
Abstract
We present a 12,6700-yr limnological history of Lake Miski, a high-elevation lake in a wet section of the Peruvian Andes. While many shallow Andean lakes dried up during the mid-Holocene, loss-on-ignition, magnetic susceptibility, and diatom analysis showed that Lake Miski was a constant feature in the landscape. Overall, fluctuations in the fossil diatom communities of Lake Miski tracked changes in insolation, but this was not the only mechanism influencing observed variability. We identify periods when insolation and interactions with the Pacific Ocean may have played a role in structuring local climate and diatom assemblages. The true mid-Holocene Dry Event (MHDE) is manifested in this record between 8000 and 5000 cal BP, but the carbonate stratigraphy and the diatom community indicated that although the level of the lake decreased, it never completely dried out, instead there was higher availability of planktic habitat and stronger mixing than in much of the Holocene. High rates of biological change observed during the late-Holocene in other records from Peru associated with human amplification of climatic signals were not observed in Lake Miski, as this lake may have been too wet and remote to be strongly influenced by human activity. Because of the presence of a woodland microrefugium, Lake Miski was suggested to have been an unusually climatically stable and wet location during the regional drying associated with the MHDE. Our new limnological information provides additional insights relating to this discussion. The presence of the observed woodland apparently withstood fluctuations that induced state changes in the lake and diatom flora, underscoring that microrefugia do not equate to ‘unchanging’ hydrologies or climates.
Introduction
The late deglacial and transition into the early Holocene in the southern tropical Andes was a time of falling lake levels. Precipitation appears to have been strongly influenced by precessional forcing for much of the glacial period, and this relationship held true until c. 9000 calibrated years before present (hereafter cal BP; Baker et al., 2001a; Cheng et al., 2013; Mosblech et al., 2012a). Subsequently, however, some paleohydrological patterns in lakes from Peru and Bolivia diverged from that of precessional forcing as increasingly evaporatic conditions appeared to influence systems more than changes in precessionally forced precipitation (Cross et al., 2000; Polissar et al., 2013). This phenomenon has been termed the mid-Holocene dry event (MHDE) and resulted in the lowest lake levels of the last c. 100,000 years on the Altiplano (Baker et al., 2001a). During the MHDE, microrefugia were probably necessary for the survival of some Andean species. Microrefugia are defined as isolated populations surviving in unusual microclimates relative to the surrounding landscape (Rull, 2009; Rull et al., 1988). Within microrefugia, species experience climatic conditions that differ from those of the adjacent climatic region. If these areas are large enough to support viable populations, the species can survive in the landscape until more favorable conditions return (Mosblech et al., 2011).
Lake Titicaca at ~6 (16° South) provides the best-documented and largest hydrological change associated with the MHDE. Submerged paleo-shorelines indicated a c. 100 m lowering of water level that resulted in a 50% loss of area of this great lake (D’Agostino et al., 2002). Fossil diatom data revealed increased salinity in Titicaca during the mid-Holocene between c. 9000 and 4400 cal BP (Tapia et al., 2003). These results were reinforced by studies of lakes Umayo and Lagunillas, both at 15 oS, which revealed lowstands between the oldest section of the core, at 7000 and 5000 cal BP (Ekdahl et al., 2008). Contrastingly, the only isotopic record from Bolivia was from the Sajama ice cap (Thompson et al., 1998), which did not portray a substantial change in precipitation associated with the MHDE. Abbott et al. (2003) studied a series of lakes between 13 and 18 oS and concluded that the MHDE was a time-transgressive phenomenon that was generally present between c. 6000 and 3000 cal BP, but started 2000 years earlier in the northernmost site compared with the southern Altiplano. Farther from the Bolivian Altiplano, the event was poorly defined temporally, as it seemed to vary regionally in its onset, peak, and termination. In southern Peru, three shallow lakes at 3400 m elevation in the upper portion of the Manu Biosphere Reserve at 13 oS all exhibited very slow sedimentation or sedimentary hiatuses between c. 12,000 and 4200 cal BP (Urrego et al., 2011). These data reflect a longer dry event that was typical for the MHDE. The much deeper Lake Pacucha, also at 13 oS in Peru, did not dry out during the MHDE. At this site, lake levels started to fall c. 15,800 cal BP, with a lowstand established c. 12,000 cal BP. A marked wet event at c. 8800 cal BP caused lake levels to rise, but between 8000 and 5000 cal BP, they fell to their inferred lowest level of the 25,000-year record. While the initial lowstand between 15,800 and 8800 cal BP coincided with the low of September–March (wet season) insolation at 13 oS, the later dry episode between 8000 and 5000 cal BP was tentatively suggested to be linked to Pacific sea-surface temperatures and the lack of El Niño Southern Oscillation (ENSO) activity (Hillyer et al., 2009). Paleo-precipitation records from speleothems and ice-cores documented that in northern Peru and Ecuador the long-term trend in precipitation tracked insolation, suggesting a general and gradual strengthening of the South American Summer Monsoon (SASM) over the course of the Holocene. The driest time, inferred from the least-depleted δ18O signal, was at c. 11,000–9500 cal BP, while the remainder of the Holocene saw a steady increase in precipitation (Bird et al., 2011; Cheng et al., 2013; Kanner et al., 2012; Mosblech et al., 2012a; Thompson et al., 1995; Van Breukelen et al., 2008).
Another source of Holocene precipitation variability in the Andes is ENSO (Haug et al., 2001; Moy et al., 2002; Rein et al., 2005; Sandweiss et al., 2001; Vuille, 1999). When ENSO is strong, it is the largest single cause of global interannual climate variability (Garreaud et al., 2009). Fluctuating irregularly, with pseudocycles of 2–7 years, positive phases induce warm El Niño conditions, while negative phases are associated with cold La Niña (Diaz and Markgraf, 1992). Warming (cooling) of sea-surface temperatures in the eastern tropical Pacific Ocean suppresses (enhances) rainfall over tropical South America, and enhances (suppresses) rainfall over the semiarid coast of Ecuador and Peru (Garreaud et al., 2009). ENSO events generally last for 18–24 months with peaks in amplitude mostly occurring in the austral summer (December–February; Grimm and Tedeschi, 2009).
Although Holocene El Niño histories are reported from a number of South American records (Conroy et al., 2008; Moy et al., 2002; Rein et al., 2005; Sandweiss et al., 2001), the inferred timing of active and inactive phases is not consistent between them. Evidence from clastic laminae in lake sediments from Ecuador point to strong El Niño activity in the early Holocene, a lull, and then resumed activity after 7 cal BP (Moy et al., 2002). In records from the Galápagos, the southern Peruvian coast and the tropical Pacific, however, diatom, seashell isotope and foraminiferal analyses showed no increase in event frequency until ~4.5 cal BP (Carré et al., 2005; Conroy et al., 2008; Koutavas et al., 2006).
Most evidence points to modern-style ENSO events being much weaker or absent between c. 8000 and 4500 cal BP (Moy et al., 2002; Sandweiss et al., 2001). Andrus et al. (2002) suggested that the eastern equatorial Pacific was 3–4°C warmer than modern between 8000 and 5000 cal BP because of reduced upwelling. Such a pattern could indicate a stronger westerly wind flow, which may have isolated the Altiplano from Amazon moisture flux (Theissen et al., 2008), that is, prevented SASM from penetrating as far west. These ideas, however, are not universally accepted, and there are also advocates of increased La Niña (cold phase) activity (Betancourt et al., 2000; Carré et al., 2012; Gagan and Thompson, 2004; Koutavas et al., 2006; Placzek et al., 2001; Rech et al., 2003; Tudhope et al., 2001), or that ENSO is not the most important explanation of climate changes of the past in areas remote from the upwelling (Baker and Fritz, 2015).
The effects of the MHDE on ecosystems have yet to be fully resolved, but increased aridity may have fostered more frequent and larger fires, causing grasslands to expand to replace woodlands or shrublands (Sylvester et al., 2014). One habitat that may have been adversely affected were Polylepis woodlands, an endemic fire-sensitive Andean tree that forms monodominant stands above treeline (Fjeldså and Kessler, 1996) providing essential habitat for high Andean woodland specialists including plants, birds, mammals, and insects (Cahill and Matthysen, 2007; Fjeldså, 1993; Lloyd et al., 2012; Yensen and Tarifa, 2002). Wet microrefugia (Mosblech et al., 2011; Rull, 2009) where the effects of the drought and fire were minimized, may have been an essential component allowing the survival and post-drought recovery of these groups. Using Polylepis pollen and charcoal (fire) records of the mid-Holocene, Valencia et al. (2016, 2018) suggested that steep landscapes with high terrain rugosity trapped moisture better than flatter ones during the MHDE, and could be used to predict microrefugial settings. Lake Miski in southern Peru was one of the sites identified as a probable microrefugium by Valencia et al. (2016), in which Polylepis forest remained a dominant component of the vegetation despite the evidence of increased fire activity. Addressing this site, our research question was: would the MHDE be evident in this system and were the changes associated with it as long-lived and strong as in other settings? Here, we present diatom, loss-on-ignition, and magnetic susceptibility (MS) data from Lake Miski that span the past 12,700 years. With these data, we investigate the effect of the MHDE on a wet, highly biodiverse, Andean ecosystem.
Study area
Lake Miski (13º 01’ 23.00” S; 72º 22’ 40.95” W, 3830 m a.s.l.) is located in a valley in the Eastern Andean Cordillera, at the head of the Huamanmarca Valley (Figure 1), 75 km northwest from Cusco city, Peru. Miski was originated by glacial activity, and thus, the valley in which it is located has a u-shape with slopes generally less than 20%. The moraine-dammed lake measures about 350 × 200 m, and lies within a catchment of about 1.6 km2 with a maximum elevation of 4200 m a.s.l. Lake Miski has a main and a secondary basin (200 and 60 m wide) separated by a shallow and narrow shelf. Two small streams feed the main basin (Zmax 10 m) from the northern and eastern shores. The lake has a single outlet with the stream flowing westward from the secondary basin (Zmax 3 m). There are multiple moraines within 2 km of the lake ranging between c. 3600 and 3900 m of elevation with most of them located above 3800 m. The largest moraines occur below Lake Miski at 3770 m a.s.l. Besides morphometric measurements extracted during the fieldwork and from satellite images, there is no limnological data available for Lake Miski.
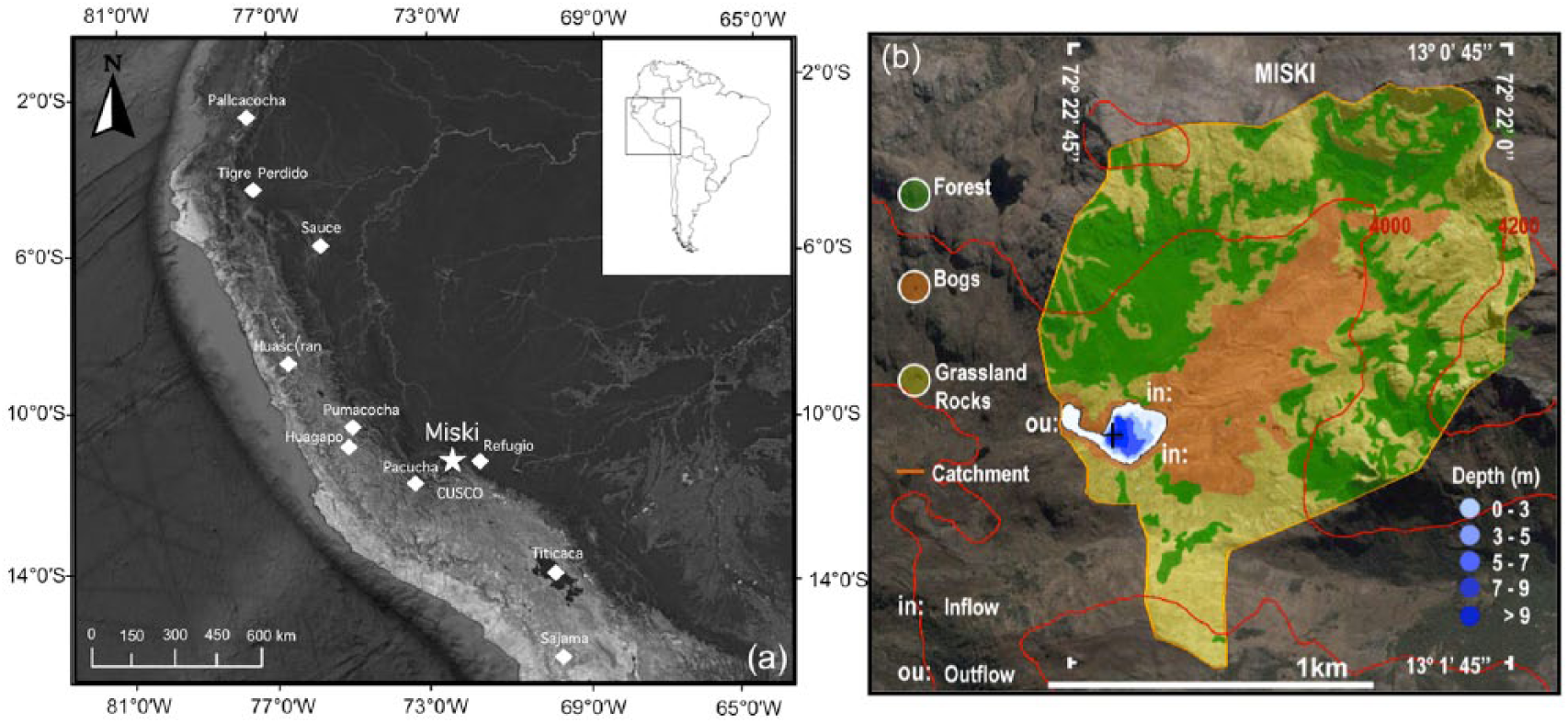
Location and bathymetry of Lake Miski, Peru. (a) Map of South America with focus on Peru. (b) Sketch showing Lake Miski catchment (orange line), surrounding vegetation, bathymetry (depths in m), and coring site (black cross). (c) Map showing the location of Lake Miski (star) in relation to other paleoclimate records (diamonds) discussed in the text.
The climate in this region is strongly influenced by the strength of the SASM, which is in turn influenced by the position of the Inter-tropical Convergence Zone (ITCZ) (Garreaud et al., 2009). During the wet season (austral summer), the ITCZ is located in its southward position bringing moisture from the Atlantic Ocean to Amazonia, fueling intense convective rainfall. The moisture is transported by a large-scale atmospheric circulation, the South American low-level jet (SALLJ), along the base of the Andes and reaches the study region. During the dry season (Boreal summer), the ITCZ reaches its northward position and brings convectional rainfall to the northern tropics (Vuille et al., 2000). Insolation controls the latitudinal movement of the ITCZ by increasing the amount of moisture convergence over the continent during periods of increased land–sea temperature differences (Biasutti et al., 2003). Correspondingly, the long-term variations of precipitation observed in paleorecords from South America, are frequently dominated by changes in insolation, thus being primarily driven by orbital forcing (Cruz et al., 2005; Kanner et al., 2012, 2013; Mosblech et al., 2012a).
The average precipitation in the region is c. 800 mm per annum. Approximately 80% of the annual precipitation in the region falls between November and April (Garreaud and Aceituno, 2001; Garreaud et al., 2003; Vuille, 1999). Besides precipitation, cloud cover is an important source of moisture. A dense cover of arboreal epiphytes on trees around Lake Miski suggests that cloud cover is a permanent feature even during the dry season. Mean annual temperatures are estimated to be c. 11°C (Zimmermann et al., 2010). While mean temperatures remain fairly constant during the year, daily fluctuations regularly exceed 20°C (Horn and Kappelle, 2009; Kessler, 2006). Night frosts are usual at this elevation, especially between June and August (Lauer, 1981).
The local treeline forms at 3740 m a.s.l., with scattered woodlands lying in sheltered locations as high as 4200 m. Andean forest exists downslope of Miski, but the woodlands within the catchment are dominated by Polylepis (Rosaceae), with Hesperomeles (Rosaceae), Gynoxys (Asteraceae) and Melastomataceae also present. Lake Miski has few trees growing within 10 m of the lake. During our field expedition, we observed that wood extraction was limited in this area, probably because of the steep slopes and wet climate (Valencia, 2014). Most of the branches on dead standing trees or woody material lying on the ground were sodden, and unsuitable for burning. Cattle grazed grasslands that formed the modern vegetation at the lake margin. Apparently, cultivation and deforestation were limited by the difficult access to the site and persistent high humidity. Earthworks, for example, terraces, were absent from the immediate surroundings of Lake Miski. Major Incan constructions such as Machu Picchu and Ollantaytambo, however, lay c. 20–30 km south of the lake.
Methods
Two sediment cores about 2 m apart from each other were collected from Lake Miski in December 2008 using a Colinvaux-Vohnout piston corer (Colinvaux et al., 1999). A total of 4.15 m (Miski-1) of sediment was retrieved in Lexan tubes from the deepest section of the lake. The cores were transported to Florida Institute of Technology for storage. MS was measured at 0.5 cm intervals using a GEOTEK core logger at the Land Use and Environmental Change Institute (LUECI) at the University of Florida. Suitable macrofossils or charcoal fragments for 14 C dating were not found, so dating was based on bulk sediments. Six subsamples for 14 C accelerator mass spectrometry (AMS) analysis were selected from sections that did not react with 10% hydrochloric acid (HCl), that is, low carbonate concentration, to minimize hard water error. Because there was no reaction to 10% HCl, samples were not pretreated with acid. Age measurements were performed at the National Ocean Sciences Accelerator Mass Spectrometry (NOSAMS). Age calibration were conducted within the Clam software (Blaauw, 2010) using the Southern Hemisphere calibration curve to generate the age model. Loss-on-ignition (LOI) analysis according to Dean (1974) with modifications made by Heiri et al. (2001) was conducted on 0.5 cm3 subsamples at 2-cm intervals. Sediment subsamples were heated at 550°C for 4 h to estimate the organic content, and at 950°C for 2 h to quantify carbonate content. Subsamples of 0.5 cm2 for diatom analysis were processed with hydrogen peroxide, according to standard digestion procedures (Battarbee, 1986) and permanent slides were mounted in Naphrax® (refractive index 1.7). Identification and quantification of diatoms were performed using a Zeiss Axioskop photomicroscope at 1000× magnification (Battarbee et al., 2001). In each sample, a minimum of 300 valves was counted. Data were plotted using the software C2 (Juggins, 1991). The overall ecological changes through time and zonation were evaluated using Detrended Correspondence Analysis (DCA) using PC-ORD program, Version 5.15 (McCune and Mefford, 1997).
Results
Age model
All six 14 C dates were accepted and used to create the age model (Table 1; Figure 2). The Lake Miski sedimentary record spanned the entire Holocene period and was consistent with continuous sedimentation since ~12,700 cal BP. Based on the age–depth model, the analysis of diatom assemblages at a 2-cm interval provided an approximate centennial-scale resolution (~ 98-year resolution).
Calibrated ages for Lake Miski.
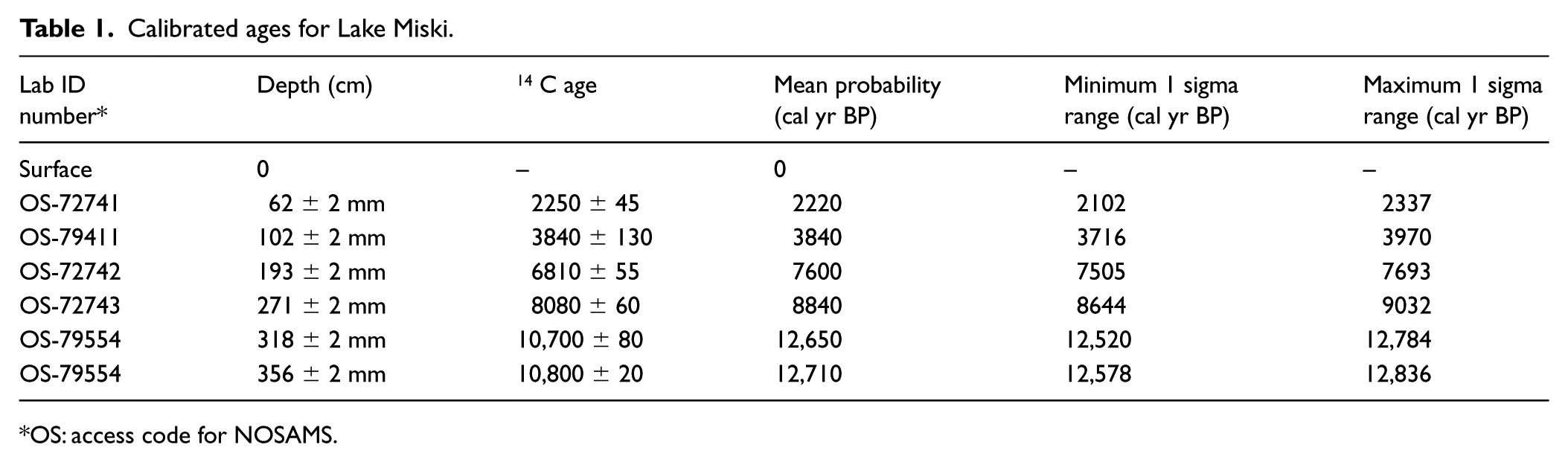
OS: access code for NOSAMS.
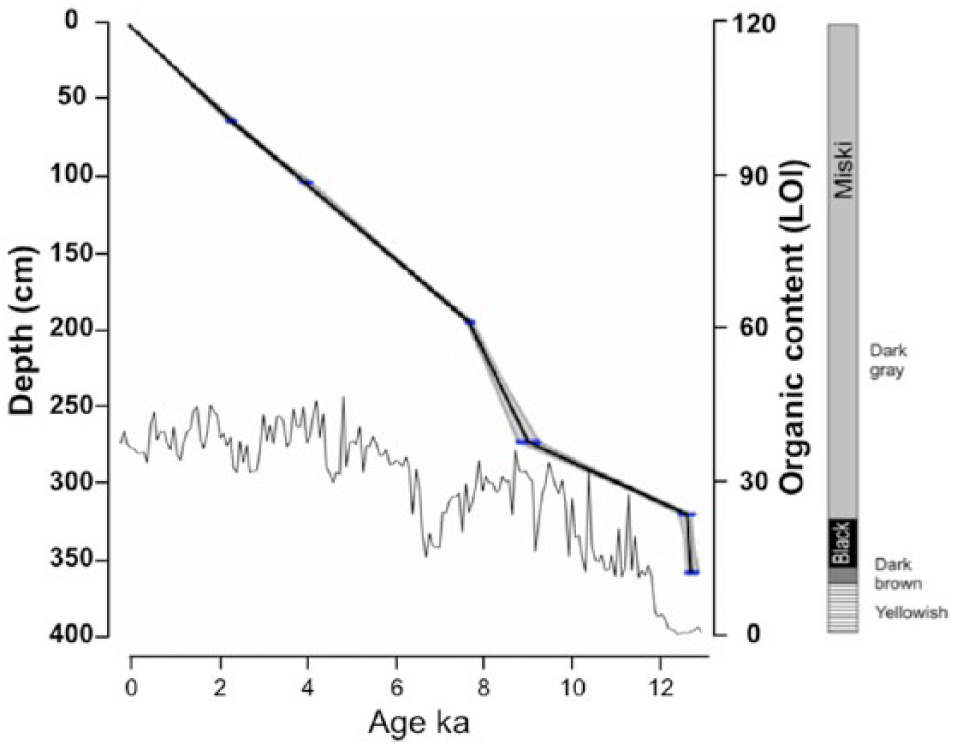
Age model, stratigraphy, and sedimentary organic content (LOI) from Lake Miski, Peru. The age model (shadowed line) was based on calibrated 14 C dates (Table 1), obtained using CLAM software (Blaauw, 2010). Transverse lines on the age model represent dates and errors. The gray area shows 95% confidence intervals based on 1000 iterations.
Diatom community changes through time
The most basal clays lacked diatoms, but between 350 and 0 cm depth the diatoms were well preserved. Over 200 diatom taxa were identified in the Miski sediments. Taxa with abundances >3% were portrayed individually in the diagram (Figure 3), however, all taxa were included in calculations. CONISS analysis provided an overly complex zonation of the record because of the large number of abrupt changes observed. Rather than follow that zonation, we use a subjective zonation supported by Detrended Correspondence Analysis (DCA) of the entire data set (Figure 4). Based on major changes in both ecological groups and DCA scores, we established six zones

Percentage fossil diatom data for selected taxa from the sediments of Lake Miski, Peru, showing zonation (Figure 4).
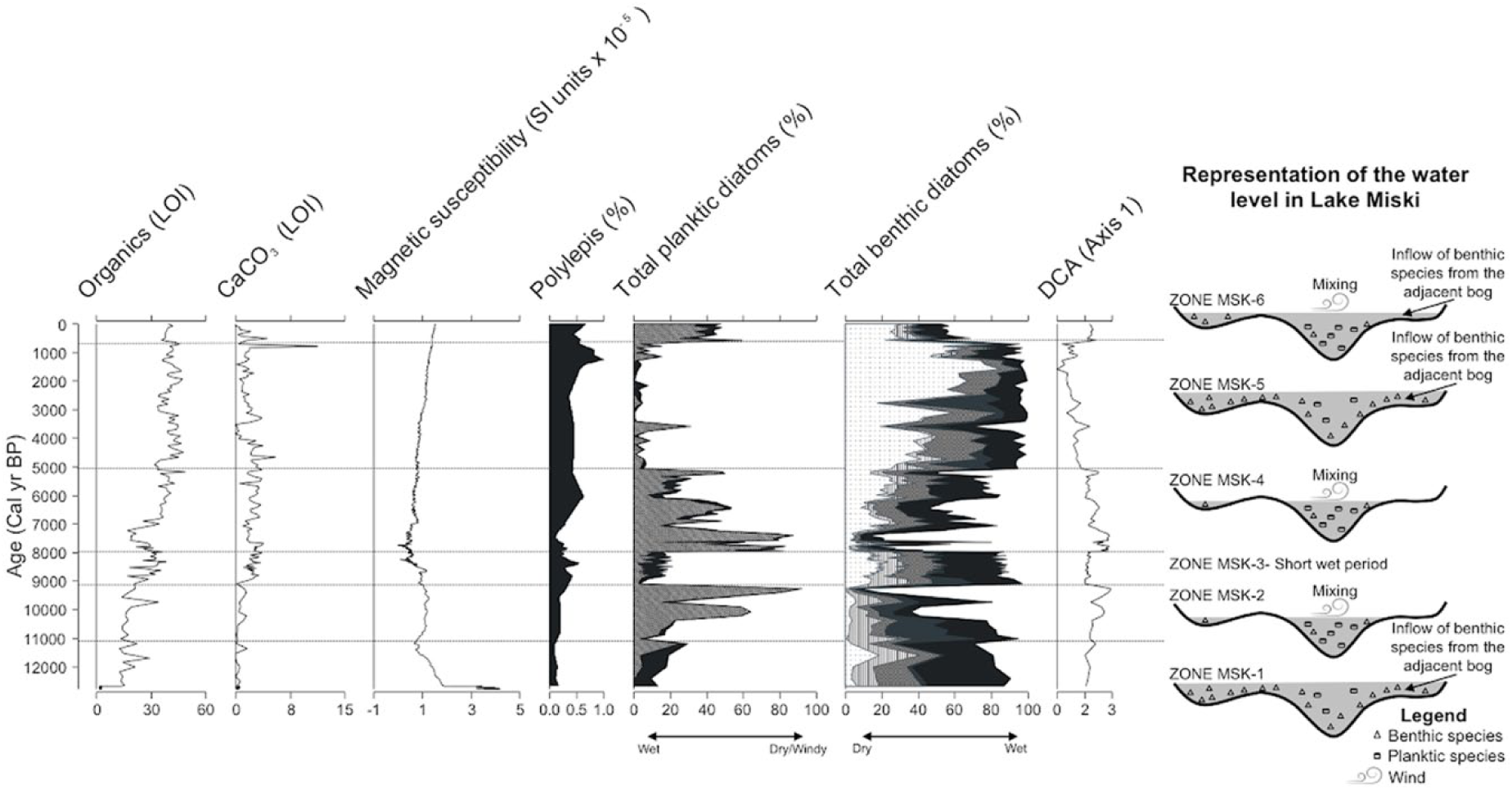
Summary diagram of loss-on-ignition (LOI), magnetic susceptibility, Polylepis (Valencia et al., 2016), diatom fossil from Lake Miski and schematic figure showing water level (in gray) oscillations over time. Dashed straight lines indicate the six zones. A square root transformation of the magnetic susceptibility data was made. Dashed angular lines:
Zone MSK-1: 350–302 cm; 12,600–11,100 cal BP
The sediment was a black-gyttja that showed fine, faint, laminations (Figure 2) with 1–15% of organics. Carbonate was lower than 1%, only reaching 1.5% in the last sample of the zone. This transition was evident in the MS values, which dropped abruptly by 15 SI units at 12,600 cal BP (Figure 4). Subsequently MS values declined steadily throughout this zone. In MSK-1, Brachysira, Achnanthidium minutissimum and Psammothidium were the most abundant diatom taxa, although Aulacoseira started to rise in abundance at 11,500 cal BP (Figure 3).
Zone MSK-2: 302–272 cm; 11,100–9100 cal BP
The sediment in this zone was a dark gray massive gyttja with fewer laminations than Zone MSK-1. Organic content reached 33%. Carbonate was still low and never exceeded 2.5%. MS did not show substantial changes in this zone. Pennate araphid diatoms from the genus Fragilaria and Tabellaria decreased from c. 13 to c. <3% after 11,000 cal BP. A small peak of Diploneis elliptica occurred at the beginning of this zone, but its abundance faded after c.10,300 cal BP. Planktic diatoms (centric) represented by Aulacoseira increased in two main pulses, at c. 10,500 and 9500 cal BP as the abundance of benthic diatoms, especially Achnanthidium and Brachysira, fell.
Zone MSK-3: 272–216 cm; 9100–7900 cal BP
The sediment in this zone was similar to that of Zone MSK-2, with a slight increase in carbonates to 3.5%. After c. 9100 cal BP, Aulacoseira decreased from c. 90% to 2% and benthics rose from c. 10% to c. 80%. MS values dropped 2 SI units at 8600 ka, presenting the highest oscillation in amplitude between samples since 12,600 cal BP.
This zone was mainly characterized by benthic taxa as Brachysira abundances increased from 11% to 25%. Achnanthidium and Brachysira had sequential peaks between c. 9100 and 8600 cal BP before decreasing again at c. 7800 cal BP. The benthic species Gomphonema, Cymbella cf. microcephala, and Psammothidium spp, and the planktic Tabellaria flocculosa increased slightly in abundance during this period.
Zone MSK-4: 216–134 cm; 7900–5100 cal BP
This zone was characterized by a dark gray gyttja sediment with organic content ranging from 16% at c. 7200 cal BP to 48% at c. 5100 cal BP, and carbonate reaching 3.8%. The high amplitude of the oscillations in MS ended at 7.5 SI. No major MS changes were observed afterwards. Zone MSK-4 started with the abrupt increase in the abundance of planktic diatoms as Aulacoseira rose from c. 8% to 80%. After c. 8000 cal BP, Aulacoseira fluctuated between 2% and 80%, but with an overall trend of decreasing abundance. Gomphonema and Tabellaria flocculosa increased in abundance during this zone.
Zone MSK-5: 134–20 cm; 5100–600 cal BP
The sediment was similar to Zone MSK-4 in terms of color. There was an increase in organic content from 35% to 49% after c. 5200 cal BP, and a decline in carbonate from 5% to 0.1% after c. 4500 cal BP. After c. 700 cal BP a peak in carbonate of 11% marked the end of this zone. This zone was characterized by the high abundance of benthic species, with the exception of a brief peak in Aulacoseira which reached 30% at c. 4500 cal BP. Multiple types of Eunotia, including E. cf. bilunaris and E. cf. incisa, reached their highest values of the record between 3500 and 1000 cal BP, reaching a total of c. >84% at c. 1500 cal BP. Brachysira abundances increased to 35% before an overall decline at c. 3000 cal BP.
Zone MSK-6: 20–0 cm; 600–0 cal BP
The sediment was similar to Zone MSK-3 with a steep decline in carbonates from 5% to 0% toward the present. Organic content dropped to 35% but increased again to previous levels in the present. This zone was defined by planktic species, as there was an increase in Aulacoseira abundance from 3% to 40% and of A. ambigua. The abundances of Achnanthidium minutissimum and Semiorbis hemyicyclus also increased during this period.
Multivariate analysis
DCA analysis of Miski (Figure 5) provided two interpretable axes with eigenvalues of 0.2 and 0.1, respectively. The negative extreme of Axis 1 consisted of samples MSK-3 and MSK-4 and was characterized by Gomphonema, Cymbella cf. microcephala, Tabellaria flocculosa, and Psammothidium spp. At the positive extreme of this axis laid samples from MSK-5 and MSK-6, characterized by Aulacoseira ambigua, Semiorbis hemyicyclus, and Eunotia cf. bilunaris. Samples from MSK-5 and MSK-6 also lay at the negative extreme of Axis 2, and were associated with several Eunotia spp. A mixture of samples from zones MSK-2 and MSK-1 lay at the positive extreme of Axis 2 characterized by Diploneis eliptica. Overall, the samples formed three recognizable groups with those of MSK-3 and MSK-4 being the most tightly clustered, while those of MSK-1 and MSK-2 showed greater scatter. Samples from the uppermost section of the core plotted similarly to some samples from the early Holocene.
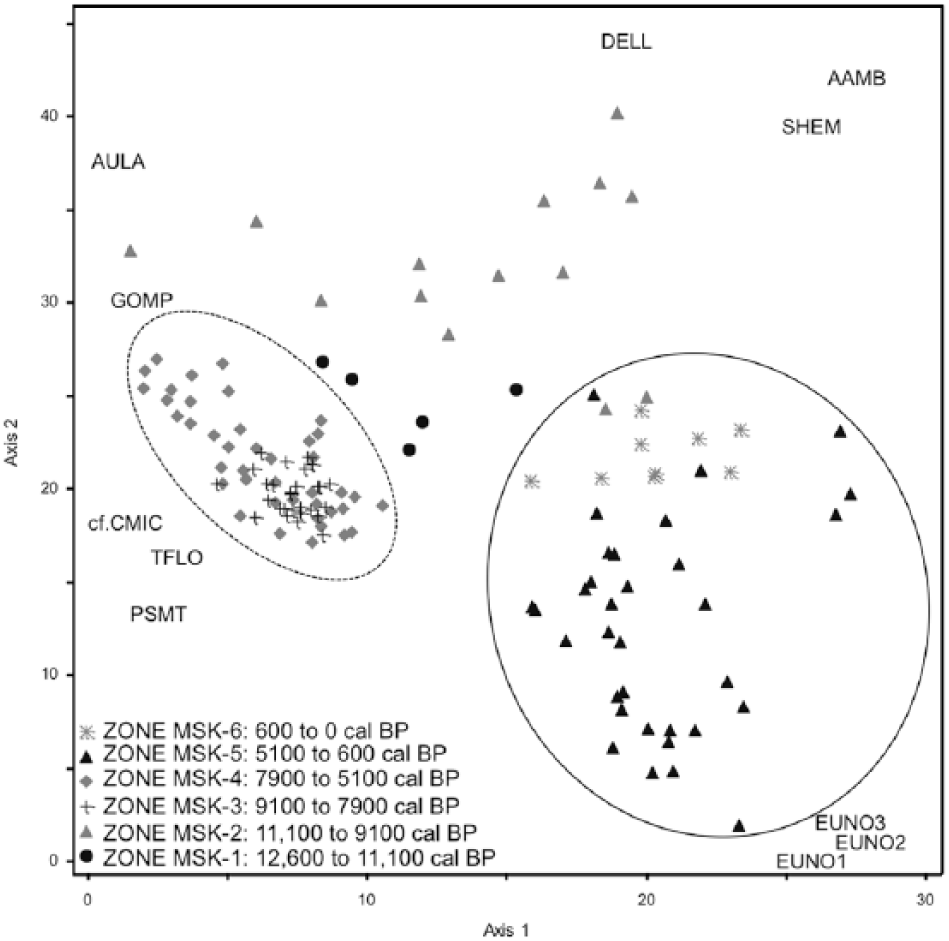
Detrended correspondence analysis of Diatom record from Lake Miski, Peru. Circles represent samples grouped by ages. Species codes – AAMB: Aulacoseira ambigua, AULA: Aulacoseira spp., cf. CMIC: cf. Cymbella microcephala, DELL: Diploneis eliptica, EUNO1: Eunotia cf. bilunaris – Group 1, EUNO2: Eunotia cf. incisa – Group 2, EUNO3: Eunotia spp. – Group 3, GOMP: Gomphonema spp., PSMT: Psamothidium spp., SHEM: Semiorbis hemicyclus, TFLO: Tabellaria flocculosa. Full circle includes all the samples from Zones MIS- 6 and MIS-5 (0–5100 cal BP). Dashed circle includes samples from Zones MIS-3 and MIS-4 (5100 to 8900 cal BP).
Discussion
Lake Miski formed when local glaciers retreated about c. 12,700 cal BP generating a decline in sediment MS, as less glacial silt washed into the system. This retreat coincided with the onset of the Younger Dryas (YD) in northern Europe (Alley, 2000; Bakke et al., 2009). Although some records from South America suggest glacier advances during this period (Clapperton et al., 1997), the pattern seen in Lake Miski is in agreement with records from the southern tropical Andes. The glacier retreat in these regions was attributed to the weakening of the Atlantic meridional overturning circulation and consequent reduced precipitation that caused abrupt regional warming through land-surface feedbacks (Jomelli et al., 2014; Rodbell and Seltzer, 2000).
An important observation in the context of regional lake records is that Lake Miski existed continuously throughout the last 12,700 years. Although there is evidence of changing limnological conditions during the MHDE, open water was consistently maintained. For the past 12,700 years, with the exception of the periods between 10,500 and 9100 cal BP, 7900 and 5000 cal BP and the last millennium when Aulacoseira spp. reached almost total dominance, benthic diatoms dominated the diatom flora of Lake Miski (Figure 3), suggesting a persistent, shallow system. The period between the formation of the lake and 11,500 cal BP (Zone MSK-1), corresponding to the YD period, was characterized by high concentrations of the benthic diatoms Brachysira spp. and Achnanthidium minutissimum, pointing to a shallow environment. In the period between 11,500 and 5000 cal BP (Figures 3 and 6a), sudden shifts in the community were evident, when the dominance of planktic diatoms indicated marked limnological changes, probably caused by changes in lake level.
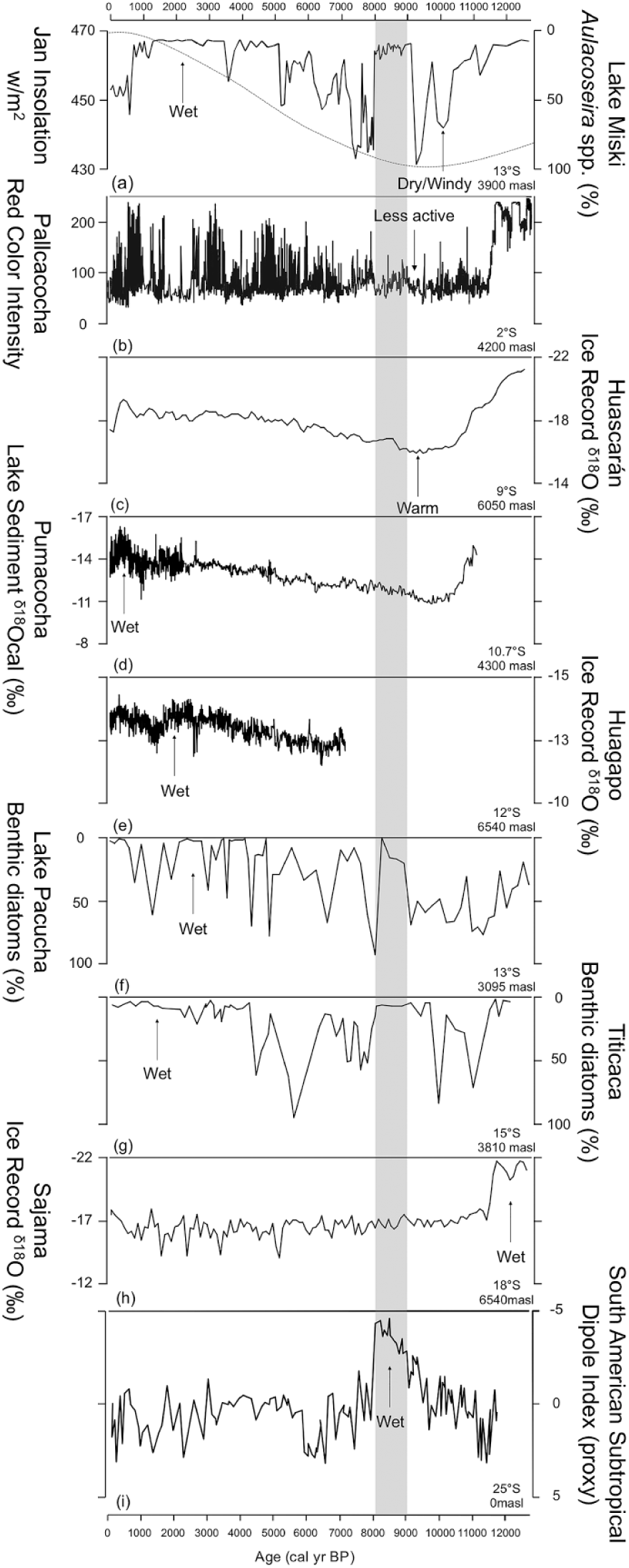
Precipitation data from selected sites from Peru showing evidence of precipitation changes at different latitudes compared with the abundance of benthic diatoms from Lake Miski, Peru (a). (a) Dashed red line represents January insolation. Other records are (b) Pallcacocha red color intensity compiled from record of sedimentation (Moy et al., 2002), (c) Huascaran δ18O ice record (Thompson et al., 1995), (d) Pumacocha precipitation inferred from lacustrine carbonate δ18O (Bird et al., 2011), (e) Huagapo Cave precipitation inferred from speleothem δ18O (Kanner et al., 2013), (f) Lake Pacucha benthic diatom relative abundance (Hillyer et al., 2009), (g) Lake Titicaca benthic diatom relative abundance (Baker et al., 2001b), (h) Sajama δ18O ice record (Thompson et al., 1998), and (i) South Atlantic Subtropical Dipole Index obtained from the Mg/Ca-derived SST record of the Northern Cape Basin and from foraminifera-based SST record at the southeastern Brazilian Continental upper slope (Wainer et al., 2014). Arrows indicate the trends of moisture balance and temperature in that record. Light gray bar indicates the 8–9 ka cold period observed in records around the world (Mayewski et al., 2004).
Although planktic species such as Aulacoseira are usually interpreted as an indicator of high lake level, the peaks in abundance of this group in Lake Miski between c. 9500 and 5000 cal BP are coincident with observed dry periods in Lake Titicaca (Baker et al., 2001b; Tapia et al., 2003) and Pacucha (Hillyer et al., 2009). If the increase in planktic diatoms is attributed to raised lake level, it suggests increased precipitation and contradicts regional records for the MHDE (Baker et al., 2001b; Bird et al., 2011; Hillyer et al., 2009; Tapia et al., 2003; Urrego et al., 2011). Thus, the Miski data raise two new questions: (1) Why do planktic diatoms dominate when most regional records suggest a period of lowered lake levels and (2) why was Aulacoseira the dominant genus of planktic diatoms in this shallow system?
Planktic diatoms and regional drought
The MHDE is observable in almost all paleoecological records from Ecuador to Bolivia. In temporal scale, it was the strongest series of droughts of the past ~100,000 years, and spatially it is believed to have been driven by insolation and ocean circulation, which are global, rather than local in nature. The dry events in other Andean lake records often left a signature of increased carbonate concentrations and benthic diatom percentages (Baker et al., 2001b; Hillyer et al., 2009b; Figure 6f and g). By comparison, the carbonate concentrations of sediments from Lake Miski (3–4%) remained close to the margin of error based on LOI analyses (Heiri et al., 2001), suggesting that the lake continued to overflow and did not become a closed-basin evaporitic system, and an increase in planktic rather than benthic species was observed (Figure 6a).
While Lake Miski does not show the usual signs of lowered water level during this period, we do not believe that our lake was decoupled from other records. The unique bathymetry of Lake Miski compared with other systems provides an explanation of the observed pattern. The lake has a flat bottom and a general depth of ~3 m, with a small deeper section, which we cored, that reached 9 m water depth (Figure 1b). We suggest that, in Miski, the decrease in precipitation during the MHDE did cause a lowered lake level, but the consequence was to reduce the area of shallows and bogs that provided a habitat for benthic species. Because most of the lake area is shallow, benthic diatoms would be favored during periods of high lake level, because of larger habitat availability. Another source of benthic diatoms is the bog around Lake Miski. Both of the Miski inlet streams flow through the bog, probably transporting benthic diatoms from it. Consequently, as the lake level falls, the shallow area available for benthic groups decreases and only the deeper portion of the water column remains, resulting in a more pelagic system, with an increase of the signal of planktic groups in the sediment record.
A gradual lowering of lake level between 11,500 and 9100 cal BP is supported by the increase in abundance of Eucocconeis/Nupela species at 11,000 cal BP. These genera are usually abundant at shorelines and in the periphyton of lakes with low conductivity and pH (Enache and Potapova, 2012; Potapova et al., 2003; Siver et al., 2007). The presence of Eucocconeis/Nupela may indicate a transitional system as the system either dries down or fills up. Thus, in this record, we infer that the peaks of Aulacoseira in the mid-Holocene could reflect times of low lake level.
Aulacoseira and the shallow lake
Aulacoseira is a tychoplanktic genus commonly associated with lake highstands in the Andes (Baker et al., 2009; Bird et al., 2011; Bush et al., 2005). Most Aulacoseira species, however, can grow in any depth of water so long as it is well mixed (Siver and Kling, 1997). Aulacoseira are drainpipe-shaped diatoms with small surface area:volume ratios that require turbulence to keep them in suspension (Padisák et al., 2009). The two most abundant Aulacoseira species in Lake Miski were A. distans-pusilla complex and secondarily, A. alpigena. The two species, which dominated the assemblage from 11,500 to 5000 cal BP, are usually associated with low pH mesotrophic systems, and well-mixed waters (Camburn and Charles, 2000; Gusev and Kulikovskiy, 2014; Holmgren et al., 2012; Morales, 2002; Saunders et al., 2009; Siver and Kling, 1997; Tsukada et al., 2006). The key here appears to be to explain how a small water body can be sufficiently well mixed to maintain Aulacoseira rather than other planktics.
Shallow, well-mixed systems can be created by surface turbulence (Lewis, 1996), which could come about through stream flow, or through cold nighttime temperatures coupled with increased wind speeds. Either of these causes is possible, though we are less inclined to believe stream flow during a lake lowstand as being sufficiently reliable to maintain mixing. The alternative (not mutually exclusive) explanation of temperature and wind effects has merit. Because of little thermal stratification, shallow tropical lakes are highly susceptible to mixing through surface winds (Lewis, 1996). Subtropical westerlies are suggested to have increased during the MHDE (Theissen et al., 2008), but the effects on wind fields in the Andes are not known. Today, high wind velocities associated with cold temperatures during the night result in very weak or no thermal stratification in high Andean lakes (Gunkel, 2000; Steinitz-Kannan et al., 1983).
Besides Aulacoseira spp., the DCA analysis (Figure 5) shows that the MHDE period was additionally associated with the benthic genus Gomphonema, and the tychoplanktic species Tabellaria flocculosa, both of which are consistent with the increase in availability of nutrients (Potapova and Charles, 2002). Although both species do not vary closely with Aulacoseira, Gomphonema and T. flocculosa were favored between 9100 and 5000 cal BP and do not reappear at the same abundances after. Taken together, the data suggest an increase in nutrient availability, caused by lowering of lake level, and stronger climatic volatility and mixing conditions during the MHDE period than in the prior or following zones (Figure 4). The MHDE was manifested in Lake Miski from c. 11,000 to 5000 cal BP as possibly drier and windier than the preceding millennia, but with lower habitat availability for benthic groups to dominate diatom assemblages.
The return of benthic species
The period after 9100 cal BP showed a remarkable change in the diatom flora with Aulacoseira falling from c. 85% close to 0% and benthic species dominating lake Miski assemblage. The high amplitude and frequency in both sediment MS and diatoms between 9100 and 7900 cal BP, suggests climatic instability and the increase in lake level with a consequent increase in habitat availability for benthic groups or inwash of benthic diatoms from the adjacent bog. As the wet event of 9100 to 7900 cal BP faded, Aulacoseira returned to dominate the system, such that Zones 2 and 4 closely resemble one another in terms of lake mixing and climatic volatility. The well-mixed, rich pelagic lake re-established at c. 7900 cal BP, but every few centuries, conditions reverted to being a benthic rich habitat. When precipitation increased and high lake level conditions returned, the dominance of Aulacoseira was reduced, leading to a stair-step pattern of state changes that gradually led to a humid benthic rich lake system after c. 5000 cal BP.
The effects of the MHDE in Miski faded abruptly at 5000 cal BP when the system shifted to a state without prior analog. Although an assemblage dominated by benthic species was re-established in Lake Miski, indicating increase in shallow area and benthic habitat availability because of increase in lake level, the dominance of Eunotia species indicates that the lake became more acidic (Pavlov and Levkov, 2013). The increase in the abundance of Eunotia had begun c. 9000 cal BP, and was consistent with the rise in organic sediment after that time (Pavlov and Levkov, 2013). Long-term acidification of lakes is a natural phenomenon, common in regions with base-poor bedrock or where the bedrock is sealed off by overlying sediment (Engstrom et al., 2000). Lake acidification can be caused by a number of factors (Fritz and Anderson, 2013). The most likely cause in this instance is that accumulated organic sediment released enough organic acids to exceed inputs of weathered bases from the country rock. The marked increase in abundance of Eunotia at 5000 cal BP coincides with organic sedimentary content reaching 40–45% dry weight from LOI indicating that this species is favored by the release of organic acids as they replaced rock flour on the bed of the lake. After 3500 cal BP, Eunotia became the most abundant genus in Lake Miski.
At c. 780 BP, carbonate content increased to c. 10% of the dry weight, a factor of four higher than prior background levels. This spike of carbonate was registered in only one sample. As Lake Miski is not located on carbonate bedrock and does not produce carbonate in the water column, it is likely that this spike represents an unidentified external source or a methodological error. Although this spike may be an artifact of analysis, the diatom community underwent a sharp shift from dominance by Eunotia spp. to an Aulacoseira alpigena-dominated system. This sudden change in the diatom community indicated that there was a genuine event that later resulted in changes in the community.
This transition to the Aulacoseira-rich system was maintained even though carbonate concentrations reverted to indicate the return of a lake system with some through flow. Regional and global data point to a period of increased precipitation during the past 700 years (Figure 6e and f; Hillyer et al., 2009; Kanner et al., 2013) associated with the increase in ENSO frequency (Figure 6b; Conroy et al., 2008; Moy et al., 2002), so the mechanism favoring Aulacoseira during this period could be different from the one observed during the mid-Holocene. Increased run-off, stream flow, or wind would be consistent with maintaining a mixed system in such a shallow lake system. The mechanism underlying this shift from a calm, acidic environment dominated by Eunotia to a mixed lake, dominated by Aulacoseira is still unknown. Nonetheless, the limnological transition that occurred in Lake Miski at 700 cal BP appears to have continued until the present.
Coherence between records
We compared the species variability in the Miski record with other paleoclimate records to derive insights into the mechanisms driving the observed changes (Figure 6). The following discussion is based on the fluctuations observed in Aulacoseira at Lake Miski. We choose to use the Aulacoseira curve because it is the most important group in disrupting the overall dominance of benthic species in Lake Miski, and because the variability observed in the DCA analyses was mostly driven by the changes in this genus. The Aulacoseira fluctuation variability seems to match regional hydrological changes.
Prior studies have argued that the low water levels of the mid-Holocene came about through the interaction of multiple processes. Here, we provide additional data that help to clarify that the dry event was composed of three stages. The first from c. 11,000–9100 cal BP was a time of falling lake levels and increase in the mixing regime coincident with declining austral summer (wet season) insolation (Figure 6a). Between c. 9100 and 7900 cal BP, this trend was interrupted by a wet period. The third stage was a return to low lake levels and a well-mixed system, even though insolation was decreasing, and thus favorable to increase in wet-season precipitation. This last phase, from c. 8000 to 5000 cal BP matches the timing of the true mid-Holocene dry event in other records, as described by Abbott et al. (2003).
Regional isotopic records show a peak of moisture between c. 20,000 and 16,000 cal BP (Baker et al., 2001b; Cheng et al., 2013; Mosblech et al., 2012b; Thompson et al., 1995, 1998). The subsequent nadir in precipitation that tracks the insolation minima at 10,000 cal BP is present in some records, for example, at Lake Junin (Seltzer et al., 2000), Lake Pumacocha (Figure 6d; Bird et al., 2011), Huascarán ice record (Figure 6c; Thompson et al., 1995), and El Condor cave (Cheng et al., 2013). In many other sequences, however, there was either no Holocene decrease in moisture, for example, the Sajama ice cap (Figure 6h; Thompson et al., 1998) and paleolake Uyuni (Baker et al., 2001a), or there was a later reduction in lake levels, for example, Lakes Titicaca (Figure 6g; Baker et al., 2001b), Taypi Chaka Kkota (Abbott et al., 2003), and Pacucha (Figure 6f; Hillyer et al., 2009; Valencia et al., 2010).
Marine and speleothem records show a weakening of the Atlantic Meridional Overturning Circulation (AMOC) and consequent strengthening of the South American Summer Monsoon (SASM) at 8200 BP and this has become known as the 8.2 ka event (Alley et al., 1997; Cheng et al., 2009). This wet event is not apparent at Lake Miski. Yet, the 8.2 ka event, expressed as a cold event in the North Atlantic, is suggested to be a part of a larger, global event throughout the 9000–8000 BP interval (Mayewski et al., 2004), which was identified in Lake Miski between 9100 and 7900 cal BP as a period dominated by benthic species. During this time, the Northern Hemisphere was more glaciated than today and changes in ice sheet extent and enhanced meltwater production would have altered ocean circulation affecting climate worldwide. The 9000–8000 BP signal is observed in the South Atlantic Ocean subtropical dipole index (Figure 6i), reconstructed from records of SSTs from the coasts of Africa and Brazil for the Holocene (Wainer et al., 2014). We hypothesize that the abrupt changes observed in Lake Miski from 9100 to 7900 cal BP was a wet event that coincided with the 9000–8000 BP interval, perhaps driven by a cooling in the North Atlantic SSTs and changes in ocean circulation modulating the South Atlantic Ocean subtropical dipole. As stated before, in Lake Miski the increase in lake level would have increased the area of shallows, and thus increased the representation of benthic species.
Between 8000 and 5000 cal BP, the last phase of the MHDE in Lake Miski matches the timing of the MHDE in other records (Abbott et al., 2003) and weakening of ENSO events (Figure 6b; Moy et al., 2002). Although the synchronicity in terms of timing and direction of change observed between records indicates that insolation and ENSO are two different mechanisms (Polissar et al., 2013), millennial-scale variability in ENSO has been suggested to be associated with orbital changes (Clement et al., 1999; Moy et al., 2002) or modulated by the northward displacement of the ITCZ favoring more permanent southeast trades and promoting cool upwelling (Koutavas et al., 2006), but the linkage between both mechanisms remains uncertain.
While other lakes were losing large amounts of water to evaporation (Cross et al., 2000), Miski retained its status as a lake, but shallowed and became well mixed. Increased wind speeds could have facilitated mixing and enhanced evaporation, generating changes that were compatible with observed records from drier settings. The reason that Miski retained water, when other lakes were dry was probably because of topographic trapping of moisture that formed low-level cloud on the Andean flank (Halladay et al., 2012). Even when windy, this setting would have frequently been cloud-saturated. During dry episodes, while flat areas desiccated, the topographically rugged terrain around Miski would have trapped orographic moisture. Another complementary possibility was that the rugosity prevented the spread of natural fires in the landscape. With such natural fire breaks, fire-sensitive forests would have been maintained (Valencia et al., 2016). Maintaining mesic forests, their soils, and microclimate would have, in turn, enhanced the evapotranspirative feedbacks that promoted cloud formation. Unsurprisingly, this small, wet setting manifested the termination of the MHDE ~ 1000 years earlier than at the much drier, and larger setting of Lake Titicaca (Baker et al., 2001a).
Although Lake Miski and Lake Pacucha were located near one another, after 5000 cal BP the responses of both lakes became decoupled. Lake Miski experienced a long period in which higher lake level caused an increase in benthic habitat availability, and acidification allowed Eunotia spp. to replace Aulacoseira spp. as the dominant diatoms. Contrastingly, at Lake Pacucha, small tychoplanktic and benthic diatoms that had dominated during the mid-Holocene dry event were replaced by Aulacoseira ambigua, whose abundance oscillated continuously with small tychoplanktic species abundance, suggesting a system that was essentially very unstable with temporary stratification and mixing (Hillyer et al., 2009).
One possible explanation for the divergence between the signals in Miski and Pacucha after 5000 cal BP is the relative role played by humans in each catchment. At Lake Pacucha, the peaks in benthic diatoms were ultimately interpreted as dry events associated with ENSO activities, but the authors noted the increased rates of change associated with this period compared with glacial times and proposed that the intensified limnological variability may have been because of human influence (Hillyer et al., 2009). A similar limnological amplification of the ENSO signal because of human activity was reported from Lake Sauce, Peru, in the same time period (Bush et al., 2017). At Lake Sauce, as agricultural activity intensified, there was amplification of an existing, but subtle climatic signal of ENSO. This signal only became apparent when the buffering of natural vegetation was replaced by erodible, deforested landscapes. Supporting this idea was evidence in the pollen record for sustained human activity both at Pacucha and Sauce for the past 5000 years, but for very limited use at Miski, in which pollen of crops were not observed (Valencia, 2014).
An event that caused a sudden change in the diatom community was observed at c. 780 cal BP from Lake Miski and this is coincident with regional records reporting strong changes in precipitation regimes. In the Lake Pacucha and Quelccaya ice cap records a dry period observed at ~ 700–800 cal BP (Hillyer et al., 2009; Thompson et al., 2000). Following that event, the Miski system reverts back to an Aulacoseira-dominated system, with abundances similar to those observed during the mid-Holocene. The high abundance in Aulacoseira could be associated with the weakening of ENSO activities observed in Laguna Pallcacocha, Ecuador (Moy et al., 2002).
Microrefugia and limnology
Local microrefugia are central to regional species persistence during adverse times, and are of increasing interest to conservation biologists (Birks and Willis, 2008; Cheddadi et al., 2017; McGlone and Clark, 2005; Rull, 2009). In the high Andes, Polylepis woodlands that exist above the treeline are critical to the survival of a set of habitat specialists (Fjeldså, 1993; Lloyd and Marsden, 2008; Terrazas and Ståhl, 2002). These woodlands are lost from fire-prone dry landscapes and are suggested to have survived the MHDE in wet microrefugia. Lakes that lie in steep terrain that would have caught Amazonian moisture are suggested as wet spots that may provide long-term moist conditions (Killeen et al., 2007). Such sites were shown to host Polylepis woodlands throughout the MHDE, whereas sites with low terrain rugosity exhibited sedimentary hiatuses or lost their Polylepis woodlands (Valencia et al., 2016). Lake Miski was identified as a potential microrefugial setting and so our new limnological information provides additional insights relating to this discussion. First, the conditions apparently needed for woodland microrefugia do not translate to ‘unchanging’ hydrologies or climates. Our evidence is consistent with the existence of an open water body throughout the MHDE, and therefore of relatively mesic conditions compared with other local settings, that is, the drying of the Refugio lakes (Urrego et al., 2011), which lie atop a flat landscape. Our data though, did suggest changes in climate.
The variability observed in Lake Miski is in agreement with other records, meaning that there were drier conditions during the MHDE (Figure 6), but the responses observed in our site were not as sensitive as in other sites, thus supporting that Lake Miski could have been a microrefugium. Factors that induced state changes within the lake were not enough to cause woodlands to disappear. Thus, for a setting to be microrefugial did not imply unchanging climates, only that conditions were ‘good enough’ to facilitate survival of the species of interest. Altered wind speeds, dry events, and cloudiness were all inferred from our data, and yet these changes were not sufficient to cause the drought-sensitive species Polylepis to be lost from the landscape. What cannot be answered with certainty was whether other plant, bird, or insect species that coexist within modern – or early Holocene – Polylepis woodlands were unaffected.
Going forward, microrefugia will have to withstand anthropogenic climatic change, and will exhibit species turnover as individual niche spaces no longer fit within the available climate space (Hannah et al., 2014). Definitions of microrefugia are, therefore, somewhat subjective in terms of which species are identified as characterizing the microrefugium. Given our data, it is probable that climate change will influence all settings, including future microrefugial habitats. Consequently, single-species conservation objectives, even associated with microrefugia, will be less likely to be viable, than planning on protecting a broad swath of species with similar (but not the same) ecological tolerances or functions.
Conclusion
Lake Miski has been a continuous feature of the Andean landscape for the past 12,700 years. Limnological changes in the diatom and carbonate records from the lake show that early Holocene precipitation variability at this highland site was influenced by orbital insolation, consistent with other records from the region, and consistent with reductions of moisture inputs from SASM as ITCZ neared its most northerly position in response to low austral summer insolation. Superimposed on this long-term trend, the decrease in ENSO activities may also have played a role in controlling precipitation variability of the Andes. Between 8000 and 5000 cal BP, and possibly for the past 600 years, the variability in Lake Miski seems to be associated with El Niño activity variability. Another possibility is that insolation also modulates ENSO (Koutavas et al., 2006) and that both are being expressed in the precipitation record.
Although probably occasionally used by humans, this site does not show strong signals of human occupation. The high rates of limnological change occurring after the end of the MHDE associated with human influence at nearby Lake Pacucha were not observed in Lake Miski.
Lake Miski was suggested to have been an unusually climatically stable and wet location during the regional drying associated with the MHDE. The presence of a woodland microrefugium apparently withstood changes that induced state changes in the diatom flora, underscoring that microrefugia do not equate to complete climatic or hydrologic stability.
Ongoing and future anthropogenic climate change poses one of the greatest threats to biodiversity, affecting species distributions and ecological interactions. In the Andes, changes in climate are expected to disrupt precipitation patterns and, of particular concern, to increase the intensity and frequency of droughts (Bradley et al., 2006; Urrutia and Vuille, 2009). In Miski the effects of recent climate change are still unknown, but the evidence is that recent warming in the region is occurring at a higher rate than the global average (Vuille et al., 2003; Wang et al., 2014), already impacting glaciers and water availability (Rabatel et al., 2013), and limnological characteristics of the water (Michelutti et al., 2015a; Michelutti et al., 2015b). Food security, water availability, and maintenance of biodiversity are all ecosystem services that are dependent upon environmental quality. Policymakers and conservationists need to understand how ecosystems responded to past climate changes so that these essential services can be maintained. From a paleoecological viewpoint, past changes in climate could have resulted in alterations that are similar to those observed under recent climate change scenarios, and thus may be a useful tool in planning for water availability and conservation in the Andes.
Footnotes
Funding
This work was supported by the National Science Foundation (Grant nos. BCS0926973, EAR-1303831, and DEB-1624207) and the National Aeronautics and Space Administration (Grant no. NNX14AD31G). The authors wish to thank the people and the government organization of Peru for allowing us to work in their country.