
Abstract
Severe aridity during the mid-Holocene, ca. 8.0–4.0 kyr BP, led to extreme ecological stress in the tropical Andes. Here, we report paleolimnological and archeological data from Lake Suches in southern Peru (70° 24’ 12” W, 16° 55’ 35” S) spanning 13.6–4.4 kyr BP. Integrated paleoclimate and archeological data reveal that moisture was locally available and the basin served as an ecological refugium throughout the mid-Holocene. Mid-Holocene aridity was established no later than 7.2 kyr BP, with maximum aridity ca. 5.5–4.8 kyr BP. However, water levels in Lake Suches were sustained throughout peak middle-Holocene aridity, even as other systems desiccated. Isotopic enrichment of water in Lake Suches (δ18Olake) and extensive wetlands (δ18Obofedal) surrounding the lake indicate prolonged residence time. These reservoirs, combined with elevation-linked hydrographic factors, mitigated mid-Holocene net decreases in atmospheric moisture. Archeological data from Suches indicate successive population increases beginning ca. 11.0–9.8 kyr BP as drier but more stable early Holocene conditions were established regionally. Population maxima in Suches during the mid-Holocene/mid-Archaic period ca. 9.0–7.0 kyr BP coincide with peak aridity in the Titicaca and Atacama systems, as well as documented archeological hiatuses in these regions. Population decreases coincide with peak aridity recorded in Lake Suches ca. 6.0–5.0 kyr BP, but the basin was never fully abandoned. Evidence for refugial microenvironments is key to understanding the persistence of human populations and other endemic Andean flora and fauna during the highly adverse climates of the middle-Holocene. We outline several mechanisms which likely explain the formation of refugia linked to bofedales and hydrographic characteristics of Suches. Understanding refugial dynamics will be key to understanding the effects of past climatic change, as well as addressing current warming and decreased precipitation trends in the tropical Andes.
Introduction
The central Andean region (9–26°S latitude, Figure 1) was strongly affected by a protracted period of severe aridity during the middle-Holocene, from approximately 8.0 to 4.0 kyr BP. Estimated reductions in precipitation of up to 40% and temperature increases of 1–2°C desiccated groundwater systems, leading to extreme ecological stress (Baker et al., 2001; Cross et al., 2001; D’Agostino et al., 2002; Ekdahl et al., 2008; Grosjean, 2001; Grosjean et al., 1997, 2001, 2003; Grove et al., 2003; Latorre et al., 2003; Núñez et al., 2002; Placzek et al., 2001; Quade et al., 2008; Rech et al., 2003; Rowe et al., 2002; Servant and Servant-Vildary, 2003; Servant-Vildary et al., 2001; Thompson et al., 1998, 2000). Despite unequivocal evidence for severe regional aridity, there is debate concerning the impacts of this middle-Holocene arid phase (MHAP) on early human populations in the south-central Andes (e.g. Moreno et al., 2009; Santoro et al., 2017). Nuñez et al. propose that severe middle-Holocene aridity led to a regional occupational hiatus or ‘silencio arqueológico’ in the Atacama region, mitigated only by localized oases that provided a toehold for much reduced populations (Grosjean et al., 1997, 2001; Núñez et al., 2002, 2013). Late Pleistocene–middle-Holocene human occupation (Figure 1a) does not show consistent patterns of settlement, abandonment, and relocation across central Andean coastal and highland areas, however (Aldenderfer, 1989, 1998, 1999; Capriles and Albarracin-Jordan, 2013; Cipolla, 2005; Craig et al., 2010; deFrance et al., 2009; Gayo et al., 2015; Grosjean et al., 2005; Klink, 2005; Latorre et al., 2013; Melgar, 2013; Neme and Gil, 2009; Osorio et al., 2011, 2017; Rademaker, 2010; Rademaker et al., 2014; Sandweiss and Rademaker, 2011).
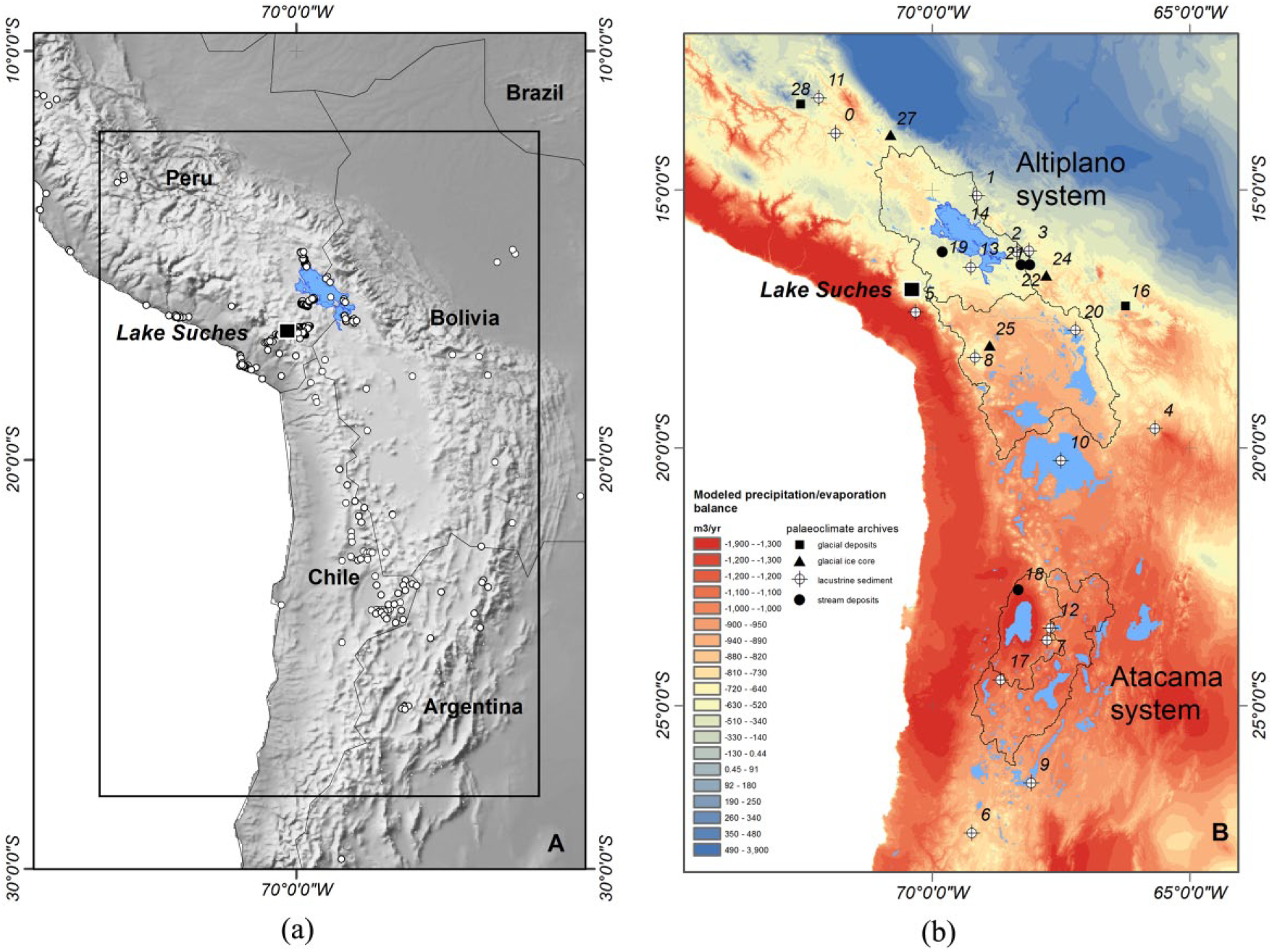
The Central Andean region, showing the location of Lake Suches (black square) relative to (a) documented Archaic-period sites (open circles) in the region and (b) modern precipitation/evaporation balances estimated from 522 instrumental records in the UN-WMO and NOAA-GCHN (Lawrimore et al., 2011) and CIARG-Global PET (Trabucco and Zomer, 2009) databases. Black lines show major catchments in the Titicaca and Atacama systems delimited from SRTM 1-arc second data. Also shown are locations of paleoclimate records.
There is growing evidence that isolated, humid microenvironments persisted throughout the MHAP, providing refugia which were crucial to the region’s ecological viability and early human settlement. The MHAP regional hiatus and shift toward local refugia in the Atacama is not mirrored in the Titicaca basin overall, where settlement regionally was more continuous and even intensified in certain locales. More intense MHAP human activity suggests the Titicaca region served as a macroregional refuge, and further provided specific microenvironmental refugia that sustained communities during periods of maximum aridity.
Here, we present evidence from the Lake Suches basin in southern Peru (Figures 1 and 2) for MHAP refugia, which sustained early human populations despite regionally adverse conditions. We reconstruct a coupled climate-human history of the basin from lacustrine and archeological archives. Sediments from Lake Suches provide evidence for sustained lake levels during the MHAP, and hyperaridity recorded in Titicaca and Atacama sequences is only weakly expressed. The Suches catchment also provides evidence of an early Holocene human occupation that persisted and even intensified during peak MHAP conditions, while the Atacama and other regions experienced a hiatus.
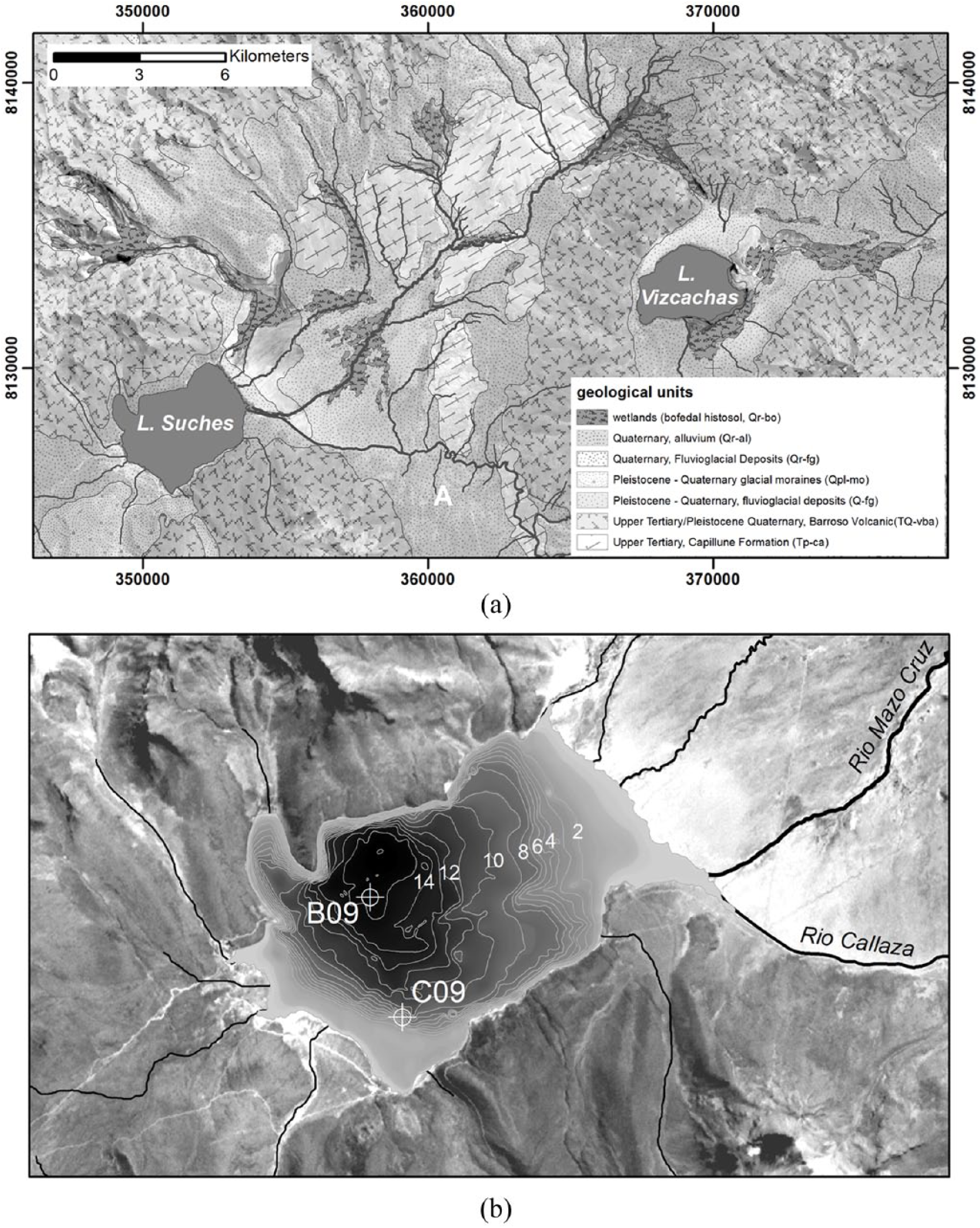
(a) Geology of the Lakes Suches and Viscachas catchments. (b) Bathymetry (m) of Lake Suches under modern conditions, showing 2009 coring locations. Coordinates are in UTM zone 19 S, WGS84.
Small, permanent lakes are rare in the Pacific-draining south-central Andean highlands, and Suches provides a unique opportunity to examine coupling between paleoclimatic signals and the human history of this region. We directly integrate paleolimnological evidence, as proxies for the precipitation/evaporation (P/E) balance, with archeological data reflecting the relative intensity and character of human occupation to understand how changes in moisture regimes throughout the south-central Andes from the late Pleistocene to middle-Holocene impacted cultural and population dynamics.
Despite the importance of climatic refugia during the MHAP and other periods of adverse climate, the mechanisms affecting their formation and human interactions are poorly understood. We outline physiogeographic mechanisms that likely contributed to stable hydroclimatic conditions around Suches, and promoted the formation of microenvironmental refugia: (1) Paleoclimatic reconstructions indicate that strongly seasonal rainfall patterns (the South American Summer Monsoon) were weakened during the MHAP, leading to overall decreased but temporally better-distributed precipitation (Abbott et al., 2003; Servant and Servant-Vildary, 2003). (2) Despite decreased atmospheric moisture, evaporitic loss at Suches was limited because of the lake’s geographic and hypsographic location, and smaller surface-to-watershed area ratio. Suches is similar to other elevation-linked refugia that maintained mesic environments under drier conditions. (3) Suches sits at the proximal end of regional groundwater systems and similar headwater lakes have been shown to maintain stable geochemical conditions during dry periods (Kratz et al., 1997; Webster et al., 1996). (4) Finally, Lake Suches is fed by extensive high-elevation Andean wetlands (bofedales) and porous aquifers that likely buffer lake-level variability. Paleoecological reconstructions of inter-Andean wetlands indicate that many are out of phase with regional climatic conditions, and maintain locally elevated water tables during climatic desiccation (Baucomb and Rigsby, 1999; Ledru et al., 2013; Servant and Servant-Vildary, 2003; Servant-Vildary et al., 2001). Archeological sites, which serve as index fossils of human responses to the MHAP, are strongly coupled with bofedales, indicating that these wetlands were key refugial microenvironments. The viability of these refugial microenvironments for human settlement suggests they may have been similarly important for other species during adverse climates.
Evidence for MHAP refugia in Suches refines mechanistic refugial models, helping to identify other refugial areas. A clearer understanding of these environments and how they relate to water resources is needed to accurately reconstruct the Holocene ecological, climatic, and human history of the central Andes. Furthermore, recent increases in aridity have been observed for the central Andes, and climatic models project overall decreases in future water availability because of continued warming (Dai, 2012; Urrutia and Vuille, 2009; Viviroli et al., 2011). Understanding the formation and role of Andean refugia will be crucial to addressing future human–climate interactions.
Site description
Regional climatic setting
Suches (70° 24’ 12” W, 16° 55’ 35” S) is within a high-elevation intermontane basin (4450–5200 m a.s.l.) located at the continental watershed of the central Andean cordillera (Figures 1 and 2). The climate of Suches is cool and semiarid, with strong diurnal temperature variations between 0°C and 25°C and high insolation rates. Modern central Andean climate is strongly affected by the South American summer monsoon (SASM; Garreaud and Aceituno, 2001; Garreaud et al., 2003, 2009; Zhou and Lau, 1998). Secular to intra-annual variations in precipitation are caused by shifting balances between humid, tropical, Atlantic/Amazonian and dry, subtropical Pacific/Antarctic air masses along the Intertropical Convergence Zone (ITCZ), influenced by orbital, solar, and oceanic-atmospheric factors (Abbott et al., 2003; Baker et al., 2009; Garreaud et al., 2003, 2009; Vuille et al., 2000; Vuille and Keimig, 2004).
The Suches basin is positioned in a region of low annual meteoric precipitation (300–400 mm isohyet) at the western edge between these two circulation systems and their hydrographic divide (Figure 1). Between 70% and 80% of precipitation occurs during the December–April austral summer wet season as orographic precipitation driven by atmospheric convection from the Amazonian basin (Figure 3). The lake is also located near the ‘arid diagonal’ – the approximate edge of a rainfall gradient separating the semiarid Altiplano and hyperarid Atacama systems (Bruniard and Moro, 1992). Consequently, Suches is an important location for understanding how shifting tropical (Atlantic-Amazonian) and subtropical (Pacific-Antarctic) air masses affected water availability in south–central Andean valleys and northern Atacama.
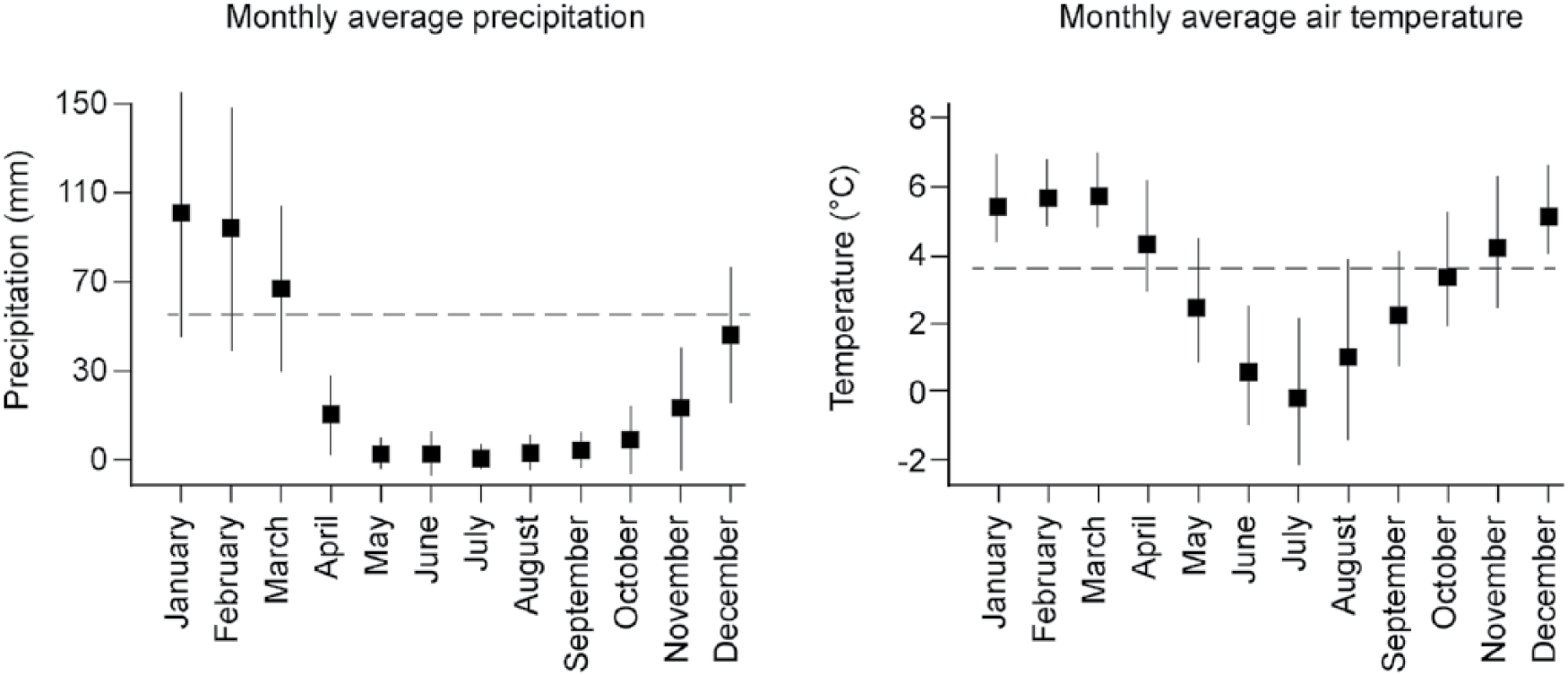
Instrumental records of monthly precipitation (1954–2008) and air temperature (1954–1984) means and standard deviations recorded at Lake Suches.
Lake characteristics
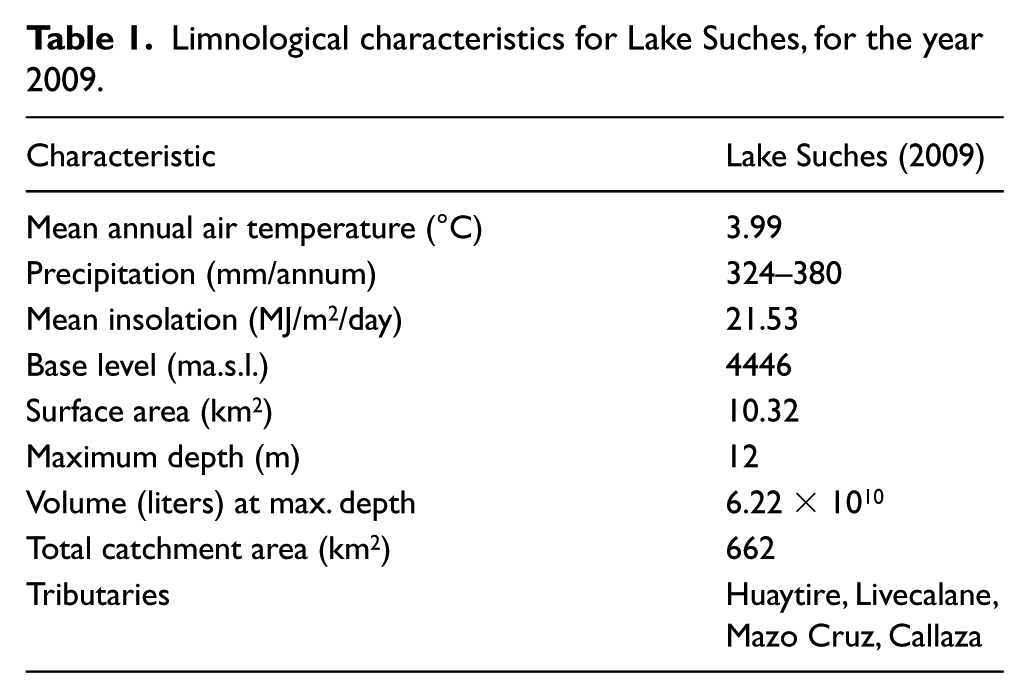
Lake Suches is a small, bowl-shaped endorheic lake (Figure 2). Estimates of the lake’s area and volume are based on bathymetric measurements made in 1983; base level was approximately 4 m lower at the time of coring, and calculations in Table 1 were adjusted accordingly. Recharge comes from perennial–ephemeral streams and associated wetlands, which are fed by meteoric sources (Figure 2). Remote sensing indicates there has been no neoglaciation in the basin since at least 1966–present; no glacigenic lacustrine sediments recovered in cores postdate the terminal Pleistocene. Two additional lakes are located in the basin. Of the three, Lake Suches perennially maintains water. The sill separating Suches from the adjacent catchment is approximately 15 m above mean modern base levels, necessitating a >2000% increase in lake volume before overflow occurs. With the exception of Miocene Maure paleolacustrine deposits in the northeastern basin (Sébrier et al., 1988), there is no indication that the basin was inundated beyond mean modern conditions, was connected to other catchments, or reached outflow conditions. Changes in lake status consequently are controlled by variations in the P/E balance.
Limnological characteristics for Lake Suches, for the year 2009.
The lake is bounded on its northern and southern shores by late Tertiary Barrosos-group andesitic massifs (INGEMMET, 1995; Sébrier et al., 1988). Pleistocene late glacial maximum moraines overly these formations and define the lake’s western shore, while the lake’s eastern shore is defined by the Pampa Huaytire, a large deltaic fan. Soil formation is minimal and consists largely of leptosols with localized bofedal histosols. Poor soils, aridity, and low mean air temperatures limit the vegetation to sparse bunch grass and shrub communities, with an important component of highly productive wetlands. These communities are a key to the water-balance and human land use history of Suches, and are described in more detail below.
Vegetation and moisture dynamics in bofedal wetlands
Vegetation in the basin consists of sparse mixed communities dominated by arid-adapted bunchgrass (Stipa spp.) or low shrubs (Parastrephia spp.). The most important communities with respect to local hydroclimates, human land use, and ecological networks are bofedal cushion bogs (Vining, 2011). Bofedales are high-elevation adapted, peat-forming wetlands that respond rapidly to changes in hydrologic regulation (Buffen et al., 2009; Earle et al., 2003). Bofedal biomass is approximately three times greater than in other high-Andean plant communities and consists principally of Juncaceae mat vegetation (Distichia muscoides) with minor suites of grasses, forbs, and flowering plants (Graf, 1981; Squeo et al., 2006). Bofedales provide necessary pasturage for Andean camelids (Bryant and Farfan, 1984; Patty et al., 2010; Reiner and Bryant, 1986). Many are further artificially enhanced by herders who improve graze quality by burning off bunchgrasses and expanding stream networks (Palacios Ríos, 1977; Verzijl and Quispe, 2013).
Autoregulatory feedbacks between moisture availability, vegetation, peat- and sediment deposition, and histosol development encourage the growth of wetlands and prolong groundwater availability (Earle et al., 2003; Graf, 1981). In turn, prolonged moisture modulates seasonal–interannual precipitation variability and sustains green vegetation through dry phases (Moreau et al., 2003; Moreau and Le Toan, 2003; Squeo et al., 2006; Vining, 2016). Selective grazing pressures suppress herbaceous vegetation with higher evapotranspiration rates in favor of rosette plants, further slowing moisture loss. These ecological factors work in concert to improve bofedales’ function as reservoirs. Isotopic values of bofedal water entering Lake Suches plot along the local evaporation line (see below), indicating that the wetlands prolong water residence time and influence lake water budgets.
Methods
Paleolimnological analysis
Oxygen isotope analysis
The effects of local evaporation and recharge on water budgets were evaluated by measuring the oxygen δ18O and hydrogen δD isotopic composition of water from Lake Suches (δ18Olake) and wetlands (δ18Obofedal) feeding the lake. A total of 30 mL water samples were collected in triple-rinsed polyethylene bottles capped underwater to remove trapped air. Oxygen isotopic ratios were measured at the University of Arizona Environmental Isotope Laboratory by CO2 equilibration with a Finnegan Delta S mass spectrometer. Hydrogen samples were reacted at 750°C with Cr metal. The precision is better than 0.1 ‰ for δ18O and 1.0 ‰ for δD (Table 2). Monthly precipitation isotope values were determined using the waterisotopes.org calculator (Bowen and Revenaugh, 2003). Values are presented in standard (δ) notation as per mil (‰) deviation from Vienna Standard Mean Ocean Water (VSMOW; Figure 4a, Table 2).
Isotope data from surface water samples collected from Lake Suches and inflowing streams and springs (coordinates are in UTM zone 19S, WGS84).
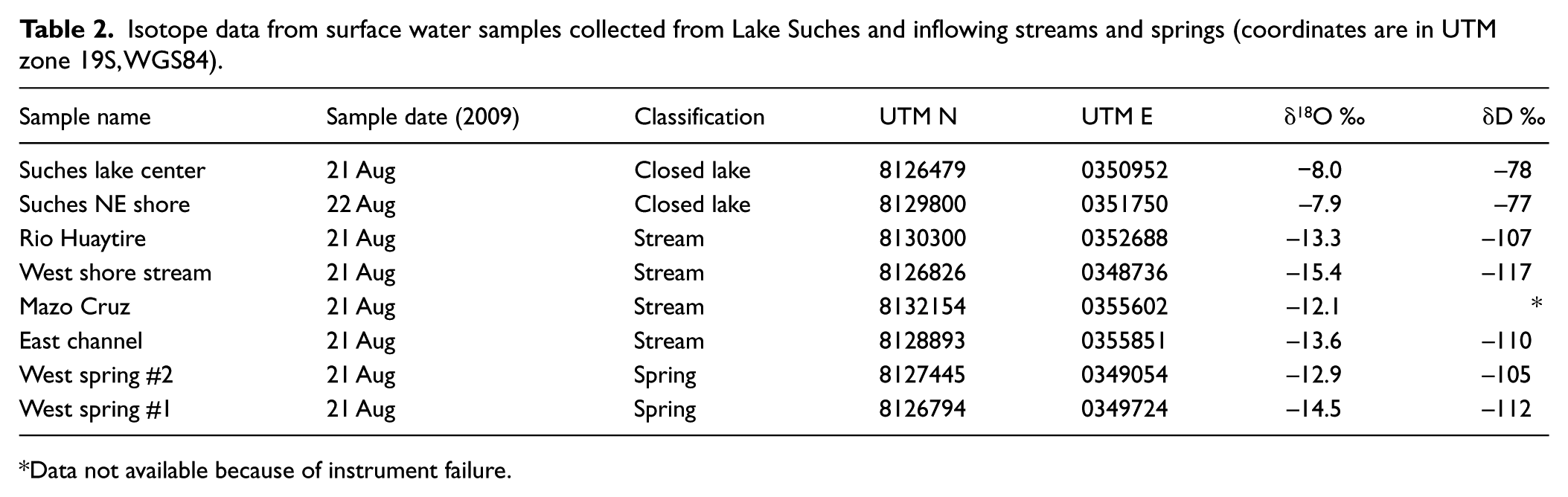
Data not available because of instrument failure.
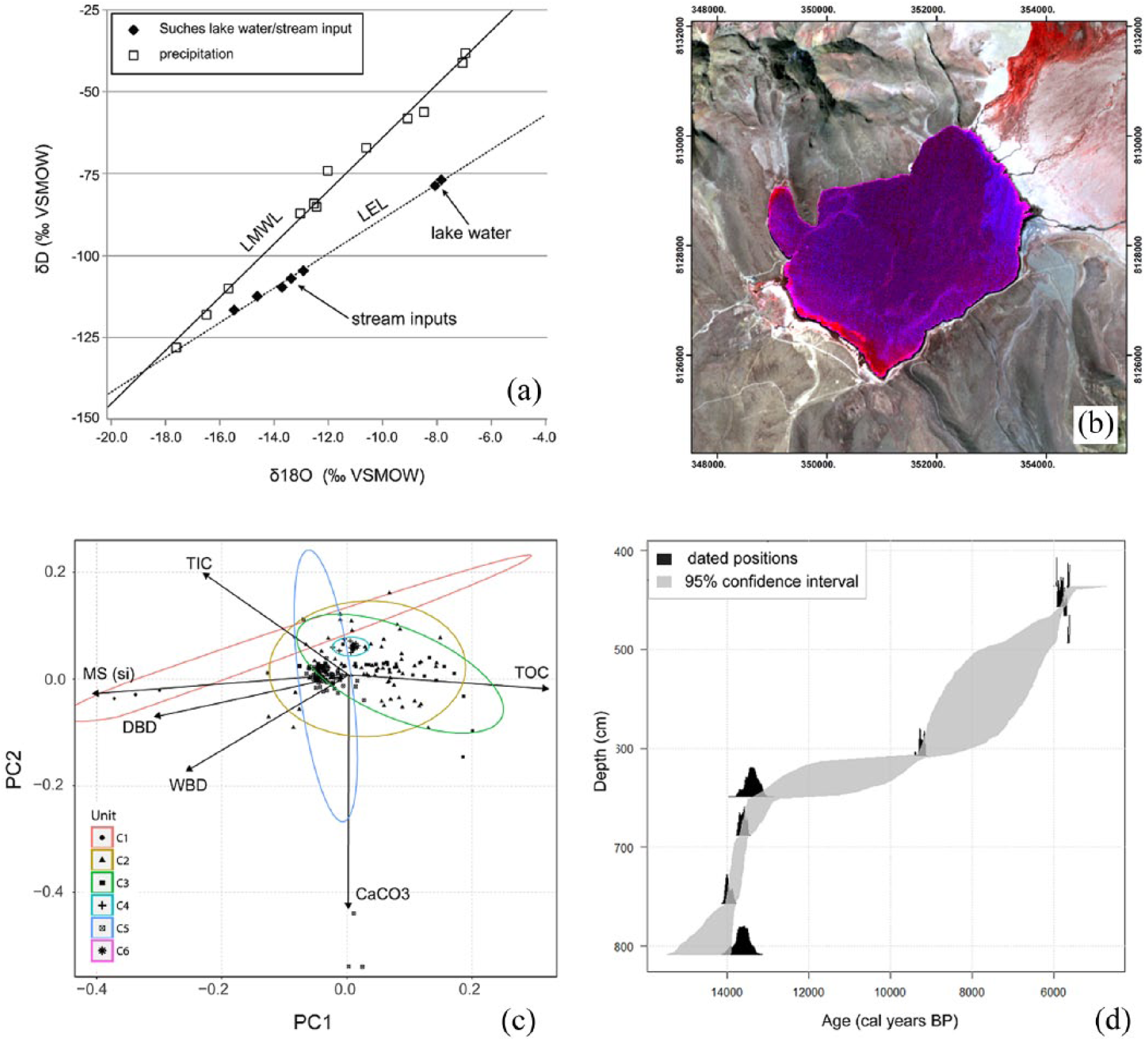
(a) Local meteoric water line (LMWL), local evaporation line (LEL), δ18O and δD values for Lake Suches water, inflowing streams and springs (open squares), and monthly precipitation (open triangles). (b) False color composite of ASTER multispectral data (3 July 2003) showing principal depositional environments in Lake Suches. Magenta shows peat-dominated near-shore deposits in shallow water (0–2 m). Lighter blue shows the extent of allogenic deposition, while deep water, authigenic environments are in darkest blue. (c) Principal components plot (PC1 and PC2) of sedimentary proxies with eigenvectors for each proxy. (d) Age-depth model (95% confidence interval) for Core C09 based on 14 C determinations in Table 3.
Core collection
Sediment cores were recovered from two locations at Lake Suches in August 2009; cores A-09 and B-09 were collected from the deepest portion of the lake to capture continuous deposition, and core C-09 was taken from the modern mean annual lowstand shoreline, to identify transgression/regression sequences (Figure 2). Coring methods followed protocols used at other high Andean lakes (Abbott et al., 1997a, 1997b, 2003; Binford et al., 1997; Bird et al., 2011b; Stansell et al., 2010). The upper 30 cm of flocculent material at the water–sediment interface (WSI) of core A09 was collected with a gravity corer and extruded in the field at 0.5-cm intervals into sterile plastic bags. Deeper sediments were collected with a 5-cm diameter Livingston piston corer in 1-m drives (Wright et al., 1984). Sediments were collected from B-09 to a depth of 4.6 m beneath WSI. Ten drives were collected from C-09 until refusal by basal glacial till at a depth of 8.2 m beneath the WSI. Here, we report on the lower portion of this core, from 4.0 to 8.2 m; the upper portion of this sequence (0–4.0 m and younger than approximately 4.0 kyr BP) was separated from import permits and destroyed during international transport.
Sediment analysis and facies identification
Physical sedimentological proxies were analyzed at the University of Pittsburgh Sediment Geochemistry Laboratory following standard laboratory protocols (Abbott et al., 1997b, 2003). Whole-core magnetic susceptibility (MS) was measured at 0.2-cm intervals using a Bartington MS2 C core logging sensor. Cores were subsequently split and one-half archived in refrigerated conditions. The work half was cleaned, photographed, and lithological units recorded using smear-slide minerology and detailed recording of Munsell color, sedimentary structure, and texture and biogenic features (Supplemental Material 1, available online). Samples measuring 1 cm3 were subsequently taken from the work half at 2.5-cm intervals. Wet and dry bulk densities (DBD) were measured on these samples prior to/after drying samples at 60°C for 36 h. Organic matter, carbonate content, and mineral matter were subsequently measured as weight% by loss on ignition (LOI) after heating samples to 550°C for 4 h and 1000°C for 2 h (Heiri et al., 2001). Results of the proxy analyses are shown in Figures 5 and 6.
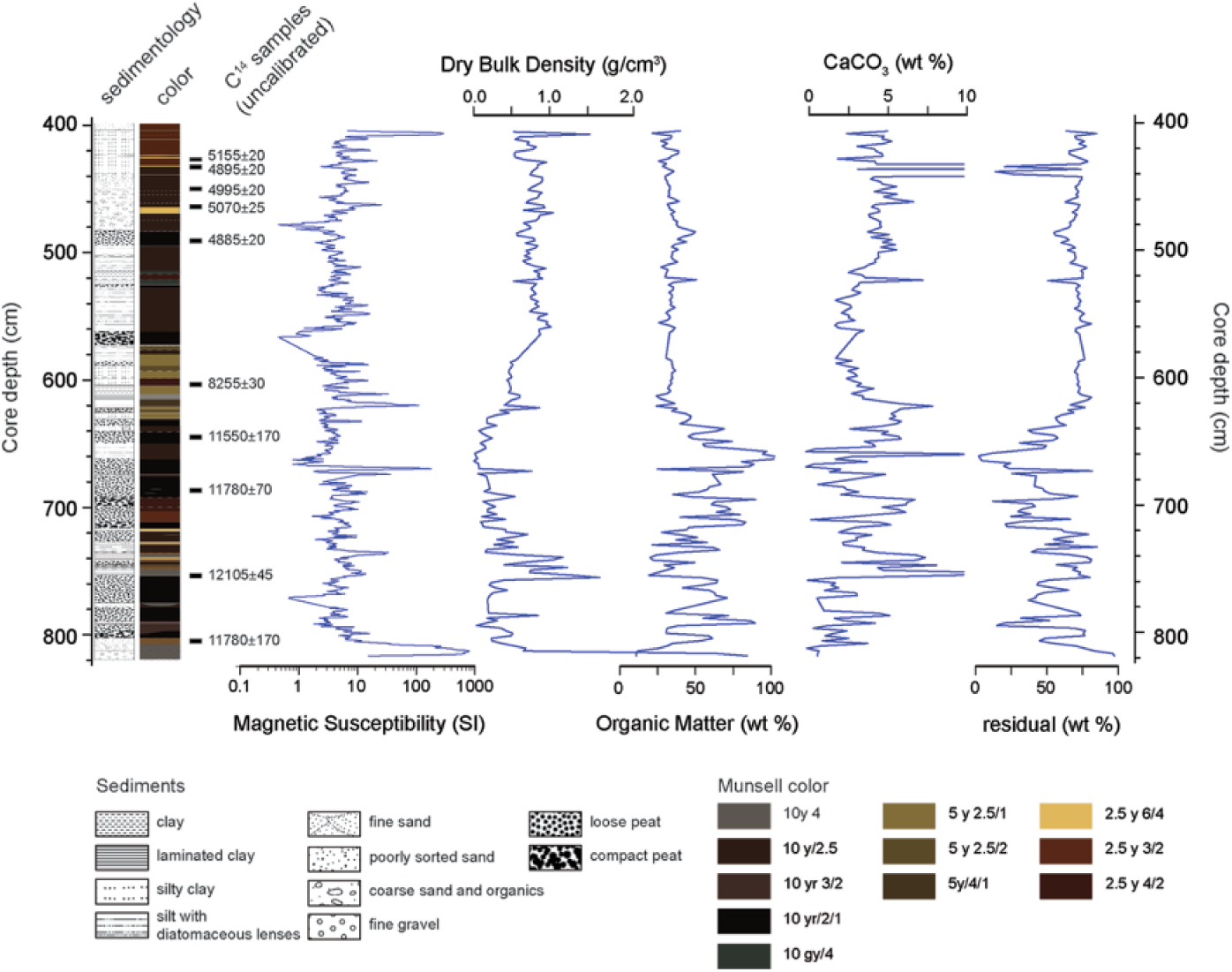
Composite proxy record for Core C09 (shoreline sequence) showing depth beneath the sediment–water interface, core lithology, locations, and ages of 14 C samples and lake-level proxies.
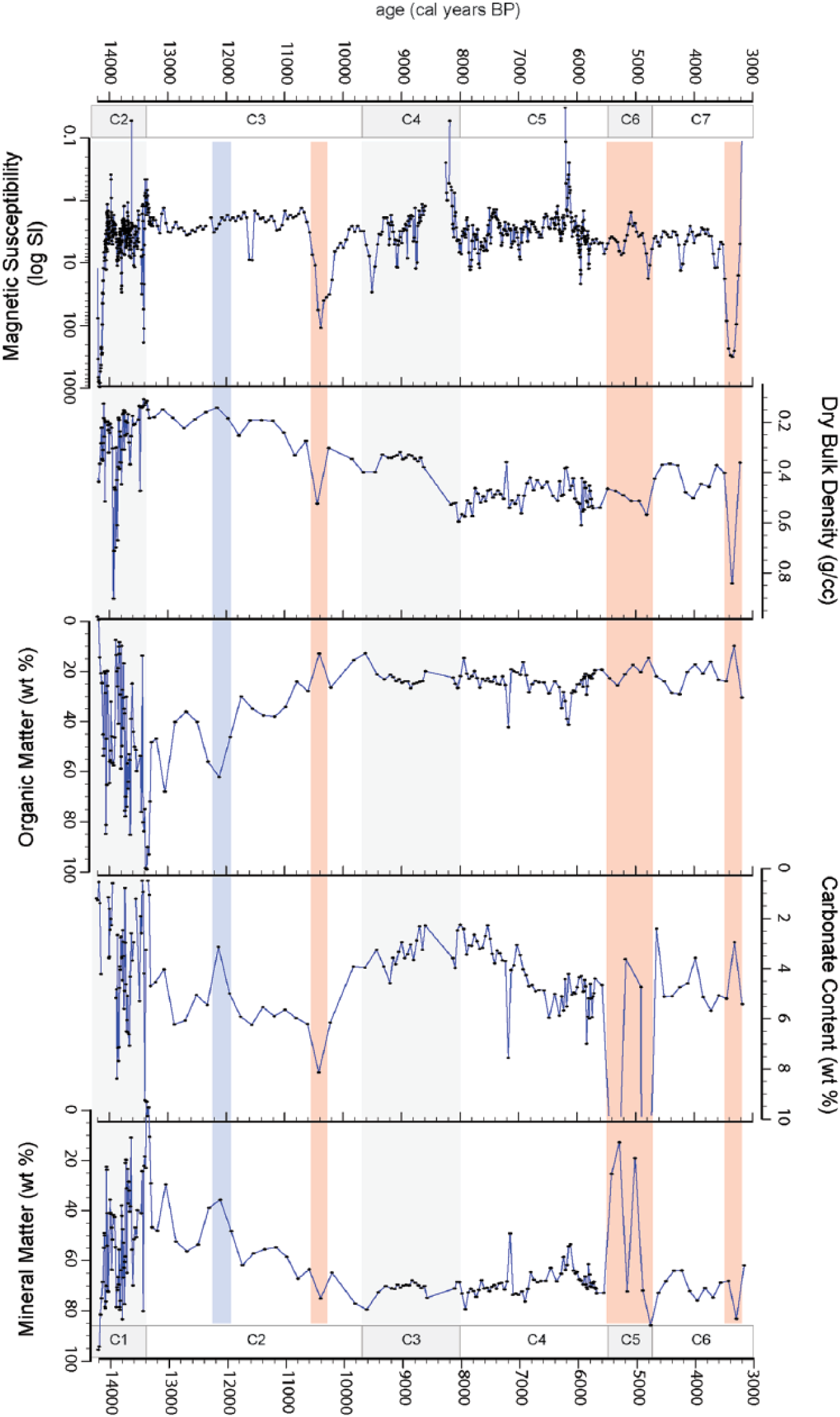
Lake-level proxies adjusted to the age-depth model with stratigraphic units indicated. Shading indicates more humid (blue) and more arid (red) interludes within each unit.
Modern depositional environments were used as analogs for relating core sediment facies to relative lake level. Exposed subaerial erosional surfaces, shallow-water facies, and deep water facies are present in core C-09 and provide approximations of higher to lower lake levels. Erosional surfaces were identified by abrupt (<0.5 cm in thickness), truncated contacts separating distinct facies. In cases where these are overlain by allochthonous sediments, these facies are identified by higher bulk density, low organic matter content, and increased magnetic susceptibility. Near-shore, higher energy shallow water facies (<2 m) are identified by silt- to sand-sized fractions and small gastropod shells. Aquatic macrophytes (Myriophyllum sp.) are abundant in low energy littoral environments at Suches (Figure 4b). Accumulations of these plants as peat in core C-09 indicate stable shallow water conditions (⩾2–3 m). Deeper water facies are dominated by fine clay–silt fractions, including authigenic diatomaceous sediments, and lower magnetic susceptibility and organic matter content. Spot sampling for diatoms confirmed facies interpretations, and showed epiphytic (Epithemia, Rhopalodia), tychoplanktonic, and benthic (Cyclotella, Cocconeis) species in shallow, intermediate, and deep waters, respectively.
To more clearly distinguish depositional environments, scaled proxy data were rotated through PCA using the R Ggplot package. The loadings for the first and second principal components, as well as the eigenvectors for the sedimentological proxies, are shown in Figure 4c.
Chronological control
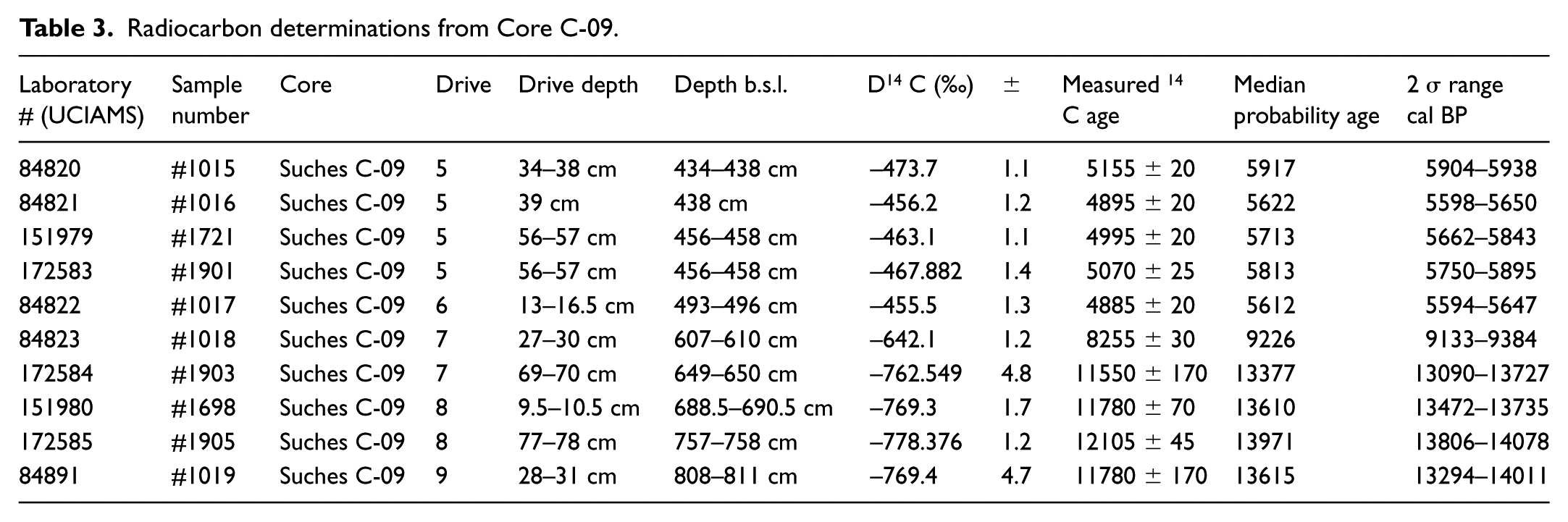
Facies age estimates are based on 10 calibrated AMS 14 C determinations. Sediments were disaggregated with 7% H2O2 and wet sieved through a 63-μm screen prior to picking materials for AMS 14 C dating. Preferred materials were carbonized stipules, seeds, and stem/leaf fragments of terrestrial grass or woody tissue. Suitable material was abundant in depths shallower than 4.96 m beneath WSI and was sporadically present in lower depths. Sample material was pretreated with a standard acid–base–acid treatment (Abbott and Stafford, 1996). Age determinations were measured at the Keck Carbon Cycle AMS Facility at the University of California, Irvine. Sample isotope fractionation values (δ13C = 0) were measured on prepared graphite samples. The resulting radiocarbon ages were calibrated using the IntCal 13 curve in CALIB v 7.0 (Reimer et al., 2013; Stuiver and Reimer, 1993), and the 2σ median probability age ranges are reported in Table 3 as years before present, calibrated (cal years BP, abbreviated below as kyr BP). Age–depth relationships and sediment accumulation rates were modeled using the BChron package in R (Ihaka and Gentleman, 1996; IHaslett and Parnell, 2008), and the 2σ range for modeled ages is shown graphically in Figure 4d.
Radiocarbon determinations from Core C-09.
Documentation of human occupation
Suches’ human history is reconstructed from two archeological surveys, the Proyecto Arqueologico Lago Suches (PALS) and Proyecto Arqueologico Qawra Thaki (PAQT; Stanish et al., 2010; Vining, 2011). Both surveys systematically inventoried all archeological loci encountered along pedestrian transects spaced ~15 m apart, for a total 2.8% random sample of all environments in the basin beneath 4900 m a.s.l. (Figure 7). Loci were classed as sites if they showed evidence of repeated use, or isolated finds if they reflected single-episode events. The PALS used a component-based system, where surface finds were cataloged in 10 m2 units to estimate the size and density of occupations during each archeological period. The age of components was inferred from diagnostic artifacts (Figure 8, see Supplemental Material, available online). We assigned absolute ages based on published typologies and radiocarbon-dated contexts at Quebrada Tacahuay and Asana, as they are closest to Suches (Aldenderfer, 1998; Keefer et al., 1998; Klink and Aldenderfer, 2005; Ravines, 1972; Tripcevich, 2010: Figures 3–10). Reported ages for diagnostic types are precise to ~100-year intervals (Klink and Aldenderfer, 2005), commensurate with the centennial-scale climatic reconstruction presented here. Published ages for these contexts were calibrated to the same IntCal13 curve used for the core chronology. Results of these surveys are provided in the Supplemental Material, available online.
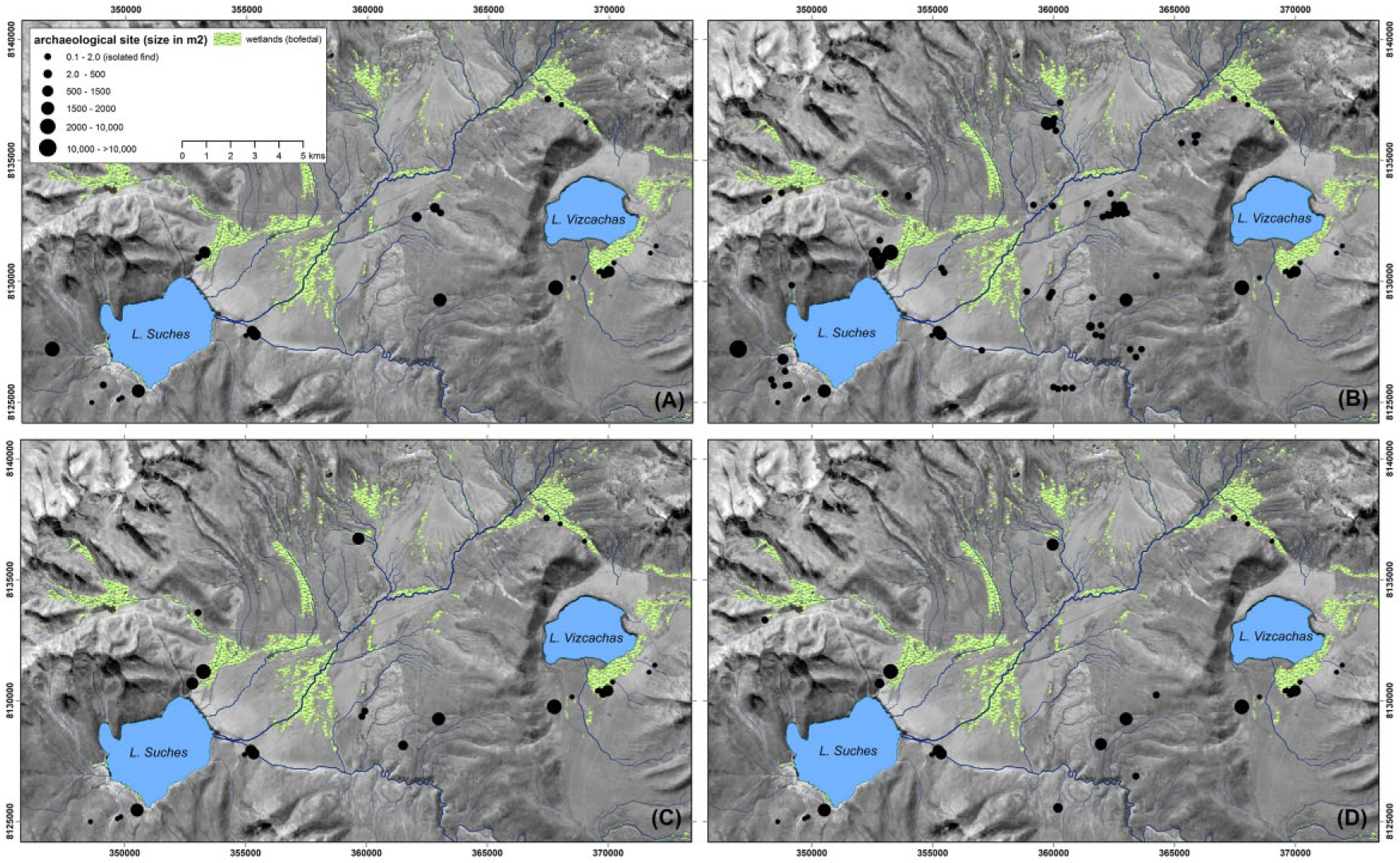
Archaic period settlement patterns in the Suches basin by subphase. (a) Early Archaic (11.2–9.0 kyr BP). (b) Mid-Archaic (9.0–7.0 kyr BP). (c) Late Archaic (7.0–5.4 kyr BP). (d) Terminal Archaic (5.4–3.9 kyr BP). Data from the PALS and PAQT surveys. Coordinates are in UTM Zone 19 S, WGS84.
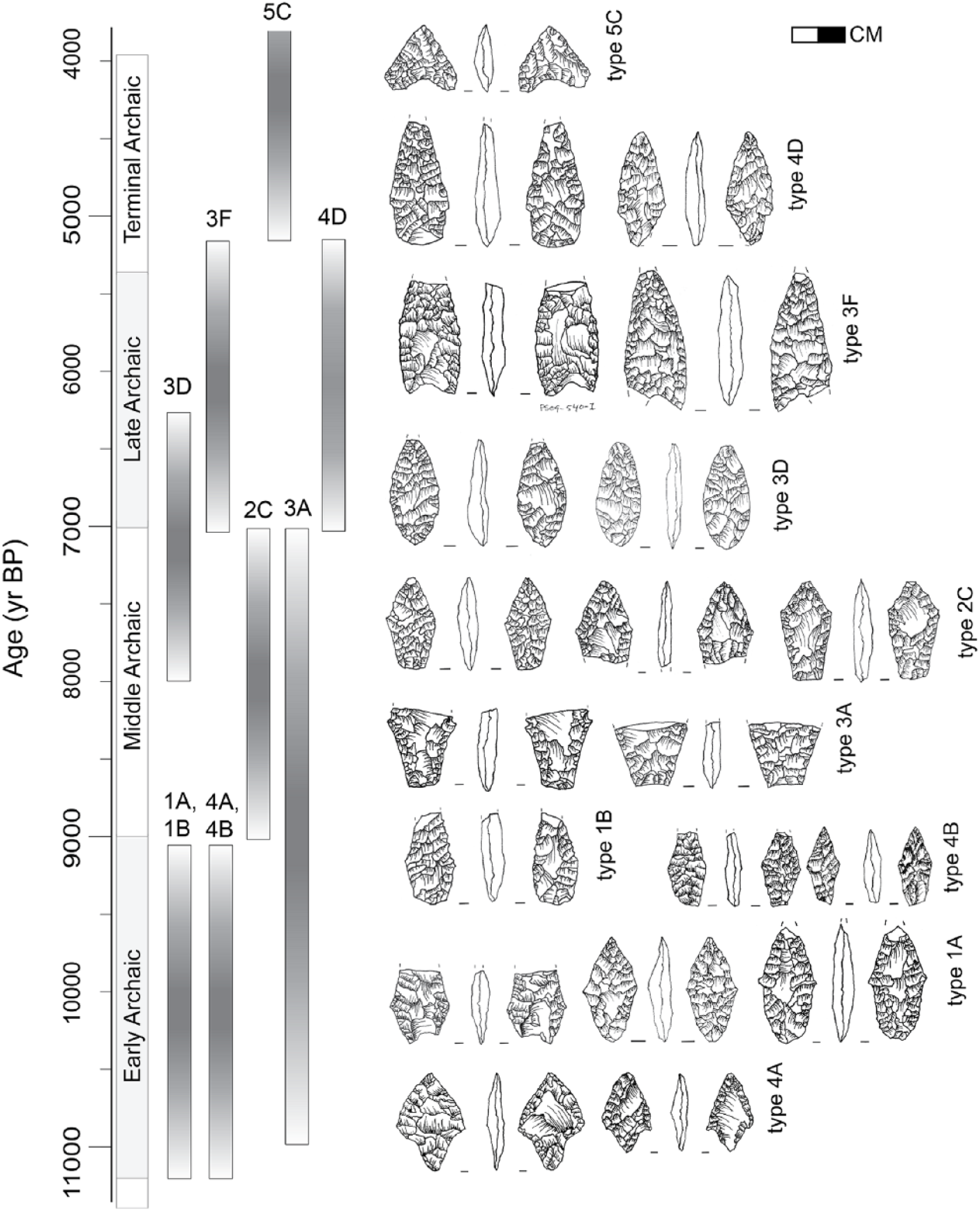
Diagnostic artifact types used to establish the chronological affiliation of archeological components. Archaic period chronology is based on 14 C determinations in Aldenderfer (1998), calibrated to the IntCal13 curve.
The relative intensity of occupation during any given period was reconstructed by summing the cumulative frequencies of diagnostic artifacts within 200-year bins (Figure 9). This follows a similar logic as using cumulative radiocarbon probability distributions (CPD) to reconstruct population histories (Rick, 1987; Shennan and Edinborough, 2007; Williams, 2012). Our use of diagnostic types adheres to the assumption that more age-dependent types will be generated by greater densities of people, and vice versa. While CPD-based population histories are subject to criticisms (e.g. Contreras and Meadows, 2014; Surovell and Brantingham, 2007), we believe they have merit as relative measures of population when used conservatively and when corroborated by other metrics.
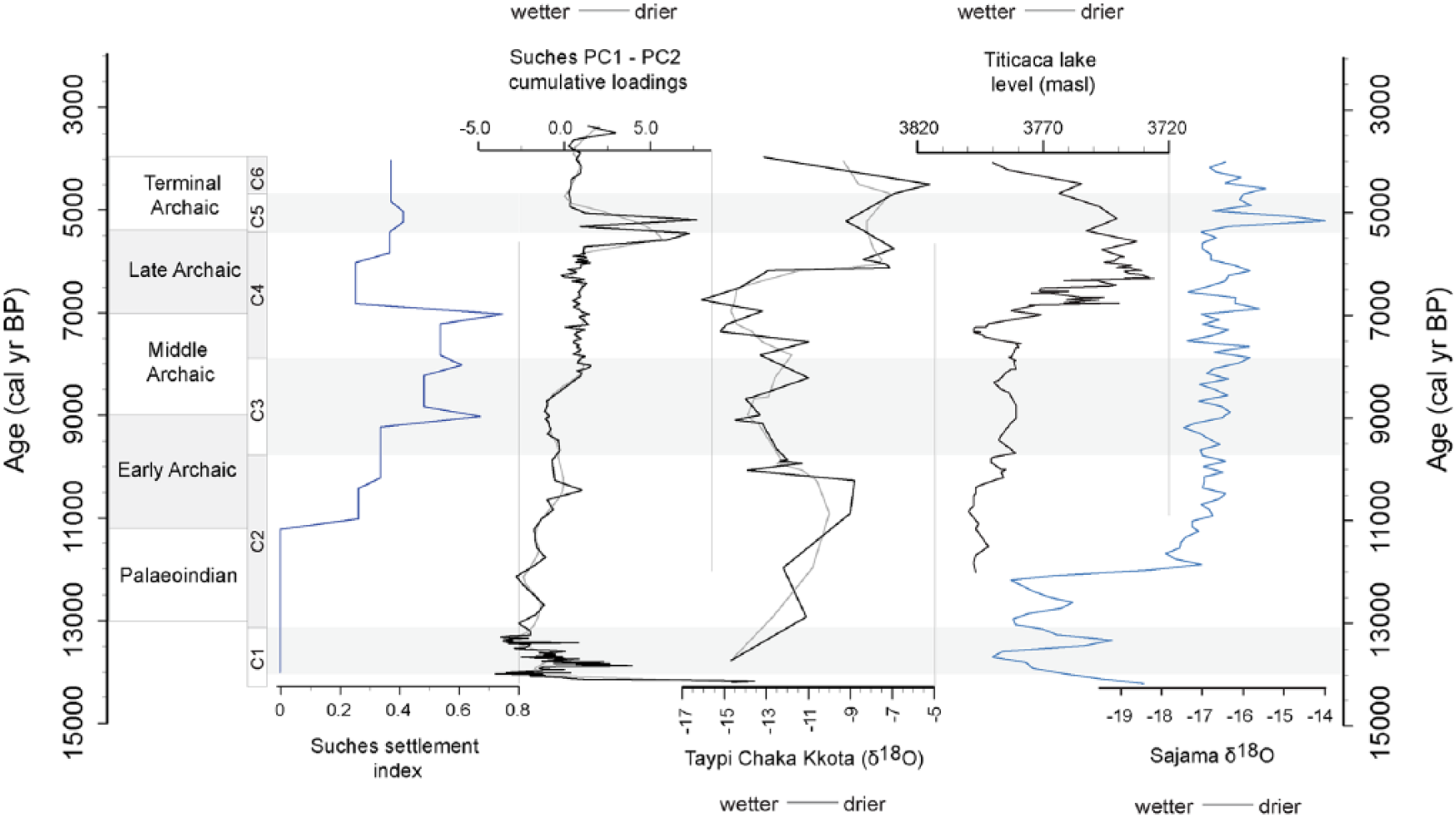
Comparison of settlement intensity index against Suches cumulative PC1 and PC2 loadings (representing approximately 73.4% cumulative variance in the analyzed proxies). Also shown are Laguna Taypi Kkota carbonate δ18O values (Abbott et al., 2003), reconstructed Titicaca lake level, and Sajama δ18O values (Thompson et al., 1998). Suches stratigraphic units (C1–C5) and Archaic-period cultural phases are provided for reference.
We normalized the observed frequencies of individual types against the variable lengths of each cultural phase during which each type was used, while preserving the cumulative frequencies of contemporaneous types. This was accomplished by dividing the frequency of each type by its chronological span (assuming a normal distribution across that age span) and then summing the total frequencies of all diagnostic types within 200-year bins. The resulting cumulative frequency curve, showing the number of diagnostic artifacts per 200-year period, is plotted in Figure 9 against the Suches and regional paleoclimatic reconstructions. The trend shown in Figure 9 is echoed in the frequencies of Archaic period components and their sizes (see Supplemental Material, available online), suggesting that it reflects a pattern of occupational intensity.
Results: Paleolimnological analysis
Isotopic composition of lake and source waters
Isotopic δ18Olake and δ18Obofedal values plot along a local evaporation line (LEL), which is substantially offset from the estimated local meteoric water line (LMLW, Bowen and Revenaugh, 2003). The isotopic enrichment of lake water compared with its source (>4‰ for 18O and >25‰ for D) indicates evaporation is a strong influence on water budgets (Gat, 1996), and premodern lake levels are strongly controlled by the P/E balance. Appreciable evaporative enrichment also occurs in bofedales and streams within the watershed, which we attribute to prolonged residence time of water in wetlands. Prolonged residence time mitigates fluctuations in atmospheric moisture on seasonal–interannual scales.
Core C-09 age–depth models
The age–depth models for Core C-09 are shown with the positions of 14 C determinations and the 2σ confidence interval in Figure 4d. Sediments date from the late Glacial ca. 14.0 kyr BP to the middle-Holocene, ca. 5.0 BP. Deposition succeeding the deposition of ice-proximal sediments was rapid (0.1–0.2 cm/yr) and then slowed to approximately 0.01 cm/yr between 13.0 and 9.5 kyr BP. Sedimentation rates remained low during the middle-Holocene from approximately 9.5 to 6.0 kyr BP. A pulse of sedimentation ca. 5.5 kyr BP and age reversals at the top of the core suggest dynamic lake levels and reworking between 6.0 and 5.0 kyr BP.
Sedimentological facies and lake levels
Core C-09 captures three broad phases of changing lake environments from the late Pleistocene to middle-Holocene. Changes in lake status are signaled by sedimentological shifts from glacial, peat-dominated, and allochthonous and authigenic sediments; carbonate throughout the sequence represents <10%, yielding insufficient material for δ18O analyses. Sedimentological proxies and derived principal components indicate a two-stepped shift toward a drier middle-Holocene climate (Figures 5 and 6). Despite evidence of drying, however, sedimentary proxies indicate that depositional environments remained relatively constant throughout most of the MHAP and lake levels were at/near mean modern shore lines. We summarize the principal depositional environments below.
Late Glacial lake formation and ice retreat (>812 cm, ca. >14,100 cal BP)
Basal sediments in core C-09 capture the late Pleistocene formation of the lake as glaciers retreated around 14.1 kyr BP. Sediments overlying the basal deposits are well-sorted, upward-fining glacial mineral sands containing abundant horneblende, biotite, feldspar, and quartz, with relatively high MS, DBD, and inorganic content. Upward fining and at least two rapid transitions from coarser to finer sediments suggest decreasing distal stream energy perhaps because of deglaciation and stabilizing periglacial environments.
Lake instability during the late Pleistocene (Unit C1, 664–812 cm, ca
13,400–14,100 cal BP).
Unit C1 captures a highly unstable postglacial lake environment. This phase of Lake Suches is characterized by predominantly organic sediments alternating with brief episodes of predominantly inorganic sedimentation. Despite fluctuations in lake environments, humid conditions prevailed and mean levels were higher than during the subsequent early–middle-Holocene. Sediments in the upper and lower portions of this unit are dominated by peat-like deposits of littoral aquatic macrophytes with minor laminae and discrete lenses of diatomaceous silts throughout. These sediments have low MS and DBD. Organic matter content in this unit fluctuates greatly, between 20% and 80%.
Between 730 and 755 cm b.s.l. (13.8–13.9 kyr BP), there is an interlude characterized by massive to laminated, glacial clays that are light gray to brown in color and have low organic content, higher MS, and dramatically increased DBD. Two beds of organic-dominated sediments intercalate with the clays, indicating reversals in depositional environments. This interlude represents increased glacial input into the lake. Similar glacial fluctuations are found in Andean lakes between ca. 14.5 and 10.9 kyr BP; glacial advance/retreat cycles between ~12.5 and 11.7 kyr BP are attributed to the Younger Dryas, while mechanisms driving earlier cycles are less clear (Jomelli et al., 2014; Mahaney et al., 2008; Rodbell and Seltzer, 2000; Rull et al., 2010; Seltzer et al., 1995; Stansell et al., 2010). Following this interlude, the alternation between organic- and inorganic-dominated sedimentation resumes.
Stepwise drying during the terminal Pleistocene–early Holocene (Unit C2, 613–662 cm, ca
9700–13,400 cal BP).
Sediments from approximately 9.7–13.4 kyr BP record a prolonged and stepwise progression toward drier early Holocene climate. Depositional environments alternate between peaty-humic and inorganic clay–fine silt sediments in couplets of similar (but decreasing) durations, and sedimentation rates slow markedly (Figure 6). Proxy values fluctuate widely, but the amplitude of these fluctuations gradually decreases over time. Unit C2 represents the gradual replacement of organic-dominated deposition with inorganic deposition. Organic matter decreases from its maximum of 95% at the Unit C1–C2 transition to a low of 17% by the end of C2. Inorganic sedimentation continues to dominate the Holocene portion of this core. Only two significant episodes of organic-rich deposition occur after Unit C2, in contrast to the organic-dominated late glacial sequence. By the end of this phase, lake environments are much more stable and this comparative stability characterizes Lake Suches for much of the subsequent early–middle-Holocene. This overall drying trend is interrupted by two centennial-scale deviations. The first, approximately 11.9–12.2 kyr BP is marked by a pronounced change to peat-dominated sediments with markedly higher organic matter (60%) and lower carbonate (2.6%) and mineral matter (~37%) content, which we interpret as a more humid episode. The second interval occurs near the Pleistocene–Holocene boundary, at 10.25–10.6 kyr BP. An abrupt, high-magnitude increase in magnetic susceptibility corresponds with increased DBD, carbonate, and mineral matter and decreased organic matter, suggesting a higher energy, near-shore depositional environment during lower lake levels. We interpret this as indicating increased allochthonous deposition into shallow littoral waters during an arid interlude. Following this event, the trend toward drier Holocene climates resumes, although similar arid/erosive conditions do not reoccur until late in the middle-Holocene.
A moderately humid early Holocene (Unit C3, 582–613 cm, ca. 8160–9670 cal BP)
The early Holocene marks the establishment of more stable lake conditions. Moderate increases in organic matter (~20–25%) and low carbonate (2–4%) and DBD indicate marginally organic dominated deposition suggesting slightly more humid conditions than the preceding terminal Pleistocene and the succeeding middle-Holocene. Magnetic susceptibility and carbonate content show low-amplitude, high-frequency variability, however, which may indicate abbreviated pulses of terrigenous sedimentation during drier episodes within this generally more humid phase. Despite these signals, proxies waver around a stable mean state. Hyperarid conditions are noted in the Atacama and Titicaca basin by ca. 8.0 kyr BP. There is a hiatus at this point in the Suches proxies ca. 8.2–8.6 kyr BP, possibly because of lake-level drawdown, and evidence of a drying environment appears shortly afterwards.
A drying mid-Holocene environment (Unit C4, 438–562 cm, ca. 5500–8160 cal BP)
The period between 5.5 and 8.0 kyr BP is differentiated from the preceding intervals by a stepwise average increase in carbonate, an increase in DBD, and declining (increasing) organic (residual mineral) matter content. Sediments during this period are predominantly silty-clays to coarse sand. Transitions between lithostratigraphic units are graded, rather than abrupt, and diatomaceous silts and gastropod macrofossils appear for the first time in the sequence. A spike in proxies signals an arid interlude ca. 7.2 kyr BP, with drying conditions after 6.4 kyr BP. This signals the onset of middle-Holocene aridity no later than 7.2 kyr BP. The lack of abrupt erosional surfaces and biogenic sediments suggest that lake levels were sustained despite more arid conditions. Peat-dominated deposits 480–490 cm b.s.l. (ca. 6050–6200 kyr BP) suggest increasing aridity was interrupted by a humid interlude.
Peak mid-Holocene hyperaridity (Unit C5, 427–437 cm, 4750–5450 cal BP)
Maximum aridity occurs at Suches late in the middle-Holocene, ca. 4.8–5.5 kyr BP. This arid phase is marked by diatomaceous lenses intercalated within fine silty-clay, and dramatic peaks in carbonate (48–60%) matched by decreases in magnetic susceptibility, organic matter, and residual mineral content. We interpret the anomalously high carbonate content and diatomaceous sediments as linked to evaporitic deposits under hyperarid conditions. Pueyo et al. (2011) similarly note increased precipitation of endogenic carbonate in Lake Chungará coinciding with middle-Holocene aridity, which they relate to blooms of halophytic microfauna. Abrupt sedimentary contacts and minor age reversals suggest erosional surfaces and reworking of material with dropping lake levels. Low organic matter, mineral content, and magnetic susceptibility suggest limited littoral macrophyte productivity as well as decreased inputs of terrigenous sediments, because of limited sediment transport under drier conditions.
Arid conditions in the latter middle-Holocene (Unit C6, 402–426 cm, 3200–4750 cal BP)
Conditions similar to those encountered during Unit C4 are established in C6. This shows an arid environment, though not as severe as during the preceding period of increased aridity ca. 4.7–5.5 kyr BP. Unit C6 ends with an influx of terrigenous sedimentation, likely from an erosive episode, marked by peaks in MS, DBD, and inorganic material and low organic matter/carbonate content.
Results: Documentation of early–terminal Archaic occupation
Early–middle-Holocene human population dynamics are inferred from a total of 113 archeological find-spots dating from the early to terminal Archaic (see Supplemental Material, available online). Because of differences in survey strategies, it is not possible to estimate the size or density of Archaic phases recorded by the PAQT beyond simple presence/absence. Despite this, both surveys recorded statistically similar profiles of sites (χ2 = 13.148, df = 19) and scores for locational preferences (t = −1.57, df = 71, p = 0.05), indicating that the spatiotemporal distributions of each survey sites recorded are coherent.
Initial colonization of the Suches Highlands
The earliest diagnostic artifacts currently documented place the initial colonization of Suches in the early Archaic period (EA), ca. 11.0–9.8 kyr BP, contemporaneous with the Asana I-II phases and coinciding with the dramatically more stable and drying conditions of the early Holocene (Figure 8). The EA occupation focused on large open-air sites along the open margins of the basin, in close proximity to modern river channels, lake shores, and, significantly, bofedales. This last relationship suggests that coupling between human land use and high-elevation Andean wetlands has an early inception. Compared with the subsequent Middle Archaic (MA), EA occupation is light and occurs as fewer, larger sites (mean = 0.41 ha), complemented by periodic but light use of upland areas (Figure 7).
MHAP settlement intensification and decline
The period from ca. 9.0–7.0 kyr BP coincides with the Atacama settlement hiatus (silencio arqueologico). Rather than a hiatus, however, the MA is one of the most important archeological periods in Suches. There is a marked increase in the number and density of MA find-spots and diagnostic types, signaling a dramatic increase in human settlement (Figures 7 and 9). MA components are more numerous but smaller in size than EA components, suggesting proliferated, less nucleated populations or patterned mobility. Marked increases in the number of isolated finds similarly indicate more intensive but ephemeral use of upland areas. Relationships between site size and location reveal an emergent bimodal settlement pattern. Larger open-air sites occur on the pampas and valley floors. These are 0.3–0.4 ha elongated scatters conforming to the margins of extant or fossil bofedales. A suite of smaller open-air sites (0.1–0.9 ha) are located in upland drainages, often on alluvial terraces above incised channels that are dry under modern conditions. Both classes of sites suggest a strengthening of relationships with localized water bodies at the onset of the MHAP.
Occupation declined notably during the subsequent Late Archaic, ca. 7.0–6.0 kyr BP. This hiatus in Suches postdates the start of the Atacama silencio arqueologico by approximately one millennium but coincides with maximum lowstands of Lake Titicaca and the onset of drying conditions around Suches. Despite indications of less occupation than in the preceding and subsequent periods, occupation was not as low as it was during the basin’s initial colonization. There was not a complete abandonment of the basin, consequently, but a contraction toward highly localized microrefugia. Late–terminal Archaic period settlement was more strongly concentrated along the edges of pampas and bofedales. Furthermore, increased frequency of diagnostic types postdating 6.0 kyr BP suggests a modest population recovery (Figure 9). In the subsequent terminal Archaic (5.4–4.0 kyr BP) site sizes remained small but increased in number, and there is an increase in the frequencies of terminal Archaic diagnostic artifacts. Both increases coincide with lacustrine evidence for peak aridity in Suches, and may represent endemic population growth or another influx of populations into the basin coinciding with regional aridification.
Discussion
Paleoclimatic and settlement dynamics in Suches generally mirror regional patterns. Yet, Suches sediment proxy data indicate that near-modern lake levels persisted throughout much of the middle-Holocene climatic deterioration, and the small lake is notable for having sustained higher lake levels than larger central Andean lakes did, in particular. We attribute this to the extensive wetlands found in the Suches basin and their close hydrological relationship with the lake. Sustained water in Lake Suches has parallels with persistent humidity recorded in other central Andean wetlands (Ledru et al., 2013; Quade et al., 2008; Servant and Servant-Vildary, 2003; Servant-Vildary et al., 2001). As an effect, Suches’ hydroclimatic response to climatic inversions is muted, and MHAP aridity that resulted in severe drawdowns of Lake Titicaca and other central Andean lakes had weaker effects on Suches. Thus, the Suches region functioned as a refugium, which enabled human occupation to persist throughout the MHAP.
Suches in a regional climatic context
Suches exhibits unstable but generally higher lake levels during the terminal Pleistocene Tauca phase, ca. <13–11.0 kyr BP. Titicaca archives indicate cooler (~5°C less than modern) and wetter (20% increase in precipitation) conditions during this period (Cross et al., 2001), as do many Atacama records (Geyh et al., 1999; Grosjean et al., 2001, 2003; Latorre et al., 2003; Valero-Garcés et al., 2003), while small alpine lakes in the Titicaca catchment formed under humid conditions (Abbott et al., 2003).
Early–middle-Holocene aridification is apparent in central Andean lacustrine and alpine glacial ice isotopic records, but the mechanisms and timing remain controversial. The timing and severity of peak aridity during the MHAP, in particular, is poorly constrained. Alpine glacial ice records indicate aridification ca. 9.0–3.4 kyr BP at Sajama; 12.0–5.0 kyr BP at Illimani; and 8.4–5.2 kyr BP at Huascarán (Ramirez et al., 2003; Thompson et al., 1995, 1998, 2000).
Drier conditions appear in Titicaca archives ca. 7.5–3.5 kyr BP, with estimates of peak aridity between 6.0 and 5.0 kyr BP (Paduano et al., 2001; Rowe et al., 2002). There is evidence for overflow conditions in the Wiñaymarka sub basin until ca. 8.3–8.0 kyr BP (D’Agostino et al., 2002; Rowe et al., 2002; Tapia et al., 2003). More arid than modern conditions are indicated until ca. 4.0–3.2 kyr BP, when lake levels rose rapidly (Abbott et al., 1997b; Grove et al., 2003). During peak aridity, Titicaca lake level dropped an estimated ~85 m beneath its modern outlet, while the smaller Wiñaymarka sub basin completely desiccated (Argollo and Mourguiart, 2000; D’Agostino et al., 2002; Mourguiart et al., 1998). Rowe et al. (2002) propose an age of 7.25–6.25 kyr BP for the Titicaca lowstand, with low water conditions until ca. 4.0 kyr BP. Microfossil assemblages, sedimentary δ87Sr, and charcoal concentrations indicate peak negative mass balance and salinized conditions ca. 6.0–4.0/3.5 kyr BP, interrupted by several centennial-scale arid/humid inversions (Cross et al., 2001; Grove et al., 2003; Mourguiart et al., 1998; Paduano et al., 2001; Tapia et al., 2003). Two ‘major’ arid events are indicated ca. 5.5 and 4.5 kyr BP by saline-tolerant diatoms and ostracod Mg/Ca ratios (Mourguiart et al., 1998; Tapia et al., 2003). Downcutting concurrent with Titicaca lowstand is found in the lake’s Rio Ilave and Rio Ramis tributaries (Farabaugh and Rigsby, 2005; Rigsby et al., 2003). These events coincide with peak aridity indicated in Suches unit C6, ca. 4.8–5.5 kyr BP.
Lacustrine archives from the Atacama region support the establishment of arid conditions ca. 8.0–3.6 kyr BP, following late Glacial–early Holocene humidity (Geyh et al., 1999; Grosjean et al., 2003). The MHAP was sufficiently severe to cause widespread human abandonment of the Atacama region (with the exception of a few ‘pronounced ecological refuges’) ca. 9.0–6.0 kyr BP (Grosjean et al., 2001, 2005; Moreno et al., 2009; Núñez et al., 2002, 2010, 2013). However, controversial arguments have been made for middle-Holocene humid phases (e.g. Latorre et al., 2003; Placzek et al., 2001; Quade et al., 2008).
Generally, paleowetlands mirrored MHAP drying trends. However, most wetland archives show the persistence of locally humid conditions, despite the desiccation of larger systems. Rios Chiarhajuiara and Chuqiaguillo paleowetlands show sustained moisture during the MHAP onset (Servant and Servant-Vildary, 2003). Sediments and aerophilic diatoms indicate stable, shallow-water conditions in Rio Baja wetlands from 8.0 to 6.3 kyr BP, followed by a humid interlude 6.3–5.3 kyr BP, until saline-tolerant diatoms indicate drying conditions 5.3–3.1 kyr BP (Servant-Vildary et al., 2001). Tiquimani wetland records show increased humidity, 5.8–4.0 kyr BP (Ledru et al., 2013). Paleowetlands persisted at Parko Kkota (at Titicaca’s outflow) during peak aridity, ca. 4.7 kyr BP (Baucomb and Rigsby, 1999). Evidence for persistent MHAP paleowetlands is also found in Quebradas Puripaca and Tulan in the Atacama region (Betancourt et al., 2000; Grosjean et al., 2001; Núñez et al., 2013; Rech et al., 2003).
Suches sediment proxies indicate it was in-phase with other records, yet the impact of MHAP aridity was not as severe. The MHAP is evident from approximately 8.0 to 3.0 kyr BP, with increased aridity after 5.8 kyr BP. This chronology is consistent with MHAP records from Lake Titicaca and Sajama (Figure 9). However, Suches sustained near-modern levels through much of this period. Fluctuations in lake status are short-lived and return rapidly to a mean stable state. The severe Lake Titicaca low stand that dropped Lago Chuchuito levels by ~85 m and desiccated Lago Wiñaymarka is not apparent. Erosional facies and major changes in lake status occur only during the most severe arid episodes, ca. 5.4–4.8 kyr BP and lacustrine sediments are present throughout the sequence.
Sustained moisture in Lake Suches during the MHAP
We hypothesize that sustained lake levels and human occupation in Suches are because of more positive P/E balances caused by interplays between four factors: (1) strong coupling between the lake’s hydroclimate and extensive bofedales, (2) watershed hypsometry and relationship to local hydrology, (3) the lake’s geographic position relative to atmospheric circulation systems, and (4) a weakened middle-Holocene SASM pattern that contributed to lower net but seasonally better-distributed precipitation (e.g. Ledru et al., 2013; Servant and Servant-Vildary, 2003). Each is discussed below in more detail.
Suches’ response is more consistent with Titicaca and Atacama wetlands records than it is with lake-level records. We attribute this to strong coupling of Suches’ hydroclimate to moisture balance in bofedales within the catchment. While these wetlands represent an estimated 2.5% of central Andean land cover (Kuznar, 1995), they comprise approximately 6% of the land cover in Suches and may have provided an extensive reservoir that buffered Suches hydroclimate from short-term atmospheric variability. Locally sustained moisture is documented in several paleowetlands, but the mechanisms contributing to this remain poorly understood (Grosjean et al., 2001; Quade et al., 2008; Rech et al., 2003). Mature bofedales have thick (>2 m) histosols, lower permeability, and greater water retention capacity than the sediments over which they develop. Autoregulation of internal hydrology additionally may be important (Earle et al., 2003; Squeo et al., 2006). Remote sensing moisture indices indicate that available water is sustained in bofedales despite seasonal–decadal dry phases (Vining, 2016). This is supported by isotopic enrichment of δ18Obofedal, indicating prolonged water residence times. Large amounts of groundwater are likely also stored in the extensive fluvioglacial fans surrounding the lake.
Three factors related to Suches’ topographic position result in more favorable P/E balances under modern conditions. First, estimated modern P/E balances indicate Suches’ high elevation results in cooler temperatures and less evaporation than other portions of both the Titicaca and Atacama catchments (Figure 1b). Second, higher elevations surrounding the basin promote orographic cloud formation and localized precipitation. Third, Suches is located at the head of regional catchments. Similar headwater lakes exhibit less pronounced biogeochemical responses to dry phases than do lakes in distal portions of regional flow systems, such as Titicaca, which depend on stream- and groundwater discharge and have greater potentials for evaporitic and through-flow loss (Kratz et al., 1997; Webster et al., 1996).
Changes in the source, type, and seasonal distribution of MHAP precipitation, rather than total amount, may have resulted in locally variable moisture balances. Multiple proxy records suggest a weakened middle-Holocene SASM until after 4.0 kyr BP (Abbott et al., 2003; Bird et al., 2011a; Haug et al., 2001; Jomelli et al., 2011; Vuille et al., 2000). A weakened SASM corresponds to weakened Amazonian convection, a decreased portion of moisture derived from Atlantic air masses, and overall more arid conditions. Microfaunal δ87Sr corroborates that increased amounts of precipitation reached Lake Titicaca from northerly/westerly portions of the basin during the MHAP than occurs under modern conditions, while lake-effect and Amazonian convection weakened (Grove et al., 2003). Paleowetland records further suggest net decreased MHAP precipitation occurred as smaller, non-stormy events that were more frequent and seasonally well-distributed throughout the austral summer and winter than occurs under the modern SASM (Servant and Servant-Vildary, 2003; Servant-Vildary et al., 2001). Combined with glacial sources and greater soil moisture capacities, seasonally well-distributed precipitation may have maintained wetland water tables even as decreased net precipitation and increased temperatures contributed to MHAP aridity (Ledru et al., 2013; Servant and Servant-Vildary, 2003).
Refugia formation in the Suches watershed during the MHAP
During the MHAP groundwater inputs from wetlands and small lakes like Suches had sustaining effects on foraging populations. Archeological evidence indicates a significant increase in human activity in the Suches watershed beginning ca. 9.0–7.0 kyr BP, and sites are increasingly tethered to wetlands and other groundwater sources throughout the MHAP (see also Cipolla, 2005; Keefer et al., 1998; Klink, 2005; Kuznar, 1989). The density of MA sites in Suches averages 2.36 sites per km2. This is approximately half the density of contemporaneous settlement in the nearby Rios Huenque and Ilave valleys, which average 4.6 sites/km2 (Craig et al., 2010; Klink, 2005). This difference is likely influenced by survey strategies, as the former surveys preferentially focused on river and wetland margins where archaeological sites are commonly found. MHAP settlement in Suches is approximately 10 times denser than that of the Lake Titicaca littoral region, which was largely abandoned at this time (Albarracín-Jordan and Mathews, 1990; Bandy, 2001; Capriles and Albarracin-Jordan, 2013; Cipolla, 2005; Kuznar, 1989; Stanish et al., 1997). Similarly, there is a hiatus in the nearby Andean sierra and coastal regions, in addition to the Atacama silencio arqueologico (Aldenderfer, 1998; Núñez et al., 2013; Wise, 1989).
There is a dramatic inflection in settlement intensity ca. 7.0–5.9 kyr BP, after which populations rebound (Figure 9). This event coincides with evidence for increased drying in Suches proxies, as well as peak aridity in Lake Titicaca. Deteriorating conditions in the latter MHAP likely had adverse effects on Late Archaic groups. Water-tethered settlement preferences strengthen as groups moved toward stable wetlands and lakeshore environments. The increase in terminal Archaic components ca. 5.2 kyr BP suggests populations recovered in Suches, even as peak hyperaridity persisted in Lake Titicaca. Several severe but punctuated arid events characterize Suches lake status ca. 5.5–3.5 kyr BP. Despite these extreme arid events, terminal Archaic groups appear to have found favorable environmental refugia around Suches.
High-elevation Andean microrefugia
Paleolimnological and archeological data indicate that Suches served as an MHAP refugia because of an interplay of factors: the Titicaca catchment, regionally, offered more amenable conditions and acted as a macro-refugium, unlike regions to the south (the Atacama) or at lower elevations (the inter-Andean Pacific valleys). MHAP human occupation further relied on discrete microrefugia found within the Titicaca macroregion. These included higher elevation wetland microenvironments, where a constellation of factors contributed to more humid, temperate environments amenable to human occupation. The persistence of humid conditions and human settlement largely undisturbed throughout the MHAP attests to the importance of high-Andean refugia for mitigating climatic variation (cf. Valencia et al., 2016).
Bennett and Provan (2008) provide a useful overview of paleoecological refugia types that illuminate MHAP Andean patterns. ‘Refugia’ describes definable, typically isolated environmental pockets that are insulated from regional conditions and consequently allow populations to persist during periods of adverse climate. Refugia are defined by changing distributions and abundances of species on long-term timescales consistent with enviro-climatic changes. A species’ dispersal ability is significant, and affects whether refugia are in situ (fragments of a taxa’s prior distribution) or ex situ (new, formerly unoccupied regions; Ashcroft, 2010). As species contract toward refugia, former ranges reduce in extent. Species abundances may increase locally, causing uneven abundance across an overall range as species concentrate into favorable refugia.
In Bennett and Provan’s classification, classic-, cryptic-, and elevational types describe elements of MHAP Andean refugia. In classic refugia types, populations descending from ancestral subsets contract to more temperate regions, subsequently rebounding when favorable conditions return (cf., ‘macro-refugia’ in Ashcroft, 2010). These differ from cryptic or micro-refugia, where taxa survive deteriorating conditions in highly localized microclimates and/or at lower densities over their former range (Ashcroft, 2010; Bennett and Provan, 2008; Stewart et al., 2010; Stewart and Lister, 2001). High-elevation regions are pivotal in shaping cryptic refugia. Paleobiogeographical research suggests elevational refugia are strongly linked to temperature (Kessler et al., 2014; Klimeš and Doležal, 2010). Temperature gradients coupled to lapse rates translate to altitudinal shifts in warm-/cold-adapted species as climates oscillate between cooling/warming phases. However, moisture availability is also coupled with lapse rates and orographic forcing, and moisture availability is a strong limiting factor in the arid–semiarid central Andes. The development of refugia in Suches can be attributed to the regionally more favorable conditions of the Titicaca/Altiplano system (‘classic’ refugia), as well as the particularly favorable microenvironments of high-elevation wetlands (showing elements of ‘cryptic’ and ‘elevational’ refugia).
Conclusion
Paleoclimatic and archeological data from Suches show coupled histories of increasing aridity but intensifying human settlement during the middle-Holocene. Sedimentological proxies indicate that glaciers retreated from the catchment around 14.0 kyr BP. This was succeeded by an initially humid late Pleistocene, with a stepwise transition to drying conditions by the early Holocene. The initial human occupation of Suches dates to the EA period, ca. 11.0–9.8 kyr BP as drier but more stable early Holocene conditions were established. Middle-Holocene aridity was established no later than 7.2 kyr BP, with maximum aridity ca. 5.5–4.8 kyr BP. While paleoclimatic patterns parallel prior paleoclimatic reconstructions of severe middle-Holocene aridity in the central Andes, the magnitude of peak aridity is significantly lower at Suches. Archeological occupation appears to have had its maximum during the MA ca. 9.0–7.0 kyr BP, coinciding with widespread evidence for abandonment in the Atacama and Titicaca regions. Occupation declined notably during the subsequent Late Archaic, ca. 7.0–6.0 kyr BP, coinciding with paleolimnological evidence of more arid conditions. Yet the basin was never fully abandoned as were other regions. There is a settlement recovery in the late–terminal Archaic, 6.0–4.0 kyr BP that coincides with evidence for peak aridity in Suches and other archives in the Altiplano region (Figure 9). Archeological sites are water-tethered, with a strengthening relationship to bofedales in particular over time.
The persistence of wetter conditions in Suches may be related to a complex interplay of factors. Extensive bofedales and porous aquifers in the Suches basin feed the lake and buffer lake-level variability. Isotopic enrichment of δ18Olake and δ18Obofedal show that water in wetlands has a prolonged residence time, mitigating seasonal–interannual fluctuations in atmospheric moisture. Favorable elevation-linked P/E balances and hydrographic relationships in the alpine basin, including localized orographic precipitation, lower evaporation rates, and weakened monsoonal patterns, further mitigated moisture variability. While diverging from other lacustrine reconstructions, evidence for ameliorated MHAP conditions in Suches agrees with sustained moisture in Titicaca and Atacama paleowetlands (Baucomb and Rigsby, 1999; Betancourt et al., 2000; Grosjean et al., 2001; Ledru et al., 2013; Núñez et al., 2013; Rech et al., 2003; Servant and Servant-Vildary, 2003).
Paleolimnological and archeological evidence indicate MHAP drying had profound regional effects on foraging societies throughout the Altiplano and Atacama systems, but that high-elevation regions like Suches provided important ecological refugia. Evidence for refugial microenvironments is key to understanding the persistence of human populations during a climatically challenging period. It is likely that refugia utilized by people benefited other endemic Andean flora and fauna and were key to the region’s Holocene ecological development. Observed and predicted warming and decreased precipitation over the tropical Andes are once again posing challenges to water availability and usage (Bradley et al., 2006; Vuille et al., 2008). The dynamics of wetlands- and elevation-linked refugia will be key to anticipating and mitigating the effects of future adverse climatic conditions in the tropical Andes.
Supplemental Material
18-Suches_Supplemental_Material – Supplemental material for Paleoclimatic and archaeological evidence from Lake Suches for highland Andean refugia during the arid middle-Holocene
Supplemental material, 18-Suches_Supplemental_Material for Paleoclimatic and archaeological evidence from Lake Suches for highland Andean refugia during the arid middle-Holocene by Benjamin R Vining, Byron A Steinman, Mark B Abbott and Arielle Woods in The Holocene
Footnotes
Acknowledgements
We thank Alex Chepstow-Lusty, Mieke Stierken, Francesco Berna, Noa Corcoran-Tadd, and Kyle Stich for their assistance during the various stages of data collection and analysis. The ASTER L1B and SRTM data products were obtained through the online Reverb Data Pool at the NASA Land Processes Distributed Active Archive Center (LP DAAC), USGS/Earth Resources Observation and Science (EROS) Center, Sioux Falls, South Dakota. Precipitation data were accessed from the National Oceanic and Atmospheric Administration’s National Climate Data Center (NOAA-NCDC) and the United Nations World Meteorological Association (UN-WMO). Southern Copper Corporation provided access to Lake Suches, logistical support and climatic data. In particular, we thank José de Pierola, César Montes de Oca, and Hugo Valdiva at Southern Copper Corporation. La comunidades autonomas de Huaytire and Japopunco and individual land holders in the Suches region were key in supporting this fieldwork. Proxy analysis was conducted at the Sediment Geochemistry Laboratory at the University of Pittsburgh.
Funding
Field and lab research was funded by the National Science Foundation (Grants BCS-0900904 to Vining, AGS-1137750 to Steinman, and the P2 C2 Program to Abbott) and a Douglas C. Kellogg Award for Geoarchaeological Research from the Society for American Archaeology (Vining).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.