
Abstract
The Songnen grasslands were traditionally thought to be dominated by Artemisia and Chenopodiaceae plants as early as the late Pleistocene. However, increasing evidence has called that interpretation into question. To shed new light on the paleovegetation evolution of the arid and semi-arid steppe in North China, phytolith assemblages preserved in the region’s sand-paleosol sequence (section Daike) are used as a proxy for paleovegetation structure. Results show that both the sand and paleosol layers in the Songnen grassland sections contain well-preserved phytoliths attributed to different families of grass. This is the first direct evidence of the nature of the vegetation that existed during the sandy layer episodes. Moreover, the phytolith evidence represented in the samples indicates that plant successions happened within the subfamilies of Poaceae through the time. Referring to phytoliths in modern plants and topsoils, and using statistical analyses, we propose that phytolith assemblages in the section Daike originated from Poaceae-dominant communities rather than an Artemisia-Chenopodiaceae ecosystem. The phytoliths, and evidence from the historical and modern pollen-vegetation relationships, lead to rejection of the hypothesis of a past widespread Artemisia-Chenopodiaceae ecosystem in the Songnen grasslands. Using published radiocarbon and thermoluminescence data, it is proposed that the present Poaceae-dominated grasslands developed as early as the early Holocene. This study also highlights the usefulness of phytolith analysis in paleovegetation reconstruction in arid and semi-arid lands.
Introduction and background
Grasslands occupy 32% of the earth’s land surface and are one of the most widespread vegetation types (Adams et al., 1990). Mainly distributed in America, Europe, and Asia, temperate grasslands are sensitive to global climate change and play an important role in global carbon cycling (Anderson, 1991; French, 1979; O’Mara, 2012; Scurlock and Hall, 1998). China has a large area of temperate steppe dominated by Stipa baicalensis (Poaceae) and Leymus chinensis (Poaceae) (Wu, 1980; Zhu, 1963). Covering about 40% of its territory, the large area of temperate steppe in North China is an important component of global grasslands and contributes to global biogeochemical cycling and associated climate effects (Ni, 2004; Wang et al., 2007).
However, many grasslands have experienced extensive land degradation associated with climate change and recent negative human interference (Gang et al., 2014). To prevent and reverse this degradation, a series of ecological protection and restoration projects have been implemented in many countries (Cao, 2011; Doren et al., 2009) including China (Cai et al., 2015). Understanding the composition of natural vegetation in geological and historical periods is crucial for assessing the impacts of climate change on grasslands and providing geological references for natural vegetation restoration in semi-humid and semi-arid regions (Liu et al., 1996; Lu et al., 2003).
Pollen analysis provides the most commonly used proxy for paleovegetation reconstruction in grasslands. For example, in the past few decades, many scholars have proposed that the Songnen grasslands were dominated by Artemisia and Chenopodiaceae plants as early as the late Pleistocene (Li, 1991a; Li and Lu, 1996; Qiu et al., 1992; Wang and Xia, 1988). The strongest evidence for that composition is the predominance of Artemisia and Chenopodiaceae pollen in the region’s paleosol sediments. However, a vegetation composition dominated by Artemisia and Chenopodiaceae is quite different from the vegetation pattern on the modern North China steppe, which is dominated by Poaceae plants (Zhu, 1963), such as Leymus chinensis, Stipa baicalensis, Filifolium sibiricum, Arundinella hirta, and others. A major difference exists between the vegetation of the present landscape and that inferred from pollen analyses for the pre-modern Holocene and late Pleistocene. This leads to some important questions. When and how did the present Poaceae-dominated meadow-steppe replace a grassland dominated by Artemisia and Chenopodiaceae? Did vegetation cover exist during the interpaleosol sand layer episodes? Did the Artemisia-Chenopodiaceae grassland actually exist, or has its fossil evidence been preferentially preserved relative to other types of vegetation?
Direct tests of the traditional hypothesis of pre-modern vegetation dominated by Artemisia-Chenopodiaceae are impeded by the scarcity of well-preserved paleoecological indicators (e.g. pollen, plant macrofossils, or charcoal). The sand-sized sediment that dominates many arid and semi-arid regions, including the North China grasslands, is abrasive and strongly associated with aerobic conditions. These qualities inhibit the preservation of pollen and plant macrofossils (Alexandre et al., 1997; Barboni et al., 1999; Bremond et al., 2008; Horowitz, 1992). The lack of precision with which pollen of species or subfamilies of Poaceae can be identified also limits reconstruction of the vegetation history in grasslands (Alexandre et al., 1997; Barboni et al., 1999; Bremond et al., 2008). To investigate the paleovegetation record more fully, we make use of an alternative and complementary proxy: phytoliths.
Phytoliths are microscopic opal-A particles that precipitate in cells and/or between cells of living plant tissues. Phytoliths occur in many plant families (Piperno, 1988), but are especially abundant, diverse, and distinctive in the Poaceae (Kondo et al., 1994; Twiss, 1992; Twiss et al., 1969; Wang and Lu, 1993). Phytoliths hold great promise for reconstructing the paleoecology of grasslands because they provide information not available through conventional pollen analysis. For example, unlike grass pollen, which is identifiable only to the family level (Poaceae/Gramineae), grass phytoliths are diagnostic to the subfamily level (Barboni et al., 1999; Twiss, 1992; Twiss et al., 1969). Moreover, phytoliths can remain well preserved in oxidizing conditions (Piperno, 1988; Wang and Lu, 1993). Because of these attributes, phytolith analyses have enabled paleoenvironmental researchers to discriminate between forest and grassland, and especially to distinguish the vegetation composition of grasslands (e.g. Alexandre et al., 1997; Barboni et al., 1999; Boyd, 2005; Bremond et al., 2008; Strömberg, 2004). Therefore, phytolith analysis has great potential for specifying the paleovegetation patterns and dynamics in grasslands.
This paper reports on a pilot investigation that tested for the existence of phytoliths, and the information about paleovegetation dynamics that could be discerned from phytoliths, in Holocene sand-paleosol sediments of the Songnen grasslands of North China. Results allow a more refined reconstruction of paleovegetation composition and distribution patterns in the Songnen grasslands as well as their evolution through time. This is also the first phytolith-based reconstruction applied to these sand-paleosol units in North China. Results of this study demonstrate the utility of phytoliths for paleovegetation reconstruction in drylands, and provide the foundation for a more detailed study of the Late Quaternary vegetation dynamics in the study area as well as other arid and semi-arid regions. By helping to improve accuracy in paleovegetation reconstructions, phytolith analyses will lead to a better understanding of vegetation change dynamics in response to past and future climate change in grassland ecosystems.
Study area and methods
The Songnen grasslands (44°45′–48°20′N, 120°40′–126°00′E) are located in the northwestern portion of the Northeast China Plain at an elevation of approximately 130–160 m (Xiao, 1995). Situated along the eastern margin of the temperate steppe in North China, its regional average annual precipitation ranges from 360 to 480 mm, and the average annual temperature ranges from 3.5°C to 5.0°C. Influenced by the East Asian monsoon, 70% of the region’s precipitation falls in summer (Li, 1994). The widespread sand found on the Songnen Plain was transported to the region by the wind from nearby Late Quaternary alluvium and lacustrine sources. Because of the wide distribution of Holocene sands across the Songnen grasslands, the region is also known as the Songnen sandy lands (Jie and Lu, 1995; Xiao, 1995).
Seven sand and paleosol samples from five different locations and a complete sand-paleosol sequence (section Daike, 2.25 m in depth, abbreviated as DK) (Table S1, available online) in the Songnen sandy lands were collected and analyzed (Figure 1). To facilitate the interpretation of phytolith assemblages, modern plants and 28 topsoil samples (collected from 18 sites) under representative plant communities were collected from the study area. Information on the plant communities and topsoil samples are listed in Table S2, available online.
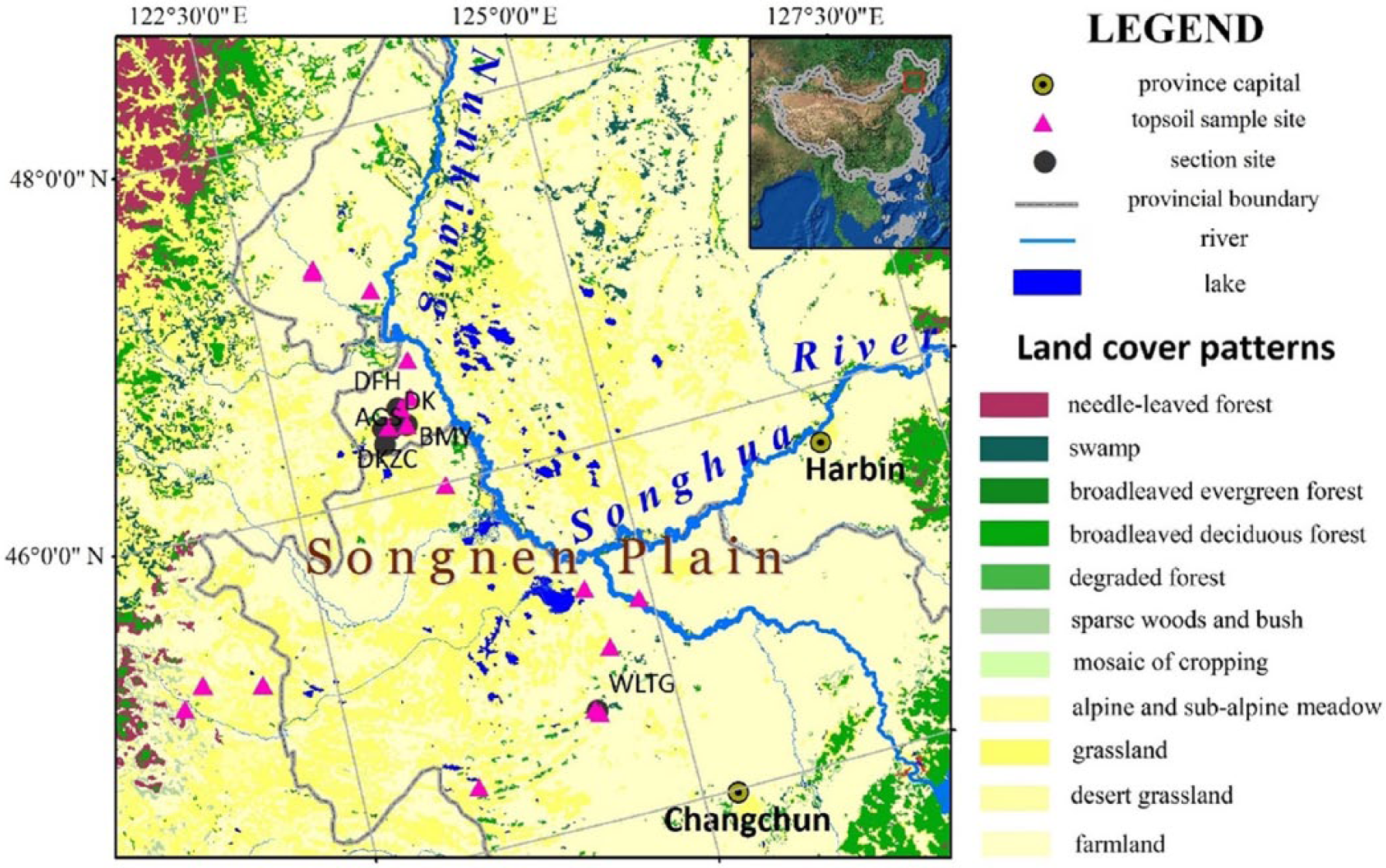
Land cover map showing the location of section and topsoil sample sites in the Songnen grasslands (Wu et al., 2003).
Phytoliths in the sand-paleosol section were extracted using a modified version of the Piperno (1988) method (Wang and Lu, 1993). According to the particular composition of sandy sediments, a subsample of ~20 g was taken from each sample (~1g for plants and ~5g for topsoil samples, see Li et al., 2017 for more details). Each subsample was sequentially processed as follows: (1) treatment with 10% HCl for 4 h to dissolve carbonates; (2) removal of organic matter with 10% H2O2 (the plants and topsoil samples were treated with 65% HNO3) at 70°C until the reaction ceased; (3) flotation of phytoliths using a ZnBr2 heavy liquid at a density of 2.35 g/cm3, and rinsing of the residua several times; (4) addition of a tablet of Lycopodium spores (27,560 spores per tablet) to determine phytolith concentration; and (5) after cleaning, dipping each sample in neutral balsam to facilitate preparing slides. For each sample, approximately 300 grains were identified and counted microscopically under an Olympus BX53 optical microscope at a magnification of 400×. Phytoliths were identified mainly following the classification system used by Lu et al. (2006), but with reference to the classification systems of Twiss et al. (1969). Phytoliths were named according to the International Code for Phytolith Nomenclature 1.0 (Madella et al., 2005). Given the total number of Lycopodium spores, total phytolith grains were determined according to the ratio of identified spores to phytoliths under the microscope. The phytolith concentration was calculated as the ratio of phytolith grains to the mass of each subsample (grains per gram). The stratigraphic diagram was constructed using Tilia (Grimm, 1992). An independent-sample t-test and discriminant analysis were performed using SPSS 19.0 software (Statistical Package for Social Sciences, Chicago, IL, USA).
Results and discussion
Phytolith assemblages of section DK and paleovegetation composition
All of the samples yielded abundant phytolith types attributed to different families of grass and occasionally trees (Figure 2), and indicated the presence of grasses in bloom. In addition, compared with phytoliths observed in other sediments, for example, peat (Lun et al., 2016) and loess (Lu et al., 2007), those extracted from the sandy sediments were typically larger and displayed evidence of having undergone some abrasion and corrosion after being released from the plants (Figure 2).

Microphotographs of selected phytolith types counted in this study. (a–e) rondel, (f–k) bilobate, (l–q) saddle, (r–u) trapeziform polylobate, (v) polylobate, (w) irregular type, (x–ab) acicular, (ac) hair cell, (ae) silicified vessel, (af–ag) blocky, (ai) sponge spicule, (aj–ak) bulliform, (al, an) bulliform-Phragmites, (ad, ah, am, ap) elongate, (ao) elongate tabular, and (aq) blocky from woody plants.
In section DK, the paleosol samples had a mean phytolith production of 4234 grains per gram, whereas the sand samples yielded an average of 2056 phytolith grains per gram of sediment (Figure 3), indicating a relative better vegetation condition during paleosol episodes than during sand layer episodes. The sand and paleosol phytolith assemblage was characterized by a high content (>50%) of elongate and acicular types (Figure 3), which are diagnostics of Poaceae (Kondo et al., 1994; Piperno, 1988; Twiss et al., 1969; Wang and Lu, 1993). Trapeziform polylobate and rondel forms, typical of C3 Pooideae (Kondo et al., 1994; Twiss et al., 1969), represented about 7% of the observed phytoliths and were more abundant in the sand samples. Bilobate forms, mainly from the Panicoideae (Kondo et al., 1994; Twiss et al., 1969), accounted for 5% on average among all the samples, whereas the saddle type, typical of Chloridoideae, represented no more than 2% on average.

Relative abundance of phytoliths (main categories) observed in section DK and calculated phytolith indices in the Songnen grasslands, northeastern China. Abundances lower than 1% are identified by a dot.
Employing the Ic and Iph indices, the present study points to paleovegetation composition changes through time within section DK. The climatic index, Ic, is the ratio of the percentage of phytoliths mainly produced by Pooideae to the combined percentage of phytoliths mainly produced by Pooideae, Chloridoideae, and Panicoideae grasses. First defined by Twiss (1992), Ic has been widely used to estimate the relative proportion of C3 to C4 grasses present in paleovegetation. In this study, Ic values show an increasing trend from the basal to the top of section DK (Figure 3), indicating a gradual expansion of C3 plants since the mid-Holocene. Previous work has demonstrated that C3 plants have a wider distribution than C4 plants in meadow-steppe and steppe areas of the Songnen grasslands, whereas the C4 species have a higher tolerance for drought (Yin and Wang, 1997). This suggests that the C3 plants increased during cooler and wetter conditions but decreased during drier and warmer episodes (Li et al., 2017). Various records show that the study region has experienced a significant climate shift since the mid-Holocene; regional temperature has declined in response to the solar radiation-driven changes (i.e. Stebich et al., 2015; Wu et al., 2011). As a result, C3 Pooideae plants, which are more tolerant of cooler temperatures than C4 plants, expanded in the Songnen grasslands.
Iph is the percentage of Chloridoideae short-cell phytolith types relative to the sum of Panicoideae and Chloridoideae short-cell phytolith types. As first defined by Diester-Haass et al. (1973), Iph was used to track the humidity-aridity tendencies in marine cores. Section DK shows significant differences in Iph values for the sand compared with the paleosol samples (Figure 3). Mean Iph value was 23% for the sand and 16% for paleosol samples, respectively, suggesting that the sand layers formed during drier conditions than existed during formation of the paleosols. Based on the variations of Iph through the section, we can infer the relative proportion of Chloridoideae to Panicoideae through time. During the sandy layer intervals, xerophytic C4 grasses (Chloridoideae) were distributed more widely than mesophytic C4 grasses in the Songnen sandy lands.
Phytolith assemblages support the notion that the Songnen grasslands were mainly covered by grasses at least since the formation of the sandy lands (Li, 1991a, 1991b; Qiu et al., 1992). The different composition of phytolith assemblages between sand and paleosol layers suggests that plant successions have taken place through time. However, the phytolith evidence indicates that most plant successions happened within the subfamilies of Poaceae, and that the Poaceae-dominated grasslands have never been dominated by Artemisia-Chenopodiaceae.
Evidence for Poaceae-dominant grasslands
Evidence from modern plants and phytolith concentration in the topsoil
Phytoliths extracted in modern plants show that the main species of Poaceae, Artemisia, and Chenopodiaceae in the Songnen grasslands produce distinct phytolith morphotypes (Figure S1, available online). Phytolith assemblages extracted from section DK suggest that most originated from Poaceae. The epidermal polygonal morphotype, which is the diagnostic phytolith of Artemisia and Chenopodiaceae, is almost absent in section DK, suggesting a relative abundance of Poaceae. However, previous studies have found epidermal polygonal phytoliths, produced by Asteraceae in contrast with Poaceae types, to be of low percentages or even absent in soil assemblages even when they dominate the surface vegetation (Cabanes et al., 2011; Cabanes and Shahack-Gross, 2015; Honaine et al., 2009). This contrast in representativeness between the Poaceae and the Artemisia and Chenopodiaceae phytoliths has been attributed to differences in the silicification process, dissolution, or physical fragmentation (Honaine et al., 2009; Madella and Lancelotti, 2012). The lower degree of silicification of Artemisia and Chenopodiaceae phytoliths would have affected their preservation, preventing their presence in the profile. Thus, simply judging from the phytolith morphotypes preserved in the soil, grass communities could be overestimated while the Artemisia and Chenopodiaceae communities could be overlooked.
Instead of using preserved phytolith morphotypes alone, a clearer picture of the paleovegetation signal can be obtained when combining that information with data on phytolith concentration. Phytolith concentration is positively correlated with the above-ground biomass, and can be used to investigate the paleovegetation density (Blinnikov et al., 2013). Thus, the more Poaceae plants there are above the ground, the higher the Poaceae-phytolith concentration will be in the soil. In the Songnen sandy lands, the phytolith concentration of topsoil samples collected under the Poaceae-dominant quadrats ranged from 921 to 16,907 grains/g (Figure 4a). This provided the modern concentration reference for determining a Poaceae-dominated community. The phytolith concentration results from section DK are plotted on the right panel of Figure 4a, and indicate that all but six samples originated from Poaceae-dominant communities. Furthermore, an independent-sample t-test comparing the topsoils to the profile samples yielded a p-value of 0.626, signifying no significant difference between these two groups. Therefore, it is more likely that the Songnen grasslands were dominated by Poaceae than by Artemisia or Chenopodiaceae.
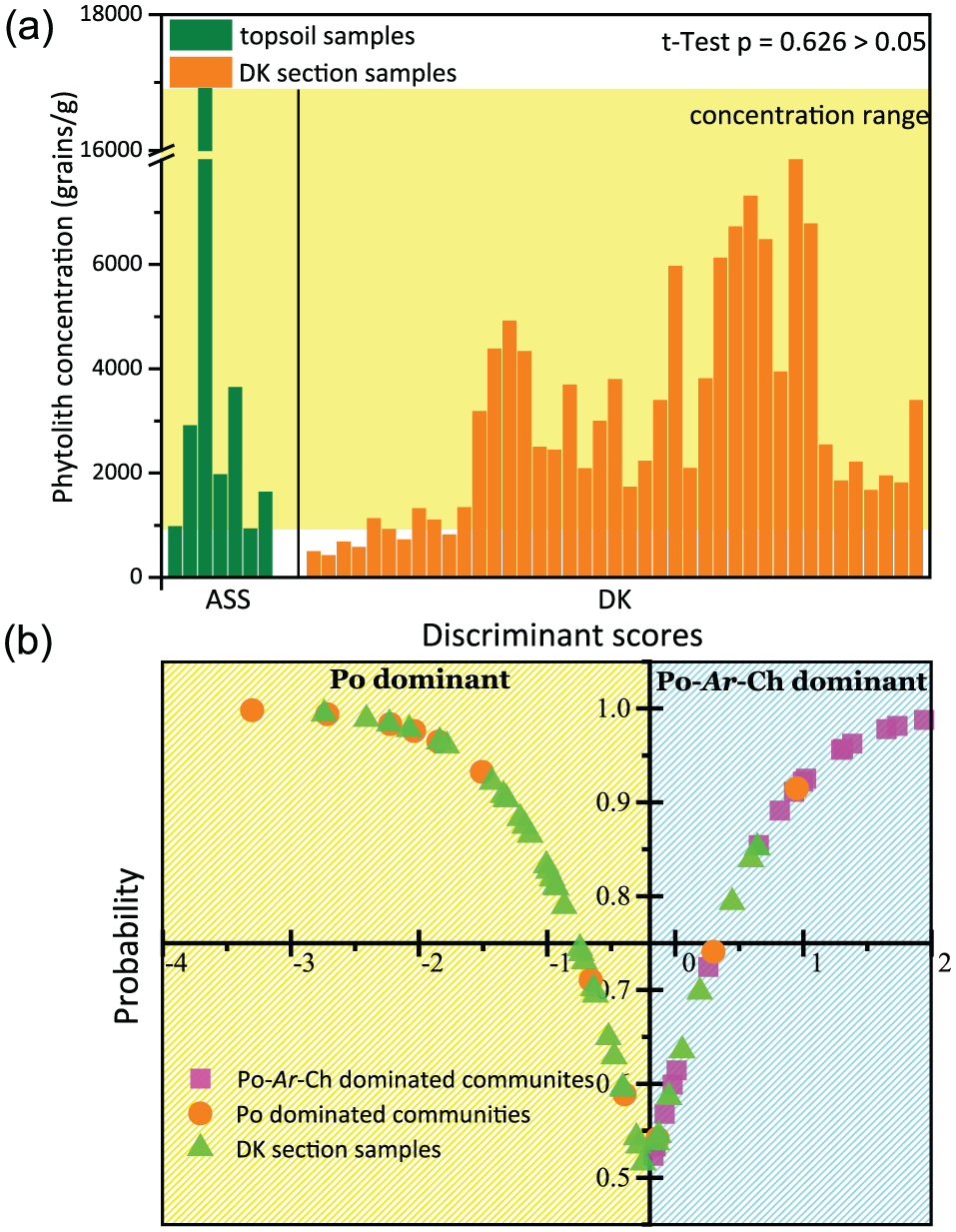
Comparisons of phytolith assemblages between the topsoil and section samples: (a) phytolith concentration and (b) discriminant analysis results.
Evidence from topsoil phytolith assemblages
Field-based study in the Songnen sandy lands indicates that Artemisia- and Chenopodiaceae-dominant communities were not as common as previous paleovegetation reconstructions proposed. Instead, they were often species found in association with a Poaceae-dominant community. To explore this further, discriminant analysis was used to establish a more precise paleovegetation composition pattern for the study region. First, we classified 28 topsoil phytolith assemblages into two major groups (Table S2, available online), Poaceae-dominant (abbreviated as Po dominant in Figure 4b) and Poaceae-Artemisia-Chenopodiaceae-dominant communities (abbreviated as Po-Ar-Ch dominant in Figure 4b). Then, a linear discriminant function was constructed based on the differences in phytolith assemblages of these two groups. The linear discriminant function successfully classified 89.3% of the samples into their original community groups. A summary of the discriminant analysis results for modern phytolith assemblages from the study region is elaborated in Table S3, available online, and Figure 4b. Using the discriminant function, most of the samples (78%) in section DK were classified as Poaceae-dominant communities. The probability that each sample was dominated by Poaceae or Poaceae-Artemisia-Chenopodiaceae is plotted in Figure 3. From this analysis, we conclude that most of the phytolith assemblages in section DK originated from Poaceae-dominant communities.
Evidence from historical documents and modern pollen-vegetation relationship studies
North China grasslands were the birthplace of ancient nomadic civilization in the Bronze Age (Chen, 2015; Qiao, 1992; Wu, 2002). The dynasties established by those nomads reigned in North China for thousands of years and even succeeded in unifying China (i.e. Yuan and Qing dynasty) (Compiling group of the History of National Relationships in Northern China (CGHNRNC), 1987; Grousset, 1970; Toynbee, 1972). Nomadic people rely primarily on livestock for their food and material needs (Bai and Kung, 2011; Bendrey, 2011), therefore abundant palatable grass was a critical requirement for the success of the region’s ancient nomadic dynasties (CGHNRNC, 1987; Grousset, 1970; Toynbee, 1972). Compared with Poaceae and Leguminosae, Artemisia and Chenopodiaceae provide few favored forage plants (The Chinese Herbage Cultivar Registration Board (CHCRB), 2008). It is unlikely that a grassland with widely distributed Artemisia and Chenopodiaceae would have been able to support the highly successful ancient nomadic economy and livestock-farming agriculture in North China. These factors call into question the notion of a grassland at that time dominated by Artemisia and Chenopodiaceae.
Recent studies of pollen-vegetation relationships conducted in steppe and desert-steppe environments (Li et al., 2011; Xu et al., 2005, 2007, 2014) also challenge the Artemisia and Chenopodiaceae hypothesis. Pollen-vegetation relationships have shown that even when Artemisia and Chenopodiaceae plants are rare or absent in vegetation communities, they still dominate the pollen assemblages, indicating strong over-representation of those plants in the pollen signal (Li et al., 2011; Xu et al., 2005, 2007, 2014). Moreover, recorded Poaceae pollen in modern Poaceae-dominated communities is usually less than 10%, indicating strong under-representation of Poaceae plants (Xu et al., 2005, 2007, 2014). Not taking into account these differences in pollen representation between Artemisia, Chenopodiaceae, and Poaceae would lead to an overestimate of Artemisia and Chenopodiaceae and an underestimate of Poaceae communities in grasslands. The predominance of Artemisia and Chenopodiaceae compared with Poaceae pollen in sand-paleosol section, therefore, does not prove the predominance of Artemisia and Chenopodiaceae over Poaceae in the associated grassland communities. The previous interpretation of the existence of a pre-modern Artemisia and Chenopodiaceae grassland could very well be an artifact of the strong over-representation of Artemisia and Chenopodiaceae pollen and under-representation of the Poaceae pollen signal.
All of the above evidence supports the notion that phytolith assemblages in section Daike originated from Poaceae-dominant communities rather than previously believed Artemisia and Chenopodiaceae grasslands. Combined with radiocarbon and thermoluminescence ages (Table S4, available online; Li, 1991a, 1991b; Qiu et al., 1992), it is proposed that Poaceae has dominated the Songnen grasslands since at least the early Holocene.
Implications for paleovegetation reconstruction in arid and semi-arid lands
In North China, especially in arid and semi-arid lands where eolian sediments are widely distributed (Liu, 1985; Zhu et al., 1980), the dearth of effective, uninterrupted evidence of paleovegetation dynamics has been a significant challenge for researchers (Yang et al., 2011). Direct paleovegetation indicators contained in the sand or sand-paleosol sequence are especially rare.
Phytoliths avoid some of the weaknesses associated with pollen analysis for paleovegetation and paleoenvironment studies in arid lands. Because they are mainly composed of SiO2, phytoliths weather slowly (Piperno, 1988; Wang and Lu, 1993). Phytolith grains are abundant and well preserved in both the sand and paleosol samples from the Songnen grasslands despite the oxidizing and abrasive conditions that occur in sand. Compared with other proxies, phytolith analysis provided this study with a more continuous paleovegetation history. It helped to refine existing knowledge about the vegetation composition in the Songnen grasslands. Phytolith assemblages provided the data to determine the existence and type of vegetation on dunes during the sand episodes. In addition, because grass phytoliths are diagnostic to the subfamily of Poaceae, they provided more information than has been available through pollen analysis, which only identifies Poaceae to the family level. Furthermore, whereas pollen of many taxa are transported far from the source plant, phytoliths enter the soil through in situ decay mechanisms; thus, phytoliths provide more local information on vegetation than pollen does. Consequently, phytolith analyses successfully refine the precision of paleoecological studies carried out in deserts. Paleoenvironmental studies of many other drylands would benefit from phytolith analyses.
Conclusion
This pilot study demonstrates that both the sand and paleosol sediments in the Songnen grasslands contain well-preserved phytoliths attributed to different families of grass. It presents the first evidence from North China about the vegetation that existed during the sandy layer episodes. Using relationships between modern vegetation cover and phytoliths present in the topsoil as references, phytolith assemblages in section Daike indicate that they originated from Poaceae-dominant communities. Independent-sample t-test and discriminant analysis results lead to rejection of the hypothesis of a widespread Artemisia-Chenopodiaceae ecosystem in the Songnen grasslands. Instead, combined with published radiocarbon and thermoluminescence ages, it is proposed that Poaceae has dominated the Songnen grasslands since at least the early Holocene. In addition, Ic values point to a C3 species (i.e. Pooideae) expansion as a response to mid-to late-Holocene climate deterioration. Moreover, compared with the paleosol sediments, more annual Chloridoideae species thrived in response to drier and likely windier conditions when the sandy layer accumulated. Because phytolith assemblages enable the differentiation of C3 and C4 grasses of Poaceae, we propose that phytolith analyses are valid and useful tools for investigating vegetation dynamics in arid regions.
Footnotes
Acknowledgements
We thank Hainan Wang, Honglong Chen, Zhuo Gao, and Hongpeng Zhang for their assistance in fieldwork. We are grateful to Yong Ge and Chutian Hu who performed the phytolith extraction experiment of some plants and photographed some representative phytolith morphotypes.
Funding
This work was financially supported by the National Natural Science Foundation of China (Grant No. 41471164, 41771214), the Shandong Provincial Key Laboratory of Depositional Mineralization and Sedimentary Minerals in Shandong University of Science and Technology (Grant No. DMSM2017007), the National Key Research and Development Project of China (Grant No. 2016YFA0602301), the National Natural Science Foundation for the Youth of China (Grant No. 41602194), and the Foundation for Public Welfare Project funded by the Ministry of Environmental Protection of China (Grant No. 201109067).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.