigs and human settlements. Archaeological data are unable to determine the number of animals in the first domestic herds. The practice of strategies based on semi-freedom would favour mixing or contact with wild animals, and cross-breeding between animals with a different status would be common (Bartosiewicz et al., 2006; Nakai, 2012), which would result in transitional phenotypes (Benecke, 1994). Several scholars agree that the systematic practice of breeding in confinement compared with a more extensive management would not have become established until the Middle Ages (Albarella et al., 2009; Ervynck et al., 2007; Thomas et al., 2013). Given the wide ecological adaptability of Sus scrofa (Scandura et al., 2008), if animals approached or entered settlements, they would have added farming sub-products or the waste of human consumption to their diet (Fraser et al., 2013). Results of carbon and nitrogen isotope studies reveal significant changes in the diet of pigs between the Neolithic and Iron Age (Hamilton et al., 2009), and a trend towards greater diversification has also been noted between the Neolithic and the Bronze Age (Madgwick et al., 2012). The seasonal amount and availability of food are key variables directly affecting the rates of growth and reproduction of animals (Férnandez-Llario and Mateos-Quesada, 1998; Halstead and Isaakidou, 2011). In this way, the gradual decrease in size of Sus domesticus from the Neolithic would be the result of increasingly constant selective pressure to which the species was subjected through a direct control over its feeding, raising and reproduction by human communities.
This decrease becomes more evident after 2500 cal. BC. As can be seen in Figure 8, the difference between the sizes of Sus scrofa and Sus domesticus in the Iberian Peninsula increases after 3500 cal. BC at the same time as the degree of variability in Sus domesticus populations decreases. In some Bronze Age settlements, such as Fuente Álamo and Cerro de la Encina, the difference between the wild and domestic forms is particularly clear (Driesch, 1987; Driesch et al., 1985). Once domesticated and subjected to conditions of artificial husbandry, the possibilities of genetic isolation between the wild and domestic populations are greater (Rowley-Conwy et al., 2012). In this context, the relatively more direct selection by Neolithic communities of animals for consumption and reproduction would have contributed in the medium or long term towards modifying the natural demographic patterns of the species. Control over reproduction patterns would undoubtedly have been a key variable, favouring significant changes in the growth rate and sexual maturity of the species. The trend for Sus domesticus seen in the box plot diagrams suggests that these selective pressures would have been increasingly intense. However, they probably occurred at different rates and in different ways depending on the means and needs of each prehistoric community. They are therefore not linear and were not the same in the whole of the Iberian Peninsula.
This differential dynamics in the management of domestic pigs, with unequal degrees of intensity and different directions of selective pressure, would probably have contributed to increasing variability in the species, as the results for the Bronze Age have shown. It should be noted that the relative homogeneity documented in Sus domesticus samples between 3500 and 2500 cal. BC may be conditioned by the small number of sites with remains that have been studied corresponding to this period. However, the trend towards a decrease in size is confirmed later, in the Bronze Age, with a mean of −0.04 compared with −0.02 in the preceding time interval. In this way, the control exercised on domestic populations led to a relaxation in the mechanisms of natural selection. This may explain the differential dynamic seen in sizes of wild boar and domestic pig in the north compared with the Mediterranean area, while the difference is practically imperceptible for domestic pigs in 2500–800 cal. BC. Differences in the mechanisms of selection are also observed in the individualised study, when possible, of dental remains and the post-cranial skeletal elements. Greater homogeneity is attested in the case of domestic populations. Studies of more recent samples, dated in the Roman, medieval and modern periods, show that after the Roman age, pig sizes vary significantly, with a large relative decrease in size in the Middle Ages and an increase in the modern age. This increase is attributed, in both the modern and Roman periods, to the application of zootechnical improvements (Grau-Sologestoa, 2015).
Conclusion
The study of animal domestication processes through analytical procedures based on biometry reveals the difficulty in explaining the diversity of situations and interactions that may have occurred between human communities and pig populations in the early- and mid-Holocene. It is clearly necessary to examine the domestication phenomenon through large conceptual categories that are not based exclusively on the wild/domestic dichotomy (Evin et al., 2015; Rowley-Conwy et al., 2012; Saña, 1999; Vigne, 2015; Zeder, 2012). Biometric analysis of the archaeological remains of Sus scrofa and Sus domesticus recovered at Iberian sites dated from 10,000 to 800 cal. BC shows that it was precisely during the early Neolithic (5700–3500 cal. BC) when the archaeological representation of pig management strategies displayed greatest variability. This should be associated with the economic and social changes derived from the adoption of livestock farming, which emphasised anthropic pressures in comparison with ecological ones. The different trajectories documented in the changes in size of Sus scrofa and Sus domesticus indicate that the adoption of pig farming was a relatively rapid process. The great capacity of adaptation of this species undoubtedly helped its breeding in artificial conditions, and it soon became a basic component in farming strategies. Specialisation in the hunting of Sus scrofa by the last hunter-gatherer societies shows that they were fully aware of its requirements in the early-Holocene and that it was an important part of their diet. Indeed, farming and hunting were not mutually exclusive but complementary activities in the Neolithic. Unlike other animals, such as goats, sheep and cattle, pigs were kept exclusively for raw materials and meat. The control of meat supplies was probably a direct cause for them being raised in artificial conditions to ensure their reproduction. From that time on, as seen in the changes in the size of the animals, domestic and wild populations followed different paths that did, however, occasionally converge. As it is a species with relatively fast gestation cycles and growth rates (Rosell et al., 2001), the differentiation from the wild population would also be more rapid than in the case of larger mammals. The differences evidenced in teeth size compared with post-cranial skeletal elements also point to the existence of flow between populations, either by direct dislocations or influenced by human action. Additionally, environmental and climate changes in the Mediterranean area at the end of the middle-Holocene (Zanchetta et al., 2016) may have conditioned new adaptations, particularly in the wild population.
Therefore, multiple and diverse variables converged during the initial domestication and adoption of pigs by the first farming communities. It has been shown that Sus scrofa and Sus domesticus responded to the various factors in different ways, which will doubtlessly facilitate the study of the dynamic and management of pigs in recent prehistory through the analysis of archaeological faunal remains.
Footnotes
Acknowledgements
The authors thank the reviewers and the Editor, whose comments greatly improved the original manuscript.
Funding
This work was developed in the frame of the project HAR2014-60081-R (Producción animal y cerámica en el neolítico peninsular. Estudio biogeoquímico integrado del consumo y las practicas culinarias) funded by Ministerio de Economía y Competitividad, Spain.