
Abstract
Sedimentological, palynological, and micropalaeontological studies carried out throughout the first half of the Holocene, during the Mesolithic/Neolithic transition in the Bay of Brest (i.e. 9200–9000 and 6600–5300 cal. BP) and in the Bay of Douarnenez (i.e. 9200–8400 cal. BP), allowed characterizing coastal environmental changes under the increasing influence of the relative sea-level rise. The gradual flooding of the two studied sites implied a transition from river valleys to oceanic bays as revealed by the gradual retreat of salt marsh environments, as detected through palynological analysis. In addition, these high-resolution studies highlight the regional imprint of the North Atlantic millennial climate variability in north-western coastal environments. Two cold climate events are indeed suggested to have been locally marked by a moisture increase, mainly detected by increases in Lingulodinium machaerophorum, Corylus, and Alnus percentages at 8550 cal. BP in the Bay of Douarnenez and at 6250 cal. BP in the Bay of Brest. Moreover, regarding the Neolithic transition timing in the Bay of Douarnenez, large pollen grains of Poaceae (i.e. Cerealia-type pollen grains) have been detected at around 8600 cal. BP, that is, 1500 years before the general accepted cereal cropping appearance in Western France. These results, consistent with other palynological studies conducted in the French Atlantic coast, could underline a Mesolithic ‘proto-agriculture’ in Brittany.
Keywords
Introduction
The Holocene is characterized by a gradual decrease in summer air temperatures and seasonality (seasonal thermal amplitude) in line with the continuing summer insolation decrease at 65°N (Berger and Loutre, 1991). Sub-millennial climate variability is superimposed on this long-term climate trend, as shown by numerous studies carried out on marine (e.g. Bond et al., 1997, 2001; Mayewski et al., 2004), terrestrial (e.g. Davis et al., 2003), and glacial records (e.g. O’Brien et al., 1995), highlighting iterated abrupt climate events also referred as ‘Bond events’ (Bond et al., 1997, 2001) or RCC (i.e. ‘Rapid Climate Change’; Mayewski et al., 2004). These events are often associated with significant climate cooling in North Atlantic surface waters and over northern Europe, as well as increasing aridity in the tropics (Hammarlund et al., 2005; Mayewski et al., 2004; Wanner et al., 2011). Finally, on decadal to multi-decadal timescales, climate over northern Europe is forced by the combined influences of atmospheric and oceanic natural oscillations, themselves driven by different teleconnections and physico-chemical exchanges at the air/ocean interface (Knight et al., 2006; McCarthy et al., 2015; Ruprich-Robert and Cassou, 2015; Tréguer et al., 2014).
The geographical exposure of the coast to the ocean and North Atlantic climate hazards thus makes the Northwest coast of France a favorite environment for the study of current and past climate. In addition to these factors, coastal areas are constantly changing shaped by different dynamic agents. The sea-level rise occurring across the deglaciation has also affected coastal environments from a geomorphological, sedimentological, and ecological point of view. On a regional scale, recent studies reconstructed and quantified the post-glacial sea-level rise on Brittany’s coasts (García-Artola et al., 2018; Goslin et al., 2013, 2015; Stéphan et al., 2015), estimated from about 10 to 15 mm/yr between the end of the last glacial period and 9000 cal. yr BP to around 4.6 mm/yr between 7500 and 6500 cal. yr BP, and less than 1 mm/yr after 6000 cal. yr BP. Moreover, palaeoecological data recently acquired on Holocene Brittany’s coastal sequences discussed past environmental variations (vegetation changes, palaeo-storm dynamics, precipitation regimes) and linked these rapid coastal changes with climate dynamics as well as human occupation on watersheds (Fernane et al., 2014, 2015). These studies especially focused on the Neolithic, a period when the development of agro-pastoral societies became more and more pronounced (Pailler et al., 2011; Visset and Bernard, 2006).
In this study, new sequences retrieved in two sites from the westernmost part of Brittany (NW France), the Bay of Brest (BB) and the Bay of Douarnenez (BD), cover the Mesolithic to Neolithic transition, a still fairly unknown period so far. They allow a multidisciplinary approach based on pollen, dinoflagellate cyst (dinocyst) and foraminiferal analyses, as well as on stable isotopes and sedimentological data. Pollen analysis provides information regarding surrounding vegetation changes on BB and BD watersheds through time. Indeed, palynological studies carried out on modern BB sediments (Lambert et al., 2017), as well as across the past 150 years (Lambert et al., 2018), evidenced the robustness of studying fossilized pollen grains in BB sediment archives to discuss both natural and anthropogenic forcings. These include hydrodynamics, fluvial discharges, pollination rates, or agricultural watershed policy. Dinoflagellates are phytoplanktonic organisms that play an important role in the trophic network. Numerous studies carried out on modern marine sediments showed specific patterns regarding the spatial distribution of fossilizable dinocysts according to sea-surface temperature (SST) and salinity, sea-ice cover duration, inshore–offshore gradient, and/or nutrient concentration (De Vernal et al., 2013; Dodge and Harland, 1991; Marret and Zonneveld, 2003; Morzadec-Kerfourn, 1977, 1979; Mudie et al., 2001; Rochon and De Vernal, 1994; Zonneveld et al., 2013). These palynomorphs have been observed with extremely low concentrations and a poor species richness in modern BB sediments (Lambert et al., 2017) but have proved to be excellent markers for reconstructing past BB sea-surface conditions across the last century (Lambert et al., 2018). Finally, benthic foraminifera are particularly sensitive to various environmental factors (temperature, salinity, oxygenation, pH, hydrodynamism, organic matter export, water depth) and are frequently used to reconstruct bottom water column conditions (Gooday, 2003; Jorissen et al., 2007). In coastal environments, foraminiferal species allow reconstructing natural or human-induced environmental changes (Debenay et al., 2006; Delaine et al., 2015), and among the rare studies conducted in the BB with this proxy, Stéphan (2008) applied it to discuss palaeobathymetric evolution in salt marsh environments related to the Holocene sea-level rise (García-Artola et al., 2018; Stéphan and Goslin, 2014; Stéphan et al., 2015).
In this paper, we thus aim at discussing the combined influences of local (i.e. watersheds) versus regional (i.e. northern European climate) factors driving palaeoenvironmental changes across the first half of the Holocene in western Brittany (from 9200 to 5300 cal. BP), thanks to a cross-correlated pollen–dinocyst–foraminiferal analysis and a pluri-decadal timescale resolution. Furthermore, the studied period covers the Mesolithic to the Neolithic transition, where the environmental changes thanks to the Neolithization must be detectable in palaeoenvironmental sequences.
Environmental contexts
Present and past sedimentological contexts of study sites
The cores selected for this study are located in the westernmost part of Brittany, in marine coastal bays located on both sides of the Crozon peninsula (Figure 1a).
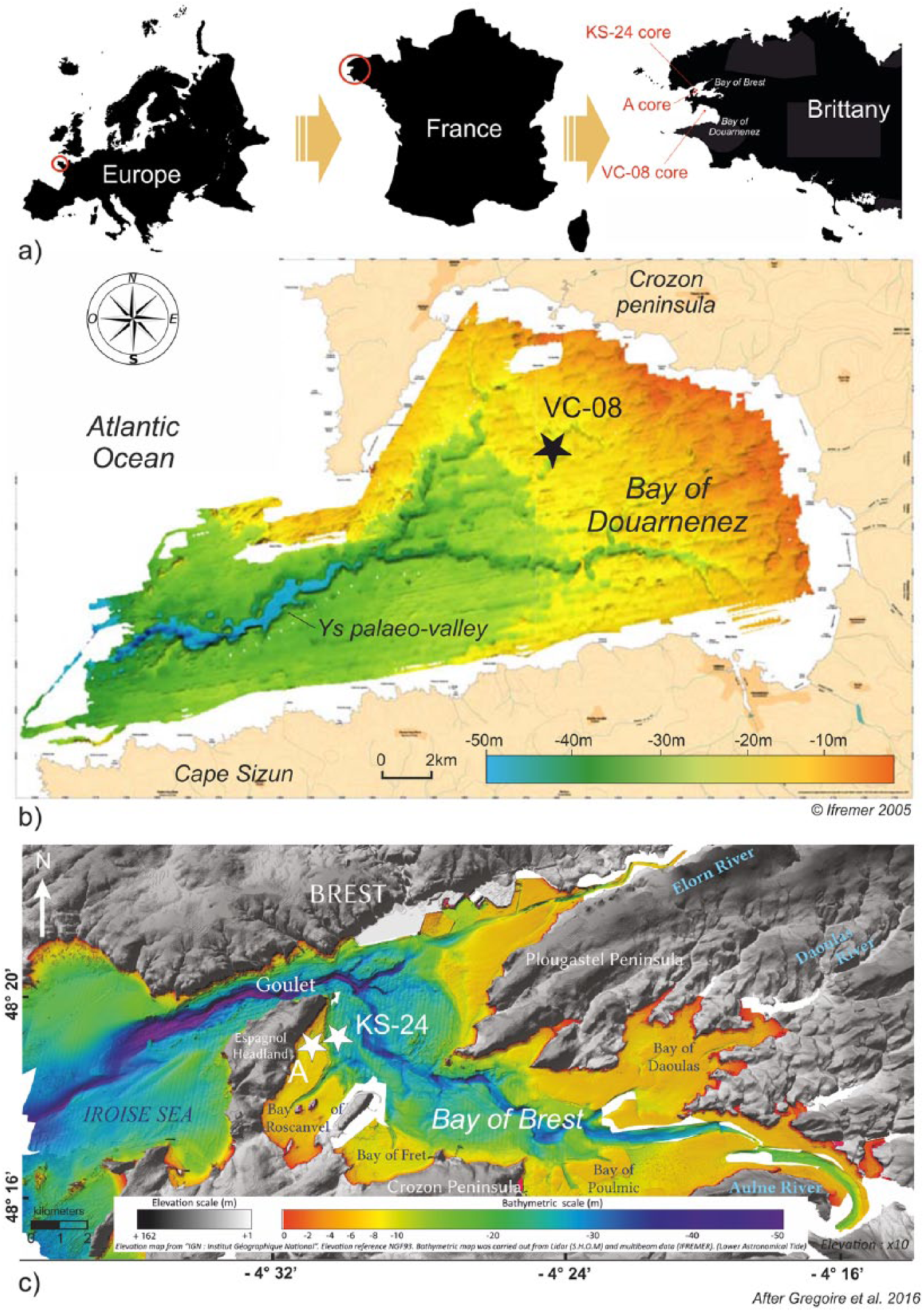
(a) Location of study sites in North Western France; location of sediment cores on the bathymetric maps of the (b) Bay of Douarnenez (black star) and the (c) Bay of Brest (white stars).
The BD
The BD is a large circular bay (350 km²) limited by the Crozon peninsula to the North and the Cap Sizun to the South, largely connected to the Iroise Sea by a passage of about 9 km wide (Figure 1b). Its bathymetry reveals a concave morphology characterized by a water depth of about 20 m, reaching 30 m in the center of the bay (Augris et al., 2005; Figure 1b). The geological basement of the BD and its surroundings corresponds to Brioverian/Palaeozoic sandstones and schist in the North and metamorphic/plutonic formations in the South (Augris et al., 2005; Mélou and Plusquellec, 1974). A major NW–SE trending fault network (Kerforne fault system) affects the BD, inherited from the Hercynian orogeny and reactivated during the North Atlantic opening (Ballèvre et al., 2013; Lefort, 1973). Coasts are mainly shaped by cliffs up to 100 m high in the Crozon peninsula and 85 m toward the Cap Sizun. Some valleys and small estuaries drain very restricted watersheds (40 km² for the larger), sometimes open between cliffs, protecting wetlands and marshes behind the shoreline (Augris et al., 2005). The BD does not receive a large amount of freshwater, implying a strong marine influence with a maximum tidal amplitude of 8 m and a relatively stable salinity (around 35‰; Augris et al., 2005).
The BD sedimentary cover mainly consists of gravels and coarse sands in the central part of the bay and fine sands in sheltered areas (Augris et al., 1988; Hinschberger and Pauvret, 1968), reaching a thickness of 18 m in palaeo-valley axes, but remaining relatively thin over the whole BD (2–4 m; Augris et al., 2005). The substratum study reveals a dendritic palaeo-channel network incising Brioverian basement and flowing into a main valley in the center of the bay named the Ys palaeo-valley (Figure 1b). These channels are connected to the main continental rivers surrounding the BD (Guilcher, 1948; Hallegouët, 1989; Jouet et al., 2003; Musset, 1934). The sedimentary infilling history of the BD is based on the analysis of seismic units (U1–U6; Figure 2) combined with analysis of numerous sediment cores (Jouet et al., 2003; Le Roy and Jouet, 2005). While the BD was totally emerged during the Last Glacial Maximum (around 20,000 yr BP), its flooding, initially confined to palaeo-valleys (10,000–8000 yr BP, meander bar units U3 and U4; Figure 2), then spread out to the rest of the BD (8000–5000 yr BP, fine estuarine sedimentation U5; Figure 2). The sea level was stabilized at about 6000–5000 yr BP (Goslin et al., 2015), and since then, reworked marine sands form the upper unit of the BD sediment cover (U6; Figure 2).

Schematic scenario of the Bay of Douarnenez sediment infilling (after Le Roy and Jouet, 2005). Four sketches related to the sedimentary units defined from seismic and sedimentological analyses (cf. details in Le Roy and Jouet, 2005).
The BB
The BB is located in north-western Brittany (NW France; Figure 1c) and consists of a shallow semi-enclosed basin of 180 km² surrounded by a 230-km-long coastline. Its basement corresponds to Proterozoic igneous rocks in the north and Brioverian/Palaeozoic sediments in the south and east. Present-day low reliefs (few hills reach 330 m high) are inherited from the peneplanation of the Hercynian chain (Ballèvre et al., 2009; Chauris and Plusquellec, 1980). The study area has been subsiding since the Eocene and still today (0.02–0.04 mm/yr; (Bonnet et al., 2000; Goslin, 2014; Ziegler, 1992), but it can be considered as negligible at the Holocene timescale (40 mm/10,000 years; Goslin et al., 2015). The river system is established since the Tertiary (Hallegouët et al., 1994). Today, the BB is characterized by a macrotidal influence with a maximum tidal amplitude of 8 m (Troadec and Le Goff, 1997). Grain size analyses of modern sediments reflect specific hydrodynamic conditions mainly related to tidal currents (Gregoire et al., 2016). In its westernmost part, the BB is connected to the Atlantic Ocean through the ‘Goulet’ (Figure 1c), a strait of about 1.8 km wide and 50 m deep. In its easternmost part, the BB receives main freshwater supplies from the Aulne and Elorn Rivers (both contributing up to 85% of the total river discharges; Delmas and Treguer, 1983), as well as from the smaller Daoulas River (Figure 1c). BB watersheds are characterized by 2000 km of waterways, and most of their runoff flows into the BB through the Aulne River (114 km long; 1224 km² of watershed and 20.40 m3/s of annual debit; Troadec and Le Goff, 1997).
The bathymetric map of the BB highlights submarine channels that attest to palaeo-fluvial systems (Figure 1c; Gregoire et al., 2016). Palaeo-channels of the two main current rivers, Aulne and Elorn, are about 30 and 15 m deep, respectively, and converge in the west at a trough of about 50 m deep (Troadec and Le Goff, 1997). Nevertheless, at present, the depth of the BB does not exceed 10 m deep on approximately 60% of its surface (Monbet and Bassoulet, 1989). The last transgressive episode corresponds to the complex and fragmented sedimentary infilling history of the BB (Gregoire et al., 2017) because of (1) the non-morphological uniformity of the substratum and (2) the non-linear post-glacial sea-level rise, combined with (3) strong and complex hydrodynamical features (Gregoire et al., 2017). Palaeo-valleys became flooded according to different steps of large fluvial terraces (Gregoire et al., 2017), considerably decreasing surfaces occupied by intertidal salt marshes and mudflats (i.e. typical environments submitted to intertidal dynamics) – the shallowest parts of the BB being flooded between 9000 and 7000 yr BP. Today, the center of the BB is predominantly under marine influence, main river mouths and intertidal areas having migrated further east.
Climatic context
Brittany is subjected to a temperate oceanic climate regime characterized by the influence of the westerlies and by low seasonal thermal amplitudes with mean annual temperatures of about 10–11°C (Belleguic et al., 2012). Annual prevailing winds (with speeds that can exceed 100 km/h during 5–15 days per year) have mainly a south-west origin (Troadec and Le Goff, 1997). Since Brittany is submitted to regular oceanic rainfalls, annual cumulative precipitation data range from 600 mm/yr to more than 1600 mm/yr in the inner continental part (Belleguic et al., 2012; Troadec and Le Goff, 1997). The climate of Brittany is due to the combined influences of atmospheric (North Atlantic Oscillation, NAO) and oceanic (Atlantic Multidecadal Oscillation, AMO) circulations detailed thereafter (Lambert et al., 2018; Ruprich-Robert and Cassou, 2015; Tréguer et al., 2014). On decadal to multi-decadal timescales, variations in North Atlantic SST control a large part of the climate variability reconstructed on the continent (Deser et al., 2010; Knight et al., 2006; McCarthy et al., 2015; Ruprich-Robert and Cassou, 2015), already discussed in the BB watersheds (Lambert et al., 2018).
Cultural evolution and territorial occupation dynamics
The Mesolithic is poorly documented in western Brittany due to the scarcity of archeological data. The French Mesolithic period is divided into two subperiods, the first Mesolithic (10th, 9th, and 8th millennia BC) and the second Mesolithic (7th, 6th, and early 5th millennia BC, depending on the studied region) according to cultural criteria based on lithic industries (Marchand, 2014). Across the Mesolithic, traces of shellfish consumption as well as of microlithic industries on the coastline suggest the presence of small communities irregularly spread over western Brittany (perhaps 25,000–50,000 people; Giot et al., 1998). Also, lithic material resulting from debitage has been found in Finistère (western Brittany), evidencing human settlements 20 km from the current coast. Human groups may have moved seasonally between coasts, riversides, and the inner land (Gouletquer et al., 1994, 1996), while recent studies rather suggest perennial settlements of restricted communities in some coastal areas (Marchand, 2005; Schulting and Richards, 2001). Previous pollen studies have not shown significant environmental impacts in Brittany due to human hunter-gatherer populations (Morzadec-Kerfourn, 1974).
The Neolithic ‘revolution’ then reached western Europe around 5500 BC (7450 BP) and the Armorican Massif around 5000 BC (6950 BP; Blanchet et al., 2010) via Danubian agricultural populations, evidenced by the western extension of the Linear Pottery Culture (i.e. Blicquy-Villeneuve-Saint-Germain, in Marcigny et al., 2010), and particularly in Southern Finistère (Marchand et al., 2006; Marcigny et al., 2010; Tinevez et al., 2015) through a diffusion model estimated at 1 km/yr (Giot et al., 1998). The contact between Mesolithic and Neolithic societies remains poorly documented, and predation practices gradually shifted to a production economy thanks to the domestication of animals and the emergence of agriculture. For many years, these cultural changes have been debated as the result of (1) the acculturation of indigenous hunter-gatherer populations or (2) a population replacement by Neolithic societies coming from the East. Recent genetic studies conducted on European populations suggest a Mesolithic population replacement, nevertheless highlighting a mosaic of scenarii according to regions, and a non-uniform demographic transition across Europe with few evidences of cultural adoption (Bramanti et al., 2009; Haak et al., 2010; Sampietro et al., 2007; Skoglund et al., 2012; Von Cramon-Taubadel and Pinhasi, 2011). Western Europe population increased sharply as a result of increasing food production. Perennial settlements of Neolithic villages are observed in western France from the first centuries of the 5th millennium BC (around 7000–6700 BP) (Marchand, 2014).
Considering the emergence of agriculture, palynological and palaeobotanical data are very rare in Finistère. Some studies carried out over Brittany are mainly based on lithic industries and funeral practices (Marchand, 2005), and the beginning of the neolithization thus cannot be precisely dated, such as in other parts of Europe (Kirleis et al., 2012; McClatchie et al., 2014). In the coastal Morbihan (Locmariaquer, SE Brittany), Visset et al. (1996) suggested ancient cereal farming occurred around 7243–5800 BC (9193–7750 BP) because of cereal pollen grain occurrences in Mesolithic levels, observed concomitantly with increases in ruderal plant and Corylus percentages, as well as a marked decrease in other arboreal pollen taxa. Other palynological studies also discussed the presence of cultivated taxa since the Mesolithic in SE Brittany and around the Loire valley (Barbier and Visset, 1997; Joly and Visset, 2005; Ouguerram and Visset, 2001; Visset et al., 2002). However, the absence of agricultural tools in archaeological sites in the direct vicinity of the above-mentioned sites did not allow confirming these assumptions based on palynological observations (Marchand, 2005). Since the early Neolithic, anthropogenic environmental disturbances and evidences of cereal cropping became more and more prevalent (Marguerie, 1991; Visset, 1979). In Finistère, the cereal cultivation practice is attested from 3500 BC (5450 BP) by the presence of cereal grains in the archeological site of Molène island (Dréano et al., 2007). On the other hand, human impact on the environment becomes noticeable from the first half of the 5th millennium BC with the first appearance of grindstones and more particularly with traces detected on lithic furniture typical from the cutting of plant fibers (Giot et al., 1998; Guéret et al., 2014).
Material and methods
Study sediment cores
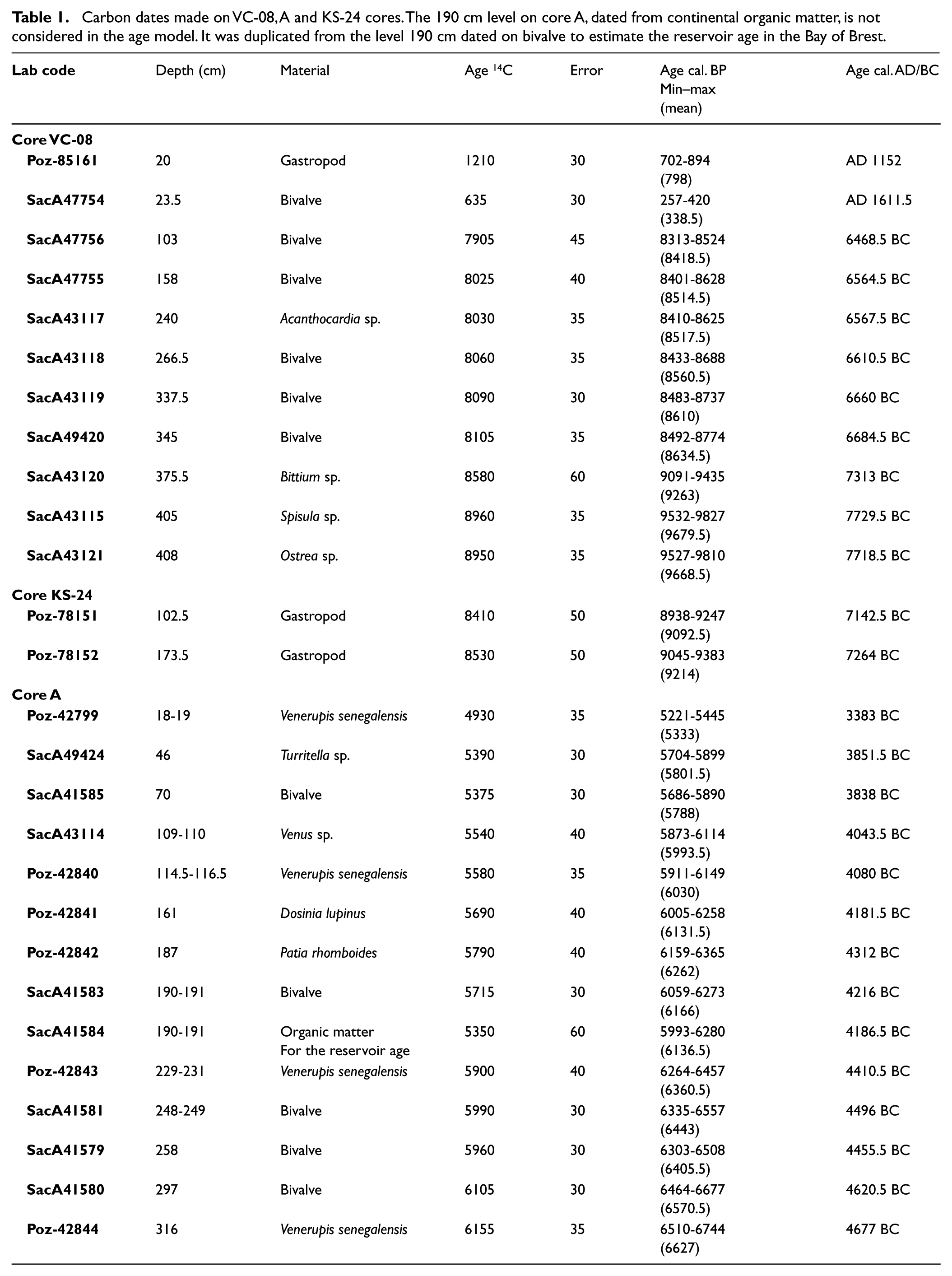
Pictures and RX radiography were carried out at the Géosciences Marines laboratory (IFREMER, Plouzané, France) for the three study cores (Figure 1), and description, granulometric analyses (laser granulometer ‘MASTERSIZER 2000’), and sampling for AMS 14C dates and palynological analyses were carried out at the Géosciences Océan laboratory (LGO-IUEM, Plouzané, France). All dates (Table 1) have been calibrated with the CALIB 7.1 program using the Marine13 calibration curve (Reimer et al., 2013; Stuiver and Reimer, 1993) and a ΔR of −40 ± 23 years (Mangerud et al., 2006). In the manuscript, when referred to a precise age, the mention to ‘Cal’. will not systematically be written, and ages in ‘yr BP’ or ‘yr BC’ will often be both specified so as to facilitate the lecture of the results for palaeoenvironmental or archeological communities, respectively.
Carbon dates made on VC-08, A and KS-24 cores. The 190 cm level on core A, dated from continental organic matter, is not considered in the age model. It was duplicated from the level 190 cm dated on bivalve to estimate the reservoir age in the Bay of Brest.
Within the BD, core ‘VC2012-08-PQP’ (48°10.2′N, 04°26.4′W; 28 m depth; 486 cm length; Figures 1b, 3 and 4) was recovered with a vibrocorer by the R/V Pourquoi Pas? during the Proteus-Dunes cruise in 2012 (Shom). Within the BB, core ‘A’ (48°19.2′N, 4°31.8′W; 8.2 m depth; 418 cm length in total but 318 cm available for this study: the top 100 cm was entirely used in 2003–2004 by biologists and bio-geochemists) was retrieved in the Bay of Roscanvel (BR; i.e. small bay in the western part of the BB; Figures 1c and 5) also with a vibrocorer by the R/V Côtes de la Manche during the ‘Défis Golfe de Gascogne’ program in 2003 (IFREMER, LEMAR-IUEM). Finally, core ‘KS-24’ (48°19.3′N, 4°31.4′W; 26 m depth; 181.5 cm length) was retrieved in the BR (Figures 1c and 5) with a gravity corer by the R/V Thalia during the ‘SERABEQ 3’ cruise in 2015 (IFREMER).

(a) Pictures and (b) XR radiography of the VC-08 core. The red strips represent the sampled core portion to perform palynological analyses.
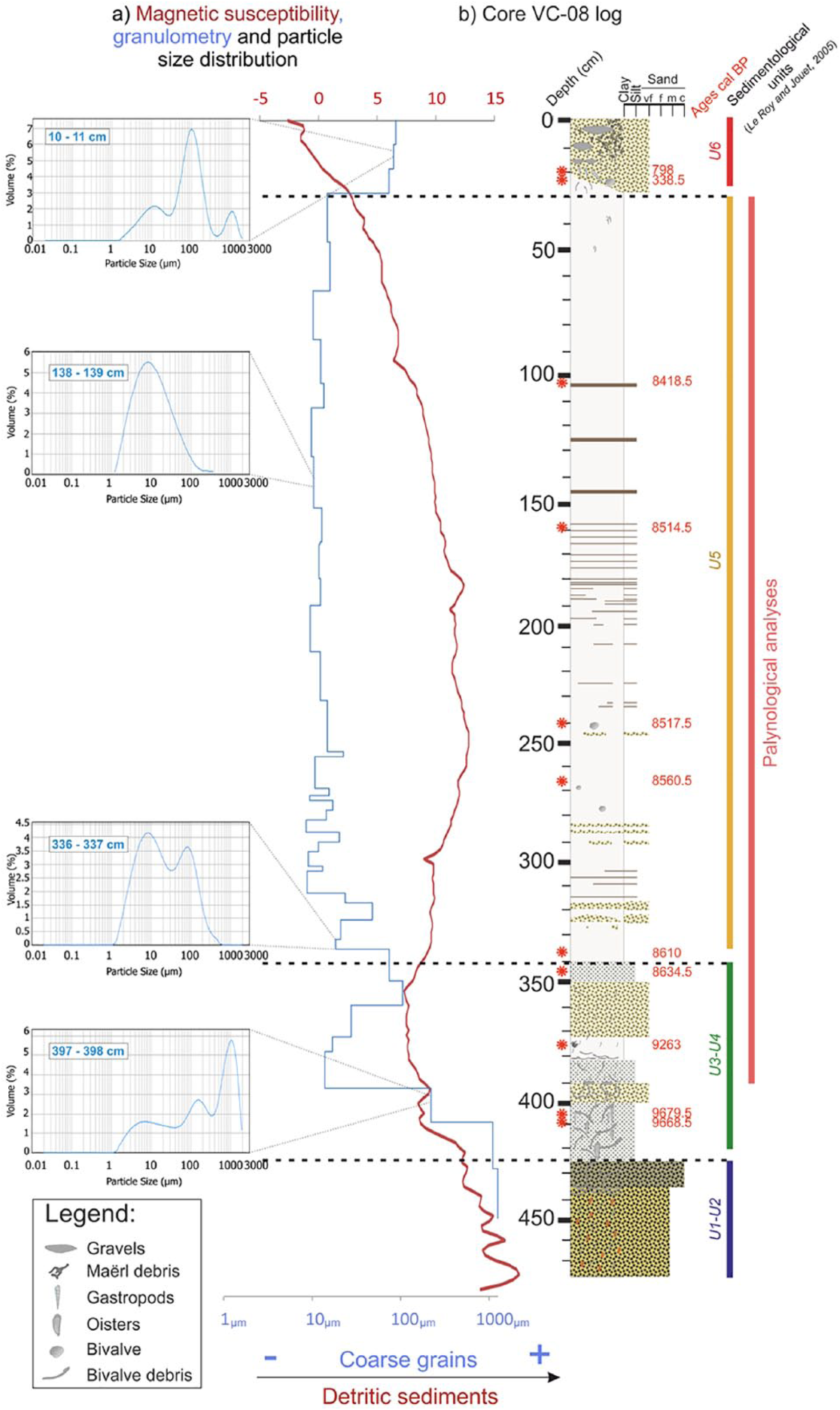
(a) Grain size evolution and magnetic susceptibility along the core and (b) sedimentological log of the VC-08 core. For the different sedimentary units (identified by color strips), a graph representing the grain percentages distribution by size was added. 14C dates are represented by red stars on the log.

(c) Cores of the Bay of Brest, ‘A’ and KS-24, and their location on an MNT of the Bay of Roscanvel with their bathymetric position. Sedimentological logs, photographs, and x-ray radiography for the (a) ‘A’ and (b) KS-24 cores. The 14C dates are represented on the logs by red stars. The red strips represent the sampled core portion to perform palynological analyses.
Palynological analyses
Palynological preparations were carried out at the EPOC laboratory (University of Bordeaux, Talence), following the procedure described by De Vernal et al. (1999) and using chemical (cold HCl and cold HF) and physical (sieving through a 10-µm nylon mesh screen) treatments in order to remove the mineral fraction and to concentrate palynomorphs (cf. http://www.epoc.u-bordeaux.fr/index.php?lang=en&page=eq_paleo_protocoles). The final residue was mounted between slide and coverslip with glycerin. Pollen and dinocysts were determined using an optical microscope Leica DMC 2900 at ×630 magnification.
In this study, 89 samples (46 for core ‘A’, 4 for core KS-24, and 39 for core VC-08) were analyzed, allowing a study resolution of about 20–30 years. Palynomorph identification followed Beug (1961), Faegri and Iversen (1989), Moore et al. (1991), and Reille (1995) for pollen and Rochon et al. (1999) for dinocysts. For each analyzed sample, a minimum of 300 pollen grains and 150 dinocysts have been counted in order to provide robust assemblages from a statistical point of view (Fatela and Taborda, 2002). Percentages were calculated on a sum of total pollen grains, or a sum of total dinocysts, without any exclusion. Concentrations (number of specimens/cm3) were obtained thanks to the Lycopodium spore method (Mertens et al., 2009). Through a camera connected to the optical microscope, measurements were also performed on the size of the annulus and of the diameter of Poaceae grains along the VC-08 core so as to differentiate Cerealia from wild grasses (Joly et al., 2007; Leroyer et al., 2004). Finally, other palynomorphs were counted, including microalgae, foraminiferal linings, spores, and copepod eggs.
Foraminiferal analyses
Prior to the palynological treatments, bulk sediments were sieved at 150 µm, and the fraction >150 µm was used for foraminiferal analyses. After dividing the largest samples with a micro-splitter, benthic foraminiferal assemblages were analyzed with a LEICA M60 binocular stereo zoom microscope at ×60 magnification. Identification followed WoRMS Editorial Board (www.marinespecies.org). Data were expressed in percentages of total benthic foraminiferal counts (at least 100 specimens counted per level). Sixteen samples (12 for core ‘A’ and 4 for core KS-24) were analyzed, allowing a resolution of about 150 years.
Monospecific stable oxygen and carbon isotopes were also measured for core ‘A’ on the Elphidium aculeatum benthic species, in the same levels than benthic foraminiferal assemblages. About five specimens were hand-picked in the 150–250 μm sediment fraction, cleaned in a methanol ultrasonic bath for a few seconds, and then roasted under vacuum at 380°C for 45 min to remove organic matter, prior to isotopic analyses (Duplessy, 1978). δ18O and δ13C (expressed in ‰ VPDB) were measured at the PSO (IUEM, BREST) using the IRMS platform: a Delta V mass spectrometer coupled with a GasBench II preparation line for benthic species. The external reproducibility (1σ) of an internal standard calibrated with NBS19 is ±0.03‰ and 0.06‰ for δ13C and δ18O, respectively.
Sedimentary context and palynological results
BD (core VC-08)
Sedimentary facies and age model
The sedimentology, granulometry, and magnetic susceptibility (MS) analyses (Figures 3 and 4) of core VC-08, taken from a BD palaeo-valley (Figure 1b), highlight different sedimentary deposits that can be related to the sediment units defined in the framework of the study of the sedimentary infilling history of the BD (Le Roy and Jouet, 2005; Figure 2).
The base of core VC-08 (from 474 to 425 cm) is made of coarse sands (mean granulometry of about 1500 µm), with high MS values illustrating a strong detrital terrigenous component (Figure 4a) that can be related to units 1 and 2 (U1 and U2 of Le Roy and Jouet, 2005; Figures 2 and 4b) and interpreted as fluvial deposits during the last glacial period. Between 425 and 340 cm, granulometry oscillates from coarse silts to very fine sands marked by a large grain size variability (from 5 to around 1500 µm; Figure 4a and b), and a fairly high concentration of broken bivalve shells, therefore probably reflecting high energetic conditions. The drop of MS values is related to major environmental change from continental to marine influences, with the sea starting to reach the westernmost part of the channels around 10,000 yr BP. This interval can be associated with units 3 and 4 (U3 and U4 of Le Roy and Jouet, 2005; Figures 2 and 4b). Between 340 and 30 cm, clayey sediments (mean granulometry of 10 µm; Figure 4a and b) and tidal laminae dominate sedimentary facies. After a small increase, MS values gradually decrease from 250 cm to the top (Figure 4a). This very thick deposit constitutes most of the sedimentary infilling of BD channels and corresponds to unit 5 (U5 of Le Roy and Jouet, 2005; Figures 2 and 4b). The last sedimentological deposit (30 cm upward), delimited at its base by an erosive surface, is characterized by marine sands (mean granulometry of 100 µm, three main modes between 10 and 1000 µm; Figure 4a and b) and reworked and remobilized under the action of waves. Extremely low MS values suggest the scarcity of fluvial detrital inputs within the bay. This deposit is related to the last unit, unit 6 (U6 of Le Roy and Jouet, 2005; Figures 2 and 4b).
Radiocarbon dates have been obtained on bivalve and gastropod shells (Table 1). The age model (Figure 6a) was then established from two linear regressions and nine stratigraphic pointers considering two parts obviously distinct considering the description of the core (Figures 4b and 6a). A significant change in sedimentation rates is therefore observed around 8600 yr BP (i.e. 340 cm), related to the Holocene sedimentary infilling history, with 0.06 cm/yr calculated before 8600 yr BP and 1.2 cm/yr calculated after (Figures 4 and 6a). Two additional dates obtained in the uppermost part of the core (i.e. 20 and 23.5 cm, 798 and 338.5 BP, respectively; Figure 4b) confirm the recent setup of the last unit made of reworked marine sands (U6 in Le Roy and Jouet, 2005). These two dates were therefore not used to build the age model (Figure 6a). The palynological study is conducted on the fine intertidal sedimentation section of the core, corresponding to units U4 and U5 described by Le Roy and Jouet (2005) (Figures 3 and 4b) and ranging from 390 to 30 cm (i.e. 9400–8400 yr BP or 7450–6450 yr BC), thus providing information on the final Mesolithic.
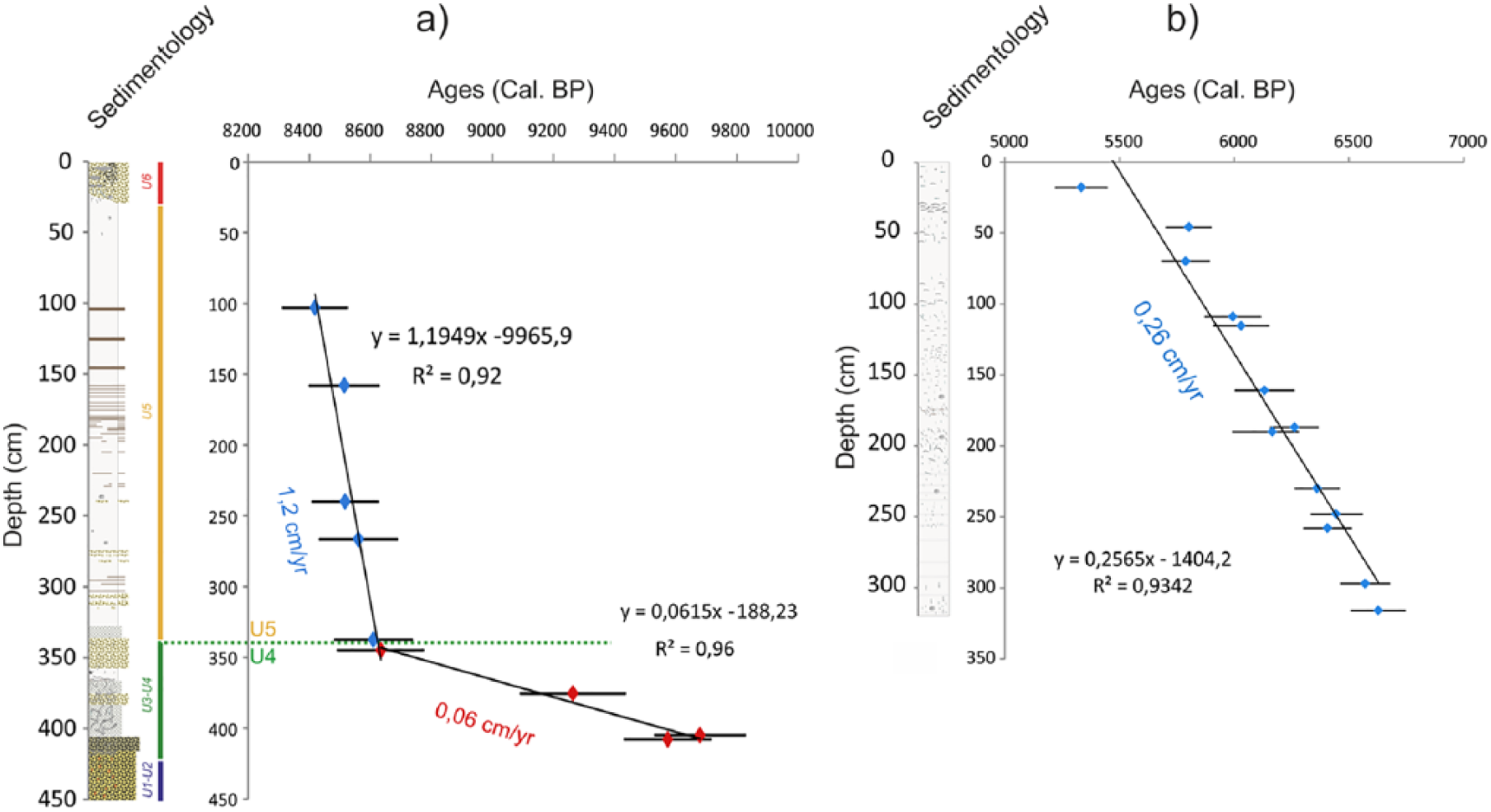
(a) VC-08 (Bay of Douarnenez) and (b) ‘A’ core (Bay of Brest) age models with their sedimentological logs, linear regression lines (in black), and sedimentation rates (cm/yr).
Dinocysts
A total of 18 taxa were recognized, with an average of nine different taxa per slide. Only main taxa (i.e. greater than 2% at least once in palynological spectra) were plotted in Figure 7a along with concentrations (cysts/cm3) and specific richness (number of different taxa per slide). Taken as a whole, Spiniferites bentorii (50% in average) and Lingulodinium machaerophorum (11%) are the most abundant taxa as it is usually described on Brittany’s coasts (Lambert et al., 2017; Morzadec-Kerfourn, 1977). The variations of the dinocyst assemblages enable to highlight three main zones, with very low diversity values at the base of the record (VC-A, 390–345 cm), followed by a slow increase of up to 250 cm (VC-B, 345–250 cm) and higher values over the entire second half of the core (VC-C, 250–30 cm). Diversity then shifts from around 5 to 12 different taxa per slide. In addition to the low dinocyst-specific richness, VC-A palynozone is defined by atypical elevated percentages of Achomosphera spp., suddenly replaced by S. bentorii. The VC-A/VC-B boundary is well marked in dinocyst assemblages by the occurrences of Spiniferites mirabilis, Spiniferites ramosus, and Spiniferites delicatus. Moreover, palynozone VC-B corresponds to increasing percentages of L. machaerophorum and a slow decrease of S. bentorii ones. Finally, palynozone VC-C corresponds to a disappearance of S. mirabilis simultaneously observed with decreasing percentages of Selenopemphix quanta and the first occurrence of Spiniferites lazus found in the study core (Figure 7a). Moreover, these biozones are highly related to the variations of dinocyst concentrations.
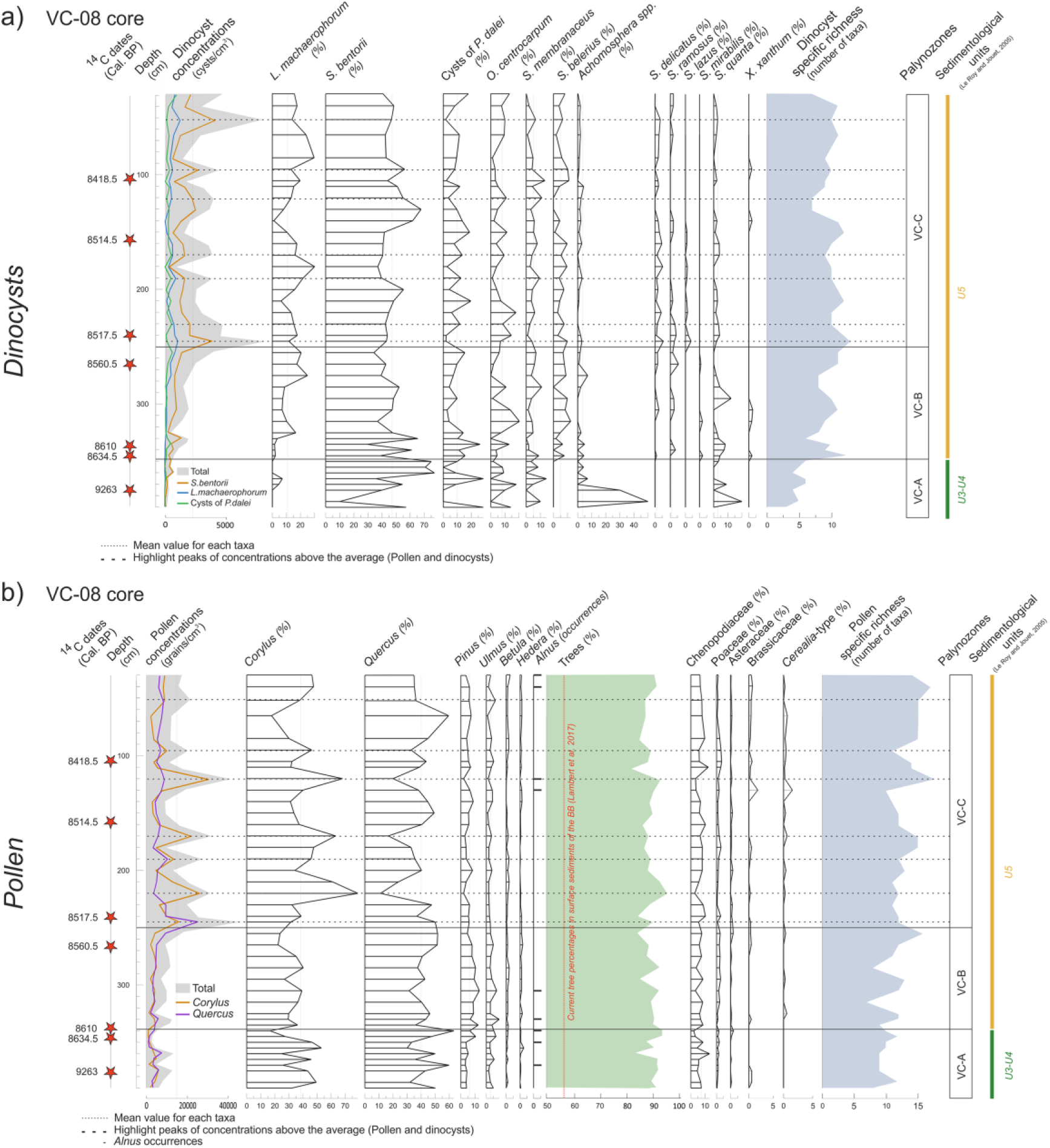
Graphs against depths with major taxa (greater than 2%) of (a) dinocysts and (b) pollen along the VC-08 core, Bay of Douarnenez. The 14C dates are represented by red stars. The dotted lines represent the pollen concentration peaks.
Pollen
A total of 33 taxa were recognized, with an average of 12 different taxa per slide. This rather low diversity due to the forest taxa dominance differs from the greater diversity observed in more recent BB pollinic sequences (e.g. Lambert et al., 2017, 2018) and resulting from the herbaceous diversification accompanying the significant anthropogenic landscape opening. In addition to the taxa characterized by percentages found at least once above 2% in assemblages, some scarce but meaningful elements of the vegetation (i.e. Alnus, Cerealia-type) are plotted in Figure 7b. In general, percentages of tree pollen grains are relatively stable, then, representing the most abundant vegetation group (average of 90%). They are predominantly represented by Quercus (42%) and Corylus (33%), both taxa being then logically anti-correlated all along the core, and accompanied by low percentages of Pinus, Ulmus, Betula, and Alnus. Among herbaceous plants, Chenopodiaceae are clearly dominant (7%) as well as Poaceae (2%).
The overall diagram displays weak variations along the core except for total pollen concentrations and diversity that allow identifying two parts corresponding, first, to both VC-A and VC-B palynozones described from the dinocysts (that we will refer to as palynozone VC-A&B in the following description) and, second, to VC-C palynozone (Figure 7b). The limit between VC-A&B and VC-C is more specifically marked by a sharp increase in pollen concentrations and specific richness. Palynozone VC-A&B also corresponds to a decreasing trend of Corylus, correlated to a Pinus increase, then replaced by Quercus. In parallel, the pollen-specific richness tends to increase. The palynozone VC-C is marked by oscillations of the two major taxa, Corylus and Quercus, up to 30%, contrasting with the lower part of the diagram. Concentration peaks (dotted lines in Figure 7b) correspond to increases in Corylus percentages and related decreases in Quercus ones. In addition, a decrease in Ulmus is noticed throughout the sequence, along with a slight increase in herbaceous plants (e.g. Poaceae, Asteraceae, Brassicaceae). Also, Cerealia-type is more pronounced in palynozone VC-C. We can note again the overlap between biozones and variations in pollen concentration, as for dinocysts.
Toward a Holocene sedimentary infilling history reconstruction of the BD
The sedimentological and palynological analyses conducted on core VC-08 (depicted in depth in Figure 7 and in age in Figure 8) allow clarifying the sedimentary infilling model discussed by Jouet et al. (2003) and Le Roy and Jouet (2005).
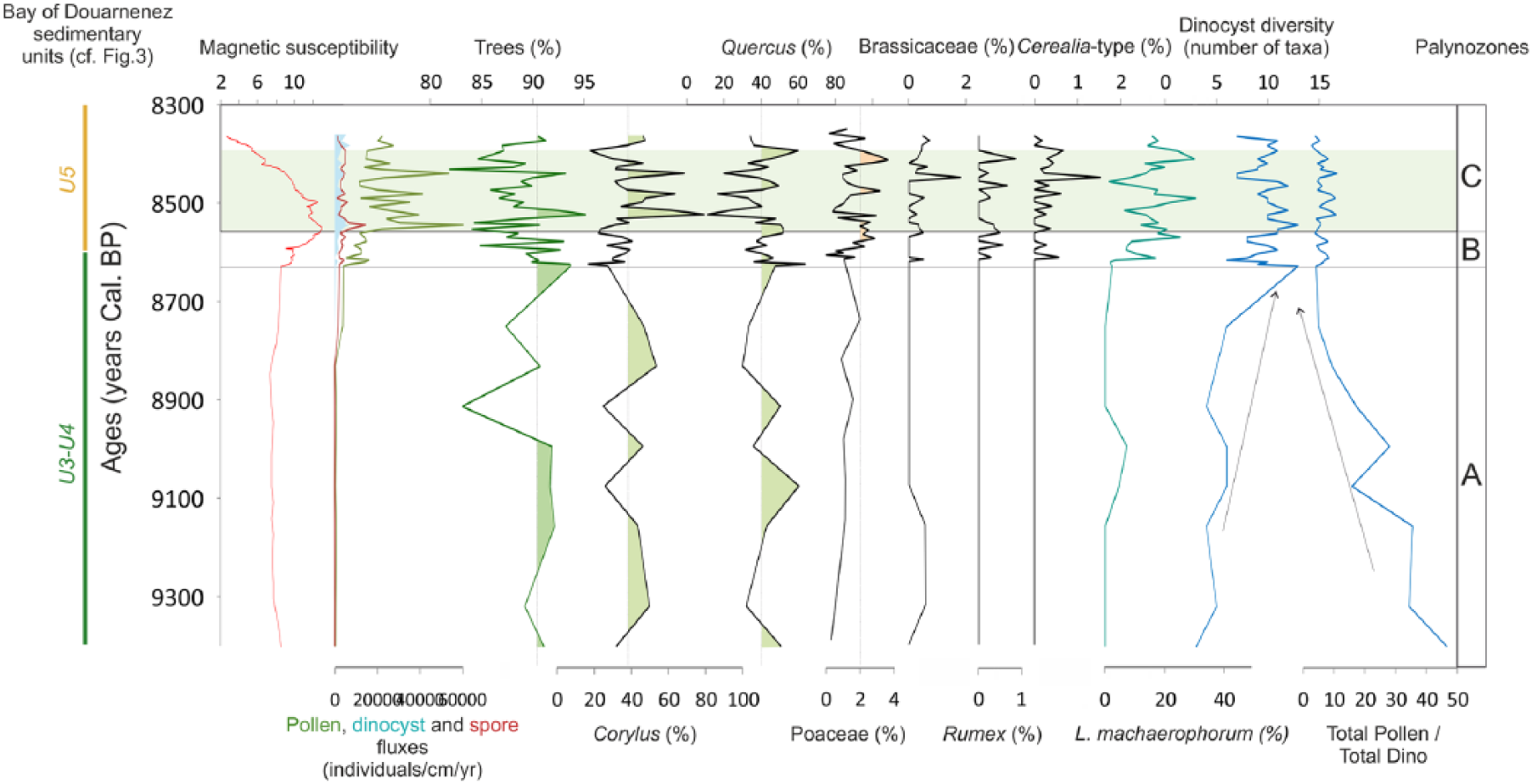
Diagram depicted in ages, from the VC-08 core, grouping together data from palynological counting (pollen, dinocyst and spore concentrations, percentages of trees, Corylus, Quercus, Poaceae, Brassicaceae, Rumex, Cerealia, the dinocyst L. machaerophorum, and the total pollen/dinocyst ratio) and the magnetic susceptibility, with respect to sedimentary units defined in Figure 4.
From 474 to 420 cm (Figure 4b), sediments are too coarse to be sampled for palynological analysis. This part could be related to the bedrock incision by rivers during the end of the last glacial period (U1 and U2; Figure 2), as also evidenced in our study by high MS values (Figure 4a). Palynological results highlight a high pollen/dinocyst ratio from 390 to 340 cm (8630 yr BP) (palynozone VC-A; Figure 8), suggesting a major pollen contribution of surrounding watersheds and therefore predominant fluvial versus marine inputs, corresponding then to the fluvial deposits (fluvial accretion bars of U3-U4 in Le Roy and Jouet, 2005; Figures 2 and 4b). The Chenopodiaceae content (Figure 7b) is likely a clue of the already settled salt marshes. Still discrete marine influences do not allow dinoflagellates to occur in large number during palynozone A, the study core being located upstream in the palaeo-river network incising the substratum of the BD (Figure 1b). Unfortunately, the ecology of Achomosphera spp. is not precisely known and the taxa thus cannot be connected to particular ecological conditions at that time.
From 340 to 250 cm (8630–8550 BP, palynozone VC-B in Figures 7 and 8; lower part of U5 in Figure 4b), the marine influence is first marked by the increased dinocyst diversity, including euryhaline taxa such as L. machaerophorum and S. belerius, together with taxa more characteristic of marine environments such as Operculodinium centrocarpum, S. delicatus, S. mirabilis and S. ramosus (Morzadec-Kerfourn, 1977, 1979). In parallel, decreasing percentages of the typically coastal species S. bentorii are probably more related to increasing percentages of the other taxa. The lower part of U5 corresponds to a thick transitional sequence between continental and marine influences, submitted to the relative sea-level rise as observed through the continuous decrease of the pollen/dinocyst ratio (Figure 8). The already mentioned breakdown in sedimentation rates at 340 cm (limit between U4 and U5; Figures 4 and 6a) corresponds to this major transition toward an estuarine sedimentation. Variations in particle size and in sediment facies are also obvious through the sudden jump in MS values, indicating greater continental intakes from 8600 yr BP (Figures 4a and 10).
From 250 cm (palynozone VC-C in Figures 7 and 8; U5 upper part in Figure 4b), marine influences are well established with a low pollen/dinocyst ratio (Figure 8), a high dinocyst diversity, and the persistence of typically coastal (i.e. S. bentorii) and marine (i.e. cysts of Pentapharsodinium dalei, O. centrocarpum, Scadoxus membranaceus, S. delicatus) dinocyst taxa, already observed in the previous zone (Figure 7a). L. machaerophorum, often associated with fluvial and estuarine brackish environments (Marret and Zonneveld, 2003), displays percentages that exceed the average of its whole dataset (Figure 7a) and are anti-correlated with S. bentorii and/or O. centrocarpum (i.e. more related to the oceanic domain; Morzadec-Kerfourn, 1976, 1979) ones.
Palynological studies conducted on core VC-08 therefore confirm the environment sketches proposed by Le Roy and Jouet (2005) on the basis of sedimentological and seismic data. Furthermore, an obvious major limit can be defined within the tidal facies of U5 at 8550 yr BP, with a lower transitional part under increasing marine influences and an upper part characterized by well-established marine conditions in a perennial flooded bay.
Western BB (A and KS-24 cores of BR)
Sedimentary facies
Core ‘A’ (Figure 5a) is characterized by clayey sediments and numerous shell debris (Figure 5b) as well as bivalve and gastropod shells in life position often sampled for dating. The age model (Figure 6b) was established through a linear regression between 13 AMS 14C dates (Table 1), allowing to consider an interval of about 1400 years (6700–5300 BP or 4750–3350 BC) and mean sedimentation rates of about 0.26 cm/yr (Figure 6b). Palynological analyses have been conducted along the 318 cm of the core. It is worth noting that in one sample, in the 190–191 cm level (Table 1), continental organic matter (vegetal fibers) has been dated in parallel with a bivalve for obtaining an estimation of the age reservoir within the BB. When calibrating the 14C ages with IntCal13 without assuming any age reservoir effect on the marine carbonate material, we obtain a difference of about 357 years between both dates, thus indicating a ΔR of −43 years, very close to the one calculated by Mangerud et al. (2006) of the Sein Island (i.e. ΔR of −40 ± 23 years) (http://calib.org/marine/).
Regarding core KS-24 (Figure 5a), sampled in a deeper but closed site, the basal part is characterized by clayey sediments sampled for palynological analyses, and the upper part consists of coarse sands and shell debris (Figure 5c). These two parts of the core are separated by a well-marked erosive surface at around 100 cm depth. Two AMS 14C dates carried out at 173.5 and 102.5 cm allow dating the clayey section (i.e. interval sampled for palynological analyses; Figure 5c) between 9214 and 9092 yr BP (7264 and 7142 yr BC).
Dinocysts
In both cores, 24 different taxa were identified, with an average of three different taxa per slide for core KS-24 and 12 for core ‘A’. Similar to core VC-08, only main taxa were plotted in Figure 9a along with dinocyst concentrations and the specific richness. These two cores constitute two temporal windows (i.e. 9200–9000 yr BP for core KS-24 and 6700–5300 yr BP for core ‘A’) on the same study site (i.e. BR, westernmost part of the BB; Figure 1c). Both cores are described together, in their stratigraphical order, with core KS-24 representing a palynozone by itself.

Diagram in depth grouping the major taxa (greater than 2%) of (a) dinocysts and (b) pollen for the ‘A’ and KS-24 cores, both taken from the Bay of Roscanvel and plotted one above the other according to the time periods covered by the two sequences. The 14C dates are represented by red stars. For the ‘A’ core, the most significant minor pollen taxa (greater than 1%) were also reported. The dotted lines represent the pollen concentration peaks.
Core KS-24 is characterized by the largely dominant S. bentorii species (69%; Figure 9a), similar to the BD at the same period (Figure 7a). Co-occurring with cysts of P. dalei, O. centrocarpum, S. membranaceus, and S. quanta at the base, S. bentorii becomes monospecific at the top of core KS-24. From the base to the top of core ‘A’, dominant taxa are first characterized by cysts of P. dalei and then replaced by L. machaerophorum that reach about 80%. Decreasing percentages of cysts of P. dalei and S. ramosus are consequently observed at the same time. Variations in percentages of major species and in total concentrations allow us to delimit three palynozones (two of them being subdivided into two sub-palynozones). The first palynozone A-A (from 318 to 250 cm; Figure 9a) displays a significant increasing trend of cysts of P. dalei, a decreasing trend of L. machaerophorum, and high percentages of S. ramosus and S. membranaceus. At the limit between A-A and A-B, total cyst concentrations as well as L. machaerophorum percentages show their lowest values, while cysts of P. dalei reach its highest percentages (i.e. around 50%). The second palynozone A-B (250–100 cm) is well marked by noticeable opposite trends, and especially by increasing percentages of L. machaerophorum and decreasing ones of cysts of P. dalei. Then, S. bentorii displays higher values than in the previous zone, while S. ramosus displays lower percentages. Moreover, palynozone A-B corresponds to the strongest occurrences of S. lazus. This palynozone has been divided into two sub-palynozones. Within zone A-B1, L. machaerophorum percentages are the lowest while those of cysts of P. dalei are the highest of their respective whole datasets. L. machaerophorum occurrences gradually rise in zone A-B2, and S. bentorii percentages stabilize with a plateau of high percentages during this interval. The third palynozone A-C (from 100 cm onward; Figure 9a) corresponds to a huge increase in L. machaerophorum percentages while those of cysts of P. dalei sharply decrease, together with a drop in S. ramosus. An additional subdivision is proposed to highlight the abrupt increase in dinocyst concentrations at 50 cm (limit between A-C1 and AC-2). Furthermore, within zone A-C1, weak but significant occurrences of S. membranaceus and S. mirabilis are observed, while percentages of cysts of P. dalei, S. mirabilis, and also S. ramosus and S. lazus drop to extremely low values within zone A-C2. Conversely, O. centrocarpum and S. belerius slightly increase in the last zone A-C2.
Pollen
In both cores, 34 different taxa were recognized with an average of 11 different taxa per slide for core KS-24 core and 12 for core ‘A’ (Figure 9b). While the two major taxa are the same for both cores (i.e. Corylus and Quercus), pollen concentrations are much higher in core KS-24 (9200–9000 yr BP) than in core ‘A’ (6700–5300 yr BP). Furthermore, trees are much more present in core ‘A’ than in the core ‘KS-24’ (i.e. 93% vs 79%). Among the herbaceous in the core KS-24, Chenopodiaceae are particularly represented with an average of 12%.
In core ‘A’, increasing percentages of Alnus (from 0% to 15%), and decreasing trends of Ulmus and Tilia, allow to delimit the same palynozones than those described for dinocysts (Figure 9a). In general, herbaceous plants are very rare, but a slow diversification is observed toward the top, with the appearance of some minor taxa (e.g. Ranunculaceae, Mercurialis, Plantago spp., Caryophyllaceae). Corylus and Quercus are anti-correlated throughout the core. More specifically, the first palynozone A-A (318 to 250 cm; Figure 9b) corresponds to low percentages of Corylus that gradually increase at the end of the palynozone and, conversely, to high percentages of Quercus that tend to gradually decrease. During this interval, Tilia and Poaceae represent a meaningful part of the pollen content. The second palynozone A-B (250–100 cm) displays the occurrence and rise of Alnus, as well as the noticeable decrease of both Poaceae and Tilia. Furthermore, at the start of palynozone A-B, Ulmus and Pinus percentages exhibit stable values when compared with palynozone A-A, while they suddenly drop around 200 cm, allowing to discriminate sub-palynozones A-B1 (from 250 to 200 cm) and A-B2 (from 200 to 100 cm). Conversely, Alnus percentages strongly increase in sub-palynozone A-B2. Palynozone A-C (from 100 cm onward) highlights a new threshold with a marked decrease in Ulmus together with a strong increase in Alnus percentages, the latter reaching its highest values within zone A-C2 in parallel with relatively continuous high pollen concentrations. Interestingly, most of the main peaks observed with pollen concentrations (dotted lines in Figure 9b) also correspond to increases in Alnus percentages along with high values of Corylus, as already noticed for the VC-08 record.
Micropalaeontological results
Benthic foraminiferal assemblages are presented in depth for cores ‘A’ and KS-24 (Figure 10), in parallel with the isotopic data measured on E. aculeatum. Since Elphidium spp. are the dominant taxa, all other taxa percentages have been calculated on a main foraminiferal sum that excludes Elphidium crispum and E. aculeatum. Similar to palynological data, only species occurring at least once with percentages above 2% have been plotted in Figure 10.

Major benthic foraminiferal taxa data plotted against depths (greater than 2%) in the ‘A’ and KS-24 cores. 14C dates are represented by red stars. Ammonia spp., Cibicides spp., P. mediterranensis, Epiphytic species, Lagena spp., and Miliolidae are represented in percentages according to a main sum excluding the Elphidium species, major taxa that greatly tainted the individual signal of each one. The isotopic data measured on shells of E. aculeatum species are added to the graph.
Along core KS-24, foraminiferal concentrations are low and assemblages are dominated by Haynesina germanica (average of about 50%), with few Elphidium spp. and important variations of Ammonia spp., here strictly assigned to the species Ammonia tepida.
Along core A, an increase in total benthic foraminiferal concentrations is observed throughout the sequence and epifauna are the most represented in benthic foraminiferal assemblages, with both E. aculeatum and E. crispum accounting for 70%. Zones previously described for dinocysts and pollen grains have been reported in the foraminiferal diagram so as to facilitate the cross-correlated approach between all fossil bio-indicators. Zone A-A is mainly characterized by Elphidium spp. that displays an increasing trend, an important content of Miliolidae, and an abrupt decrease in Cibicides spp. In addition, A. tepida and Planorbulina mediterranensis are observed with significant percentages. Palynozone A-B corresponds to still high Elphidium spp. percentages and occurrences of A. becarii. Within this interval, two subzones correspond first (i.e. A-B1, 250–200 cm) to decreasing values of Cibicides and Miliolidae, together with the occurrence of Lagena spp., and then (i.e. A-B2, 200–100 cm) to increasing percentages of Cibicides (reaching 20%) and drastically low Lagena percentages. Within palynozone A-C (from 100 cm onward), A. becarii is replaced by A. tepida, while Cibicides spp., P. mediterranensis, and epiphytes (i.e. with a flat face that allows them to be attached to sediments or plants) show approximately a same increasing profile, opposite to the Elphidium spp. trend.
Regarding isotopic analyses (Figure 10), δ13C and δ18O show opposite trends with a slight decrease toward lighter values for the δ18O signal (amplitude of 1‰ between minimal and maximal values in the dataset, ranging from 1.8‰ to around 0.8‰) and a slight increase toward heavier values for the δ13C signal (amplitude of 2‰ between minimal and maximal values in the dataset, ranging from −1.5‰ to around 0.5‰).
Environmental evolution in the BR under the rising sea-level influence
Environments of the BR were greatly affected by the relative sea-level rise between 9000 and 5000 yr BP. The general palaeoenvironmental evolution of the BR can be reconstructed thanks to two Holocene windows obtained on core ‘A’ (6700–5400 yr BP) and core KS-24 (9200–9090 yr BP) and with averaged palynological (pollen and dinocysts) and foraminiferal results compiled in pie charts (Figure 11).

(a) Mean palynological (pollen and dinocysts) and micropalaeontological (benthic foraminifera) data for the ‘A’ and KS-24 cores represented in pie charts. (b) Diagrams representing two models of the landscape evolution in the Bay of Roscanvel following the sea level rise. On each model, the typical foraminifera encountered in the different environments according to their ecology were added.
Throughout core KS-24, dinocyst assemblages show a low general diversity with major occurrences of S. bentorii (Figures 9a and 11), arguing for a strictly coastal environment. Among pollen assemblages (Figures 9b and 11), core KS-24 displays an important content of Corylus, a species considered as pioneer during the post-glacial temperate forest (mixed oak) reconquest that mainly marked north-western European landscapes around 9000 yr BP (Huntley, 1993; Tinner and Lotter, 2001). Among herbaceous plants, Chenopodiaceae are significantly present, certainly coming from nearby salt marshes. In addition, foraminiferal assemblages are dominated by H. germanica, A. tepida, and Elphidium williamsoni. A. tepida and E. williamsoni are closely related to intertidal sheltered mudflat environments, while H. germanica is related to continental organic matter inputs (Debenay et al., 2006; Rossi et al., 2011). These three species indicate a highly tidal-influenced environment and local small runoffs rich in continental organic matter (Debenay et al., 2006; Perez-Belmonte, 2008; Redois, 1996; Rossi et al., 2011). However, the noticeable absence of L. machaerophorum and Alnus suggests low major freshwater supplies. The surrounding continental context may then correspond to small watersheds, while the Aulne River was still flowing to the north of core KS-24 location taking into account the low sea level position at that time (i.e. –26 m; Gregoire et al., 2017; Figure 11). In this context, the marine influence remains confined to the axis of the main channels of the Aulne and Elorn Rivers (Gregoire et al., 2017). The higher river terraces (i.e. where core ‘A’ was taken; Figures 1c and 5c) are entirely emerged at that time, while the lower site where core KS-24 has been retrieved (Figures 1c and 5c) is characterized by the development of abundant maritime marshes (schorre and slikke) with halophilous plants (i.e. Chenopodiaceae) and a rather weak fluvial dynamic. Considering the current 26 m depth location of core KS-24 and its 1.81 m length, it allows deducing a relative sea level between 27 and 28 m under the current sea level, which is very similar to the 26 m proposed at 9000 yr BP by Gregoire et al. (2017), taking into account the macrotidal context of north-western Brittany.
Throughout core ‘A’, dinocyst assemblages display a general higher diversity than in core KS-24 with 24 taxa, mostly oceanic to coastal, along with occurrences of about 35% of L. machaerophorum (estuarine taxon). Predominant marine influences, favorable for diversified dinoflagellate blooms, are associated with the BR marine flooding in a context of superimposed fluvial influences, as evidenced by L. machaerophorum, mainly associated with Aulne River inputs, here perceptible compared with the previously marsh environment described with core KS-24. Concerning pollen assemblages, the mixed temperate forest is settled (90% of tree pollen grains; Figures 9b and 11) with a higher representation of Quercus when compared with core KS-24. This indicates a connection of the BR to the mainstream of the Aulne River due to the flooding of the BB, thus highlighting a more regional signature (i.e. BB watersheds) in core ‘A’, also corresponding to the colonization of temperate tree taxa in western Europe during the middle Holocene (Brewer et al., 2002; Ruddiman and McIntyre, 1981). The Chenopodiaceae signal is less pronounced in core ‘A’ due to the disconnection of the study site from marsh environment, as also confirmed by the drastic reduction in marsh-related foraminiferal species (i.e. H. germanica, A. tepida, and Elphidium williamsoni). Also, benthic foraminiferal assemblages display a stronger diversity in core ‘A’, suggesting a greater marine influence in agreement with dinocyst results. Moreover, the presence of some epiphytic and fixed species (i.e. Cibicides spp. and Planorbulina mediterranensis) suggests a higher hydrodynamism. In addition, the absence of benthic infauna could indicate a lack of very important organic matter inputs. The environment would therefore be characterized by a sufficient organic matter supply to allow the development of algal coverings (allowing the presence of epiphytes), and hydrodynamism would disperse this organic material, thus oxygenating the bottom. In summary, around 6000 yr BP, the relative sea level was relatively close to the present-day one (Goslin et al., 2015) and the BB as a whole (and thus the BR) is flooded (Figure 11). The environment is then marked by a clearer marine influence and the remoteness of salt marsh areas. Tidal currents erode sediments on terrace slopes from the main channels, leading to erosive surfaces separating muddy and sandy facies (i.e. upper part of KS-24 core from 100 cm onward not sampled for palynological analysis; Figure 5c), as also identified by Gregoire et al. (2017).
Climate impacts and land-use changes
Local imprints of the 9000–8000 and 6000–5000 yr BP RCC events
Superimposed on the palaeoenvironmental reconstitutions discussed in the context of the rising sea-level context, climate trends are well expressed in our palynological records due to our high-resolution study carried out on cores characterized by high sedimentation rates.
In the BD, from 8550 to 8400 yr BP (zone VC-C; Figures 7b and 12), Corylus becomes clearly dominant. Moreover, this interval displays a slight decrease in Ulmus, a disappearance of the thermophilous taxon S. mirabilis, and significant occurrences of S. lazus, a dinocyst associated with cold SST (Zonneveld et al., 2013). These palynological evidences may support a cooling event. Indeed, this cooling associated with the appearance of a strong seasonal contrast between 8855 and 8000 yr BP observed by Naughton et al. (2007) favors the development of Corylus woodlands and contributes to the decrease in deciduous Quercus forest expanse. At the same time, the obvious increase in pollen fluxes, coeval with a marked increase in L. machaerophorum percentages, and Alnus ones to a lesser extent, may argue for increasing fluvial discharges (as previously discussed in Lambert et al., 2017) and thus of humidity. It is worth noting that low percentages of Alnus may here be ascribed to its later settlement in the vegetal reconquest succession, as observed in the Armorican Massif by David (2014). Each L. machaerophorum increase (Figures 7a and 12), also concomitant to the withdrawal of coastal (S. bentorii) and marine (O. centrocarpum) species, coincides with increasing percentages and concentrations of Corylus (Figures 7b and 12).
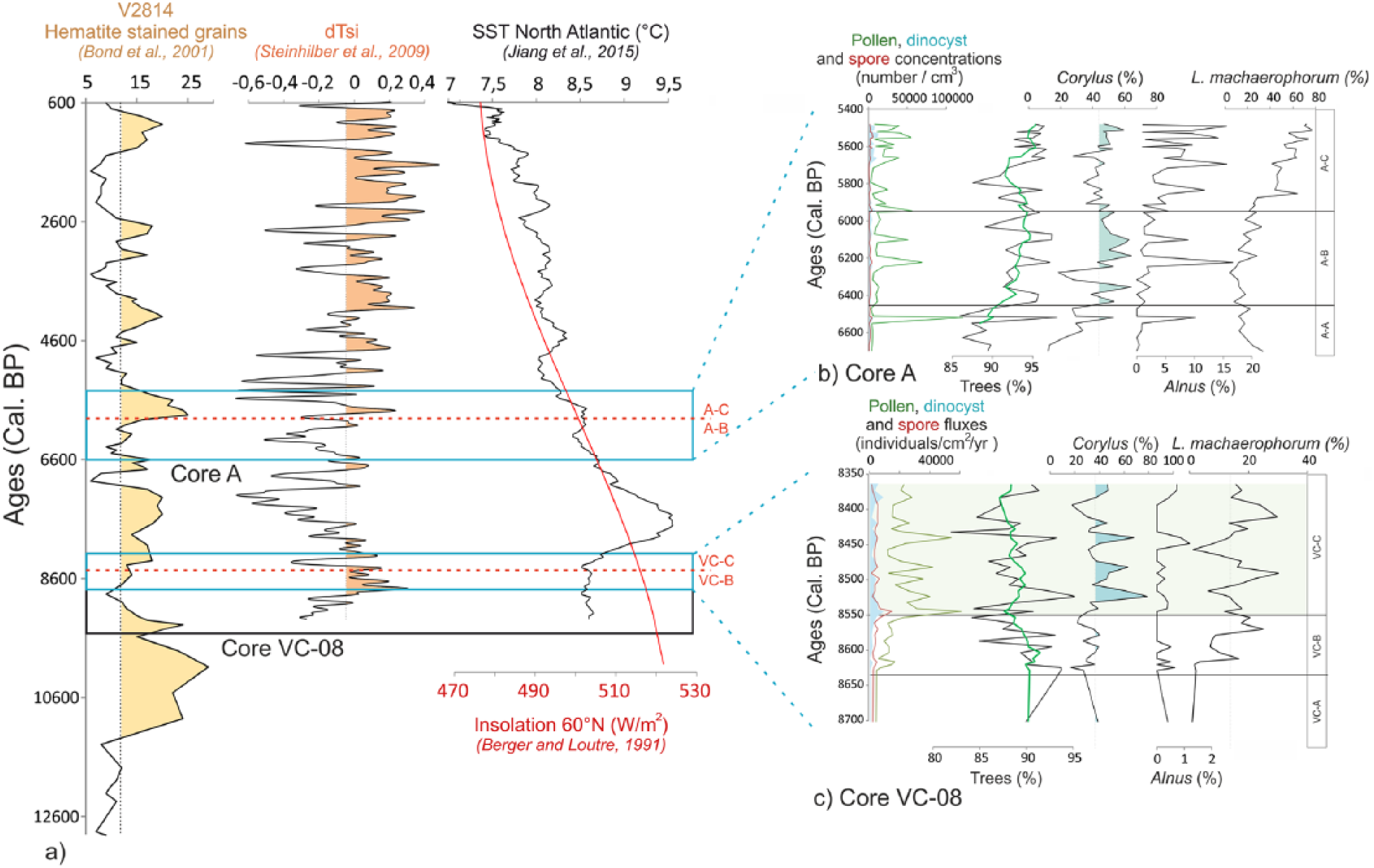
(a) Published reconstituted palaeoclimatic data across the Holocene in the North Atlantic region (solar irradiance (dTSi) by Steinhilber et al., 2009, sea-surface temperature (SST) by Jiang et al., 2015, and Northern Hemisphere summer insolation by Berger and Loutre, 1991) and the detrital hematite-stained grains by Bond et al., 2001. (b) Diagram showing the palynological data of the ‘A’ core between 6600 and 5400 cal. yr BP (from left to right: pollen and dinocyst concentrations, tree percentages, Corylus and Alnus percentages, and the L. machaerophorum/S. bentorii ratio). (c) Diagram showing the palynological data of the VC-08 core between 8700 and 8400 cal. yr BP with the same succession of palynological proxies. The temporal extent of each core is represented by a black rectangle in (a) and the temporal extent of the graphs (b) and (c) is represented by a blue rectangle. The limits of palynozones A-B/A-C and VC-B/VC-C have also been reported in (a).
In the BR, dinocyst assemblages were dominated by S. ramosus (oceanic taxa; Morzadec-Kerfourn, 1977) and cysts of P. dalei before 6000 yr BP and are characterized after this limit (between zones A-B and A-C) by an obvious and sharp increase in L. machaerophorum percentages from 30% to 70% (Figures 9a and 12), a species usually encountered in estuarine and fluvial environments of Brittany’s coasts (Morzadec-Kerfourn, 1977). This could emphasize the establishment of stratified waters related to increasing fluvial inputs. Furthermore, L. machaerophorum is accompanied by increasing percentages of the heterotrophic taxa S. quanta (Figure 9a) that could support increasing fluvial nutrient inputs to the BB. Moreover, since Alnus began to appear on the territory, we now observe an increase of this tree in parallel with L. machaerophorum. As previously observed for core VC-08, Corylus increases are concomitant with peaks of pollen concentrations and of L. machaerophorum percentages. Finally, higher percentages of H. germanica and A. tepida (Figure 10), benthic foraminiferal species subordinate to environments under fluvial influences (Debenay et al., 2006; Estournès et al., 2012; Perez-Belmonte, 2008), confirm previous observations of increasing fluvial discharges probably related to increasing moisture.
For both cores VC-08 and ‘A’, increasing fluvial inputs would correspond to a local detection of the large-scale Holocene millennial-scale climatic variability (i.e. ‘Bond events’, Bond et al., 2001; or RCC, Mayewski et al., 2004). Most of these cold Holocene events, initially identified in North Atlantic sedimentary cores by detrital grains drained by glaciers (Figure 12a), correspond to Scandinavian glacier advances (Nesje et al., 2001), colder northern hemisphere temperatures, and drier conditions in the tropics (Mayewski et al., 1997, 2004; Meeker and Mayewski, 2002). Also, the strengthening of the westerlies (Bradbury et al., 1993; Mayewski et al., 2004) may have resulted in recurrent positive modes of the NAO, leading to increased precipitations in north-western Europe. Interestingly, Mojtahid et al. (2013) associated the 6000–5000 interval with persistent positive NAO conditions in the Bay of Biscay. A solar irradiance decline may be responsible for this 6000–5000 yr BP event (Steinhilber et al., 2009; Figure 12a), also leading to a North Atlantic SST decrease (Jiang et al., 2015; Figure 12a). Thus, core VC-08 around 8550 yr BP, and core ‘A’ around 6000 yr BP (Figure 12c), may then evidence for the first time the regional imprint in western Brittany’s coasts of two RCC events through significant increases in moisture and fluvial discharges.
Local signal of Cerealia-type pollen grains during the Mesolithic: coastal Poaceae or early traces of a proto-agriculture?
Between 8600 and 8300 yr BP, VC-08 core shows a very homogeneous laminated facies, which attests to a stable environment aggradation related to a deltaic alluvial plain characterized by intertidal regimes (Le Roy and Jouet, 2005; Figure 4). Constant percentages of Chenopodiaceae all along the record, associated with few and constant percentages of Poaceae (Figure 7b), suggest a coastal marsh (slikke and schorre-type) environment, confirming the aggradation of intertidal deposits. In this sedimentary context, the palynological analysis interestingly records iterative occurrences of large pollen grains of Poaceae (Figure 13).
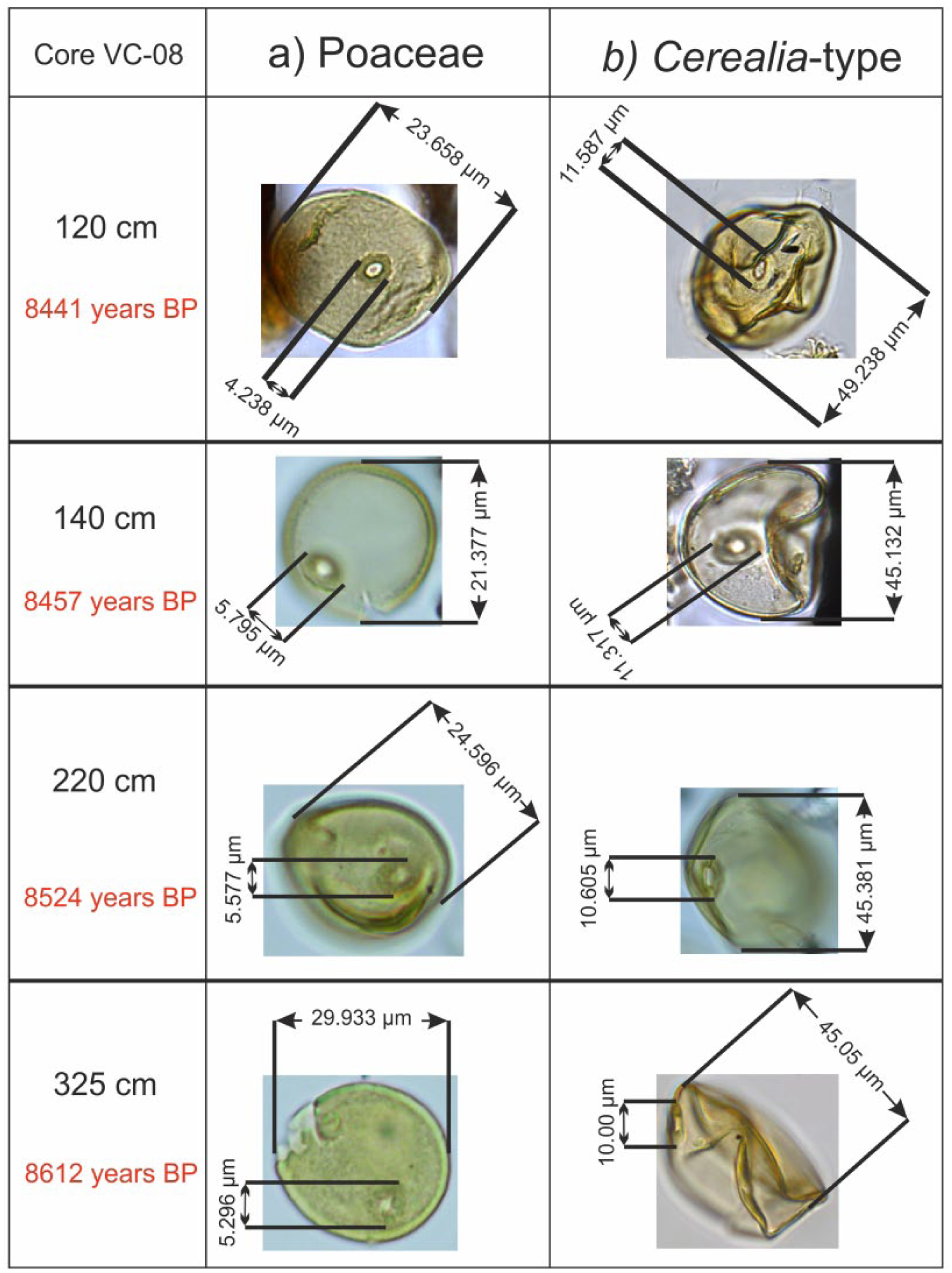
Pollen grain photographs of wild (a) Poaceae and (b) cereals, with grain and annulus diameter measurements for each one, on four levels of the VC-08 core.
Cereal pollen identifications are usually based on the diameter criteria (grain and annulus) following Beug (1965), Leroyer et al. (2004), and Joly et al. (2007), because of the impossibility to separate wild grasses and Cerealia-type pollen grains using exine sculpture types (Beug, 2004). In western Europe, the commonly accepted thresholds of 45 µm for the grain diameter and 8 µm for its annulus (that we will refer as the 45–8 µm criteria) theoretically enable the distinction between cereal pollen grains and those of indigenous wild grasses from the French western Atlantic coast (Leroyer et al., 2004). Considering these criteria along the whole sequence, seven levels delivered cereal-type grains with one to three occurrences in palynological slides. These 12 large pollen grains recognized in total, and distributed all along the core (325, 295, 220, 170, 140, 120, and 65 cm; Figure 13), have been carefully measured, providing average values of 46 µm for the grain diameter and 9.9 µm for the annulus. Nevertheless, the ‘45–8 µm’ criteria have been criticized (Joly et al., 2007). The statistical study conducted by Joly et al. (2007) on modern plants from the Atlantic coast reveals that the ‘45–10 µm’ criteria make it possible to identify 91% of cereals (but 9% of pollen grains are from wild grasses). In their study, Joly et al. (2007) then proposed to increase the thresholds to the most discriminant level of ‘47–11 µm’, thus avoiding to consider, as much as possible, larger pollen grains of coastal grasses from the Atlantic coast. The ‘47–11 µm’ criteria then make it possible to discriminate 96% of cereal pollen grains (i.e. 4% of wild poaceae fall within this criterion) (Joly et al., 2007). However, with these new criteria, the sensibility to detect Cerealia pollen grains significantly declines. More precisely, among 100 modern Cerealia pollen grains, only 59 would be detected as such with the ‘47–11 µm’ criteria, the other grains being then supposed as wild Poaceae. Despite this low sensibility and the fact that some cereals are probably not taken into account with these thresholds, the applied ‘45–11 µm’ criteria to our data confirm discreet peaks of Cerealia-type pollen grains, from around 8600 to 8300 yr BP (at 65, 120, and 170 cm; Figure 13).
Considering that indigenous coastal grasses may have larger pollen grains than continental species, it is also worth noting that these weak occurrences observed in the BD record do not co-occur in parallel with variations of other Poaceae or Chenopodiaceae, suggesting no significant changes in landscape and/or pollen transport at that time. Moreover, to be more confident about these atypical and early observations, many other clues must be considered. Some plants often associated with crops (e.g. adventitious taxa) are necessary to discuss anthropogenic signatures in the vegetation cover (Behre, 1981; Willcox, 2005). It is precisely interesting to note that slight increases in Plantago lanceolata, Rumex sp., and Brassicaceae percentages are observed in VC-08 core during main occurrences of Cerealia-type (Figures 8 and 14). A slight long-term decrease can also be highlighted in tree percentages between 8600 and 8300 yr BP, perhaps arguing for an early start of a small human-made landscape opening (Figures 8 and 14).
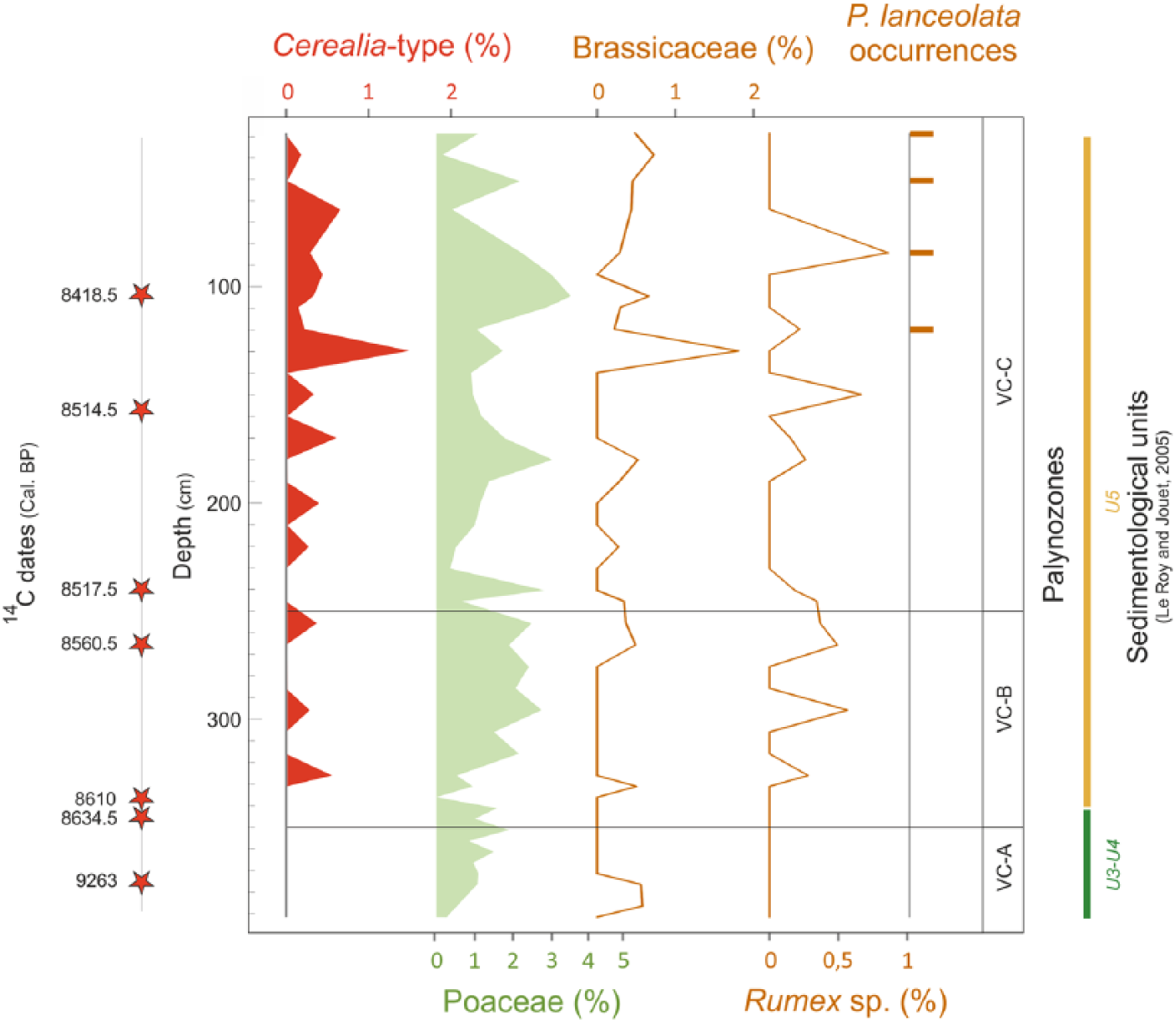
Diagram in depth from the VC-08 core, representing variations of some herbaceous taxa percentages (i.e. Cerealia-type, other Poaceae, Brassicaceae, Rumex sp.) and the occurrences of Plantago lanceolata.
Our findings are, however, in marked opposition with the regional neolithization model admitted by the archaeological community and based on clues related to changes in cultural practices identified by lithic (and ceramic) industries (Blanchet et al., 2006; Cassen et al., 1998; Hamon, 2008; Marchand, 2005, 2007; Marchand et al., 2006; Pailler et al., 2007, 2014). In addition, studies on Mesolithic archeological sites in Brittany have highlighted traces of plant cuttings, but tools did not show the classical polishes associated with cereal cuttings (Guéret, 2013; Guéret et al., 2014). Even if our results raise questions and encourage caution, they are consistent with other palynological studies carried out in western France that also evidenced early discreet appearances of Cerealia-type pollen grains (Figure 15). As early as 1996, analyses conducted by Visset et al. opened the debate about the existence of a very ancient cereal farms as highlighted by low occurrences of Cerealia-type pollen between 7200 and 5800 yr BC (9200–7800 yr BP), concomitantly with increases in Corylus and ruderal plant percentages and decreasing Quercus percentages in the coastal Morbihan region (SE Brittany; Figure 15). Other studies along the Loire River revealed the presence of cereal pollen taxa as well as Juglans between 6400 and 5900 yr BC (8400–7900 yr BP; Ouguerram and Visset, 2001) and between 6600 and 5800 yr BC (8500–7800 yr BP; Carcaud et al., 2000). Moreover, Joly and Visset (2005, 2009) reported cereal and ruderal pollen grains on the southern Atlantic coast (Vendée) between 7500 and 6300 yr BC (9500–8200 yr BP) and between 7500 and 6200 yr BC (9400–8100 yr BP). Beyond Brittany, some early Cerealia-type occurrences have already been observed as co-occurring with ruderal plants between 6400 and 5800 yr BC (8400–7800 yr BP) in the northern Pyrenean region by Galop and Vaquer (2004) or between 6600 and 5800 yr BC (8600–7800 yr BP) in the Parisian Basin (Leroyer and Allenet, 2006). In addition, occurrences of Cerealia-type pollen grains have been reported from many Swiss sites between 7750 and 5800 cal. BC (Erny-Rodmann et al., 1997; Lotter, 1999; Tinner et al., 1999, 2007); Beckmann, 2004; Figure 15). Taking into account our data and the numerous references cited above, we underline the great timing coherence of large Poaceae pollen grain appearances, with sizes usually interpreted as resulting from a cultivated origin. Among the reservations that are often opposed to these detections (e.g. reliability of dating, long-distance transport or laboratory contaminations; Behre, 2007), we can consider minimal errors in carbon datings (see the core stratigraphy) or in laboratory treatments that are identical for all cores (i.e. Roscanvel ‘A’ core, without any large Poaceae pollen grain). Regarding the long-distance transport of pollen grains from the eastern areas where cereals were already cultivated, as already mentioned by Tinner et al. (2007), it is surprising that we do not detect pollen grains from other exotic species with a better pollen dispersion than cereals (e.g. Quercus ilex, Pistacia). Moreover, the BB and BD watersheds are too small to receive long-transported pollen grains from eastern or southern sites. We can thus suggest two hypotheses:
These atypical large pollen grains are not inherited from cultures and represent a signature of littoral grasses. Indeed, some coastal Poaceae on the Atlantic coast may have pollen grains that can reach large sizes (Joly et al., 2007). The size threshold which is considered by palynologists as a robust identification criteria for the determination of cereal pollen grains may be revised again (cf. Joly et al., 2007) in light of the particular pollen sizes of some regional littoral grasses. Regarding this first hypothesis, it still appears curious not to detect any significant increase in total Poaceae or Chenopodiaceae when the most relevant peaks of Cerealia-type pollen grains are recorded (Figure 7b). Furthermore, in the BR records (A and KS-24 cores; Figure 9b), studied from 9200 to 5300 yr BP in same environmental sedimentary context, the total absence of these large pollen grains of Poaceae before 8600 yr BP implies no peculiarity of size among pollen grains from littoral grasses.
A discreet proto-agriculture signal would be recorded for the first time during the Mesolithic in western Brittany. Discussed by the Mesolithic archaeological community, this type of ‘agriculture’ would be characterized by its discretion in the landscape and the absence of specific hard rock–made agricultural tools (Marchand, 2005). The work of plants seems indeed to be grown toward the end of the Mesolithic, but regional use-wear analysis on archeological material suggested that wear traces result from scrapping of wood and soft plants, with, however, no traces of cereal plant work (e.g. Gassin et al., 2013; Beg an Dorchen, SW Brittany). In the previous palynological studies mentioned above (Figure 15), Cerealia-type pollen grain occurrences are always scarce and isolated in space and time (Visset et al., 2002). Our study, conducted on a core taken in the marine coastal domain, enables to record a temporal BD watershed signal with iterative occurrences of these large pollen grains (8482; 8440; 8395 yr BP or 6532; 6490; 6445 yr BC) associated with some other cultural indicators as Plantago lanceolata and Rumex spp. (Figure 14). The scarcity of Mesolithic data on the territory, and the difficulties to identify proto-agriculture evidence, both with pollen or lithic data, does not allow to discuss the local introduction of this proto-agriculture imprint as a result from a meridional migration movement (Retzian culture via the Loire estuary; Visset et al., 2002) or from the main Neolithic cultural in Brittany from a Danubian origin (Dubouloz, 2003; Gomart et al., 2015; Figure 15).
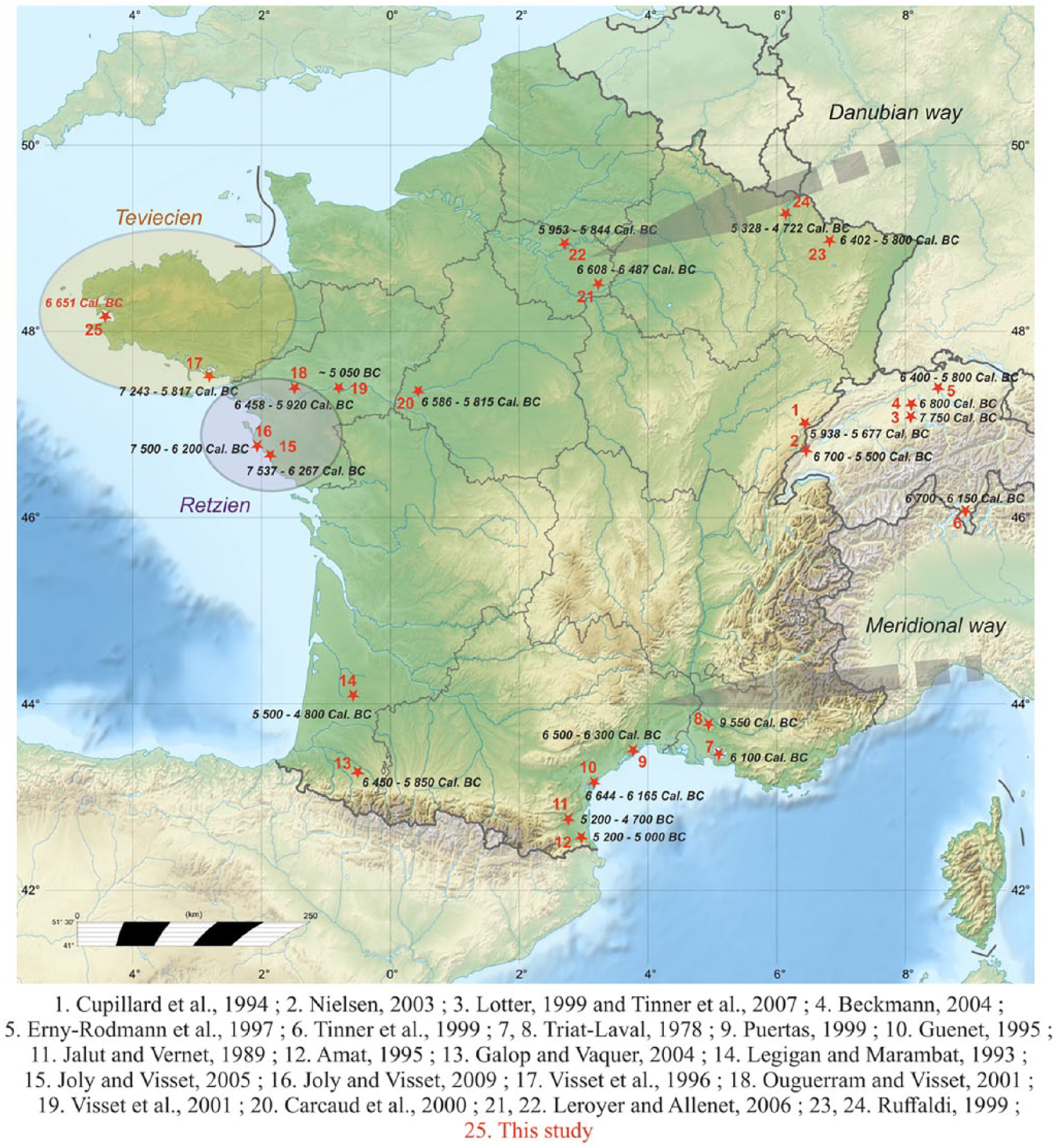
Map showing the first occurrences of Cerealia in published pollen records, with related dates. The two circled areas (Teviecien and Retzien) delimit the cultural specificities of the end of the Mesolithic after Marchand (2005). The gray arrows correspond to the different neolithization paths reaching Western Europe.
In order to progress, future studies must be conducted (a) to newly establish the size variation of the current littoral wild grass pollen grains and (b) to confirm the timing of the first large pollen grain detection at a broader regional scale.
Conclusion
Our results allowed characterizing Holocene coastal palaeoenvironments of NW France (BB and BD, western Brittany) over two given periods (9200–8400 and 6600–5300 yr BP). Various factors forced coastal environmental changes during the early to middle Holocene, especially including the relative sea-level rise that modified sedimentation processes and the post-glacial recolonization of temperate trees. Sedimentological and palynological analyses carried out in this study allowed us to characterize and specify the environmental variations that affected coastal environments of western Brittany. The relative sea-level rise influenced the sedimentary infilling history of shallow marine environments that transited from river valleys to oceanic bays. In addition, high-resolution studies enabled us to detect the regional response to more global events arising from the millennial-scale climate variability in the North Atlantic. Thus, around 8600 and 6000 yr BP, onsets of two major abrupt climate events are locally detected by a moisture increase marked by strong occurrences of Corylus, Alnus, and L. machaerophorum dinocyst taxon. In addition, the gradual decrease in continental summer temperatures is marked by the withdrawal of thermophilous species Ulmus and Tilia. Moreover, during this time period also marked by the appearance of the neolithization, Cerealia-type pollen grains have been detected in the BD, about 1500 years before the advent of agriculture commonly accepted. These local results, while being consistent with other palynological studies conducted on the French Atlantic coast, must be taken with caution and should open the way to further studies so as to confirm these ‘pre-domestic’ agriculture indices.
Footnotes
Acknowledgements
We thank the UMR CNRS 5805 EPOC (Talence) for palynological laboratory procedures (Muriel Georget), the UMS 2572 LMC14 (Saclay) for carbon dating via Artemis project fundings, the sedimentology laboratory of the Shom (‘Service Hydrographique et Océanographique de la Marine’) for the VC-08 core taken from the Bay of Douarnenez (PROTEUS-DUNES cruises; 2012), the laboratory IFREMER-Marine Geosciences (Plouzané) for the KS-24 core collected in the Bay of Brest (SERABEQ cruises; G. Gregoire PhD thesis, 2016), and the LEMAR laboratory (‘Laboratoire des Sciences de l’Environnement Marin’; IUEM, Plouzané) for the A core taken from the Bay of Brest (Défis Golfe de Gascogne cruise; 2003). Main issues of this project are integrated within the theme ‘Dynamics of Human Settlement and Paleoenvironments’ of the Zone Atelier Brest Iroise (ZABrI, INEE-CNRS).
Funding
This study was supported by the French CNRS and is a contribution to the 2015-2016 INSU project EC2CO-LEFE: ‘CAMOMI: Convergences/Approches croisées des signaux MOléculaires et MIcropaléontologiques pour décrypter les forçages anthropiques et climatiques en milieu côtier (Rade de Brest)’ and the UBO-BQR project: ‘PARADE: Signature PAléoenvironnementale des séquences holocènes en RADE de Brest’. This work was supported by the ‘Laboratoire d’Excellence’ LabexMER (ANR-10-LABX-19) and co-funded by a grant from the French government under the program ‘Investissements d’Avenir’.