
Abstract
Identifying evidence for hunter-gatherers in the palaeoenvironmental record is far from simple. Despite decades of research, few studies have demonstrated unambiguous palynological evidence of hunter-gatherers. This paper presents the results of high-resolution palaeoecological analyses of a peat sequence located within the vicinity of the pre-historic Yup’ik village of Nunalleq in southwestern Alaska. The aim of this research was to examine whether there are any discernible palaeoenvironmental impacts associated with the 15th–17th century occupation of the site. Presuming an ephemeral character to any palaeoecological signal, this study selected a sampling location approximately 30 m east of the limit of archaeological remains. Pollen, non-pollen palynomorphs and microscopic charcoal analysis were then used to generate a highly resolved (contiguous 1 cm) environmental history for the site. The results are striking and indicate that the activities of prehistoric Yup’ik hunter-gatherers at Nunalleq did leave a clear material trace in the palaeoenvironmental record. Through the application of high-resolution Pb210 and C14 dating and Bayesian modelling, these impacts were found to be concurrent with the occupation of the archaeological site. These findings suggest that not only can circumpolar hunter-gatherers leave a material palaeoenvironmental trace but that these traces may be used to accurately date such site activity in lieu of excavation.
Keywords
Introduction
The use of pollen analysis to detect signals of human activity in palaeoenvironmental archives is well established. Since Iversen (1941) first outlined evidence for the impact of agricultural societies on the forested environments of Denmark, palynological data have been deployed to illustrate the impact of agriculture in hundreds of palaeoenvironmental archives over a variety of timeframes and biomes across the globe (e.g. Deevey et al., 1979; Iversen, 1956; Ledger et al., 2014a; Li et al., 2009; Miras et al., 2015; Whitney et al., 2014). In contrast, records of the environmental impacts of hunter-gatherer societies have been less forthcoming (Edwards, 1998). Unlike agriculturalists – who are sedentary and actively change ecosystems by clearing or burning forests to plant crops and create grazing lands (Iversen, 1956) – hunter-gatherers are typically highly mobile and regularly move camp in order to exploit the seasonal flora and fauna of a region. Any palynological signal associated with such activity is therefore likely to be limited and highly localised, that is, only visible within a few hundred metres of the activity area (Bos and Urz, 2003). For many years, this fostered the perception that, except for increases in microscopic charcoal deposition, hunter-gatherers are near invisible in sedimentary records (Bos and Janssen, 1996). As a consequence, they have attracted far less interest than agriculturalists in palynological enquiry (Hicks, 1993).
Research on Mesolithic landscapes in central and northern Europe has not significantly altered this perspective. Despite numerous studies at varying spatial scales, an unambiguous signal for hunter-gatherers – comparable to that seen for farmers – has yet to emerge (Edwards, 1998). Most evidence centres on relative changes in tree pollen types, elevated deposition of microscopic charcoal and subtle increases in some herbaceous taxa (e.g. Edwards, 1995; Edwards and Sugden, 2003; Hornberg et al., 2006; Innes and Blackford, 2003; Ralska-Jasiewiczowa and Van Geel, 1992; Smith et al., 1989). A major difficulty is the likely small-scale and highly localised nature of any impacts, making finding them akin to searching for the proverbial needle in a haystack. Therefore, it has been argued that in order to identify hunter-gatherer activity in the palynological record, it is necessary to (1) know where such activity sites were and (2) examine profiles from highly localised depositional environments in close proximity to such activity areas (e.g. Bos and Janssen, 1996; Bos and Urz, 2003; Kuneš et al., 2008). Unfortunately, even this approach has had mixed results. While being sensitive to localised vegetation disturbance, depositional environments with small relevant source areas for pollen (RSAPs; sensu Sugita et al., 1999) are also dominated by local pollen deposition (Davis, 2000). In landscapes where trees, which are high pollen producers, were dominant, this poses problems as they have the potential to obscure any anthropogenic impacts (Bos and Janssen, 1996; Kuoppamaa et al., 2009). Consequently, even when profiles are derived from locations within the vicinity of known archaeological sites, difficulties have been encountered in identifying a clear signal for human activity beyond elevated concentrations of microscopic charcoal (e.g. Behling and Street, 1999; Woelders et al., 2016).
Forested environments therefore appear to represent a significant challenge to identifying hunter-gatherer impacts in palynological records, but what about more open landscapes? At circumpolar latitudes, the hunter-gatherer lifeway survived late into the recent historical period and the location of numerous sites is well known. Despite this resource of sites, few researchers have examined their potential. Hicks (1993) conducted one such study at a Sami winter settlement in northern Finland, identifying slight rises in Poaceae, other herbs and an increase in microscopic charcoal as a signal for hunter-gatherer activity. More recent work from North America has presented slightly more equivocal patterns. At sites in Labrador, the palynological signal for human impact associated with Thule Inuit occupation is weak, although macrofossil evidence indicates the expansion of apophytes such as Silene and Montia (Roy et al., 2012, 2015). In Newfoundland, Renouff et al. (2009) record similarly subtle palynological changes but identify evidence for the impacts of hunter-gatherers on limnological processes.
The open, treeless tundra landscape of the Yukon–Kuskokwim (Y-K) delta in sub-Arctic southwestern Alaska presents, in many ways, a similar geographic and cultural setting to these studies. The region is home to the Yup’ik who are part of the wider Inuit cultural and language group that dominates circumpolar North America from Alaska in the west to Greenland in the east (Dumond, 1987). A semi-sedentary hunter-gatherer lifeway, focused on exploiting marine mammals, fish and game and centred on permanent winter villages and seasonal summer camps, continued to be practised in this region until the mid-19th century (Oswalt, 1992). Modern vegetation surveys from abandoned settlements across the circumpolar north have revealed a discernible impact on the immediate environment whereby the floral composition of such sites is markedly different from that of the surrounding environment (Forbes, 1996; Fredskild, 1961, 1988; Porsild, 1932). Therefore, there is strong reason to suspect that any such impacts would be recorded and detectable in the palynological record. To investigate this proposition, this paper presents pollen-analytical data from a peat profile located within the vicinity (30 m) of the archaeological site of Nunalleq. Primarily, the paper aims to examine the following: Do sub-Arctic hunter-gatherers leave a clear palynological footprint in the palaeoenvironmental record? If so, what are the nature of such impacts in northern environments and can they be used as a means to date such activity?
Regional setting
Physical geography
The study site is located within the Y-K delta of southwestern Alaska approximately 5 km south of the village of Quinhagak (Figure 1). The topography of the region is characterised by flat, low-lying treeless tundra dissected by rivers and dotted with small- to medium-sized lakes and ponds. The geology is uncomplicated and comprises recent alluvial deposits of silts, sands and clays of Quaternary age (Wilson et al., 2015). Climatically, the site is located in the sub-arctic and the west coast climate division of Bieniek et al. (2012) within a region of sporadic permafrost (Jorgenson et al., 2008). The nearest observational climate data come from the weather station at Quinhagak airport (c. 5 km northeast at co-ordinates; 59°45′18″N 161°50′43″W) where respective mean summer and winter temperatures for the period 1961–1990 were 12.4 (July) and –11.7 (February) with annual precipitation of 548 mm (SNAP, 2016).
Location of the Nunalleq within Alaska.
Archaeology
The archaeological site of Nunalleq was discovered in 2009 and has subsequently been the subject of five seasons of excavation which have revealed the partial remains of an extensive multi-phase Yup’ik village truncated by erosion from the Bering Sea. To date, tens of thousands of exceptionally well-preserved wood and grass artefacts have been recovered, along with a variety of unique ecofacts such as human hair, insects and coprolites (Forbes et al., 2015). The recovered material culture indicates that the site is associated with the Yup’ik culture and bears strong similarities to ethnographic materials collected from the region in the 19th century. Zooarchaeological and isotope analyses of human hair indicate that subsistence was centred on fishing and hunting of marine mammals and caribou (Britton et al., 2013, 2016). Canid bone is frequent at the site, pointing to substantial numbers of dogs that were likely used for hunting and transportation, but occasionally as a source of food/pelts (McManus-Fry et al., 2016). The majority of rangefinder radiocarbon data span the 14th–19th centuries and suggest occupation of the site from between 1300 and 1405 CE to between 1650 and 1750 CE (Britton et al., 2016). However, the assays presenting the earliest ages were undertaken of materials which may produce unreliable ages, and a 15th–18th century occupation period may be more plausible (Ledger et al., 2016). The presence of extensive burned wood and charred sod deposits in the terminal layers of the site suggests that the village was destroyed by fire. This has been suggested to be associated with a period of warfare – known locally as the Bow and Arrow wars – that afflicted the region from the mid-17th century until contact with Russian fur traders in the early 19th century (Fienup-Riordan and Rearden, 2016).
Sampling location
In order to identify a suitable sampling location, a gouge auger survey was undertaken within the vicinity (<100 m) of the archaeological remains. This identified peat deposits to a depth of 25–35 cm with permafrost present between 30 and 34 cm. A sampling location approximately 30 m east of the archaeological site (Figure 2) was chosen on the basis of the thickness of the peat deposits (>30 cm), depth to permafrost (34 cm) and level of topography. The immediate environment surrounding the sampling location and the archaeological site of Nunalleq is characterised by the dwarf shrub–lichen–sphagnum permafrost plateau and undulating dwarf shrub tussock tundra vegetation communities of Boggs et al. (2014). The shrub flora is dominated by Rubus chamaemorus, Betula nana, Empetrum nigrum and Ledum palustre, while monocots such as Eriophorum angustifolium, Luzula spp., Hierochloe spp. and Festuca spp. are common within the herbaceous component.

The stratigraphy of the sampled profile illustrating its proximity to the archaeological remains at Nunalleq.
Materials and methods
Fieldwork and sediment sampling
Fieldwork was undertaken in July 2015. In the first instance, a small (80 cm × 70 cm) trench was excavated until permafrost was encountered at 34 cm. Thawing of the permafrost was then monitored and recorded to be occurring at approximately 3 cm/day. The trench was subsequently expanded to 350 cm × 150 cm (Figure 2) and maintained daily to promote thawing of the active layer. Sampling was undertaken on 16 July at which point the active layer had thawed to a depth of approximately 60 cm. Before sampling, a horizontal datum was established following a level section of the western face of the excavation trench to ensure accurate measurement of sampling depths. A series of six 10 cm × 75 cm × 15 cm monolith tins were then inserted into the western face of the trench to sample between the depths of 0 and 50 cm as illustrated in Figure 2. The tins were extruded from the trench and wrapped in polyene before being transferred to Quinhagak for sampling. Sub-sampling was undertaken in two phases. Initially, a series of samples for pollen, macrofossil and radiocarbon analysis were sub-sampled at 1 cm resolution from the adjacent sides of each monolith tin. Sampling at 2 cm resolution was then undertaken to acquire approximately 2 L samples for coleopteran (beetle) analysis (subject of a forthcoming paper). Following sub-sampling, samples were stored at 4°C before being transferred to the palaeoecology laboratory at the University of Aberdeen. Loss-on-ignition (LOI) was measured in the laboratory on approximately 1 cm3 subsamples following 3 h of combustion at 550°C.
Pollen, NPP and charcoal analysis
Contiguous 1 cm subsamples of sediment were treated for pollen analysis following standard procedures using NaOH, sieving, acetolysis and floatation stages (Moore et al., 1991; Nakagawa et al., 1998). Lycopodium tablets (Stockmarr, 1971) were added prior to treatment to enable the calculation of concentration and pollen accumulation rate (PAR) data. Residues were suspended in silicone oil before being mounted on slides and examined under a light microscope and counted until the total land pollen (TLP) sum of 500 had been achieved, except in instance where the pollen concentration was very low. Pollen and spores were identified with the aid of Moore et al. (1991) and reference material held at the University of Aberdeen. Nomenclature follows Bennett et al. (1994) and Bennett (2016), with taxa absent from these lists following Moore et al. (1991). Microscopic charcoal in pollen residues was enumerated during pollen counting. The x and y axes of each observed fragment were measured using a microscope eyepiece graticule at a magnification of 400× to establish its approximate surface area. These data were then used to calculate charcoal concentration and accumulation values. Coprophilous fungal spores, derived from fungi that grow on animal dung, were identified with reference to Van Geel et al. (2003). Pollen percentage and influx diagrams were constructed using Tilia and TGView software (Grimm, 1993, 2013) with percentages based upon the TLP sum. The pollen and fungal spore content of subsamples of three presumed dog coprolites recovered from the terminal layers of the archaeological site was also examined. This analysis was primarily undertaken to identify whether coprophilous fungal spores such as Sporormiella-type were present. Consequently, only low counts of up to 150 TLP were undertaken on these samples.
Numerical analysis
CONISS (Grimm, 1987) was used to assist with biostratigraphic zonation of the pollen diagram following square root transformation of the percentage data. Ordination of the pollen samples was performed using CANOCO 4.5. An initial exploration of the dataset was undertaken using detrended correspondence analysis (DCA). This generated a gradient length of 1.912 for the primary axis, indicating a linear rather than unimodal response in the dataset (Lepš and Šmilauer, 2003). Therefore, principal components analysis (PCA) was used to explore the pollen-analytical data (Ter Braak and Šmilauer, 2002). Scaling was focused on inter-species scores, species scores were divided by the standard deviation and species vectors were centred by species.
Chronology
Ten plant macrofossil samples and two peat (humic acid) samples were submitted for radiocarbon dating. Plant macrofossils were isolated by gently disaggregating small peat subsamples in weak NaOH and subsequently washing them through a 125-μm sieve using distilled water. Residues were examined under a binocular microscope and cleaned to remove roots of Ericaceae, monocots and fungal mycelium. The remaining macrofossils – leaves and branches of Sphagnum moss and other bryophytes – were stored in distilled water containing a drop of HCl and sent to the Oxford Radiocarbon Accelerator Unit (ORAU) for AMS 14C dating. Lead dating was used to establish the chronology of the past c. 100 years of accumulation through the analysis of 1-cm-thick contiguous samples from the surface to a depth of 14 cm. The unsupported 210Pb activity within samples was ascertained by subtracting the supported component (measured as 214Pb at 295.22 and 351.93keV) from the total 210Pb activity measured at 46.54keV (Wallbrink et al., 2002). 210Pb and 214Pb activities were measured using EG&G ORTEC hyper-pure Germanium detectors in a well configuration (11 mm diameter, 40 mm depth) housed at Coventry University. Calculation of the depth–age relationship followed methods outlined by Appleby and Oldfield (1978) and Appleby (2001). A CRS dating model was used to calculate ages as accumulation rates varied down-core (Appleby, 2001; Appleby et al., 1988). Radiocarbon dates were calibrated using the IntCal13 calibration curve (Reimer et al., 2013). Age–depth modelling of all the chronological data was undertaken using a Bayesian approach in Bacon (Blaauw and Christen, 2011).
Results
Lithostratigraphy
The profile is divided into a highly minerogenic and a peat-rich phase. From the base of the excavated trench (60 cm) to c. 33 cm, LOI is between 11% and 25% and the deposit comprises brown sandy silt containing infrequent, highly degraded organic remains (Figure 2). At 33 cm, a marked increase in the LOI from 25% to 82% coincides with a relatively sharp lithological change to moderately humified herbaceous peat containing occasional woody remains. The pronounced nature of this stratigraphic transition is suggestive of a depositional hiatus (discussed further below) or truncation of the profile. Peat or sod blocks were the primary construction material for Yup’ik winter dwellings (Oswalt, 1992), and the proximity of the profile to the archaeological remains raises the prospect that the profile may have been truncated through peat cutting. However, there is no visual evidence for this. At 19 cm, there is a transition to red brown/orange brown poorly humified Sphagnum peat, which persists until the surface (Figure 2).
Chronology
Radiocarbon dating results are presented in Table 1. All of the dates fall within the last five centuries and have broad errors, owing to the unfavourable shape of the radiocarbon calibration curve for this period. The data generally overlap at the 2σ level and form a conformable sequence with the Pb210 results. Only two dates present possible age reversals: the first of these occurs at 18–19 cm (OxA-33582, 315 ± 34; 1479–1649 cal. CE) and the second at 28–29 cm (OxA-33586, 129 ± 26; 1677–1941 cal. CE). These dates are potentially incongruous; however, there is no clear basis to suspect they are inaccurate (i.e. the materials assayed are typically reliable dating materials), and therefore, they were retained for age–depth modelling. Despite the lithological evidence (see above), there is no convincing chronological evidence for a depositional hiatus at 33 cm. Although the 14C assays from immediately above and below this depth (OxA-33589, 231 ± 26; OxA-33625, 387 ± 26) are statistically different (T = 18 where χ2 statistic=3.84 (cf. Ward and Wilson, 1978)), there is a wide degree of temporal overlap between dates (OxA-33888, 269 ± 26; OxA-33590, 315 ± 27) that bracket them. The minor age discrepancy between OxA-33589 and OxA-33625 could therefore be attributed to taphonomic differences between dating materials. The lower of these dates (OxA-33625) was undertaken on humic acid, which in northern environments can provide older age estimates than macrofossils from the same depth (see Holmqvist et al., 2016; Oswald et al., 2005; Schofield et al., 2008). Bayesian age–depth modelling in Bacon (Figure 3) was therefore conducted without an inferred hiatus. The results suggest accumulation of the deposit was near linear and occurred at a rate of between 0.05 and 0.12 cm yr–1. The basal sample is estimated to date from 1420 to 1585 CE, indicating that the analysed sequence is contemporary with the occupation of the winter village.
Radiocarbon dates for the Nunalleq profile.

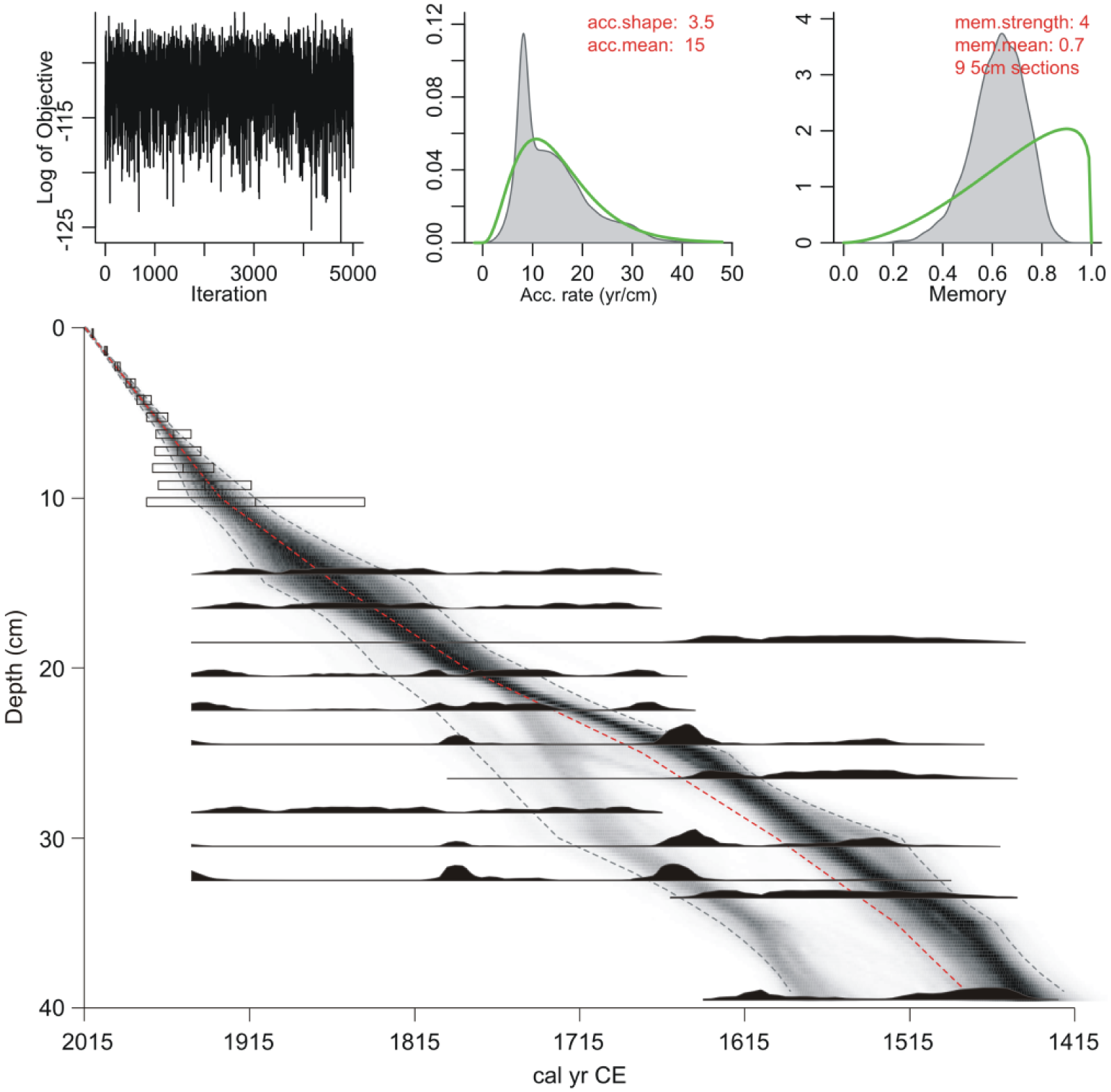
Age–depth model for the Nunalleq profile generated using Bacon v2.2. The mean accumulation was set to 15 yr cm–1 with memory strength and mean, respectively, set at 4 and 0.7. The shaded area in the model indicates all possible age–depth models and the dotted lines show the 2σ age range; darker areas indicate increased certainty and the dotted line indicates the weighted mean of the model.
PCA
PCA (Figure 4a and b) indicates a good separation of data with the first two axes accounting for 37.5% and 20.3% of the respective variance. The analysis identifies three broad clusters of samples (Figure 4b). The broadest of these clusters of samples is characterised by positive responses along Axis 1 in herbaceous pollen types such as Poaceae, Achillea-type, Rumex, Polemonium-type, Epilobium-type and Asteraceae, which are common to meadow-like environments (Hultén, 1968). Axis 1 is also associated with strong positive responses in the fungi Sordaria-type and Sporormiella-type, the latter of which is exclusively coprophilous (Van Geel et al., 2003). The most pronounced negative responses along Axis 1 are associated with Ericaceae and Betula which are typical to tundra environments. The secondary axis is associated with highly negative responses in Sphagnum, Cyperaceae and Artemisia and broadly positive responses in plants that are generally associated with aquatic/humid settings such as Myrica gale, Hippuris vulgaris and Ranunculus-type.
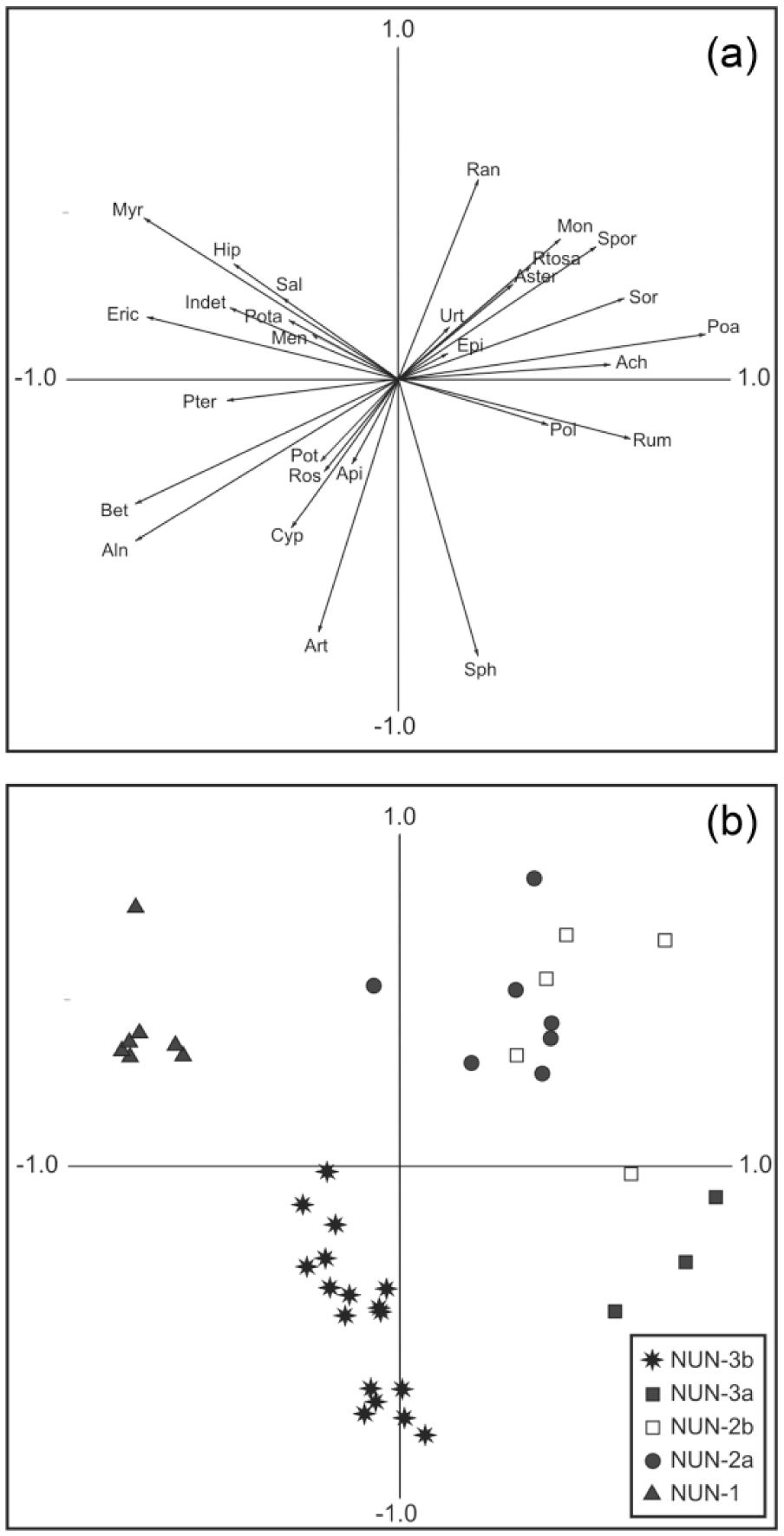
(a) PCA of pollen types and associated proxies (selected taxa only). Key to taxon labels: Ach, Achillea-type; Aln, Alnus; Api, Apiaceae; Art, Artemisia; Aster, Aster-type; Bet, Betula; Cyp, Cyperaceae; Epi, Epilobium-type; Eric, Ericaceae; Hip, Hippuris vulgaris; Indet, unknown and indeterminable; Men, Menyanthes; Mon, Montia fontana; Myr, Myrica gale; Poa, Poaceae; Pol, Polemonium-type; Pota, Potamogeton; Pot, Potentilla-type; Pter, Pteropsida (monolete) indet.; Ran, Ranunculus-type; Ros, Rosaceae; Rtosa, Rumex acetosa-type; Rum, Rumex; Sal, Salix; Sor, HdV-55a Sordaria-type; Sph, Sphagnum; Spor, HdV-113, Sporormiella-type; Urt, Urticaceae. (b) PCA plot of sample scores for the Nunalleq pollen and spore dataset.
Palynology
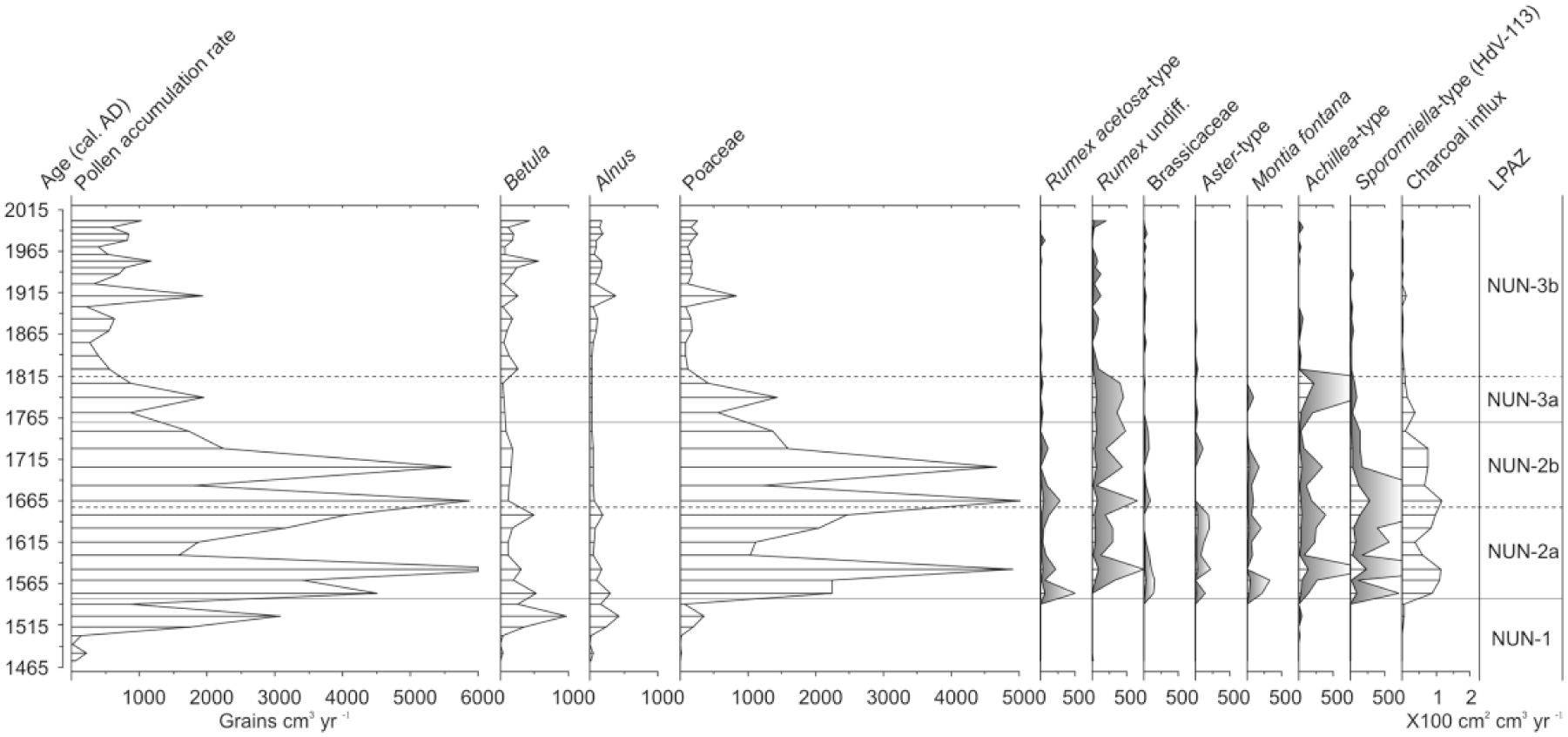
LPAZ NUN-1 dates from approximately 1420–1585 cal. CE. Both microscopic charcoal and pollen concentrations are low, and pollen from shrubs and heaths dominate the zone at 60–80% (Figure 5). Low LOI values (c. 20–25%); the presence of Myrica gale (c. 20%), which favours swamps and shallow waters (Hultén, 1968); and pollen from aquatic plants such as Hippuris vulgaris and Potamogeton indicate a lacustrine depositional environment. Alnus percentages of c. 20% may reflect copses of alder growing alongside humid locations, but – given the absence of alder in the modern environment – more reasonably derive from extra local and regional sources. Betula pollen comprises 15–30% TLP and likely derives from Betula nana growing locally, although the open nature of the landscape would suggest a regional component from Betula papyrifera is also probable. The herbaceous pollen component dominated by Cyperaceae is also indicative of shallow wetland areas. A steady contribution from rarer herbs such as Ranunculus-type and Potentilla-type and Rosaceae may, respectively, reflect wetland plants such as Ranunculus hyperboreus and Comarum palustre.

Percentage pollen and spore diagram for the Nunalleq profile displaying selected taxa (minimum sum = 500 TLP except where noted). Also displayed are the 14C dates, lithology, loss-on-ignition, microscopic charcoal concentration and total pollen concentration. + indicates <1% TLP.
A pronounced lithological and biostratigraphical transition marks the beginning of LPAZ NUN-2a between 1490 and 1660 cal. CE. LOI increases to c. 80% (Figure 5) and there is a shift from highly minerogenic sandy silt to organic-rich, herbaceous peat. Shrub and heath pollen percentages fall at the expense of herbs (Figure 5), although this is mostly an artefact of proportional data related to a fourfold increase in the PAR (Figure 6) driven by Poaceae pollen deposition. The appearance of a suite of herbaceous pollen types, including Rumex, Brassicaceae, Aster-type, Achillea-type and Urticaceae, suggests that the sharp increase in PAR and Poaceae deposition records the emergence of a meadow-like environment (Hultén, 1968). The zone is notable for elevated microscopic charcoal concentrations and the presence of pollen from Montia fontana – a nitrophilous plant common to habitation sites in northern environments (Blondeau and Roy, 2004; Forbes, 1996). Sporormiella-type percentages are also high (1–4% TLP) and are likely indicative of an increase in dung in the landscape.
Pollen accumulation rate (PAR) diagram for Nunalleq showing total PAR and PARs for selected pollen types and Sporormiella-type plotted against the median probable age from the Bacon model. Charcoal accumulation rate is also presented (exaggeration curves where present are 10×).
The opening of LPAZ NUN-2b is marked by a final increase in Poaceae percentages to c. 80% and a minor decrease in the LOI from 93% to 70%, which may be indicative of erosion. Sporormiella-type and microscopic charcoal deposition remains high, and both peak in the zone (Figure 6). Aside from these changes, LPAZ NUN-2b is generally unremarkable from the previous. Shrubs and heath pollen continues to decline; the suite of herbs, indicative of meadow environments, remains well represented; and there are rare occurrences of previously unrepresented types such as Polemonium and Artemisia.
The transition to LPAZ NUN-3a is primarily characterised by a steady decline in the PAR and Poaceae from 80% to 48% TLP, increases in Rumex and Achillea-type percentages, consistent representation from Polemonium and a spike in Sphagnum spores. Concurrent with these developments is the decline in the frequency of other types such as Montia fontana, Aster-type and Brassicaceae and a sharp decline in microscopic charcoal deposition and Sporormiella-type (Figures 5 and 7). Taken together, these changes may be indicative of the beginning of a succession from a meadow-like to a tundra environment.

Pollen and NPP content of presumed dog coprolites (minimum sum = 150 TLP), + indicates <1% TLP.
LPAZ NUN-3b is noteworthy for the continued decline in Poaceae and PAR (Figure 6 and 7) and increased representation from Cyperaceae, Betula and Alnus which is concurrent with a lithological shift towards poorly humified Sphagnum peat. Microscopic charcoal concentrations fall to values previously seen in LPAZ NUN-1, and Sporormiella-type is sporadically represented. The zone is also notable for the increased representation of herbs such as Artemisia, Apiaceae and Potentilla-type.
Dog coprolites
The pollen content of each of the presumed dog coprolites was remarkably similar (Figure 7). Poaceae was present in all the samples at between 30% and 60% TLP, and pollen from shrubs and heaths such as Betula, Ericaceae and Alnus were also frequent. NPPs were also present in each of the coprolites, with the most common types being Chaetomium-type (HdV-7a), Sordaria (HdV-55a), Podospora (HdV-368) and Sporormiella-type (HdV-113). Sporormiella-type is considered to be an obligate coprophilous fungus as it requires passage through an intestinal tract and excretion to complete its lifecycle through germination on dung (Baker et al., 2013). Typically, this occurs through the ingestion of plant materials by obligate herbivores (e.g. caribou in Alaska) rather than carnivores such as dogs. Nevertheless, both wild canids and domesticated dogs consume plant matter (Sueda et al., 2008), and in some South American cultures, plants are regularly incorporated into canid diets to improve health and hunting ability (Bennett and Alarcón, 2015). Therefore, the presence of Sporormiella-type in the dog coprolites, although unexpected, is perhaps unsurprising and implies that, at Nunalleq at least, it may reasonably be considered a proxy for dogs. In addition to the coprophilous fungi, eggs of the parasitic fish tapeworm Diphyllobothrium spp. were also noted to be abundant in two of the samples. This parasite can cause severe and potentially fatal conditions in dogs and have been noted to be common along the northwest coast of North America (Bathurst, 2005). The presence of Diphyllobothrium spp. in the coprolites therefore suggests that dogs at Nunalleq were either fed or had access to raw or undercooked fish.
Discussion
Is a hunter-gatherer footprint evident?
Establishing what constitutes a palynological signal for hunter-gatherers is far from straightforward (e.g. Bos and Urz, 2003; Edwards, 1998; Renouff et al., 2009; Roy et al., 2015). Hunter-gatherer societies are, and were, diverse, interacting with the environment in myriad ways that differ between cultural groups. Indeed, until relatively recently many North American hunter-gathering societies were presumed to have exerted little to no influence upon nature (Denevan, 1992). Despite these challenges, it should be possible to identify palynological evidence that can be ascribed to human activity. Most palynologists would agree that microscopic charcoal concentrations – when consistently elevated above a baseline level for any given location – can be considered as indicative of human impact (e.g. Edwards, 1990; Keeley, 2002). Similarly, the concerted presence of people or animals in a geographic space is accepted to result in trampling and general disturbance that favours the propagation of geographic specific apophytes (cf. Behre, 1981). Incidental nitrogen enrichment also has the potential to favour the growth of nitrophilous plants that may otherwise be absent (e.g. Fredskild, 1988). More recently, it has become established that coprophilous fungi are a powerful additional proxy for identifying human impact (e.g. Baker et al., 2013; Ledger et al., 2014a; Van Geel et al., 2003).
Taking the diagram from Nunalleq, it appears clear that the pollen-analytical changes occurring at the boundary between LPAZs NUN-1 and NUN-2a mark the appearance of evidence for Yup’ik activity. In LPAZ NUN-1, microscopic charcoal concentrations are very low and the pollen assemblages compare favourably with other sites in southwestern Alaska that exhibit no evidence for human activity (e.g. Hu and Brubaker, 1996; Hu et al., 1995). In contrast, LPAZ NUN-2a is characterised by a significant and sustained increase in microscopic charcoal deposition. This almost certainly reflects fires associated with common domestic activities at a Yup’ik dwelling, such as heating, cooking and the curing of meat/fish (Oswalt, 1992). A diversification of the pollen assemblages is also evident. NUN-2a marks the first appearance, or increased presence, of a number of pollen types such as Rumex, Ranunculus-type, Brassicaceae, Aster-type, Achillea-type, Urticaceae and Montia (Figure 5). It is likely that these occurrences reflect indigenous plants spreading from natural associations and behaving as apophytes in response to disturbance such as trampling and sod cutting (cf. Fredskild, 1988; Ledger et al., 2014a). Species within these families/types – such as Rumex graminifolius, Rumex acetosa, Rorippa islandica, Rorippa hispida and Achillea borealis – are often associated with ruderal/disturbed habitats in Alaska (Hultén, 1968; Pratt, 1989) and the circumpolar north in general, while plants within the Asteraceae and Urticaceae families are commonly associated with human disturbance (e.g. Behre, 1981). In addition, the patterns recorded from the beginning of NUN-2a to the end of NUN-2b are strikingly similar to those observed to be associated with the arrival of humans in pollen diagrams from sub-Arctic locations such as Greenland and Iceland (Schofield et al., 2013). Particularly noteworthy is the expansion in Montia fontana. This nitrophilous plant has been recorded in palaeoecological studies from hunter-gatherer sites in Labrador and Nunavik (e.g. Bhiry et al., 2016; Roy et al., 2012, 2015), Norse sites in Greenland (e.g. Ledger et al., 2014a, 2015) and is common in habitations in the Arctic region of Nunavik (Blondeau and Roy, 2004). Concurrent increases in the coprophilous fungi Sporormiella-type (Figures 5 and 7) are also indicative of increased amounts of faeces in the environment. Typically, Sporormiella-type is associated with obligate herbivores (Baker et al., 2013); however, the results from dog coprolites (Figure 6) indicate that the increases through NUN-2a/b likely derive from excrement from the packs of dogs that were integral to Yup’ik hunting practices (McManus-Fry et al., 2016).
Palaeoenvironmental change at Nunalleq
It appears clear that the presence of hunter-gatherers at Nunalleq resulted in a material trace in the palynological record from NUN-2a onwards. Nevertheless, the extent to which subsequent palaeoenvironmental change reflects human activity or natural environmental processes is uncertain. Evidence for indigenous hunter-gatherer groups as active agents in ecosystem development is suggested from a variety of locations across North America (e.g. Keeley, 2002; Oliver, 2007), although there is no ethnographic evidence for such activity in prehistoric Yup’ik populations (e.g. Nelson, 1899). Any human influence on palaeoenvironmental change at Nunalleq is therefore likely to have been incidental.
The most pronounced palaeoenvironmental shift at Nunalleq occurs at the NUN-1/2a LPAZ boundary (Figure 5). The lithology changes from organic sandy silt to herbaceous peat and the LOI shows a sharp increase in the organic content of the deposits. These developments are indicative of the beginning of peat development and appear to record a successional transition from an open water to a meadow-like environment (cf. Treat et al., 2016). Biostratigraphic changes similarly point to the emergence of a monocot-rich meadow; however, they also present the first evidence of human impact. The synchronous nature of these developments is intriguing and may imply a causal link. Sod cutting associated with the construction of the dwelling – and the destruction of the integrity of the palaeoenvironmental record – could account for the observed changes (cf. Schofield et al., 2008). However, the radiocarbon dating is not suggestive of a hiatus (see above), and absence of stratigraphic evidence for a hiatus makes it unlikely the sequence is truncated. More likely, the NUN-1/2a LPAZ zone boundary reflects a real successional transition. The change dates to 1490–1660 cal. CE and is concurrent with ‘Little Ice Age’ in Alaska (Hu et al., 2001; Overpeck et al., 1997) which may have induced permafrost aggradation (cf. Treat et al., 2016). Frost heave associated with climatic cooling may have uplifted the ground surface altering the hydrological regime, leading to the development of a meadow environment (e.g. Kanevskiy et al., 2014). The development of permafrost would then limit water infiltration and enable the development of peat (Treat et al., 2016). Indeed, the ‘Little Ice Age’ has been implicated in permafrost aggradation and the initiation of peat accumulation elsewhere in Alaska and the Arctic (Bhiry et al., 2007; Kultti et al., 2004). Alternatively, the successional change observed may have been triggered by human activity. Drainage systems have previously been noted to have been incorporated into the construction of sod houses in Alaska (Knecht, 1995), and such features were identified at Nunalleq (Knecht, personal communication). It is therefore plausible that drainage systems, incorporated into the structures at Nunalleq, led to a drying of the environment that results in the rapid establishment of a monocot-rich meadow community.
Regardless of the causes for the shift in the depositional environment and initiation of peat growth, it is likely that the development and propagation of a meadow-like environment were favoured by human activity. The consistent presence of Montia fontana and Sporormiella-type percentages through LPAZs NUN-2a and NUN-2b are indicative of nitrogen enrichment that may account for a fourfold increase in the PAR. This increase in deposition and vegetation productivity is plausibly related to the proximity of the profile to the dwelling (c. 30 m). There was almost certainly an element of casual sheet middening within the immediate vicinity of the village associated with daily activities such as food processing and woodworking. Similarly, the area would have been subject to consistent trampling and disturbance that would favour the appearance and establishment of apophytes such as Rumex, Brassicaceae and Achillea-type.
Determining exactly when the human influences cease is more complicated. The response of vegetation in sub-Arctic regions to changing environmental pressures often occurs very slowly, if at all (e.g. Forbes, 1996). Vegetation changes alone are seldom a reliable indicator of the cessation of human activity in northern environments (Edwards et al., 2011; Ledger et al., 2014b; Roy et al., 2015). Nevertheless, declining Sporormiella-type and microscopic charcoal deposition at the transition between LPAZs NUN-2b and NUN-3a appears to reflect the cessation of human activity at Nunalleq and are concurrent with falling productivity (Figure 6) and the disappearance of Montia fontana (Figure 5). Despite this, there is continued representation of taxa such as Rumex and a significant expansion in Achillea-type through LPAZ NUN-3a. Both these taxa, and others such as Brassicaceae, Aster-type and Urticaceae, continued to be well represented in LPAZ NUN-3b (Figure 5). These observations chime with those from other abandoned archaeological sites in the circumpolar north (e.g. Forbes, 1996; Fredskild, 1961; Porsild, 1932), suggesting that disturbance associated with human settlement had an enduring influence on the vegetation at Nunalleq.
Are palynological records a reliable means to date hunter-gatherer activity?
Identifying the establishment of the Yup’ik village at Nunalleq in the palaeoenvironmental record is relatively straightforward. On the basis of microscopic charcoal, coprophilous fungal spores and the appearance of apophytes such as Montia fontana, it is clear that the NUN-1/2a boundary marks the beginning of palynological evidence for hunter-gather activity. The modelled age for this zone boundary is 1490–1660 cal. CE, post-dating the oldest radiocarbon data from the archaeological site (see Britton et al., 2016). Given the coincidence of the palynological evidence for human activity with pronounced lithological changes, this could suggest that the profile is truncated (cf. Schofield et al., 2008). However, the earliest assays from the archaeology are on wood and grass artefacts which have been demonstrated to provide unreliable results on archaeological sites in the circumpolar north (Ledger et al., 2016). If these data are omitted, the oldest 14C date from the archaeology provides a calibrated age of 1430–1620 CE (Britton et al., 2016; Ledger et al., 2016). The remarkable consistency of this date with that of the modelled age from the palaeoenvironmental record (1490–1660 cal. CE) suggests it is unlikely that the profile has been truncated through sod cutting.
Identifying and dating the point at which evidence for human activity ceases in the palaeoenvironmental record from Nunalleq are complicated by the asynchronous nature of changes in key indicators of human activity. The disappearance of Montia fontana and Sporormiella-type poses solid evidence that the NUN-2b/3a (Figure 5) reflects abandonment, while deposition of microscopic charcoal – although much reduced – into NUN-3a (Figure 6) may indicate continuity of activity. Nonetheless, it is clear that by the NUN-3a/b boundary, pollen-analytical indicators return to pre-activity baselines (Figures 5 and 6) and that permanent occupation has ceased. Modelled age ranges for the end of NUN-2b (1725–1815 cal. CE) and NUN-3a (1780–1850 cal. CE) should therefore be considered the terminus post quem and terminus ante quem for the end of activity at Nunalleq. Both these dates provide slightly more recent age estimates than the archaeology, which suggests abandonment of the village somewhere between 1650–1750 cal. CE. This apparent time lag, between the abandonment and an obvious palaeoenvironmental signal, is unsurprising given that Arctic ecosystems do not readily recover from disturbance (e.g. Forbes, 1992, 1996). The persistent microscopic charcoal signature in NUN-3b is less easily explained. However, the weak nature of this signal – an order of magnitude lower than in the previous zone – does not appear indicative of regular local fires. Microscopic charcoal deposition in NUN-3b may therefore be related to more distant or temporary occupation in the vicinity of the site.
Conclusion
Evidence for hunter-gatherers in the palynological record is far from widespread. Despite many years of research in Europe, and more recent attempts in North America, few standalone sequences present unequivocal evidence for such activity. This owes much to the ephemeral nature of any impacts and the taphonomic issues relating to the responsiveness of palynological evidence to them. Consequently, this paper adopted the perspective that maximising the palynological signal for such activity was essential to definitively identifying hunter-gatherers in the palaeoenvironmental record. An inherent risk of this approach is that samples derived from very close to human activity areas may be adversely influenced by the impacts one wishes to examine. This study is no exception. Despite possible evidence for the truncation of the profile – which on balance is discounted – this paper outlines convincing evidence for a pollen-analytical signal associated with pre-historic Yup’ik activity at Nunalleq. Aside from the well-established microscopic charcoal signal, there are indications of the emergence of meadow-like communities in the areas immediately surrounding the site. Once established, the meadow community may then have been sustained by the activities of humans and animals, such as fish processing, trampling and defaecation. Chronological evidence which dates these changes as being contemporary with radiocarbon data derived from the archaeological site serves to reinforce these conclusions. Indeed, the findings presented here highlight the prospect of using palynological sequences to independently date palaeo- and neo-eskimo sites across the circumpolar north. This approach offers the opportunity to circumvent taphonomic issues associated with such sites (e.g. Ledger et al., 2016) and a novel means to improve our understanding of the chronology of the recent past of circumpolar North America.
Footnotes
Acknowledgements
The author thanks Professor Ian Foster for providing Pb210 dates and Véronique Forbes, John Randall and Roman Meunier for their assistance in the field on a stormy Saturday afternoon. Special thanks are also due to Qanirtuuq Incorporated and Warren Jones for logistical support and warm hospitality in Quinhagak. Finally, the author would like to acknowledge the helpful and insightful comments of two anonymous reviewers.
Funding
This research was funded by the Arts and Humanities Research Council (AH/K006029/1) awarded to Drs Rick Knecht, Kate Britton and Charlotta Hillerdal and an NERC Radiocarbon Facility grant (NRCF 1965.1015).