
Abstract
The southern Benguela upwelling system near the St. Helena Bay has been proposed to be affected by various factors, while few investigations about the late-Holocene oceanic conditions has been carried out in this area. To determine the oceanic variability and its forcing mechanisms in the southern Benguela region during the late-Holocene, we examined organic-walled dinoflagellate cyst (dinocyst) records from two marine sediment cores located in the southernmost and central Benguela upwelling system. We compare our results with other proxies including alkenone-derived SSTs, grain size, and coccolithophore assemblages from the same samples. The results indicate a distinctive behavior between the southernmost Benguela system and the central Benguela area. We infer that the oceanic conditions in these two regions are primarily governed by an interplay of wind-induced upwelling, fluvial discharge, and advection of cold sub-Antarctic waters, which is consistent with the current understanding of the paleoclimate conditions in this area. However, the findings also suggest that the southernmost Benguela system also receives additional effects of warm and saline waters via the Agulhas leakage, which has a clear influence on the oceanic conditions in this area.
Keywords
Introduction
The Benguela upwelling system along the west coast of southern Africa is one of the four major eastern boundary upwelling regions of the world driven by the southeast trade winds (Shannon, 1985). The strongest upwelling takes place near Lüderitz (27°S) which naturally divides the Benguela into two subsystems: the northern and central Benguela region with seasonal to perennial upwelling (17–25°S) and the southern Benguela region with seasonal upwelling (25–35°S). Modern variability relating to upwelling conditions and wind stress differs between the two subsystems (Farmer et al., 2005; Petrick et al., 2015a; Tim et al., 2015; Zhao et al., 2017), while a clear picture of upwelling variations during the late-Holocene is not yet established. The southern Benguela upwelling system (Figure 1) is driven by the interaction of several factors, that is, coastline direction, southeast trade winds, southern westerlies, and the influence of the Agulhas Current system. The Agulhas Current is one of the largest currents of the Southern Hemisphere, which flows southward along the east coast of southern Africa and then flows back (retroflection) into the Indian Ocean when it passes the southern tip of South Africa. Warm and salty waters are leaked through eddies, rings, and filaments into the Atlantic Ocean from the retroflection area, which is called the Agulhas leakage (De Ruijter et al., 1982; Lutjeharms et al., 1991). The Agulhas leakage crossing the South Atlantic Ocean leaves behind a trail of mixed and stirred water, whose frictional energy transfer heightens the background energy field of the ocean basin (Duncombe Rae, 1991). It plays a crucial role in the inter-ocean heat and salt exchange affecting the long-term Atlantic meridional overturning circulation and global climate (Biastoch et al., 2008; Gordon et al., 1992; Weijer et al., 2002). Thus, the variability of the leakage is of particular interest on all timescales. For the Cape Basin, numerous studies have been carried out on millennial, orbital, and glacial–interglacial timescales (Caley et al., 2011, 2012, 2014; Dickson et al., 2010; Dyez et al., 2014; Franzese et al., 2006; Marino et al., 2013; Martínez-Méndez et al., 2010; Peeters et al., 2004; Petrick et al., 2015b; Rau et al., 2002).
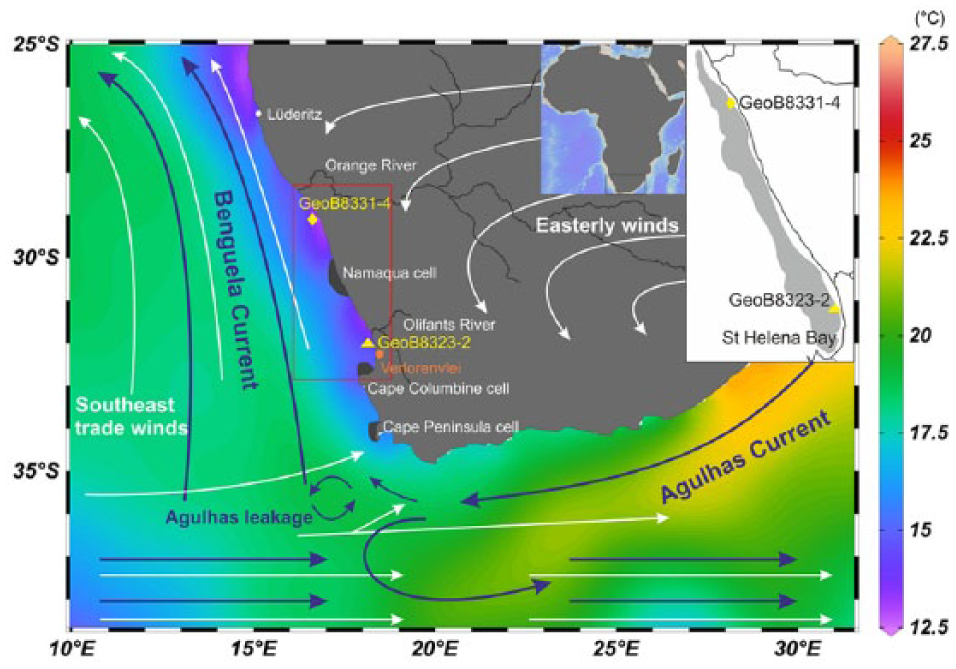
Map of the research area showing the locations of gravity cores GeoB8323-2 (yellow triangle) and GeoB8331-4 (yellow diamond) retrieved from the Namaqualand mudbelt (gray shaded area in the upper right corner) off the west coast of South Africa, the Orange and the Olifants Rivers as well as the terrestrial record Verlorenvlei (orange circle) mentioned in the study. The arrows indicate the major atmospheric (white arrows) and oceanic (blue arrows, Agulhas Current, Benguela Current) circulation systems over southern Africa with the mean annual sea surface temperatures in colors (Ocean Data View, U.S.NODC World Ocean Atlas 2013). The locations of three primary upwelling cells in the southern Benguela upwelling system are denoted with black shading: Namaqua cell, Cape Columbine cell, and Cape Peninsula cell.
It has been demonstrated that the Agulhas leakage, besides its impact on the ocean circulation and global climate change, has strong local effects on the Benguela system. The Alkenone-derived sea surface temperatures (SST) together with grain size data and element ratio in the southernmost Benguela region suggests an influence of the Agulhas leakage during the late-Holocene (Granger et al., 2018; Hahn et al., 2016). Coccolithophore data from the same core were obtained to reconstruct surface water conditions such as water temperature and nutrient availability (Weiser et al., 2016). The coccolithophore assemblage showed five different phases of characteristic upwelling conditions driven by the interaction of southeast trade winds and the latitudinal shift of the southern westerlies, although no signal based on the coccolithophore data can indicate the effects of the Agulhas leakage (Weiser et al., 2016). However, the organic-walled dinoflagellate cyst record from the vicinity of the Orange River demonstrated that warm waters of Agulhas leakage unlikely reached as far north as the central Benguela region (Zhao et al., 2017). Thus, until now, modulations of the Agulhas leakage during the Holocene and its possible influence on waters of the inner shelf (Namaqualand mudbelt) remain uncertain. Therefore, a detailed paleoceanographic reconstruction for the Holocene is an important contribution to the study area.
A widely used method of depicting upwelling history is through paleoceanographic reconstruction based on organic-walled dinoflagellate cyst (‘dinocyst’ hereafter) analysis. Dinocysts are composed of resistant organic material that is generally well preserved in most marine sediments (e.g. Dale, 1996). Dinocyst species have different environmental preferences and their distribution in surface oceanic sediments has been shown to correlate well with sea surface conditions such as productivity, temperature, salinity, and sea-ice cover duration (e.g. Dale, 1996; Dale et al., 2002; De Vernal et al., 1997; Marret and Zonneveld, 2003; Zonneveld et al., 2013). Therefore, the fossil dinocyst association has become a valuable tool to reconstruct past marine environments, especially in neritic (shallow marine environment on the continental shelf) highly productive environments (De Hardy et al., 2018; De Vernal et al., 1997; Pospelova et al., 2006, 2015; Verleye and Louwye, 2010). No dinocyst studies so far have covered the late-Holocene oceanographic changes over the southern Namaqualand mudbelt. To address this lacuna, we investigated a centennial scale organic-walled dinoflagellate cyst record of marine sediment core GeoB8323-2 located in the southernmost part of the Benguela system (Figure 1). We compared our data with dinocyst records of marine sediment core GeoB8331-4 located in the central Benguela region (Figure 1) and other proxies to address the following questions: (1) How did the oceanic conditions vary during the late-Holocene in the southern and central Benguela upwelling region? (2) What factors drove this oceanic variability? (3) What is the effect of Agulhas leakage on the Benguela upwelling system?
Southern Benguela upwelling system
The characteristics of the southern Benguela upwelling system have been described in detail by Lutjeharms and van Ballegooyen (1984), Nelson and Hutchings (1983), and Shannon (1985). Wind-driven seasonal upwelling occurs in the southern Benguela region with three primary upwelling cells: the Namaqua cell (NC, 30°S), the Cape Columbine cell (CC, 33°S), and the Cape Peninsula cell (CP, 34°S; Figure 1). The southern study site is located in the St Helena Bay region near to the Cape Columbine cell. Seasonal upwelling intensity is related to the strength and direction of the southeast trade winds, which are controlled by the position of the South Atlantic High Pressure System (Nelson and Hutchings, 1983). Thus, the maximum upwelling occurs when the southeast trade winds are the strongest in this area during austral spring and summer, while upwelling minima are associated with prevailing westerly winds during austral winter (Shannon, 1985; Shannon and Nelson, 1996).
Along the southernmost part of the Benguela upwelling system, water of the Agulhas Current enters the South Atlantic Ocean as the ‘Agulhas leakage’ in the form of eddies or filaments, which are further transported in a north-westerly direction through the South Atlantic Ocean (Figure 1). The leakage is determined mainly by the latitudinal position of the southern westerlies. A widespread belief is that the southward shift of the southern westerlies would enable more inflow of the Agulhas Current water to the South Atlantic Ocean (Biastoch et al., 2009; De Ruijter, 1982). However, recent modeling studies of Durgadoo et al. (2013) emphasized that increased leakage corresponds with increased intensity of the southern westerlies. Despite apparent contradictions, these two hypotheses do not conflict with one another, since the southern westerlies show a latitudinal contraction as well as intensification within the core during austral summer, while during austral winter they extend northward but with reduced wind intensity in the core zone (Lamy et al., 2010). Therefore, during austral summer, the southward contraction and intensification of the southern westerlies would favor more Agulhas leakage, coinciding with increased upwelling in the southern Benguela region.
Materials and methods
Age-depth model
The gravity core GeoB8323-2 (32°01.70’S, 18°12.21’E, 285 cm) was recovered in 2003 during the M57/1 Meteor cruise (Schneider et al., 2003) from the southern Namaqualand mudbelt near St Helena Bay off western South Africa (Figure 1). The core consists of homogeneous, dark greenish-gray sandy mud (Schneider et al., 2003). The age model of GeoB8323-2 has been published by Herbert and Compton (2007) and was updated by Hahn et al. (2016). Because of a hiatus somewhere between 252 and 234 cm and the absence of material (such as gastropods) between 252 and 227 cm that could be radiocarbon dated, the age model for the lower part is uncertain. Therefore, only the undisturbed upper part of the core (above 227 cm) was analyzed. An age-depth model was established based on four AMS 14 C dates above 227 cm (39.5, 111, 124, and 227 cm; Table 1) based on a Bayesian model using Bacon (Blaauw and Christen, 2011).
Radiocarbon ages of marine organisms and bulk TOC for core GeoB8323-2 recalibrated after Herbert and Compton (2007) and Hahn et al. (2016). Ages were calibrated using OXCAL software (Bronk Ramsey, 2001) and marine13 radiocarbon calibration model (Reimer et al., 2013). An extra reservoir age (ΔR) of 146 ± 85 14 C years (Dewar et al., 2012) was applied.

Dinoflagellate samples preparation and identification
A total of 35 samples were treated with cold ca. 12% HCl and ca. 40% HF. All particles smaller than 10–15 µm were removed by ultrasonical sieving. Samples were stored in water, mounted in glycerol, and identified under a light microscope (magnification 400× and 1000×). For each sample, at least 400 dinocysts were counted and percentages of each taxon were calculated based on the total number of dinocysts. Accumulation rates of dinocysts (cysts/cm2/yr) were calculated by multiplying the dinocyst concentrations (cysts/cm3) with the sedimentation rate (cm/yr) for each sample. Water loss might have affected the absolute volume measurements, which introduces a systematic error in the accumulation rates but without consequences for the interpretation of relative changes. All counts of dinocysts are available and stored in the Pangaea database (https://doi.pangaea.de/10.1594/PANGAEA.886221).
The identification for the dinocyst taxa follows the online modern dinoflagellate cyst determination key (Zonneveld and Pospelova, 2015 and references therein). Identification of the cysts of Polykrikos kofoidii and Polykrikos schwartzii was made according to Matsuoka et al. (2009). All Brigantedinium species were grouped in Brigantedinium spp., because often the orientation of the cysts on the microscope slides precludes identification of the species within the genus. Also, other cysts were not always identifiable at species level because of unsuitable orientation, adherence of debris, or the inability to spot typical taxonomic features. These were grouped in ‘spp’. for instance, Spiniferites spp. Cysts of Echinidinium and Islandinium that could not be identified free of doubt have been counted as spiny brown cysts.
Dinocyst identified in this study are ecologically grouped into heterotrophic taxa or autotrophic taxa according to their metabolism mechanism (Table 2, Supplementary Figures 1–3). The ratio Peridiniaceae /Gonyaulacaceae (P/G) was first documented by Lewis et al. (1990) to illustrate changes in the upwelling intensity offshore Peru, which has been broadly used and adapted with the inclusion of other heterotrophic taxa like Echinidinium (Esper et al., 2004) and Polykrikos (Marret, 1994). Considering that heterotrophic and autotrophic groups are more comprehensive and better linked to productivity than the Peridiniaceae and Gonyaulacaceae groups (e.g. Bringué et al., 2014; Verhoeven and Louwye, 2013), the proportion of autotrophic taxa of the total dinocyst sum is calculated as A/(A + H).
List of dinoflagellate cyst taxa identified in core GeoB8323-2. Thecal equivalents are referred to in Head (1996), Zonneveld et al. (1997), Head et al. (2001), Matsuoka et al. (2009), and Zonneveld et al. (2013).

It has been found that dinocysts are prone to species-specific aerobic degradation (McCarthy et al., 2004; Prebble et al., 2013; Versteegh and Zonneveld, 2002; Zonneveld et al.,1997, 2007, 2008). Because of the large differences in degradation of sensitive cysts (e.g. Brigantedinium spp., Echinidinium spp., Selenopemphix quanta) and resistant cysts (e.g. Impagidinium spp., Nematosphaeropsis labyrinthus, Polysphaeridium zoharyi), it is necessary to separate degradation from past bioproductivity signals. To investigate the degree of degradation of sensitive cysts (S-cysts) in our samples, the degradation of S-cysts expressed by their ‘kt’ values has been calculated assuming a first-order decay process:

where Xi = initial cyst concentration (cysts/cm2/yr) and Xf = final cyst concentration (cysts/cm2/yr) (Versteegh and Zonneveld, 2002; Zonneveld et al., 2007).
Generally, a linear relationship can be observed between the dinocyst accumulation rates (AR) of different cyst species groups in sediments that have not been prone to aerobic degradation, which is AR group A = α × AR group B (Versteegh and Zonneveld, 2002; Zonneveld et al., 2007). Zonneveld et al. (2007) determined a constant, regionally determined value ‘α’ for species group A (resistant species) and species group B (sensitive species) and then successively reconstructed the initial accumulation rates of species group B. Then the X in Eq. (1) was replaced by AR and the ‘kt’ values can be calculated, which turned out to be strongly related to the bottom water oxygen concentrations.
In addition, we address the modern dinocyst distribution pattern using a dinocyst database based on 111 marine surface sediments in the Benguela upwelling region from Holzwarth et al. (2007), Zhao et al. (2017), and Zonneveld et al. (2001a).
Ordination methods
An ordination analysis based on dinocyst data of core GeoB8323-2 has been performed with the CANOCO software (Canonical Community Ordination: version 5; Ter Braak and Šmilauer, 2012). A Detrended Correspondence Analysis (DCA) was carried out first to test the distribution of the dataset. The longest gradient of DCA analysis was found to be shorter than 3 standard deviations (0.99), indicating that a linear model is more suitable. Thus, a principal component analysis (PCA) was performed on the percentages of all dinocyst species and on the 35 samples. The percentage data used for analysis were log-transformed to ensure that each dinocyst taxon contributes equally to the PCA. In addition, constrained Incremental Sum of Squares Cluster Analysis (CONISS, TILIA 2.0.41; Grimm, 2011) was also used to define dinocyst assemblage zonation.
Results
Chronology
Different calibrated ages were reported by Hahn et al. (2016) and Granger et al. (2018), although with minor differences. Recalibration applying an extra reservoir age of 146 ± 85 14C years (Dewar et al., 2012) confirmed those of Hahn et al. (2016; Table 1). The results of AMS 14C dates based on marine organisms are presented in Table 1. A Bayesian model using Bacon (Blaauw and Christen, 2011; Figure 2a) was used in which linear accumulation has been assumed between 38 vertical sections of 5 cm. Prior sedimentation time was set to 10 yr cm-1 with a normal distribution (Shape parameter 1.5). For the memory parameters the default setting was used. Linear sedimentation rates were calculated based on weighted mean calibrated ages with the depth interval divided by the age interval between each sample. The age-depth model show a basal age of 268 BCE. The alkenone-derived SSTs, grain size (Granger et al., 2018), and coccolithophore data (Weiser et al., 2016) were also plotted on the new age model.
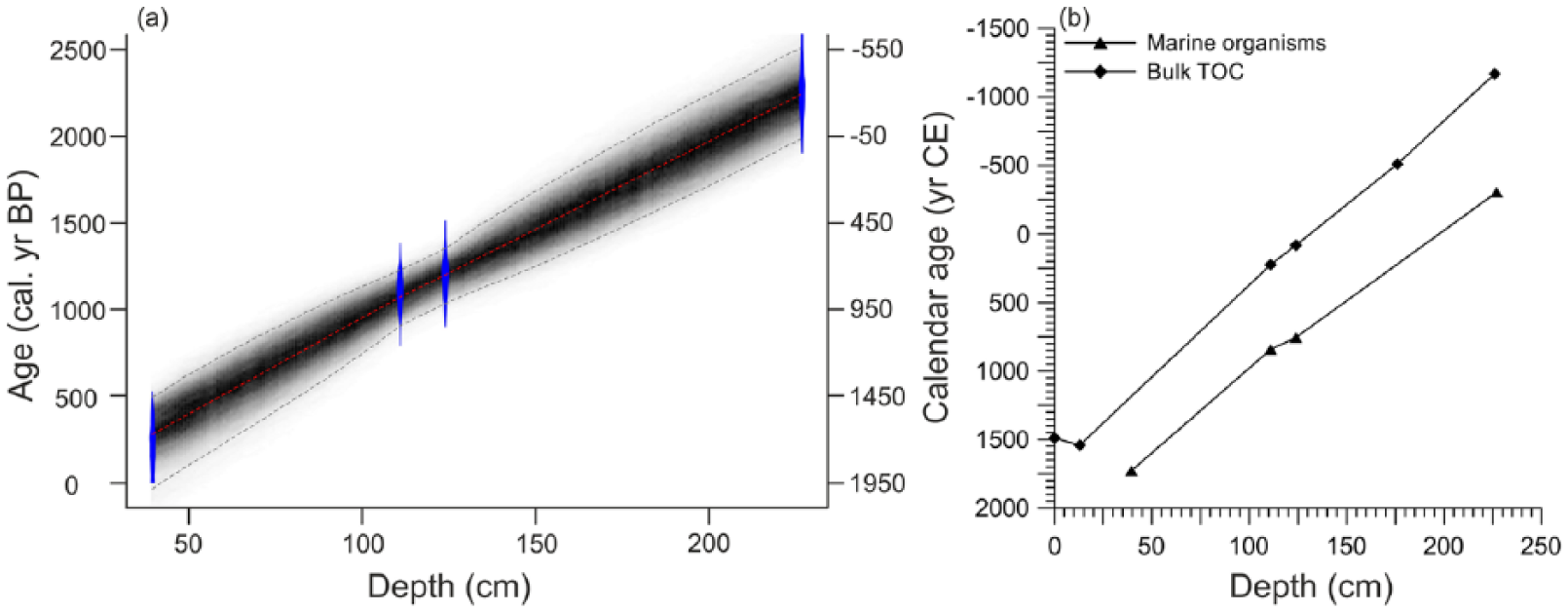
(a) Bayesian age-depth model of core GeoB8323-2: the blue areas represent the 95% probability distributions of the calibrated 14C ages; the thin red line shows the weighted mean ages and the blackish-gray area shows the 95% age-depth relation as modeled by the R software package Bacon 2.2 (Blaauw and Christen, 2011). (b) Comparison of calibrated ages based on marine organisms (triangles) and bulk organic carbon (diamonds). Ages were calibrated using marine13 radiocarbon calibration model (Reimer et al., 2013). An extra reservoir age (ΔR) of 146 ± 85 14C years (Dewar et al., 2012) was applied.
Because of the limited number of radiocarbon ages, there is about a 500 year range for the 95% probability in the age-depth model, which should be regarded with caution. The calibrated ages based on bulk organic carbon (Figure 2b) are generally ca. 500 years older than the calibrated ages in marine organisms from the same horizons, suggesting a relatively constant sediment accumulation rate through the late-Holocene (Hahn et al., 2016). However, the age-depth model between ca.450 CE and 300 BCE is not constrained by data points.
Dinocyst results in core GeoB8323-2
Dinocysts were abundant in core GeoB8323-2 and a total of 36 cyst taxa (including types and morphotypes) were identified. No reworked dinocyst from older material was observed in the core samples. All dinocyst taxa identified in this study are listed in Table 2. Records of selected dinocyst taxa are shown in Figure 3. Dinocyst concentrations varied from ~19,000 to ~113,000 cysts/cm3 (average of ~66,100 cysts/cm3) and accumulation rates varied from ~1870 to ~11,100 cysts/cm2/yr (average of ~6900 cysts/cm2/yr). Both showed a declining trend prior to 1600 CE and increased again to high values after 1600 CE (Figure 5f). The most dominant dinocyst taxa consistently throughout the record were Brigantedinium spp. (max of ~49%), Dubridinium spp. (max of ~30%), Selenopemphix quanta (max of ~26%), cyst of Protoperidinium monospinum (max of ~16%), cyst of Protoperidinium americanum (max of ~11%), and Echinidinium aculeatum (max of ~9%; Figure 3). Dinocysts produced by Gymnodiniales (cysts of P. kofoidii and P. schwartzii) contributed up to 22% of the assemblage at the beginning of the record but quickly decreased to low values after ca. 0 CE. Percentages of cyst of P. schwartzii reached a minimum (~2%) at ca. 1600 CE and then increased gradually up to ~7% at the end of the record (Figure 3). The autotrophic taxa included Spiniferites mirabilis (max of ~14%) and Spiniferites ramosus (max of ~13%), both with high values between ca. 400 and 800 CE and Spiniferites hyperacanthus (max of ~7%) with high values between ca. 110 BCE and 800 CE. The other autotrophic taxa such as Operculodinium centrocarpum (max of ~11%), P. zoharyi (max of ~7%), and Operculodinium israelianum (max of ~6%) all showed similar trends with high values between 25 BCE and 250 CE (Figure 3). These three taxa (particularly O. centrocarpum and P. zoharyi) started to increase again after ca. 1800 CE.
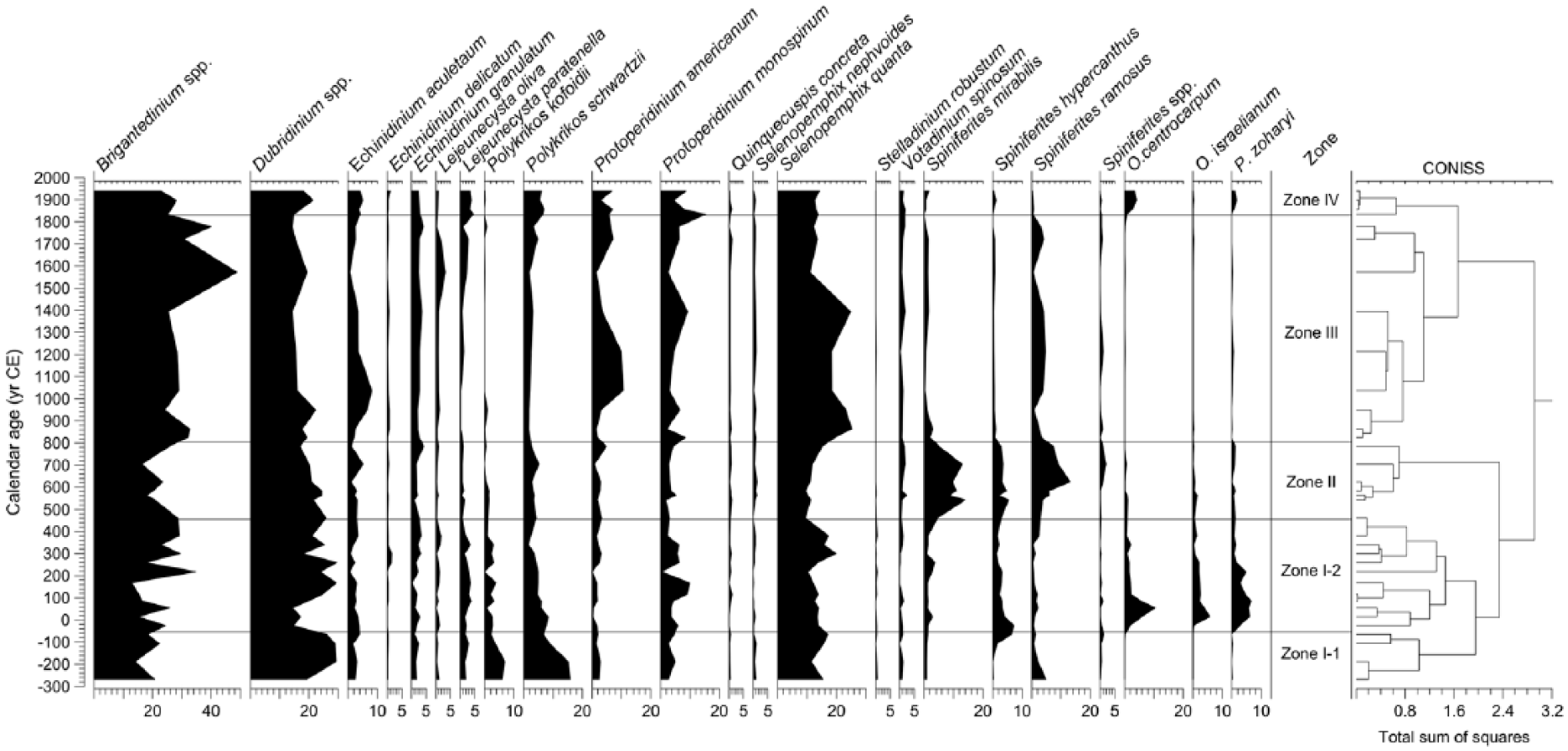
Relative abundance (%) of selected dinoflagellate cyst taxa in gravity core GeoB8323-2 from the southern Namaqualand mudbelt. Dinocyst assemblage zones were assigned using CONISS (Grimm, 2011).
Therefore, dinocysts produced by heterotrophic dinoflagellates dominated the assemblage with the representation between 70% and 98% (average of 88%) throughout the whole record, while dinocysts produced by autotrophic dinoflagellates were only abundant before ca. 800 CE, representing up to 30% (average of ~12%) of the assemblage (Figure 5d). The kt values calculated for the whole record ranged between 0 and 3 with relatively high values between ca. 25 BCE and 580 CE (max of 3, average of 1) and after 1900 CE (max of 0.8, average of 0.7; Figure 5e).
The PCA resulted in the first and second axes accounting for 33% and 23% of the total variance, respectively (Table 3). Figure 4 shows the ordination of the most common cyst taxa based on the first two axes (PCA1 and PCA2). Four groups of cyst taxa were distinguished based on taxa scores along the axes of CA results (Figure 4).
Total variances explained by different axes resulted from principal component analysis (PCA).
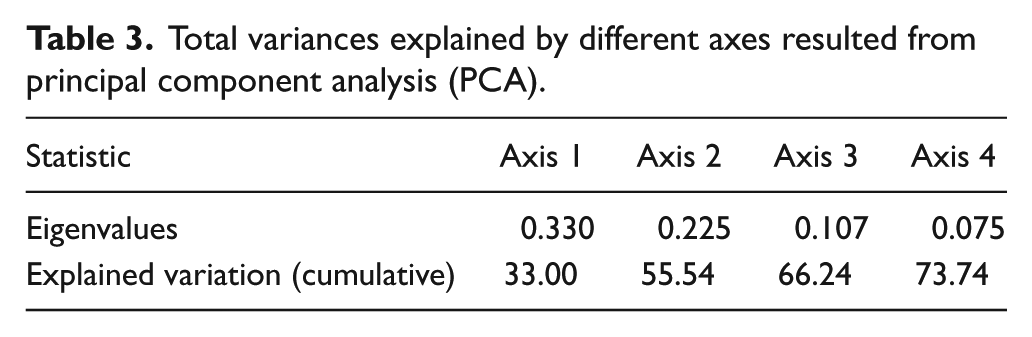

Results of the principal correspondence analysis (PCA) illustrating the ordination of dinoflagellate cyst taxa from gravity core GeoB8323-2 with colored symbols referring to the groups.
(1) Agulhas group: O. centrocarpum, O. israelianum, and P. zoharyi
The Agulhas group of core GeoB8323-2 (Figure 5a) includes O. centrocarpum, O. israelianum, and P. zoharyi which all have higher values at the same period (between ca. 30 BCE and 250 CE). Although O. centrocarpum is a cosmopolitan species, it is also documented to increase in abundance in unstable waters (Dale, 1996; Wall et al., 1977). Davey (1971), Davey and Rogers (1975), and Wall et al. (1977) found that high concentrations of O. centrocarpum in southern Benguela shelf sediments are influenced by warm waters influenced by Agulhas leakage. O. israelianum can be observed in high salinity surface water conditions (Zonneveld et al., 2013) as well as P. zoharyi, which often dominates in areas with elevated surface water salinities (Marret and Zonneveld, 2003). Currently, the warm and saline Indian Ocean waters derived from the Agulhas Current are the main source of warm and salty water masses in the southeast Atlantic Ocean. Although eddies of Agulhas Current do not propagate onto the continental shelf (Pilo et al., 2015) from where core GeoB8323-2 has been retrieved, strong exchange and interaction of warm and salty Agulhas water with South Atlantic water can still affect the properties of the southern Benguela region, even it is not under the direct pathway of the Agulhas leakage (Duncombe Rae, 1991; Granger et al., 2018; Lutjeharms et al., 1991). Thus, we can use this group as an indication of changes in the upper water column over the northern Namaqualand mudbelt associated with increased Agulhas leakage.
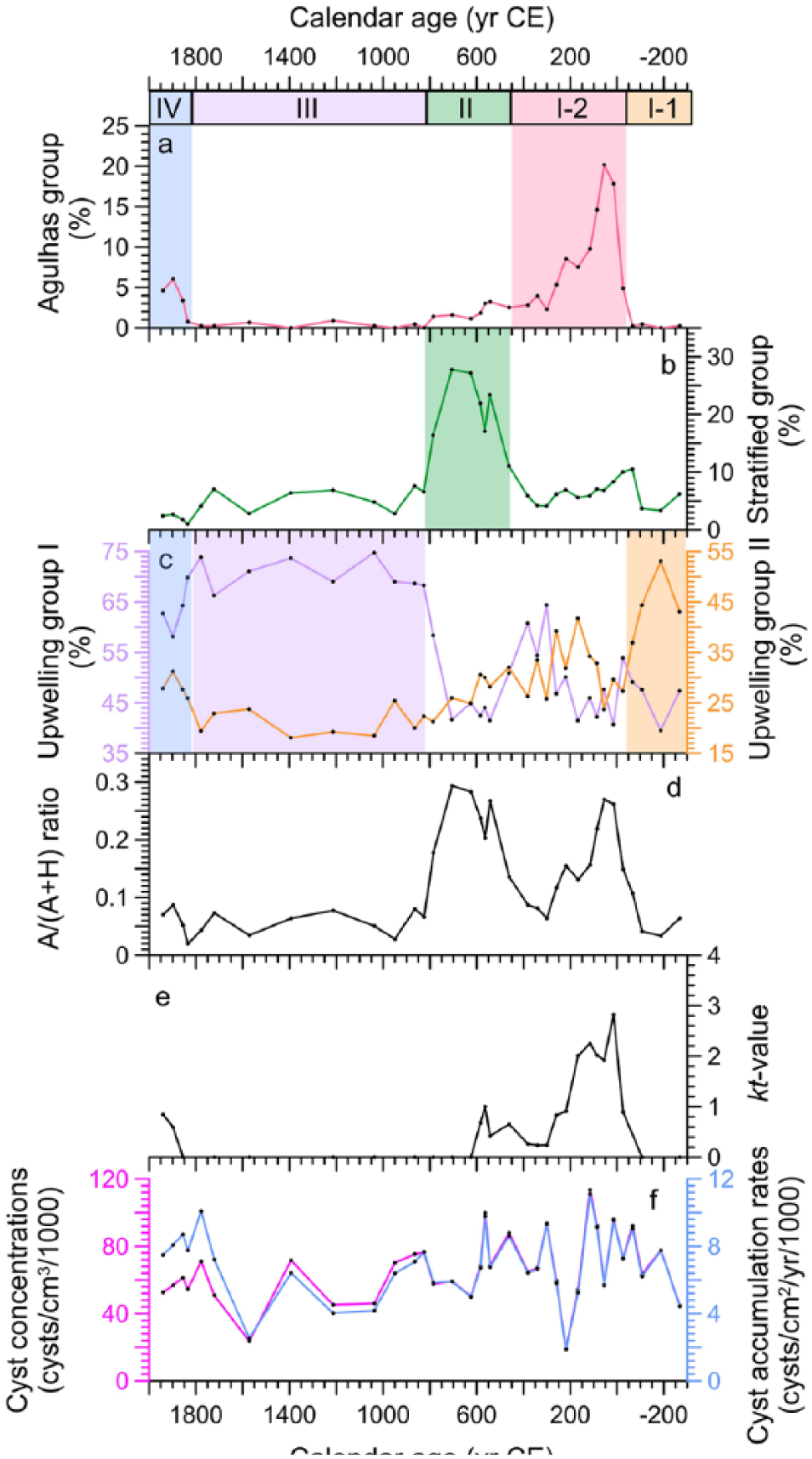
Dinoflagellate cyst percentages of (a) Agulhas group, (b) Stratified group, (c) Upwelling groups I and II, (d) the proportion of autotrophic taxa of the total dinocyst sum (A/(A + H) ratio), (e) kt value of sensitive cysts, and (f) cyst concentrations and accumulation rates for gravity core GeoB8323-2. Agulhas group (O. centrocarpum, O. israelianum, and P. zoharyi); Stratified group (S. mirabilis, S. hyperacanthus, S. ramosus, and Spiniferites spp); Upwelling group I (Brigantedinium spp., E. aculeatum, E. delicatum, E. granulatum, L. oliva, cyst of P. americanum, S. quanta, cyst of P. monospinum, Votadinium spinosum); Upwelling group II (Dubridinium spp., cyst of P. kofoidii, cyst of P. schwartzii and L. paratenella).
(2) Stratified group: S. mirabilis, S. hyperacanthus, S. ramosus, and Spiniferites spp
The Stratified group (Figure 5b) is Spiniferites group including S. mirabilis, S. hyperacanthus, S. ramosus, and Spiniferites spp. Pospelova et al. (2018) based on eight sediment trap time series in different geographic regions from the Pacific Ocean, Indian Ocean, and Atlantic Ocean demonstrated that different Spiniferites species react in a similar way to local environmental triggers at each site and the highest values of Spiniferites indicate minimal turbidity or stratification and availability of nutrients. In core GeoB8331-4, Spiniferites spp. and S. ramosus together with O. centrocarpum were inferred to indicate warm and stratified waters, while in core GeoB8323-2 Spiniferites taxa are distinguished in one separate group. This suggests that the Stratified group indicates different warm conditions as those indicated by Agulhas group, which is warm and stratified waters with stable conditions likely because of upwelling relaxation.
3. Upwelling group with two subgroups: Upwelling group I (Brigantedinium spp., E. aculeatum, E. delicatum, E. granulatum, L. oliva, cyst of P. americanum, S. quanta, cyst of P. monospinum, and Votadinium spinosum), Upwelling group II (Dubridinium spp., L. paratenella, cysts of P. kofoidii, and P. schwartzii)
The taxa in the Upwelling group (Figure 5c) are all produced by heterotrophic dinoflagellates, which most likely indicate cold and nutrient-rich waters with active upwelling. However, the trend in percentages of Dubridinium spp., Polykrikos, and L. paratenella cysts is different from those of the other taxa in the Upwelling group (Figures 3 and 5c) and the ordination of these four taxa differs from other heterotrophic taxa (Figure 4). Therefore, we distinguish two subgroups: Upwelling group I including Brigantedinium spp., E. aculeatum, E. delicatum, E. granulatum, L. oliva, cyst of P. americanum, S. quanta, cyst of P. monospinum, and Votadinium spinosum and Upwelling group II with Dubridinium spp., Polykrikos cysts and L. paratenella. The change in dominant dinoflagellate community suggests differences in upwelling conditions between two different periods, one before 50 BCE and the other after 800 CE.
Four zones can be distinguished based on the CONISS calculation (Figures 3).
Discussion
Investigation of modern dinocyst distribution along the Namaqualand mudbelt and fossil dinocyst records in core GeoB8331-4 indicated that neither species-selective preservation nor lateral transport significantly affected the dinocyst assemblage in the central Benguela (Holzwarth et al., 2007; Zhao et al., 2017; Zonneveld et al., 2001a). In core GeoB8323-2 the protoperidiniaceans (composed mainly of Brigantedinium spp., Dubridinium spp., S. quanta, and cysts of P. kofoidii and P. schwartzii) being sensitive to post-depositional degradation (Gray et al., 2017; Zonneveld et al., 1997, 2007, 2001b, 2008) were well preserved and represented up to 98% of the dinocyst assemblage. The kt values associated with dinocyst preservation never exceeded 3 (Figure 5e), suggesting no alteration of the dinocyst assemblage by selective degradation of cysts. Dinocyst assemblages thus can be used as a reliable tool to reconstruct oceanographic conditions in this region.
Late-Holocene oceanographical reconstruction
Active upwelling and Agulhas leakage (300 BCE–450 CE, Zone I-1 and Zone I-2)
High representation of the Upwelling group II at the beginning of the record (Figures 4 and 5c) with the highest values prior to 0 CE suggest active upwelling conditions. The relatively high dinocyst concentrations and accumulation rates during this period also indicate high productivity (Figure 5f). However, the dominance of the Upwelling group II was interrupted by high representation of the Agulhas group between 30 BCE and 300 CE (Figures 4 and 5a). This coincides with high percentages of coccolithophore Gephyrocapsa ericsonii (Figure 6h; Weiser et al., 2016). However, these results are not consistent with the decline of alkenone SSTs (Figure 6e; Granger et al., 2018) and low Gephyrocapsa-based SSTs (Figure 6g; Weiser et al., 2016) from 70 BCE to 700 CE. The iron concentrations from the Chilean continental margin at 41°S (Lamy et al., 2001) provides a record of the latitudinal position of the southern westerlies. On one hand, the low iron concentrations suggests a southward shift of the southern westerlies from ca. 50 BCE to 300 CE reaching its southernmost position of the past 2200 years (Figure 6j). On the other hand, Lamy et al. (2010) found that northward expansion and weakening in the core zone (50–55°S) of the southern westerlies would also result in wetter conditions and lower iron concentrations in the marine sediments off central Chile (33–40°S) and vice versa. Thus, combining the iron concentrations in Lamy et al. (2001) and further interpretation of Lamy et al. (2010), an alternative interpretation could be a contraction of the southern westerlies but intensification of the wind stress in the core zone during this period. The record of terrestrial input by Granger et al. (2018) also indicates a weak influence of rain-bearing westerly winds during this time (Figure 6f). The contraction of the southern westerlies at that time would allow for the influence of strong southeast trade winds to promote upwelling in the southern Benguela region. This would cause a decline of SSTs, which is observed in both cores (GeoB8323-2 since 230 BCE and GeoB8331-4 since 30 BCE, Figure 6a and e), suggesting that the SST variations are dominated by upwelling during this period (Granger et al., 2018). However, low SSTs do not explain the high percentages for the Agulhas group dinocysts (Figure 6c) and G.ericsonii (Figure 6h).

From top to bottom: records for gravity core GeoB8331-4 (a) alkenone-derived sea surface temperature (SST, °C, blue; Leduc et al., 2010) and (b) dinoflagellate cyst percentages of cold and nutrient-rich water indicator (%, blue; Zhao et al., 2017); dinocyst records for gravity core GeoB8323-2, dinoflagellate cyst percentages of (c) the Agulhas group (%, pink) and (d) the Stratified group (%, pink; this study); other records for gravity core GeoB8323-2 (e) alkenone-derived SST (°C, purple) and (f) grain size variation (purple; Granger et al., 2018), (g) Gephyrocapsa-based SST (°C, green) and (h) coccolithophore percentages (%, green) of Gephyrocapsa ericsonii (Weiser et al., 2016); (i) dilute-water diatom (orange, %) and brackish-water diatom (magenta, %) from Verlorenvlei core VV09 (Stager et al., 2012); (j) iron concentrations (Fe intensity, cps) from the Chilean continental margin at 41°S (Lamy et al., 2001); (k) calibrated carbon ages for gravity core GeoB8323-2 (this study).
The seemingly contrasting results for the period between 300 BCE and 450 CE could be the expression of differences between seasons, in which upwelling intensified during austral spring and summer and Agulhas inflow increased during austral summer. The intensified upwelling with high nutrient conditions would have favored the growth of heterotrophic dinoflagellates, which indeed are well preserved (Upwelling group II until 30 BCE in GeoB8323-2 and indicator of cold and nutrient-rich water in GeoB8331-4, Figures 4, 5c and 6b). Despite intensified upwelling, the Agulhas leakage signal (Agulhas group) and warm water indicator G. ericsonii coccoliths are abundant in core GeoB8323-2 after 30 BCE, suggesting an influence of warm waters to the southern Benguela region. Currently, the warm and saline Indian Ocean waters derived from the Agulhas Current are the main source of warm water masses to the southern Benguela region. Therefore, the most likely source for the warm water contribution is increased Agulhas leakage. This can be explained by a contraction of the southern westerlies together with intensified wind stress in the core zone, which would enable higher inflow of the warm Agulhas water (Durgadoo et al., 2013). Although it has been demonstrated that the Agulhas waters are unlikely to flow as far north as site GeoB8331-4 (Zhao et al., 2017), this mechanism may well have influenced the properties of the southern site GeoB8323-2 near the Cape Columbine.
Suppressed upwelling in the southern Benguela region (450–800 CE, Zone II)
The higher representation of the Stratified group (Figures 4 and 6d) and higher alkenone SSTs (Figure 6e) in the southern core GeoB8323-2 suggest warm and stratified waters, which is probably because of decreased upwelling conditions. A northward expansion of the southern westerlies during this period and synchronous northward shift of southeast trade winds would have suppressed upwelling (Figure 6j). Such a mechanism would effectively result in higher SSTs causing wetter conditions in the winter rainfall zone. Stager et al. (2012) demonstrated that high percentages of moderately brackish-water diatom record of Verlorenvlei indicated relatively high rainfall, which was sufficient to favor planktonic forms over littoral assemblages, but not enough to favor dilute-water taxa (Figure 6i). However, the record of terrestrial input at GeoB8323-2 indicates that conditions on land were still dry (Granger et al., 2018). This mismatch was possibly caused by variable proxy response, suggesting that the rainfall might be enough to cause wetter conditions on land but still not higher to result in increased fluvial discharge to the ocean. An increase of G. ericsonii and higher Gephyrocapsa-based SSTs (Figure 6h and g) started later at ca. 740 CE in core GeoB8323-2. The timing of warming based on alkenone SSTs, the abundance of the Stratified group, and Gephyrocapsa-based SSTs do not align, which is difficult to explain. The relative abundances of the Stratified group reaching maximum values at ca. 700 CE are followed by high alkenone SSTs at ca. 920 CE with Gephyrocapsa-based SSTs finally peaking at ca. 950 CE. One possible explanation for the temporal offsets between the different SST reconstructions might be that dinoflagellates and coccolithophorids (alkenone producers) are associated with different living habitats and growth seasons, and that there is some degree of competition between them. Dinoflagellates in the Stratified group (e.g. S. mirabilis) are all autotrophic species which depend on the availability of light in order to photosynthesize using nutrients, which are taken up directly from the water column (Schnepf and Elbrächter, 1992). Although in this study Spiniferites spp., S. mirabilis, and S. ramosus are determined to indicate warm and stratified water, they are generally reported as a cosmopolitan species which are not solely controlled by water temperature (Marret and Zonneveld, 2003). However, S. mirabilis are also typically formed during upwelling relaxation periods and can be present in high relative abundances in areas where salinity is reduced either seasonally or throughout the year (Zonneveld et al., 2013). This could well explain the low relative abundances of S. mirabilis between 30 BCE and 300 CE within the period of increased Agulhas leakage and higher salinity (discussed in section ‘Active upwelling and Agulhas leakage (300 BCE–450 CE, Zone I-1 and Zone I-2’), as opposed to the period of maximum abundance between 450 CE and 800 CE when Agulhas leakage decreased, and was accompanied by upwelling relaxation (Figure 3).
Between 450 and 800 CE, northern core GeoB8331-4 is characterized by low SSTs and, conversely, a steady decline in the percentages of cold and nutrient-rich water indicators (Figure 6a and b). Considering that core GeoB8331-4 is located in the central Benguela region, we infer that the northern location was too far away to be influenced by the southern westerlies and still experienced upwelling during this period.
Fluvial discharge (800–1850 CE, Zone III)
Since ca. 800 CE, the minimum representation of the Agulhas group and Stratified group and maximum representation of the Upwelling group I in the southern core GeoB8323-2, Figures 4 and 5a–c) indicate a cooling in the southern Benguela region, which is supported by a declining trend of alkenone-based SSTs (Figure 6e). The relatively high representation of the Upwelling group I (Figure 5c) indicates a return to cold and high productivity waters. This might be attributed to higher fluvial input from the Olifants River during this period, which would bring more nutrients from terrestrial sources that favor the growth of heterotrophic dinoflagellates (Graham and Wilcox, 2000). On the other hand, high nutrient input coupled with more freshwater might lead to low cyst concentrations because of competition with diatoms during enhanced productivity (Dale and Fjellså, 1994), which is indeed observed in both the cyst concentrations and accumulation rates of core GeoB8323-2 (Figure 5f). This interpretation is supported by the terrestrial input record (Figure 6f; Granger et al., 2018) and the diatom record in Verlorenvlei characterized by high percentages of dilute-water taxa (Figure 6i; Stager et al., 2012). These conditions are associated with a northward expansion of the southern westerlies, in particular after 1150 CE, resulting in increased humidity and high fluvial input to the southern Benguela region. Meanwhile, less Agulhas leakage resulting from decreased intensity of the southern westerlies (Durgadoo et al., 2013) together with the advection of cold sub-Antarctic waters into the Benguela system during this period might also have contributed to the lower SSTs (Granger et al., 2018). The continued northward expansion of the southern westerlies after 1150 CE influenced the central Benguela region, resulting in suppressed upwelling and higher SSTs (Figure 6a). This offers a possible explanation for the second SST maximum at ca. 1200 CE. Subsequent, cooling at both sites was characterized by a minimum representation of cold and nutrient-rich water indicators and maximum representation of fluvial-nutrient signals in the northern core GeoB8331-4 (Zhao et al., 2017). The declining trend of cold and nutrient-rich water indicators could be explained by increased fluvial input from the Orange River which showed an increase after 50 BCE (Hahn et al., 2016; Zhao et al., 2017). The higher fluvial input favors the growth of fluvial-nutrient indicators resulting in a decline of cold and nutrient-rich water indicators.
Upwelling return (1850 CE–present, Zone IV)
After ca. 1850 CE, the fluvial input started to decrease in both regions (cores GeoB8323-2 and GeoB8331-4). In the southern core GeoB8323-2, however, both of the Agulhas group and the Upwelling group II as well as alkenone SSTs slightly increased around this time. On one hand, the increase of the Upwelling group II suggests upwelling with partly comparable dinoflagellate communities as during the period 300 BCE–30 BCE, while the slight increase in percentages of the Agulhas group suggests some increase in Agulhas leakage, likely because of southward contraction and increased intensity of the southern westerlies. In the northern core GeoB8331-4 the cooling trend continued, characterized by minimum alkenone SSTs and increased cold and nutrient-rich water indicators suggesting increased upwelling.
On the other hand, the increase of the Agulhas group and the Upwelling group II might be attributed to human activities. Previous studies (Pospelova and Kim, 2010; Pospelova et al., 2002, 2005) of polluted estuarine sediments from the US and South Korea found dinocyst taxa being sensitive to changes caused by human activities (e.g. sewage or industry pollution) including cysts of P. schwartzii and P. kofoidii and Dubridinium spp. Thus, an alternative explanation for the increase of the Upwelling group II (in particular cyst of P. schwartzii and Dubridinium spp. in the southern core GeoB8323-2 would be increased nutrient enrichment because of human activities. In addition, the clear increase of O. centrocarpum, a species which is capable of forming large populations under certain conditions, might indicate an opportunistic response to environmental change marking the first signal of human influence (Dale et al., 1999; Thorsen and Dale, 1997). Moreover, salt production has been found as one of the few economic activities from the period prior to 650 CE in South Africa (Antonites, 2016), thus the increase of P. zoharyi (which notably inhabits high-salinity surface water (Marret and Zonneveld, 2003)), could be likely attributed to the influence of the salt industry on the west coast of South Africa.
Conclusion
Organic-walled dinoflagellate cyst records for oceanographic conditions were used in conjunction with published data (alkenone-derived SSTs, grain size, and coccolithophore records) from two marine sediment cores (GeoB8323-2 and GeoB8331-4) to document the late-Holocene oceanic variability of the southern Benguela upwelling system over the Namaqualand mudbelt off SW Africa. The results indicate a distinctive behavior of the southernmost Benguela system compared with the central upwelling area. From ca. 300 BCE to 450 CE, the oceanic conditions varied synchronously in both areas, reflecting intensified upwelling in the entire southern Benguela region, which was proposed to be caused by a southward contraction of the southern westerlies. An influence of warm and saline waters from the Agulhas leakage was detected, but only in the southern core GeoB8323-2. Between 450 and 800 CE, less upwelling prevailed in the southern core GeoB8323-2, which was associated with a northward expansion of the southern westerlies. Contrary to the southern core GeoB8323-2, the location of GeoB8331-4 still experienced upwelling conditions until 1000 CE, suggesting that the position of the southern westerlies was too far away to influence the northern region. Between 800 and 1850 CE, higher fluvial input was observed in both regions. Since 1200 CE, the southern westerlies expanded further north, suppressing upwelling at both sites, and blocking Agulhas leakage as well as causing advection of cold sub-Antarctic waters into the southern Benguela, leading to a synchronous SST decrease at both locations. After 1850 CE, the southern core was characterized by increased SSTs with either intensified upwelling conditions and more influences of the Agulhas leakage or increased human influences in the southern core, while the northern core is characterized by a continued decline of SSTs, increased cold and nutrient-rich water indicators suggesting intensified upwelling in the northern core.
Based on these findings, we infer that the oceanic conditions in the Benguela upwelling region are driven by an interplay of wind-induced upwelling, fluvial discharge, and advection of cold sub-Antarctic waters with additional effects of warm water via the Agulhas leakage in the southernmost Benguela. Our study indicates that the signature of the Agulhas leakage on the water properties on the inner shelf (Namaqualand mudbelt) opens possibilities to trace Agulhas leakage modulation with its effects on the Atlantic meridional overturning circulation during the late-Holocene. Further work (more high-resolution core studies in the entire southern Benguela region) in this region will help us to better understand the behavior of the Agulhas leakage.
Supplemental Material
Supplementary_file_for_The_Holocene – Supplemental material for Late-Holocene oceanic variability in the southern Benguela region driven by interplay of upwelling, fluvial discharge, and Agulhas leakage
Supplemental material, Supplementary_file_for_The_Holocene for Late-Holocene oceanic variability in the southern Benguela region driven by interplay of upwelling, fluvial discharge, and Agulhas leakage by Xueqin Zhao, Lydie Dupont, Enno Schefuß, Robyn Granger and Gerold Wefer in The Holocene
Footnotes
Acknowledgements
Thanks to the captain, the crew, and the scientists of the Meteor cruise M57-1 for recovering the studied samples, and the GeoB core repository in MARUM (Center for Marine Environmental Sciences) for retaining the samples. We thank GLOMAR (Bremen International Graduate School for Marine Sciences) for supporting the first author’s PhD project. We would like to thank Karin Zonneveld for her help with the dinoflagellate cyst identification. Thanks to Annette Hahn and Gesine Mollenhauer for useful discussions. We thank the two anonymous reviewers and the editor Fabienne Marret for their helpful and constructive comments.
Funding
This work was supported by the German Federal Ministry of Education and Research (BMBF) (Grant/Award Number: ‘03G0840A’). The investigations were conducted within the collaborative project ‘Regional Archives for Integrated Investigations’ (RAiN), which is embedded in the international research programme SPACES (Science Partnership for the Assessment of Complex Earth System Processes).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.