
Abstract
The article presents the results of a multi-proxy study investigating an alluvial deposit located in the coastal plain of Genoa (Liguria, Northwest Italy). A very deep core was sampled from 15 to 25 m below ground level to analyse sedimentological properties, pollen assemblages, non-pollen palynomorphs, plant macrofossils and micro-charcoal. The deposit, spanning from the second half of the 5th Millennium to the end of the 7th Millennium cal. BC, was characterized by an alternation of silty and peaty layers, and proved to be rich in plant remains. Pollen data allowed the confirmation of a gradual increase in Fagus sylvatica to the detriment of Abies alba during the mid-Holocene of the region, and showed the presence of brackish lagoons, as well as the progressively stronger incidence of anthropogenic taxa, indicating the use of the plain for agricultural purposes. The study of non-pollen palynomorphs led to the taxonomic identification of some previously unidentified types, and pointed to the occurrence of marshy environments, which were probably grazed on by domestic herbivores. Macrofossil analysis provided the first record of Triticum new glume wheat type in Liguria. Moreover, the finding of a grain of Triticum dicoccum dated to the second half of the 6th Millennium cal. BC allowed us to pre-date by seven centuries the agriculture in the region. To date, this represents the oldest evidence of human activity in the urban area of Genoa.
Keywords
Introduction
The mouth of the Bisagno lies within a currently densely populated area situated immediately east of the natural amphitheatre that, as early as the Iron Age, was occupied by the early settlement of Genoa (Melli, 2014). A marked and uninterrupted urbanization since the second half of the 19th century (Manganelli and Melli, 2014) changed completely the morphology of this small coastal plain. As a result of this process, the stream bed underwent modifications, so that nowadays the water flows underground inside an artificial structure extending from the railway station of Genova Brignole to the sea (Figure 1).
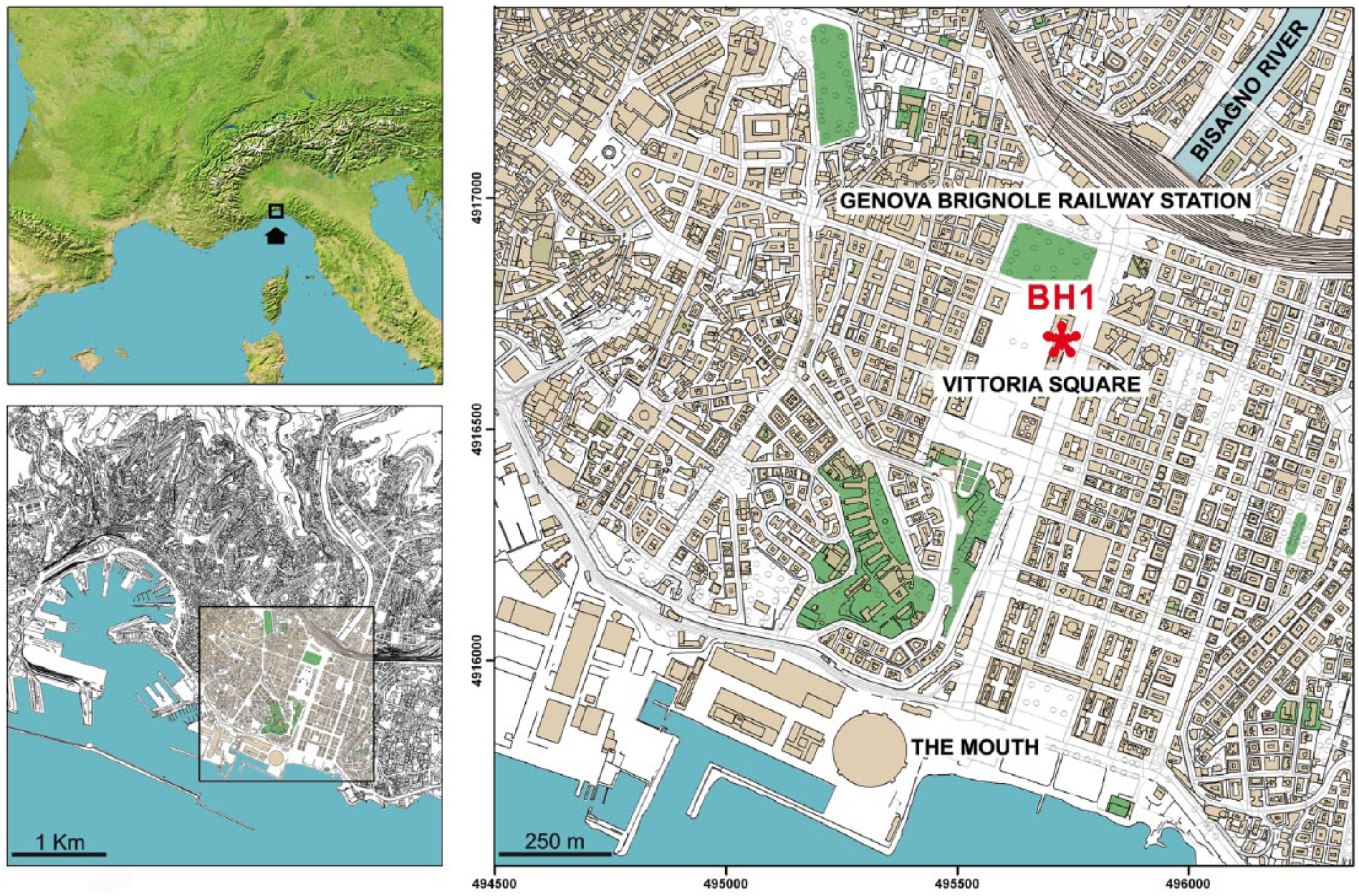
Location of the city of Genoa and core BH1 at Piazza della Vittoria.
The intense degree of urbanization is one of the major factors in the periodic and dramatic floods that, even recently, have affected the city causing great damage (Faccini et al., 2015, 2016), thus making extremely difficult the study of Holocene environmental change in the plain of Bisagno.
During the last decades, a number of archaeological finds have been recovered following public construction work in the area of the valley floor and on the nearby hills. These data pointed to human occupation of the plain as early as the late 6th Millennium cal. BC. The occupation of the territory and the resulting human impact since the mid-Holocene (Del Lucchese, 2014) are under investigation, due to the availability of several geognostic boreholes (c. 400). This work has been undertaken in the last 50 years and has allowed the University of Genoa to carry out archaeological and bioarchaeological investigations. The data recovered by multidisciplinary studies on the first anthropization of the urban area of Genoa turned out to be essential for the reconstruction of the changes occurred in the plain of Bisagno during the middle Holocene (Arobba et al., 2016, 2017a; Bonci et al., 2014; Montanari et al., 1997). The study has allowed us to identify the presence of currently locally extinct species and to recognize new non-pollen palynomorph (NPP) types, as well as to reveal the earliest traces of human presence in the area, in relation to agricultural practices documented by cereal macro-remains.
Materials and methods
The study area
The drainage basin of the Bisagno extends over an area of c. 95 km2 and shows a complex form following the two main axes N-S and E-W, the highest point having an altitude of 1037 m a.s.l. (Mt. Candelozzo). Due to neotectonic processes, the basin shows asymmetrical transverse sections, and much more developed secondary catchment areas on the right bank. The principal axis runs over c. 25 km, and the underground stretch near the mouth is still subject to abundant sediment accumulation, in spite of a number of intake structures. This area is nowadays comprised between the railway station of Brignole and the trade fair centre of Genoa.
The core BH1 was extracted between April and May 2006 in the urban area, at Piazza della Vittoria (44°24′12″N; 08°56′47″E) (Arobba et al., 2016) and reached a depth of 150 m from the current ground level (7.3 m a.s.l.). The stratigraphic sequence of BH1 starts at a depth of 3.4 m, below the crawl space of a modern building. The first horizon, about 1.1 m thick, consists of backfill sediments followed by coarse alluvial levels formed by brown sandy-silty gravels, up to 14.3 m. From this depth to 24.8 m, a series of dark grey and clayey poorly sandy silts occur, intercalated by organic-rich peaty levels.
Between 29.4 and 44.6 m, greyish/brown clayey and poorly clayey/poorly sandy gravelly silts occur, interrupted from 36.5 to 39.4 m by a succession of gravelly sands embedded in an ochre silty matrix. At a depth of 45 m, the Pliocene formation of Ortovero shales occurs, replaced at a depth of 111.5 m by the Mt. Antola Flysch Cretaceous bedrock, which persists up to the end of the borehole, 150 m below ground level.
The core was taken with a sampler having a diameter of 101 mm, and 43 samples averaging 10 cm in thickness were taken between the depths 15.10 and 25 m for sedimentological and palaeobotanical analyses (Figure 2). The part of the core between 15.25 and 16.05 m was collected as an undisturbed sample for preliminary geognostic assessments, and therefore it was not available for the additional sampling carried out for this study.

Photograph and schematic lithological description of core BH1 (25/15 m) with 14C dates, sedimentary facies, depth of the samples, organic matter content and texture (clay, silt, sand).
Radiocarbon dating
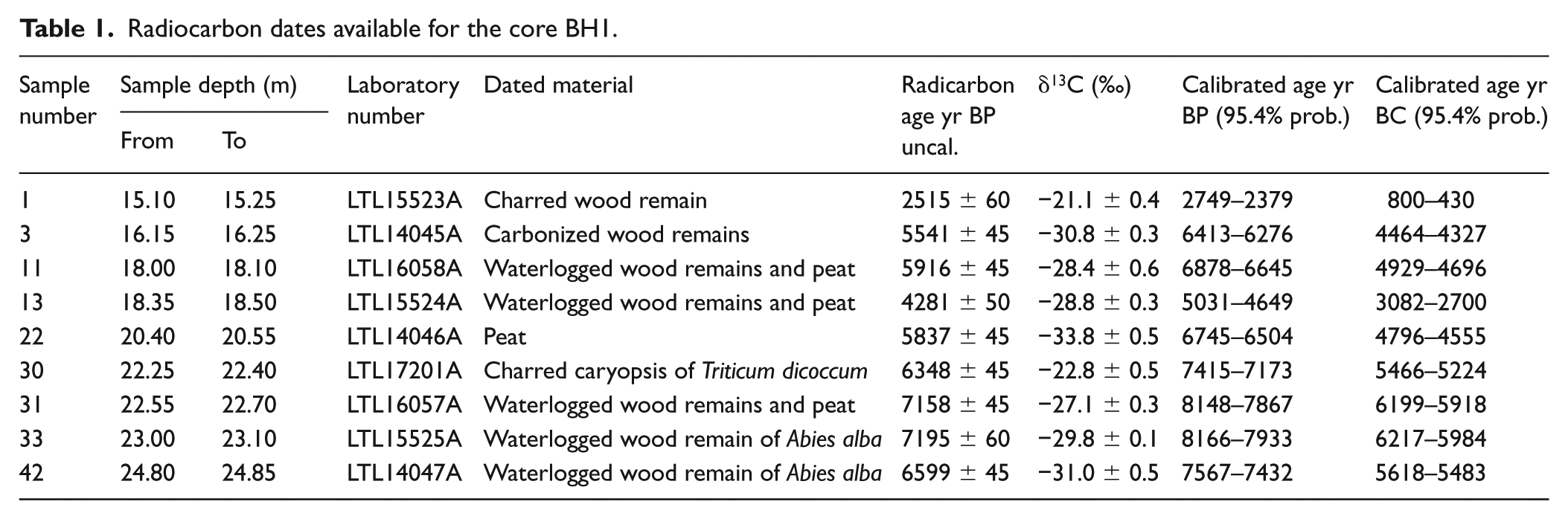
Overall, nine samples consisting of plant macro-remains were radiocarbon-dated by accelerator mass spectrometry (AMS) at the Cedad Laboratory of the University of Salento. The samples were collected from the part of the core between 15.10 and 24.85 m. Due to the state of preservation, it was possible to determine the species only for two uncharred wood fragments of Abies alba (LTL15525A, LTL14047A) and a charred caryopsis of Triticum dicoccum (LTL17201A). The remaining dates were obtained from charred, carbonified and uncharred wood from unidentified species and peat fragments.
Particle size analysis
In total, 43 samples were taken along the sequence for granulometric analysis. The coarse fraction, up to 63 µm, was analysed with a standard sieve column using a mechanical siever, whereas the finer portion was determined with the aid of an x-ray Sedigraph.
Loss on ignition analysis
All of the samples were subject to loss on ignition (LOI) analysis, in order to estimate the organic matter content (%OM) (Ball, 1964; Dean, 1974; Heiri et al., 2001).
Pollen, NPP and micro-charcoal analysis
The 43 samples used for sedimentological analysis were also used for palaeobotanical investigations. For pollen analysis, 10 to 20 g of sediment were treated according to standard procedures (Moore et al., 1991) and heavy liquid separation (Goeury and de Beaulieu, 1979; Nakagawa et al., 1998). Lycopodium spore tablets were added to each sample to evaluate pollen, NPP and micro-charcoal concentration (Stockmarr, 1971). Pollen and spore types were identified under the light microscope Leitz Diaplan at 400–1000× using the reference collection of the Archaeological Museum of Finale, as well as pollen keys and photo atlases (Beug, 2004; Moore et al., 1991; Reille, 1992, 1995, 1998). In each pollen slide, about 300 pollen grains were counted. Percentage values were calculated based on the total arboreal (AP) and non-arboreal (NAP) terrestrial pollen excluding spores of Filicales, Equisetum and NPPs, due to their high abundance compared with the other components.
For the determination of pollen of Cerealia-type, the parameter indicated by Joly et al. (2007) was followed, based on the use of a biometric threshold of two values (47 µm for grain diameter and 11 µm for annulus diameter). This allows accidental inclusion in the cereal category of some halophytic or wild grasses (e.g. Ammophila littoralis, Glyceria fluitans, Hordeum murinum) to be avoided.
As regards pollen of Cichorieae, only fenestrate grains smaller than 44 µm have been observed (Florenzano et al., 2015). The Human Impact Index was calculated based on the pollen sum of synanthropic species and cereals relative to the total AP (Accorsi et al., 1989).
Pollen Assemblage Zones (PAZs) and Non-Pollen Palynomorph Assemblage Zones (NPPAZs) were detected with the aid of the program ZONE 1.2 (Juggins, 1991), and to facilitate distinction they were named BH1-1/3 and GPV-1/12, respectively.
For the study of NPPs, a count of a minimum of 200 fungal/algal spores and hyphopodia per sample were reached (averaging 244, ranging from 201 to 659), except for seven samples less rich in microfossils, whose counts range from 110 to 183 NPPs (mean 141). A Leica DME compound microscope was used for the analysis (400×). A high number of small or particularly significant microfossils have been identified in oil immersion at 1000×. NPP values are expressed as percentages of total fungal/algal spores + hyphopodia (TNPP sum), following Cugny et al. (2010) and Mazier et al. (2009). Chironomid remains, testate amoebae, parasite eggs, flatworm eggs, stomata, sporangia, other microfossils of animal origin and fungal fruit bodies are reported as TNPP%. Newly identified microfossils were labelled with the code UR (University of Reading), followed by a sequential number. In order to avoid unnecessary multiplication of type codes, when the identification was certain, the codes that appeared in the previous literature have been used (Miola, 2012; Revelles et al., 2016; Shumilovskikh et al., 2015).
The abundance of microscopic charcoal, counted on the pollen slides following Tinner and Hu (2003) and Finsinger and Tinner (2005), was calculated as concentration values (mm2/g), obtaining data for regional (50–125 µm) or local (>125 µm) fire events.
All the diagrams (pollen, NPPs and micro-charcoal) were drawn using TILIA Software (Grimm, 2011).
Plant macrofossil analysis
This category of remains was extracted from the residues of the samples used for palynological and sedimentological analyses, by sieving through a 0.2-mm mesh. The amount of sediment analysed is between 40 and 280 g per sample (90 g on average). Plant macro-remains were observed and identified under an Olympus SZH10 (10–70×) stereomicroscope, using the reference collection of the Archaeological Museum of Finale and specific atlases (Anderberg, 1994; Berggren, 1969; Bojnanský and Fargašová, 2007; Cappers et al., 2006; Ercole et al., 2012; Jacomet, 2008). The results were elaborated using TILIA Software (Grimm, 2011) to create a histogram with values expressed as number of remains/100 g of sediment.
Results
Radiocarbon dating
Table 1 shows the results of the AMS radiocarbon dates from the nine selected samples. Calibration was carried out with the aid of OxCal, version 4.3.2 (Bronk Ramsey, 2009), using the IntCal13 atmospheric curve (Reimer et al., 2013).
Radiocarbon dates available for the core BH1.
Sedimentology and LOI
The largely prevailing granulometry consists of organic clayey silt, often having a high percentage of clay (see Figure 2).
Overall, the stratigraphy of the sampled part of the core appears as a succession of dark grey and poorly sandy clayey silt levels frequently intercalated by very organic-rich levels, sometimes actual peaty levels.
These organic horizons begin immediately underneath fluvial deposits consisting of sandy gravels that were found until the depth of c. 14.3 m. These deposits record a marked change in sedimentary processes and are probably related to a phase of river transgression.
The upper part of the sequence, between 16 and 17 m from the ground level, is formed by a dense alternation of silty-clayey levels with sporadic malacological remains and very organic-rich silty-clayey levels with numerous plant macro-remains. Between 18 and 20 m, three peaty levels occur, overlaid by light grey silty levels with poor organic content, concentrated into millimetre-thick laminations. Further deep, dark grey organic-rich silty-clayey levels prevail again. The deepest part of the sampled core is characterized by slightly sandier levels with sporadic rounded clast and rare organic components, a composition similar to the lower part of the sequence down to the Pliocene deposits.
The average %OM is around 13%, but higher values were recorded in a few peaty levels (from 21% to 32%, Samples 11, 13, 14, 22, 31 and 35). Sample 43, taken from the lowest sampled part of the core (depth 24.9/25 m), marks the beginning of a textural change highlighted by the presence of poorly gravelly and gravelly silts, almost lacking any organic matter. Plant macro-remains included in the peat account for an over-representation of the sand fraction in Sample 22 (see Figure 2).
Pollen analysis
Figure 3 shows the dendrogram used for the identification of different PAZs, whereas Figures 4 and 5 show the complete palynological diagram and the curves of a number of pollen categories of particular ecological interest. A specific curve represents the Human Impact Index.
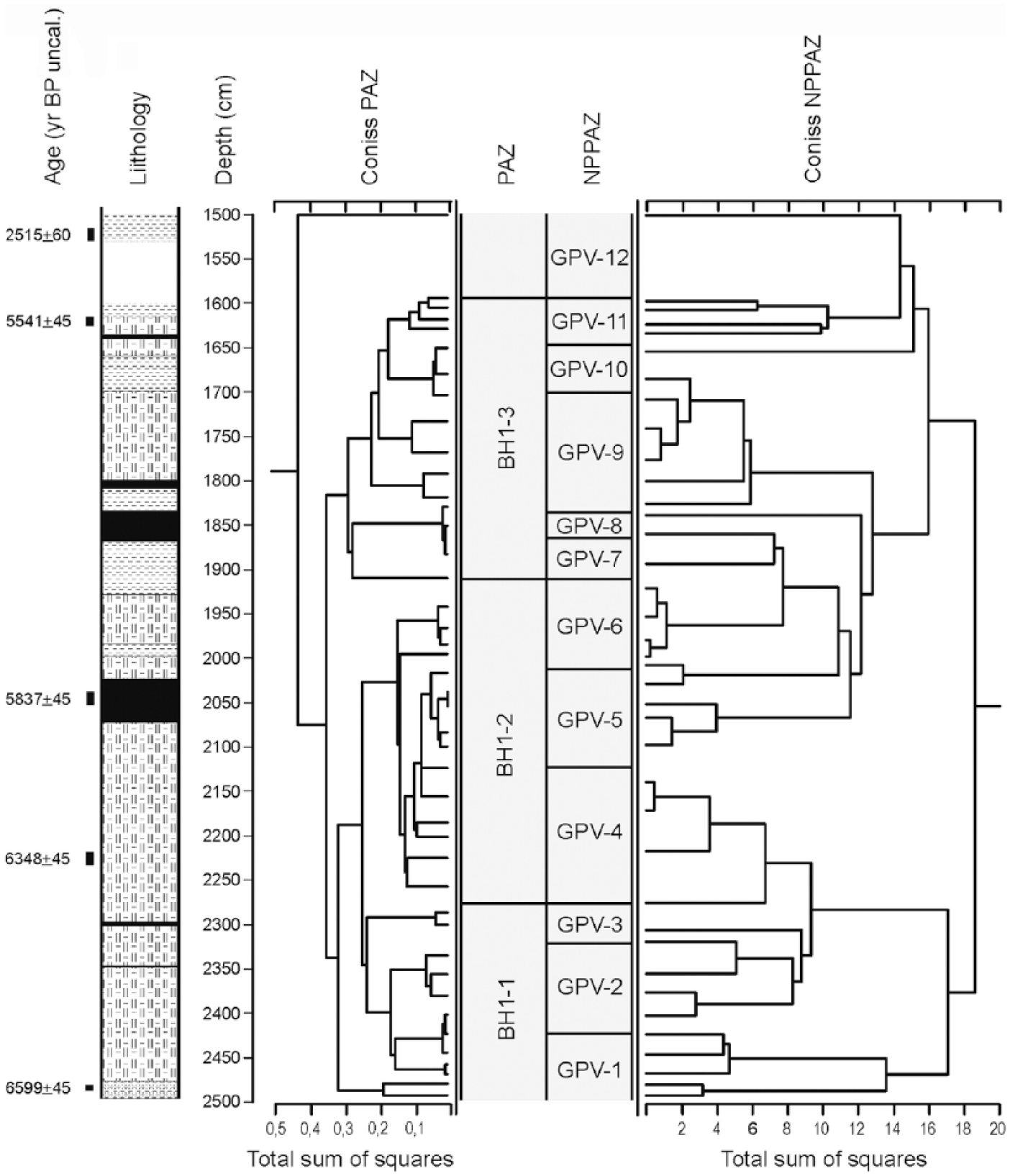
Dendrograms used as an aid to the subdivision into PAZs and NPP-related subzones (NPPAZs).
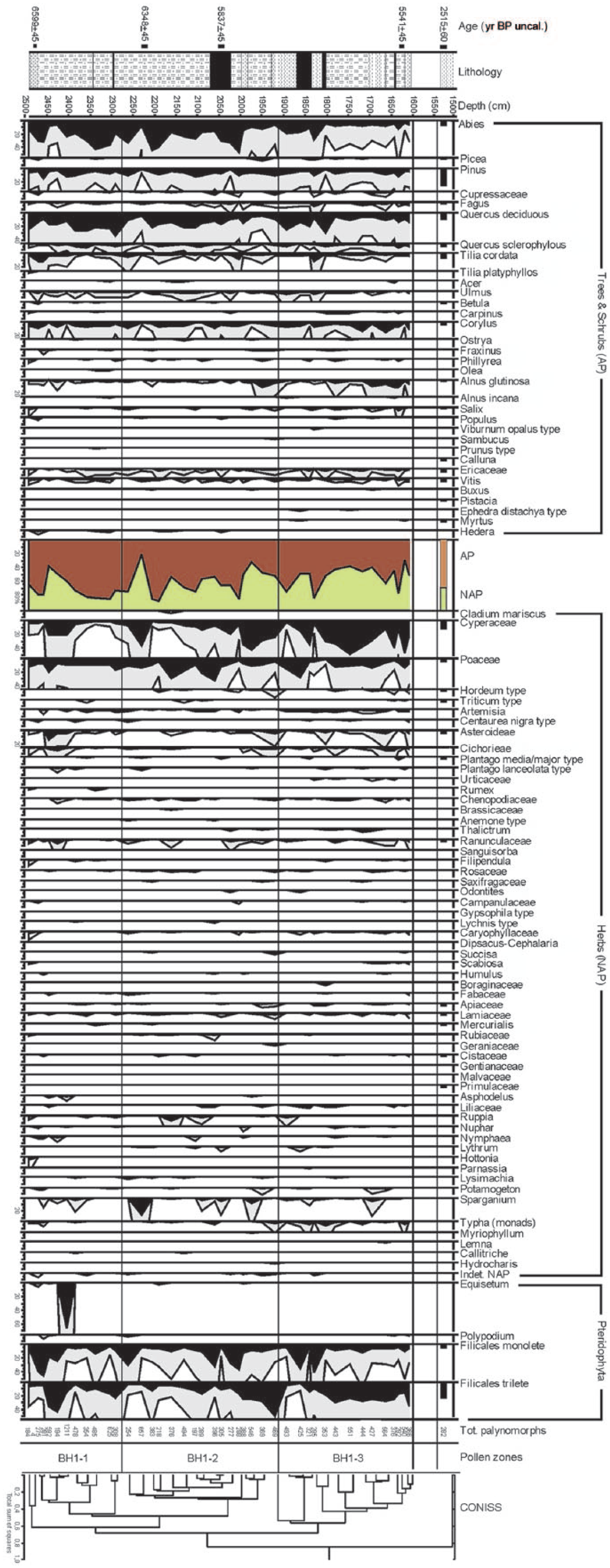
Pollen diagram (grey curves represent a 5× exaggeration factor).
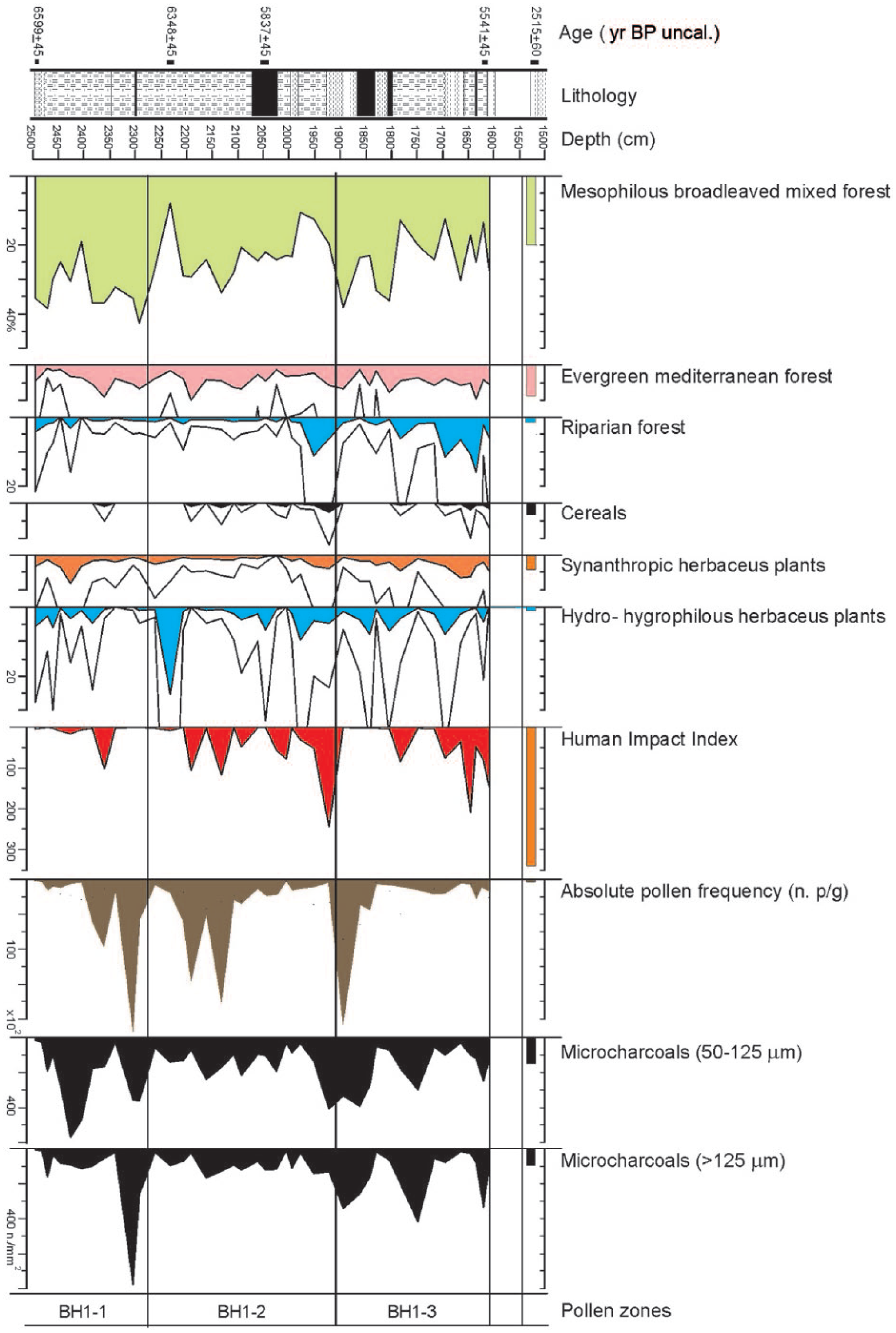
Pollen diagram showing selective cumulative curves of ecological interest, Human Impact Index, total pollen and micro-charcoal particles of regional (50–125 µm) and local (>125 µm) provenance.
The same diagram shows as a histogram of the pollen spectrum from the isolated sample taken at the top of the sequence (15.17 m, Sample 1), in order to show its discontinuity with the continuous sequence comprised between 25.00 and 16.05 m.
A large number of palynological taxa (93) were identified, including 32 arboreal and 61 herbaceous entities, with high pollen concentrations (averaging 36,308 palynomorphs per gram of sediment, ranging from 57 to 221,071).
PAZ BH1-1 (from 25.00 to 22.77 m.; c. 5500–5000 cal. yr BC)
Looking at the total AP/NAP ratio, a relatively high tree cover stands out, with maximum values around 86%, interrupted by short periods showing a drop to 39%. The dominant taxon is Abies alba, with peaks reaching 43%, followed by Pinus and Quercus deciduous (up to 35%). Along with these, we find a group of broad-leaved species typical of the mesophilous and thermophilous oak wood (Tilia, Acer, Ulmus, Carpinus, Corylus and Ostrya), followed by Betula, heliophilous colonizing species, and by riparian trees such as Alnus and Salix. Pollen of Vitis vinifera (1.8%) also occurs, an element of humid mesophilous wood. Bushy taxa typical of the Mediterranean maquis are also relevant (4%). The herbaceous component mostly consists of Cyperaceae, whose maximum values reach 21% at the beginning of the zone and 5% towards the end, and of Poaceae, which are steady around 11%. The presence of cereal pollen at 23.60 (Triticum t. 1%) is also worth noting. Numerous hydro-hygrophilous herbaceous taxa occur (Nymphaea, Potamogeton, Hottonia, Nuphar and Sparganium), some of which relate to brackish environments (Ruppia). Synanthropic taxa (Artemisia, Centaurea nigra t., Cichorieae, Plantago media/major t., Plantago lanceolata t., Urticaceae, Rumex and Chenopodiaceae) occur with an average value of 2.6%.
PAZ BH1-2 (from 22.77 to 19.10 m; c. 5000–4500 cal. yr BC)
In this intermediate zone, the pollen percentages of arboreal species remain high, up to 88%, with the exception of drops with minimal values around (32%). These decreases are driven by the fluctuations of Abies alba and Quercus deciduous, while Pinus remains steady around 8%. Fagus, although already sporadically recorded in the lowest zone, reaches now relevant values, up to 3.4%. The taxa of the mixed oak woodland and Vitis vinifera, relatively infrequent species and low pollen producer, remain on similar values as before, while the arboreal riparian vegetation, formed mostly by Alnus glutinosa, expands slightly in the second half of the zone alongside a rise in Cyperaceae, accompanied by a reduction in Poaceae. Among the cereals, both Triticum t. (0.9%) and Hordeum t. (2.4%) are present. The Mediterranean vegetation does not change (4.1%), whereas hydro-hygrophilous plants show values higher than in the previous zone: in particular, Ruppia peaks in the initial part (7.5%) and Typha in the final part of the zone (up to 3.3%). Synanthropic species markedly decrease up to 1.3%.
PAZ BH1-3 (from 19.10 to 16.10 m; c. 4500–4200 cal. yr BC)
The arboreal component appears to be decreasing compared with the two earlier zones, with values averaging 53%. There is clear drop in Abies alba, while Pinus, Fagus and the mesophilous oak wood keep steady on the previous values. The Mediterranean bushy-arboreal component (Quercus ilex t., Phillyrea and Myrtus) shows a slight increase, averaging 5%. The riparian vegetation appears more common, as indicated by Alnus glutinosa (up to 10.6%) and Salix (up to 5.3%). Vitis vinifera is constantly present, with a maximum value of 2.2%. Cereals are slightly decreasing, being Triticum t. at 0.4% and Hordeum t. at 1.6%. Among the other herbaceous taxa, grains of Cyperaceae are particularly abundant and reach here their highest peak (48.8%), whereas Poaceae (10%) occur with values similar to the previous zone. In spite of a decrease in the percentage values, the above-mentioned hydro-hygrophilous species are still present, except for the disappearance of Ruppia. In this phase, synanthropic species appear to rise significantly (3%).
Sample 1 (from 15.17 to 15.10 m)
The histograms show a total AP value of 69.5%, still maintained high by Abies alba, as well as by Pinus and elements of the mixed oak wood and Mediterranean flora (Pistacia, Myrtus, Ericaceae), while the components of the riparian wood decrease (1%). Within the NAP, the presence of Cerealia (3.3%) and synanthropic species (4.3%) should be stressed, as well as the floral dearth.
NPP analysis
A separate zonation was adopted to represent the main NPP assemblages and related ecological changes (NPPAZ; Carrión and Navarro, 2002; Cugny et al., 2010; Mazier et al., 2009; Miras et al., 2010). Therefore, the phases identified do not necessarily overlap with the pollen zones, as fungal, algal and plant communities respond to different driving factors (Baker et al., 2013). However, NPPAZ GPV-1 to 3 appear to be included within BH1-1, NPPAZ GPV-4 to 6 within BH1-2 and NPPAZ 7 to 11 within BH1-3, as highlighted in Figure 3, where pollen and NPP dendrograms appear correlated. The diagram in Figure 6 shows the trends of identified and undetermined NPPs. Some of the types mentioned in the text are illustrated in Figure 7. All new types, their possible biological identification and equivalence with previously described types are listed in the corresponding table (see Supplementary materials, available online).

NPP diagram showing selective cumulative curves of ecological interest (coloured curves represent a 5× exaggeration factor): 1. algae/aquatic organisms, 2. aquatic fungi, 3. saprophytes, 4. largely coprophilous, 5. occasionally coprophilous, 6. pathogenic/parasitic, 7. erosion, 8. plant material, 9. zoological and 10. intestinal parasites.
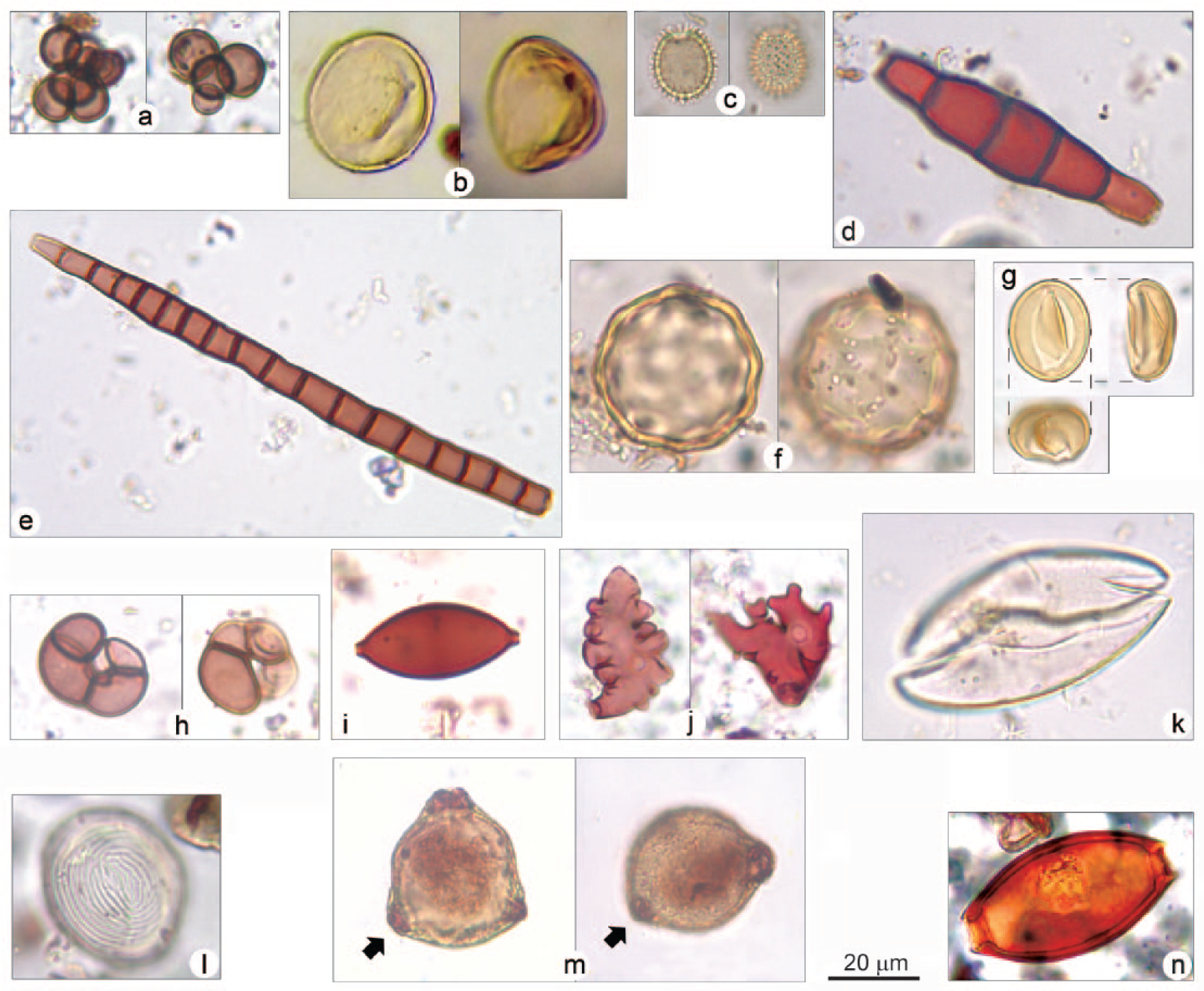
Non-pollen microfossils from BH1 core: (a) UR-12, cluster of three to seven nearly perfectly spherical and relatively thick-walled cells, psilate, pale brown; (b) UR-14, rounded, always concave on one side, psilate, yellowish; (c) UR-15, 1-2-porate, covered with short appendages, hyaline; (d) UR-17, 4-septate, slightly constricted at the septum, psilate, brown; (e) UR-18, multiseptate, not constricted at the septum, psilate, brownish/reddish; (f) UR-20, spherical, thick-walled, with slightly wavy outline due to a dense pattern of irregular depressions covering the surface, psilate, yellowish; (g) HdV-119, elliptical/rounded, always with an elliptical/polygonal rupture pattern on one side, concave–convex in lateral view, bilobate when seen from above, psilate, yellowish. The orthogonal projection illustrates its morphology and the relationship between the three views; (h) Cirrenalia basiminuta (HdV-120), four-celled, constricted at the septa, psilate, brownish/reddish; (i) HdV-55B, elongated with two large protruding apical pores, psilate, brown; (j) polylobate fungal hyphopodia (Buergenerula spartinae?); (k) Spirogyra; (l) Pseudoschizaea; (m) pollen grain of Corylus showing darker fungal material inside the pores; (n) egg of Trichuris sp.
NPPAZ GPV-1 (from 25.00 to 24.05 m)
This zone is characterized by the dominance of UR-15 (25%) and a rise in UR-20 (14%), coupled with a slight increase in Glomus (7%). Among the aquatics, Zygnema t. spores are recorded.
NPPAZ GPV-2 (from 24.05 to 23.05 m)
Pseudoschizaea increases (2.5%), along with Spirogyra (3%). Coniochaeta decreases (7%), while UAB-31 and Cirrenalia basiminuta reach high values (15%).
NPPAZ GPV-3 (from 23.05 to 22.77 m)
A peak in fungal hyphopodia occurs (5%), but the dominant taxa are Coniochaeta and UR-17, reaching 24%.
NPPAZ GPV-4 (from 22.77 to 21.32 m)
Coniochaeta occurs in lower percentages (c. 10%), whereas Cirrenalia basiminuta (22%) and UR-12 (15%) become dominant. A rise in fern sporangia occurs (5%).
NPPAZ GPV-5 (from 21.32 to 20.05 m)
The zone is characterized by a sharp increase in Spirogyra (16%), along with a new rise in Coniochaeta (37%), a peak in Kretzschmaria deusta (12%) and the first considerable rise in Sporoschisma (7%).
NPPAZ GPV-6 (from 20.05 to 19.10 m)
A new rise in Pseudoschizaea occurs (8%), along with the first notable increase in Glomus (12%). Sporoschisma reaches its highest value (10%), coupled with Cercophora t. (7%).
NPPAZ GPV-7 (from 19.10 to 18.62 m)
Glomus decreases (less than 3%), while Cirrenalia basiminuta becomes dominant (25%), along with UAB-31 (20%).
NPPAZ GPV-8 (from 18.62 to 18.30 m)
The zone is defined by a new peak in Spirogyra (9%), paralleled by a sharp rise in Sordaria t. (8.5%) and Coniochaeta (40%).
NPPAZ GPV-9 (from 18.30 to 16.95 m)
A second notable rise in Glomus occurs (30%), along with an increase in Cercophora t. (7%). Among the aquatics, Pseudoschizaea rises again (4%). UR-18 reaches remarkable values (4%).
NPPAZ GPV-10 (from 16.95 to 16.45 m)
The zone is largely dominated by HdV-119, which reaches extremely high percentages (69%).
NPPAZ GPV-11 (from 16.45 to 16.10 m)
A very sharp rise in fungal hyphopodia occurs (46%), along with a rise in Coniochaeta (38%), UAB-4 (13%) and Pseudoschizaea (12%). HdV-200 reaches high values (18%).
NPPAZ GPV-12 (from 15.25 to 15.10 m)
The zone is dominated by UR-14 (53%), preceded by a rise in UR-15 (12%). The main aquatic taxon is represented by Pseudoschizaea (5%).
Micro-charcoal analysis
Figure 5 shows the concentration curves for microscopic charcoal remains. During BH1-1, charred particles from a regional scale prevail, with a maximum peak of 572 mm2/g. On the other hand, the local component shows in the initial part a maximum of 115 mm2/g and then a very marked peak (778 mm2/g) related to fires in the close surroundings of the zone of sedimentation. In the middle zone BH1-2, the trend of the two curves appears to nearly overlap, with a variation of 240 and 409 mm2/g at most for the regional component and averaging 91 mm2/g for the local one. During BH1-3, we observe a varying and synchronous trend of the curves, with maximum values of 395 mm2/g for the regional component and 420 mm2/g for the local component. However, any consideration regarding the significance of these components needs to take into account the type of deposition characterizing deltaic environments, such as our site, where low-energy sedimentation occurs. The incidence of charred particles, including the larger fraction, may then partly result from water transport following hillslope run-off within the catchment area, subjected to forest fires.
Plant macrofossil analysis
The diagram in Figure 8 shows the abundance of different carpo-remains, while Figure 9 shows their sum, calculated on the basis of the categories of provenance: wet meadows/marsh, aquatic plants, fresh/salt marsh, riparian forest, deciduous trees, cereals and other habitats. Leaf remains only belong to Abies alba and Pinus sp. Most of macrofossils are waterlogged (n = 1329); only in a few cases (n = 8), they are charred (cereals, Euphorbia helioscopia and Vitis vinifera ssp. sylvestris). Figure 10 shows some seeds/fruits of particular interest.

Plant macrofossil diagram (charred remains marked by asterisks).
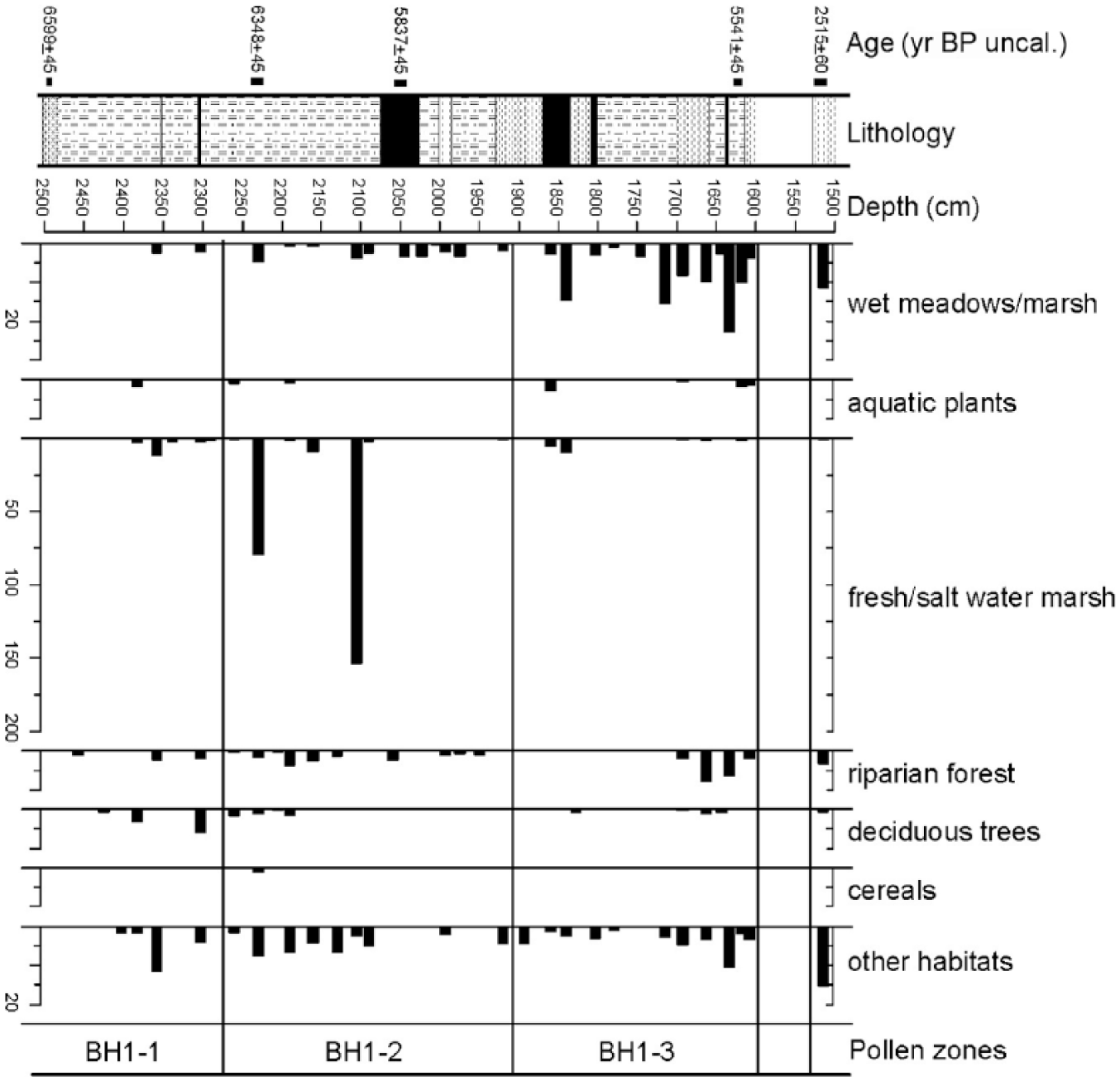
Plant macrofossil diagram showing cumulative curves of selected taxa.

Waterlogged and charred plant remains (bar = 1 mm): (a) Abies alba (leaves), (b) Cornus sanguinea, (c) Rubus fruticosus s.l., (d) Rubus caesius, (e) Rubus idaeus, (f) Fragaria vesca, (g) Alnus glutinosa, (h) Sambucus ebulus, (i) Vitis vinifera ssp. sylvestris, (j) Physalis alkekengi, (k) Sanguisorba officinalis, (l) Rumex conglomeratus, (m) Verbena officinalis, (n) Zannichellia palustris, (o) Lycopus europaeus, (p) Potamogeton perfoliatus, (q) Polygnonum amphibium, (r) Bolboschoenus sp., (s) Cladium mariscus, (t) Mentha aquatica, (u) Baldellia ranunculoides, (v) Euphorbia helioscopia, (w) Hordeum vulgare, (x) Triticum dicoccum, (y) cf. Triticum NGW type.
PAZ BH1-1
The samples from the earliest zone appear relatively poor in macro-remains. The presence of leaves of Abies alba is particularly worth noting, along with seeds/fruits of plants from the understory of Abies forests (Rubus idaeus) and from mesophilous coenoses of hillslopes (Rubus fruticosus s.l. and Rosaceae cf. Prunus), along with taxa testifying to the existence of marshy environments with a various degree of salinity (Bolboschoenus sp., Carex divisa, Potamogeton coloratus). The finding of seeds of Vitis vinifera ssp. sylvestris and carpo-remains of Sambucus nigra/racemosa, mesophilous elements of occasionally relatively humid uplands, deserves a particular mention.
PAZ BH1-2
The components of oxbow lakes and fresh/brackish water prevail (Cladium mariscus, Bolboschoenus sp., Carex sp. and Stratiotes aloides), and along with elements of the riparian wood (Alnus glutinosa, Sambucus sp., Physalis alkekengi, Rubus idaeus and Rubus sp.), they partially replace the deciduous wood. Leaf remains of Abies alba show a peak at 21.91 m and progressively decrease until 21.20 m. An increase in the herbaceous cover and floral richness can be observed (Medicago littoralis, Campanula sp., Galium sp., Plantago sp., Allium sp., Cirsium sp., Arctium sp., Asteraceae).
At the bottom of this zone, two peaks of Bolboschoenus sp., a species inhabiting humid areas with fresh or brackish water, should be stressed. At a depth of 22.32 m, evidence for cereal cultivation was found, as shown by the only charred remains recovered: a radiocarbon-dated caryopsis of Triticum dicoccum, a spikelet base perhaps belonging to the Triticum NGW type, and a caryopsis of Hordeum vulgare var. tetrastichum.
As regards Triticum new glume wheat type in particular, this glume base is extremely large and differs from the measurements known for Triticum dicoccum (Kenez et al., 2013). It seems to also have a massive and distinct keel that does not typically occur in emmer and einkorn. This attribution, based on a single fragmented remain, for the moment should be considered hypothetical and would be confirmed in case of newer and clearer evidence from the area of the site.
At the same depth, a few charred seeds of Vitis vinifera ssp. sylvestris and Euphorbia helioscopia were found.
PAZ BH1-3
A marked increase in the floral components of wet meadows markedly occurs (Blysmus compressus, Carex sp., Cladium mariscus, Mentha aquatica, Allium cf. schoenoprasum, Lycopus europaeus, Sanguisorba officinalis, Hydrocotyle vulgaris, Baldellia ranunculoides), as well as in elements of the riparian wood (Alnus glutinosa, Rubus caesius, Rubus sp.) and of still or slowly moving water (Zannichellia palustris and Potamogeton perfoliatus). Nitrophilous and trampling-related species appear (Atriplex sp., Ranunculus sp. p.p., Solanaceae, Verbena officinalis and Senecium sp.).
Sample 1
In the separate sample, among the species favouring humid environments, only Blysmus compressus and Carex sp. occur. The abundance of carpo-remains of the riparian wood (Physalis alkekengi, Rubus caesius, Sambucus sp.) and deciduous wood with Vitis vinifera ssp. sylvestris is low. Coastal halophytes and synanthropic species, among which Chenopodium sp., Atriplex sp. and Verbena officinalis, remain present.
Discussion
Dating
The radiocarbon dates available for this portion of the coring are apparently distributed over a rather long chronological period from the early 1st Millennium to the end of the 7th Millennium cal. BC (see Table 1). The result for the lowermost sample, taken between 24.80 and 24.85 m and obtained from an uncharred wood fragment of Abies alba (LTL14047A, 6599 ± 45 BP, 5620–5480 cal. BC), shows a more recent date than those from the two overlying samples, comprised between 23.10 and 22.55 m (LTL15525A, 7195 ± 60 BP, 6220–5980 cal. BC and LTL16057A, 7158 ± 45 BP, 6200–5920 cal. BC) and must therefore be considered intrusive. The two following dates, obtained by a caryopsis of Triticum dicoccum and a peat sample (LTL17201A, 6348 ± 45 BP, 5470–5220 cal. BC and LTL14046A, 5837 ± 45 BP, 4800–4555 cal. BC), should instead be considered correct, whereas various factors suggest to consider the dates from the samples between 18.50 and 18.00 m as unreliable. Being 1500 years later than the adjacent dates, the first date (LTL15524A, 4281 ± 50 BP, 3080–2700 cal BC) could be related to intrusive material, while the second one (LTL16058A, 5916 ± 45 BP, 4930–4700 cal. BC) may be due to the presence of a multi-centennial tree. The sample taken between 16.25 and 16.15 m, dated to 5541 ± 45 BP (LTL14045A, 4470–4330 cal. BC), appears to be reliable.
Finally, the date from the most superficial part of the sequence, between 15.25 and 15.10 m (LTL15523A, 2515 ± 60 BP, 800–430 cal. BC), appears to be 3500 years more recent than the immediately underlying one, taken from a not much deeper sample (c. 1 m). This result, which does not appear to be justified by the presence of sedimentary gaps at this depth, is likely to be considered as an intrusive episode from the above levels of sandy gravels.
Considering the level of detail of the analyses, the relative scarcity of dates and their irregular distribution in the profile impeded any attempt to build a reliable age-depth model, either using Clam 2.2 (Blaauw, 2010) and Bacon (Blaauw and Christeny, 2011), and therefore it was decided to avoid presenting this kind of model for BH1. The part of the core comprised between 16.15 and 23.10 m of depth seems thus to pertain to a time span of almost 1900 years between the end of 7th and the first half of the 5th Millennium cal. BC.
Geomorphological setting and sedimentology
The series of boreholes carried out since 1993 for public construction work at Piazza della Vittoria has allowed an evaluation of the formation processes that created the delta plain of the Bisagno (Bonci et al., 2014; Colombi, 1996; Firpo, 1996).
The sequences recorded in the area in the previous years, up to a depth of 33 m below road level, already showed the occurrence of coarse sediments in the upper levels and silty-clayey horizons in the lower part, often rich in organic matter. These data point to an alluvial depositional environment and marshy areas, related to the mouth of the Bisagno (Montanari et al., 1997, 2014).
Similar horizons have been found also in the core BH1, where they are clearly evident at lower depths. In general, the final part of the alluvial plain of the Bisagno between the end of the 7th and the first half of the 5th Millennium cal. BC appears to be characterized by a diverse range of sedimentary sub-environments showing a highly varied textural and structural composition, even in areas located at a short distance from one another, as depending on the transport energy.
Reconstruction of landscape evolution by means of palaeobotanical analysis
First, the floristic richness recorded by the analysis of palynological taxa is a fact resulting from a mosaic of vegetation environments occurring in the same time span, and from an alternation of short-lived episodes that deeply impacted the forms of the territory.
BH1-1 and BH1-2 are characterized by a dense tree canopy. Indeed, Abies alba is present both in the pollen and in the macrofossil record, coupled with elements of oak deciduous wood capable of colonizing lower areas extending to the plain, as well as other features of the humid mesophilous wood (Guido et al., 2013; Muller et al., 2007). Being an element of the mesophilous understory, Vitis vinifera ssp. pollen and macro-remains also fit within this scenario. In both zones, micro- and macro-remains point to the presence of freshwater and brackish lagoons, connected by short watercourses along which riparian vegetation could thrive.
Evidence for cereal cultivation is already found in the pollen record since BH1, with grains of Triticum t. at 23.60 m, whereas Hordeum t. occurs in BH1-2 with values ranging from 1% (21.92 m) to 2.4% (19.22 m).
The rise in pollen of Fagus during BH1-2 is of interest, as in spite of being present in the previous phase, it reaches now substantial values, which will last in BH3. This situation, as well as the progressive decrease in Abies, is compatible with the changes in the environment occurring between 5800 and 3000 cal. yr BC, and probably also with an early human intervention affecting the tree cover of the hilly and montane belt of the backcountry (Branch and Marini, 2013; Cruise et al., 2009; Dubar et al., 2011).
Human impact is confirmed also by traces of agriculture, as shown by various macrofossils of Triticum dicoccum, Triticum new glume wheat type, Hordeum vulgare var. tetrastichum and seeds of Vitis vinifera ssp. sylvestris.
The presence of charred cereal caryopses coupled with a charred seed of Vitis vinifera is relevant, as it informs on the relationship between agricultural products and those presumably obtained through hand picking. As for Vitis vinifera, the finding of pollen across the whole sequence and the occurrence of macro-remains point to a special interest of the local population towards this species, to the point where a form of protection aimed at ensuring harvesting of the fruit may have been in place (Arobba and Caramiello, 2018; Cremaschi et al., 2016). These macrofossils were all recovered from the same level (22.32 m), and they are the only charred carpo-remains present in the whole sequence, along with a seed of the synanthropic species Euphorbia helioscopia.
A slight increase in riparian pollen and macro-remains can be observed in BH1-3, as well as the highest value of Cyperaceae and elements characterizing humid meadows. The other pollen categories examined are herbaceous hydro-hygrophilous freshwater and brackish species and synanthropic taxa, all of them present in the three zones with a slight increase in the third one.
Among the hydrophytes, two species, currently absent in Liguria according to the recent floristic literature, deserve a special mention: Stratiotes aloides, only documented in BH1-2, and Hydrocotyle vulgaris, appearing in BH1-2 and then more abundant in the last part of BH1-3 (Pignatti, 2017). Both of them were not recorded in the latest uppermost level.
In the top sample, arboreal species consist of Pinus, elements of mixed oak wood and Mediterranean flora. Here, the presence of cereal pollen and synanthropic species also increases significantly.
Ecological interpretation by means of NPP analysis
The abundance of aquatic microfossils allows some remarks on the hydrology of the site. A first sharp increase in Spirogyra during GPV-5 points to shallow and stagnant water, as supported by a rise in fungi predominantly known from submerged habitats (Sporoschisma and Bactrodesmium sp.; Hu et al., 2010). The data from GPV-8, showing a new increase in Spirogyra spores, suggest a shallow and eutrophic water body, about 0.5 m deep or less (Van Geel, 2001). This environment is likely to have attracted herbivores as a watering place, as shown by a sharp increase in Sordaria t., coupled with Sporormiella t. and Podospora decipiens t. (Schlütz and Shumilovskikh, 2017). In this regard, it is worth mentioning that although Spirogyra can also inhabit brackish pools (Aleem, 1961), drinking salty water has no significant negative effects on cattle health (Visscher et al., 2013). These variations in the abundance of algal microfossils across the deposit can be accounted for by the presence of temporary oxbow basins, formed from meanders of the Bisagno over subsequent flood events.
Increases in Pseudoschizaea can also be observed. Ecologists do not have a univocal view on its indicator value, although it is mostly thought to originate from an unknown aquatic organism (Grenfell, 1995; Scott, 1992; uncertain results possibly pointing to a relationship with angiosperms based on DNA extraction were reported by Milanesi et al., 2006). This microfossil seems to be an indicator of summer drought and periodical drying out of wet areas in warm climates (Carrión and Navarro, 2002), and it is normally considered to represent a freshwater algal cyst. Encystment is a well-known strategy used by phytoplankton species in order to survive phases of extreme environmental conditions (Blackburn and Parker, 2005; Bold and Wynne, 1985). It is thus likely that alternate desiccation and flooding of the area triggered algal encystment during dry periods.
The lower part of the sequence is characterized by higher percentages of wood saprophytes, which might point to a denser tree canopy, as well as by sporadic peaks in a range of unidentified microfossils: UR-12, UR-15, HdV-1036 and UAB-31 (Revelles et al., 2016). The presence of HdV-120 is noteworthy. The type was first recorded in a brackish lake by Pals et al. (1980). It seems possible to associate this conidium with the occurrence of relatively salty pools, as its morphology points to identification with Cirrenalia basiminuta (Marine Lignicolous Fungi Database, 1999; Raghu-Kumar et al., 1988), a cosmopolitan hyphomycetes consistently collected in environments characterized by some degree of salinity (El-Sharouny et al., 2009; Leong et al., 1991).
Infection of pollen grains by fungal hyphae filling the apertures was also noted. The process has unknown causes and was first reported in the palaeoecological literature by Shumilovskikh et al. (2015). A relationship with dry phases, as suggested by these authors, may also apply to our site, subjected to wet-dry cycles.
The remarkable dominance (69%) of HdV-119 during GPV-10 requires further investigation. This microfossil shows a characteristic opercule-like aperture, somewhat resembling the one found on certain dynoflagellate cysts (e.g. Brigantedinium sp.; Leroy and Albay, 2010), along with a concave–convex lateral view (Deforce, personal communication, 2017).
The type has been recorded in Mediterranean and northern European contexts, especially in aquatic environments characterized by stagnant or slow-moving waters and a certain degree of salinity (Carrión and Navarro, 2002; Demey et al., 2013; Pals et al., 1980).
An important episode of plant parasitism occurs at the top of the sequence (GPV-11). Previous research has highlighted the match between polylobate fungal hyphopodia and Carex species (e.g. Van Geel et al., 1983). However, it is not possible to observe this correlation in our study, and it should be pointed out that very similar hyphopodia may be produced by different genera (Pirozynski et al., 1988). It is thus possible that the sharp rise here recorded resulted from a different host–pathogen association. To this end, it is worth stressing that closely resembling hyphopodia of Buergenerula spartinae are typical of brackish environments along the coasts of northern and south-western Europe (Kohlmeyer and Gessner, 1976).
Data on the local presence of omnivores are provided by eggs of intestinal parasites. In GPV-8, a Trichuris egg measuring 48.6 µm in length falls within the size range of whipworms infecting humans (Trichuris trichiura) and pigs (Trichuris suis) (Beer, 1976). The other eggs occurring at the top of the sequence (GPV-12) appear particularly small (ranging from 31.6 to 40.8 µm), whereas on average eggs of Trichuridae range from c. 50–65 µm (Trichuris trichiura, Trichuris suis, Capillaria sp.) to c. 70–85 µm (Trichuris vulpis, Trichuris ovis) (Beer, 1976; Brinkkemper and van Haaster, 2012; Florenzano et al., 2012; Fugassa et al., 2008; Sondak, 1948; Traversa et al., 2011), and only a few other cases of unusually short eggs are known in the literature (Beer, 1976; Sondak, 1948). The eggs from GPV-12 also appear to be rather degraded and it is not possible to observe features such as surface ornamentation and slight asymmetry of polar plugs, elements necessary to confidently separate between Trichuris and Capillaria spp. (Lartigot-Campin, 2014; Traversa et al., 2011).
Although the size of the eggs is compatible with Trichuris suis, domestic pigs were introduced in Liguria only during the Late Neolithic (Rowley-Conwy, 1997). However, Trichuris suis can also infect wild suids and humans (Beer, 1976), and bones of wild boar/pig were found very near the coring spot in archaeological layers at Brignole (Fontana et al., 2010). Therefore, in our case, both suid and human infections are plausible.
A wide range of obligate and occasionally coprophilous fungi occurs throughout the sequence, in particular Delitschia sp., Sordaria t. and Cercophora t. In the absence of specific data to address herbivore composition such as lipid/bile analysis or DNA metabarcoding (D’Anjou et al., 2012; Giguet-Covex et al., 2014), the coastal location of our site allows us to restrict the number of possible species whose dung was affected by fungal growth. The most common even-toed ungulates documented in the Ligurian Holocene (Capreolus capreolus, Cervus elaphus, Rupicapra rupicapra, Capra ibex) all have preferences for elevated habitats (up to 2000 m a.s.l.) and only sporadically graze at lower altitudes. No hills exceeding 500 m a.s.l. occur in a range of 5 km from the site, suggesting only a limited contribution to the coprophilous assemblage from these species. Moreover, it is not certain that fallow deer (Dama dama), typically Mediterranean, was diffused in the region in prehistory (Minelli, 2002). Leporidae and Mustelidae are likely to be represented in our fimicolous record, but presumably only on a limited scale. As regards equid populations, horses are almost non-existent in the Italian zooarchaeological record prior to their reintroduction as domesticated species from eastern Europe during the Eneolithic (Wilkens, 2012). Omnivores such as wild boars should also be ruled out, given the rarity of fungi isolated from non-herbivore dung (Lundqvist, 1972).
In light of this, it is most likely that the only significant contribution to the coprophilous record results from bovine and ovi-caprine dung, a substrate very frequently affected by fungal growth (Doveri, 2011; Richardson, 2001). The alluvial plain may have been particularly suitable for domestic flocks, given the presence of handy watering places with still water and salt availability, a precious element for bovine and ovi-caprine diet (Tasić, 2000).
Human impact on the delta area during the Neolithic Age
The finding of charred cereal carpo-remains at the beginning of the phase BH1-2 and the date obtained by a caryopsis of Triticum dicoccum are the earliest evidence of human presence available to date for the urban area of Genoa.
The pollen curve of Human Impact Index shows a particularly marked frequency and height of the peaks since the initial part of BH1-2, when cereal cultivation seems also more evident in the carpological record.
As regards the curve of small micro-charred particles, thought to represent burning events on a regional scale, the trend correlates with the curve of larger particles, resulting from local fires. Although a clear differentiation between the two sources is difficult, this situation seems to point to a higher number of fires at the end of BH1-1, during the period of preparing and preceding cereal cultivation. In BH1-2, the situation seems to become more steady, with a relatively low incidence of micro-charcoal, followed by a new increase during BH3.
These dates, obtained from botanical finds, seem to witness to human presence in the Bisagno floodplain in the first half of the 5th Millennium BC, in agreement with the artefacts attributed to the Square-Mouthed Pottery Culture that were found on the ridges around the current city of Genoa (Del Lucchese, 2014; Fossati and Starnini, 2008).
Traces of Neolithic occupation first emerged in 1993, when a borehole in Piazza della Vittoria (S13) hit a fragment of carved wood from deciduous oak at a depth of 12.5 m (5.16 m b.s.l.). The find was dated between 4780 and 4460 cal. BC (GrN21525, 5770 ± 70 BP) and a relationship with a few small prehistoric potsherds recovered in correlated levels from a nearby borehole (S13 bis) dated between 5300 and 5030 cal. BC (GrA 5156 6200 ± 50 BP; Montanari et al., 1997), dug during the same construction work to build an underground car park, was put forward (Maggi, 1996a, 1996b).
A decade later, between 2007 and 2008, during the excavation of a ventilation pit for the tube station of Piazza Brignole, at a depth of 10 m below road level (3.65 m a.s.l.) and c. 500 m far from the core BH1, uncharred ash twigs showing clear evidence for shredding were found. Tree pruning is a traditional pastoral practice, which is used to obtain fodder. The remains were dated between 4780 and 4540 cal. BC (LTL3700A, 5806 ± 50 BP), allowing us to place the finding within the chronological horizon BH1-2 of Piazza della Vittoria (Arobba and Caramiello, 2014).
Conclusion
The radiocarbon dates place the investigated sequence between the end of the 7th Millennium and the first half of the 5th Millennium cal. BC, allowing us to record in detail the mid-Holocene palaeoenvironmental evolution of a deltaic area where materials from the whole drainage basin of the Bisagno were deposited.
An element characterizing the pollen and macrofossil diagram is the presence of Abies alba, the dominant arboreal taxon during BH1-1 and BH1-2, then decreasing since c. 4500 years cal. BC, in agreement with other papers on the Ligurian mountains and coastal belt (Bellini et al., 2009; Branch, 2004, 2013; Branch et al., 2014; Branch and Marini, 2013; Lowe et al., 1994; Montanari et al., 1997, 2014). This reduction on the first Apennine slopes and in the near-coast areas is interpreted as the effect of a cooler and more humid climate that would have favoured the expansion of beech. Aside from climate change, the simultaneous decline in silver fir seems to be due also to human activity, which in this area is indicated by an increase in agricultural and clearance practices for the management of woodland resources and above all for wood collection (Mercuri et al., 2012).
Moreover, the landscape is characterized by elements of the mesophilous wood, accompanied by hydrophilous arboreal species along the impluvia, while the Mediterranean component appears still poorly represented. The presence of wetlands is shown across the whole sequence by herbaceous coenoses dominated by freshwater hydro-hygrophilous taxa, sporadically coupled with elements typical of brackish environments (Ruppia, Bolboschoenus) pointing to occasional marine ingressions in back dunal zones.
Floristic changes caused over time by deep modifications in the water regime of the territory, affected by urbanization and management of the coastal environment, are clearly shown by the finding in the carpological record of BH1-1 and BH1-3 of two hygrophilous species currently unknown in Liguria (Hydrocotyle vulgaris and Stratiotes aloides).
Human presence is made evident mainly by the finding of cereal carpo-remains dated to 5470–5220 cal. BC, to date the earliest evidence of agricultural activities in the area of Genoa. Triticum dicoccum and Hordeum vulgare var. tetrastichum are the most commonly found cereal taxa in Western Liguria since the 6th Millennium BC (Arobba et al., 2017b; Nisbet, 2008). The finding of a spikelet base probably from Triticum new glume wheat is of particular interest and represents the first record in Liguria. This is a tetraploid wheat, nowadays extinct and similar to Triticum timopheevi, appearing in the Early Neolithic of Central Europe and France with the Linear Pottery Culture (Linearbandkeramik-Kultur, LBK, 5500–4500 cal. BC) and then decreasing until its disappearance in the early 1st Millennium BC (Rottoli and Castiglioni, 2009; Toulemond et al., 2015).
Additional elements pointing to anthropogenic disturbance around the borehole area result from the finding of herbaceous synanthropic species and from the analysis of micro-charcoal particles, which, even though they can derive from the whole catchment area through fluvial transport, may also be the product of local fires.
A further aspect concerns the NPPs, some of which can be related to lagoonal environments and human presence in the area through possible pastoral practices.
For the area under investigation, it is rather uncommon that about two millennia of palaeoenvironmental history are preserved in such a long sedimentary sequence, for which a particularly detailed analysis was possible. However, at the moment, the sequence cannot be compared with the ones obtained from other boreholes, where the same time span is comprised within thinner deposits. In the sedimentary sequence analysed, no major changes in environment connected to large-scale climatic events can be observed. The fluctuations responsible for the formation of various habitats comply with the usual condition of a deltaic area, naturally subject to continuous modifications, related to seasonal variations in the flow rate of the watercourse. This is also highlighted by the stratigraphy, showing a remarkable depositional homogeneity interrupted only by a few episodes, mainly characterized by the formation of peat deposits related to the presence of swampy oxbow lakes.
Supplemental Material
Supplementary_material – Supplemental material for New evidence on the earliest human presence in the urban area of Genoa (Liguria, Italy): A multi-proxy study of a mid-Holocene deposit at the mouth of the Bisagno river
Supplemental material, Supplementary_material for New evidence on the earliest human presence in the urban area of Genoa (Liguria, Italy): A multi-proxy study of a mid-Holocene deposit at the mouth of the Bisagno river by Daniele Arobba, Rosanna Caramiello, Marco Firpo, Luca Mercalli, Lionello F Morandi and Stefano Rossi in The Holocene
Footnotes
Acknowledgements
Particle size analysis with the aid of the Sedigraph was carried out by Alberto Demergasso of the Laboratory of Sedimentology and Marine Geology of the University of Genoa. We would like to thank two anonymous reviewers for their constructive comments.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.
Supplementary material
List of new non-pollen palynomorph (NPP) types, their possible biological identification, equivalencies with previously published types and references.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.