
Abstract
Here we present an almost complete and integrated Holocene record of marine and terrestrial palaeoenvironmental change from the NE shelf of the Black Sea. A dinoflagellate cyst record used to reconstruct Holocene sea-surface conditions highlights that the NE shelf was a brackish water environment, with a minimum salinity of 7 psu in the early-Holocene before changing at a gradual rate to a more saline environment with maximum salinities of ~18 psu being reached around 3 cal. ka. A warming phase was detected from 6 cal. ka BP, with warmest conditions between 3 and 2.5 cal. ka BP. A pollen record is used to examine the major climate and land-use changes in the eastern Black Sea region. Biome reconstructions show that the temperate deciduous forest dominates throughout the record, although with an overall decline. From early-Holocene to the first hiatus around ~9 cal. ka BP, Pinus pollen dominates, while taxa representing a mixed oak-hornbeam-beech forest are less abundant, indicating relatively cool and dry conditions. Between ~7.9 and ~6.1 cal. ka BP, a thermophilous deciduous forest established, suggesting an overall warming trend and humid conditions. From 4 cal. ka BP, Pinus dominates the pollen record, accompanied by an increase of herbs, implying an opening of the landscape, which would coincide with the beginning of the Meghalayan Age. The integrated record of the marine and terrestrial climate indicators supports the notion that this change in landscape may have been triggered by a combination of warmer and drier conditions and human activities in this region.
Introduction
Environmental change in the Black Sea during the Holocene has been increasingly investigated using various proxies over the past two decades. Numerous published studies agree that the reconnection of the Black Sea to the global ocean, via the Bosphorus Strait, initiated between ~9.6 and 9 cal. ka BP (e.g. Aksu et al., 2016; Hiscott et al., 2002, 2017; Ivanova et al., 2015; Lister et al., 2015; Major et al., 2006; Yanchilina et al., 2017). The initiation and development of the reconnection and the establishment of a two-way exchange of water between the Black Sea and Marmara Sea was the primary control on sea-level rise in the early- and mid-Holocene (Balabanov, 2007; Brückner et al., 2010; Hiscott et al., 2017). However, the baseline conditions prior to the establishment of the two-way flow and the rate of change after ~9.6–9.0 cal. ka BP are still contested. Two recently published studies highlight this disagreement. According to Yanchilina et al. (2017), the Black Sea palaeoshore was around 120 mbsl around ~9.3 cal. ka BP, with a Black Sea disconnected to the Marmara Sea. It was estimated that the filling of the Black Sea to reach the level of the Bosphorus Strait took less than 40 years. In contrast, the recent study of Aksu et al. (2016) demonstrated that there was a strong Black Sea outflow in the Marmara Sea in the early-Holocene, suggesting that the sea level was either at the Bosphorus sill level or slightly higher. Most recent studies agree that modern sea levels, albeit minor fluctuations, were established ~4 cal. ka BP (e.g. Balabanov, 2007; Brückner et al., 2010).
Secondary controls on the conditions of the Black Sea during the Holocene include regional climate change and human activity. In comparison with studies examining the reconnection to the global ocean, these secondary influences are less well investigated. However, a few records show some interesting pattern of climate change during the Holocene, sometimes conflicting. The Sofular Cave record (Göktürk et al., 2011) highlighted enhanced and more intense precipitations in the southern Black Sea region between 9.6 and 5.4 cal. ka BP, in agreement with regional climatic trends reconstructed from pollen records from the east Mediterranean (e.g. Kotthoff et al., 2008). However, further west, in the Marmara Sea, in the Gulf of Gemlik, sedimentary records suggest dry conditions between 8.2 and 5.4 cal. ka BP followed by alternation of wet and dry climate conditions (Filikci et al., 2017). Pollen record from Varna Lake suggests relatively humid conditions early-Holocene until 5.6 cal. ka BP with continental conditions replacing oceanic ones (Filipova-Marinova et al., 2016). Few climate reconstructions are available for the NE region of the Black Sea. A recent study based on pollen records located in the Taman Peninsula from Bolikhovskaya et al. (2018) shows alternations of warm/arid phases with humid intervals for the last 7400 years. Finally, the pollen-based climatic reconstructions for Europe by Mauri et al. (2015) highlight the contrast between the northern and southern regions of the Black Sea. Holocene human occupation around the Black Sea has been relatively well documented by archaeologists (e.g. Anthony, 2007), notably in the west and north. A pollen record from the coastal NW Bulgaria region of the Black Sea evidences human impact on the vegetation from around 7250 cal. year BP (Tonkov et al., 2014). Other record of significant land use (deforestation) was presented by Giosan et al. (2012) who showed the effects of catchment processes in the late-Holocene in the area surrounding the Danube Delta, including on dinoflagellates.
Palynological records yield fundamental information about past vegetation, biogeography and palaeoclimate reconstructions (e.g. Mauri et al., 2015). The Black Sea region has very few Holocene terrestrial pollen records, despite being of high interest as a region of vegetation refugia during the LGM (e.g. Leroy and Arpe, 2007). However, marine cores have provided suitable palynological records, in particular in the NW and SW parts of the sea (Cordova et al., 2009; Marinova et al., 2018). In the SE, Black Sea basin (core 22-GC3/8) has provided insights on the late Quaternary evolution of the vegetation, with evidence of warmer than present-day conditions during the last interglacial period, ca. 125 ka, as well as an increase of precipitation by 100–200 mm/year (Shumilovskikh et al., 2013). A detailed pollen record from this core covering the entire Holocene interval (Shumilovskikh et al., 2012) has been analysed using a quantitative method of vegetation reconstruction, also known as ‘biomization’ (Prentice et al., 1996). This globally tested method (Prentice et al., 2000; Wanner et al., 2008) uses plant functional types (PFTs), which are broad classes of plants defined by growth and leaf form, phenology and climatic adaptations. The PFT concept allows objective assignment of palaeobotanical data (i.e. pollen and plant macrofossils) to major regional vegetation types (i.e. biomes) and selection of a dominant biome by calculating/comparing the numerical biome scores (Prentice et al., 1996). The same PFT concept is also used in all climate-driven vegetation models, which can simulate global and regional vegetation distributions (Prentice et al., 1992). The use of a common set of PFTs in the biome reconstruction and simulation allows more robust climatic interpretations of the analysed palaeodata and facilitates data-model comparisons (Prentice et al., 1996).
In contrast to the SW and NW, the NE region of the Black Sea has a paucity of Holocene pollen records, either on the land or in basin (Bolikhovskaya et al., 2018; Mauri et al., 2015). However, ostracod- and mollusc-based studies of benthic changes in this area show three periods of differing environments (Ivanova et al., 2007, 2012, 2015; Zenina et al., 2017). These studies have documented the dominance of Caspian-type assemblages from early-Holocene to 7.4 cal. ka BP followed by a relatively short interval of cohabitation of Caspian and Mediterranean types (between 7.4 and 6.8 cal. ka BP). From 6.8 cal. ka BP, ostracod and mollusc fauna are progressively dominated by Mediterranean species. Very similar assemblages were found in the SW shelf (Williams et al., 2018), except that the disappearance of Caspian type occurred around 6.3 cal. ka BP, about 500 years later than in NE. Contrary to the ostracod records in the NE, proxy record of surface conditions is limited to the incomplete record of dinoflagellate cyst (dinocyst) assemblages in core Ak-2575 published in Ivanova et al. (2015). Recent Holocene dinocyst records in the Black Sea (e.g. Bradley et al., 2012; Marret et al., 2009; Mudie et al., 2001; Shumilovskikh et al., 2012, 2013; Verleye et al., 2009) have demonstrated the richness in species and detailed environmental changes in salinity conditions. Early-Holocene assemblages show a dominance of brackish species, Spiniferites cruciformis and Pyxidinopsis psilata, although presence of the euryhaline species Lingulodinium machaerophorum indicated marine influence and salinity above 7 psu (e.g. Marret et al., 2009; Mertens et al., 2012). Full modern conditions were reached around 4.1 cal. ka BP.
This study aims to evaluate whether changes in water conditions (amplitude and timing) across the Black Sea basin were contemporaneous during the Holocene as well as to assess the impact of climatic changes in the NE region. To address these issues, we have analysed core Ak-2575 retrieved on the northeastern shelf of the Black Sea (e.g. Ivanova et al., 2012) for its integrated record of pollen and dinoflagellate cysts.
Modern regional setting
The Black Sea is a relatively deep-water basin, with a mean depth of 1315 m, and a maximum of 2258 m at its centre (Figure 1); the sea shelves are in general narrow (only occupying 25% of the basin) with the exception of the NW region where they extend for more than 200 km. Its only connection to the world ocean is through the Bosphorus Strait which is about 30 km long, with a depth varying between 20 and 100 m. The Sea of Azov in the NW region is a relatively shallow basin, with a maximum depth around 15 m and it is connected to the Black Sea via the Kerch Strait. Three major rivers (Danube, Dnieper and Dniester) in the NW region comprise most of the river input (80%); other important rivers are the Caucasian rivers Kodori, Inguri, Rioni and Chorokhi which are classified as midsize rivers with a drainage area from 2000 to 50,000 km2 (Mikhailov and Mikhailova, 2008).
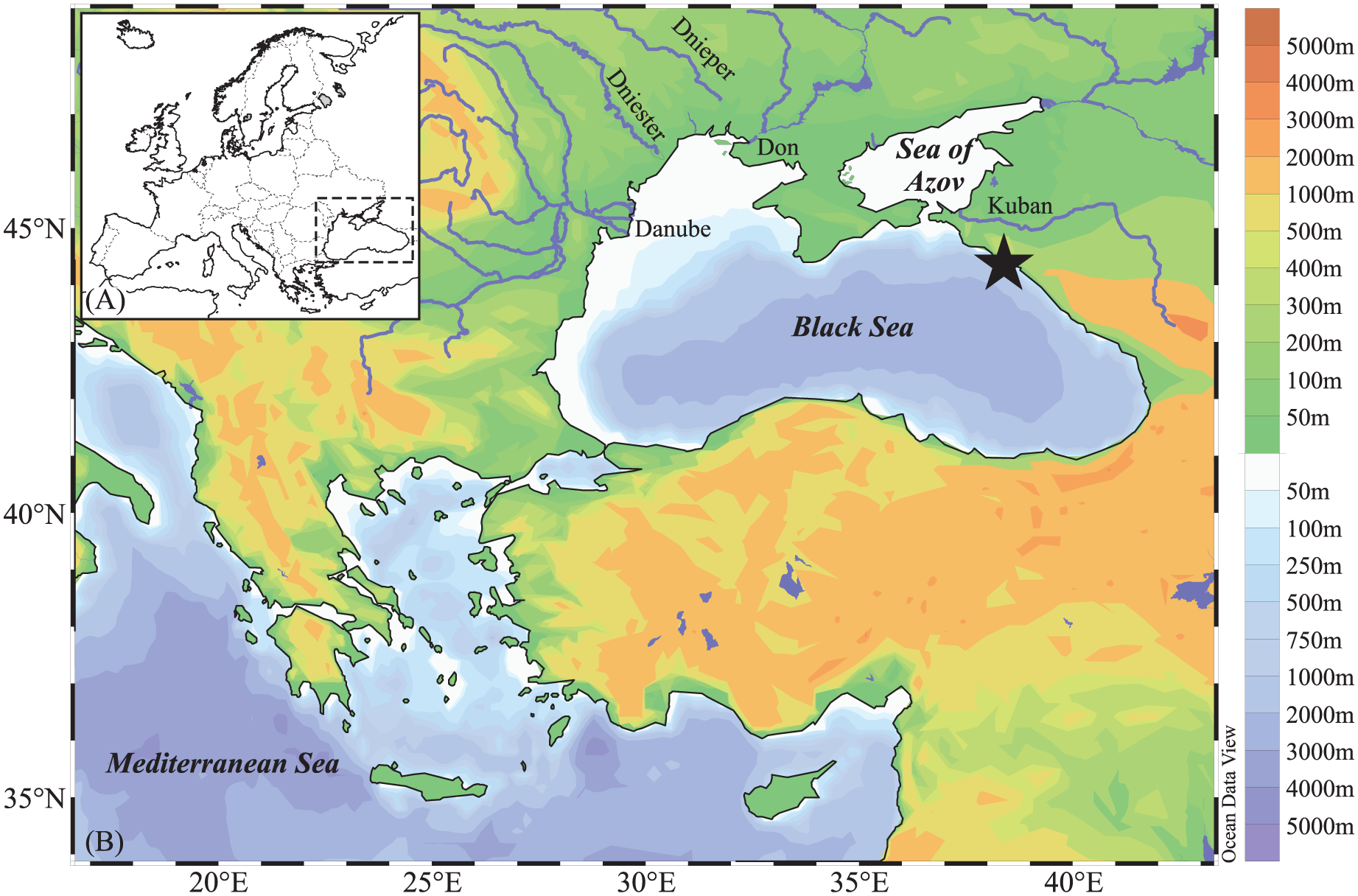
The Black Sea: bathymetry, rivers, core location and relief.
The Black Sea is characterised by a permanent halocline and is strongly stratified, with a well-oxygenated surface water (up to 50 m water depth) and anoxic conditions and high sulphide concentrations from 100 m to the bottom. Annual surface salinity ranges from ~15 psu (in the NW region, near river mouths, and south of the Kerch Strait) to 18–19 psu in the middle of the basin (Figure 2b; Boyer et al., 2013). Bottom salinity is influenced by Mediterranean waters flowing and averages 22 psu. Annual sea-surface temperatures (SST) show a west-east gradient, with the lowest values in the Odessa Bay (~10°C) and the highest off Sochi (~18°C), in the eastern region of the sea (Figure 2a). Movement of the surface water mass is controlled by wind, river input and climate (temperature, evaporation and precipitation). A cyclonic circulation, the Rim Current, is composed of two main cyclonic gyres in the western and eastern regions, respectively, and a number of anticyclonic eddies (Toderascu and Rusu, 2013).
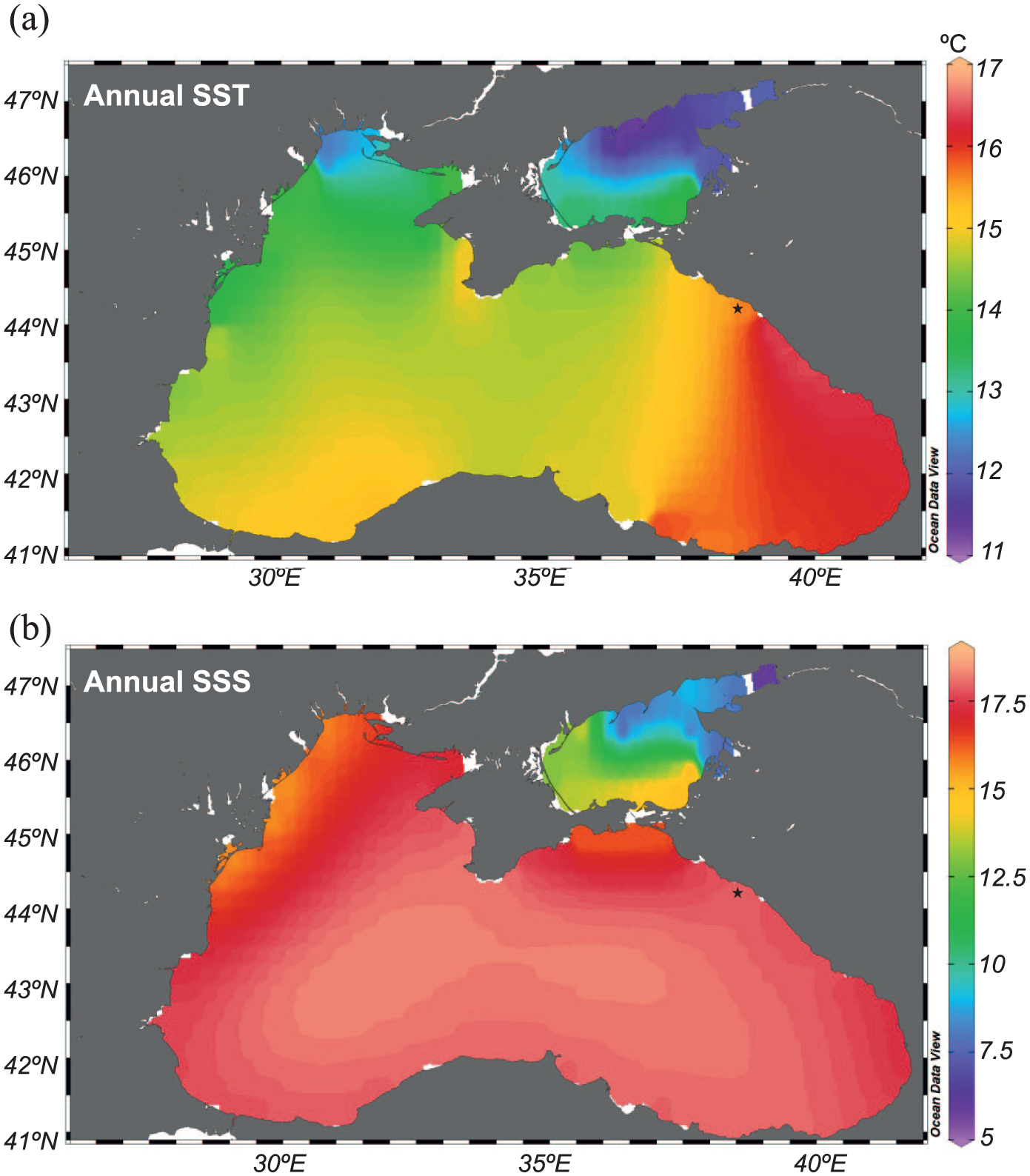
Maps showing annual sea-surface (a) temperature and (b) salinity based on the World Ocean Atlas 2013 (Boyer et al., 2013) and created with Ocean Visual Display software.
Topography around the Black Sea is quite diverse, with plains in the west and north, and mountainous in the east (Caucasus) and the south (Pontic) (Figure 1). The northwestern region is under the influence of a humid continental climate, whereas the southwest region reveals Mediterranean-type climate. The northeastern side is characterised by humid subtropical conditions, and the southern coast is dominated by an oceanic climate (Peel et al., 2007). The studied region is the only coastal region in Russia to have subtropical landscapes as well as humid to semi-arid landscapes (Petrooshina, 2003). Winter temperatures average 3–5°C in winter up to 23–24°C in summer.
The vegetation in the studied region is characterised by the Colchic forest which is a relic of tertiary forest of Western Eurasia (Nakhutsrishvili et al., 2011). A succession can be observed according to altitude, within the lowland area, up to 500 m, a dominance of oak-hornbeam (Quercus pubescens–Carpinus orientalis) forests, accompanied by Fraxinus excelsior (Figure 3). From 300–800 to 1000 m, a mixed sessile oak (Quercus petraea)–hornbeam (Carpinus betulus, C. orientalis) forest is observed. Above this vegetation unit, the Caucasian oriental beech forests dominate the landscape, up to 1800–2000 m. Fagus sylvatica subsp. orientalis is the dominant tree, accompanied by Tilia, Acer, Ulmus, Castanea sativa. This forest also alternates with the west Caucasian fir-spruce forests (Abies, Picea). High altitude areas are covered with grasslands with some patches of Pinus forests, although pine can also be found in lower latitudes as they are eurythermic trees and their presence may be associated with human activities, with sandy and/or stony locations (Bohn et al., 2004).
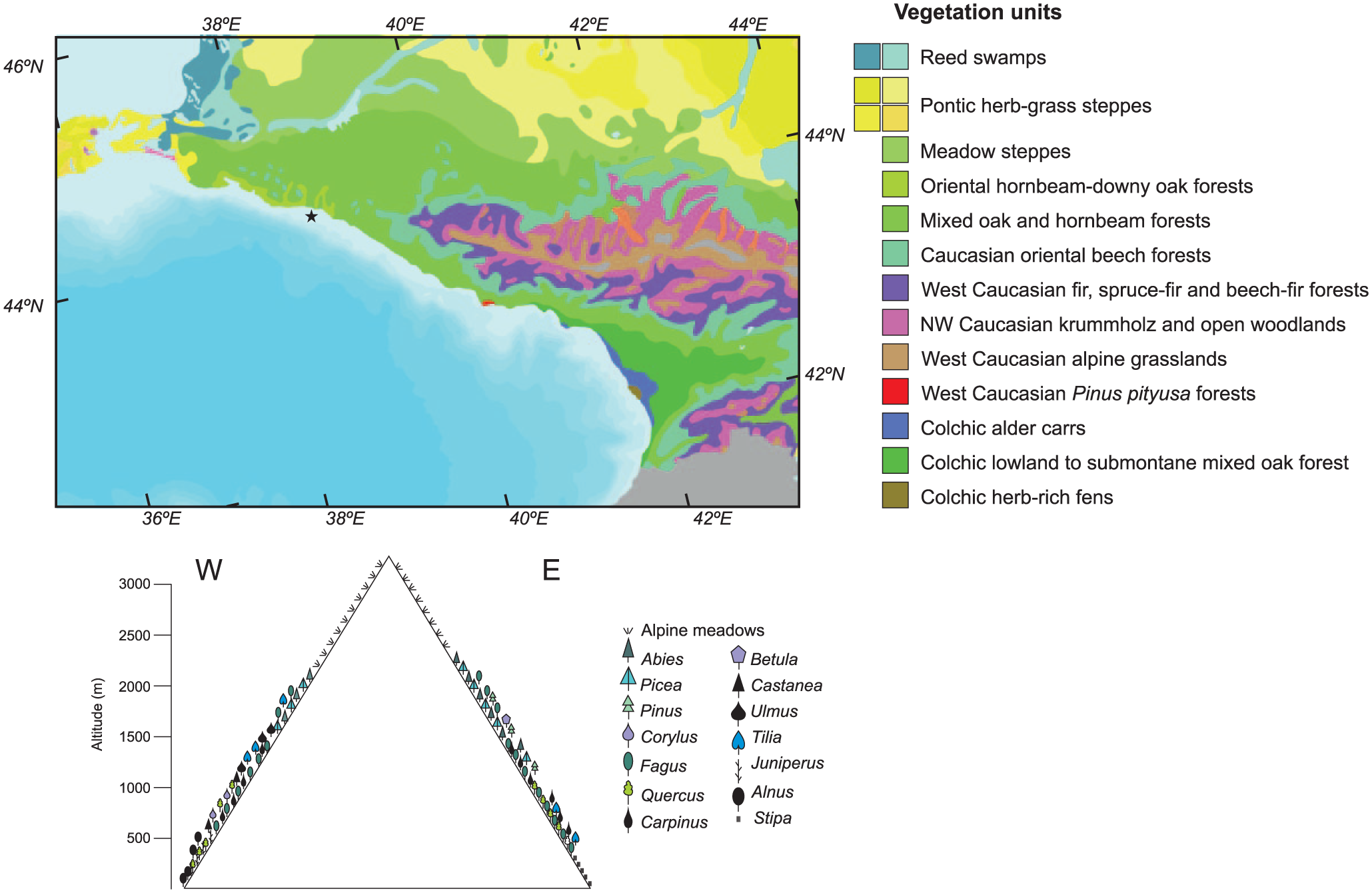
Vegetation map modified from Bohn et al. (2004) with location of core Ak-2575 (star) and vegetation profile from west to east across the Greater Caucasus.
Materials and methods
The 186-cm long sediment core Ak-2575 was collected in 2007 from the NE Caucasian outer shelf (44°13.46′N, 38°38.03′E, water depth 99 m), during the cruise by RV Akvanavt. A semi-liquid surface layer of unknown thickness, ~10 cm, was likely lost during the core Ak-2575 processing. Detailed lithology, an age model based on molluscs only as well as a preliminary dinoflagellate cyst record, can be found in Ivanova et al. (2015). The core was sliced into 2 cm thick samples, and a total of 45 samples were used for palynological analysis.
Age model of core Ak-2575
In total, 18 AMS radiocarbon ages were obtained on samples from core Ak-2575. These include 11 dates on bivalve molluscs and 7 from monospecific ostracod samples (Table 1). For the extraction of the dated material, sediment samples were wet sieved and ostracods were picked out and rinsed several times before being placed in glass vials and dried overnight at 40°C. Samples were analysed at Scottish Universities Environmental Research Centre (SUERC), Keck C-Cycle AMS Laboratory and Poznań Radiocarbon Laboratory (UCIAMS). 14C dates were calibrated using IntCal13 (Reimer et al., 2013). A reservoir correction of 404 ± 91 years for radiocarbon dates <7500 14C years, 300 ± 125 for radiocarbon dates between 7500 and 8400 14C years, and 258 ± 55 years for ages >8400 14C years were applied (see Ivanova et al., 2015). The age-depth model for Ak-2575 (Figure 4) was constructed using Bacon (Blaauw and Christen, 2011) in R (R Development Core Team, 2018) using all samples listed in Table 1. Three hiatuses were inserted into the model using sediment data and seismic profiles presented and discussed in Ivanova et al. (2015). Ages used in this paper are given as mean ages and are reported as calibrated ages BP (0 cal. BP = AD 1950).
Details of radiocarbon dates, calibrated age range (2σ) and details of original publication of radiocarbon dates.


Age-depth model for Ak-2575. Red = ostracods; Blue = molluscs; Purple = two samples overlapping.
Palynological preparation and analysis
Sample preparation follows that of Marret et al. (2009). Volume of sediments was first estimated and followed by the addition of exotic markers (Lycopodium clavatum) enabling to calculate palynomorph concentrations (specimens/ml). Sediments were then treated with cold 10% hydrochloric acid (HCl), followed by cold 40% hydrofluoric acid to remove carbonate and silicate content, respectively. Residues were then treated with 10% HCl to remove fluorosilicates. Samples were neutralised between acid treatments with distilled water. Residues were sieved with a 10-µm mesh, and the larger fraction was mounted on a microscope slide in glycerine jelly stained with safranine. Where possible, a minimum of 100 dinoflagellate cysts and a minimum of 300 pollen grains were counted. Dinocyst taxonomy is based on Marret et al. (2004, 2009). Spiniferites spp. and Brigantedinium spp. comprise of specimens that could not be identified at the species level due to poor orientation or hidden diagnostic characteristics. Specimens of L. machaerophorum were often broken or folded, and they were rare in many samples, so attempting to reconstruct past salinity using the equation from Mertens et al. (2012) would not have been statistically reliable. Pollen identification follows that of Beug (2004) and pollen reference collection at the Department of Geography and Planning, University of Liverpool and Department of Palynology and Climate Dynamics of the University of Göttingen. Dinocyst percentages were calculated on the whole sum of dinocysts, and pollen percentages are based on arboreal and non-arboreal pollen grains; but due to the over-representation of Alnus, the pollen percentages were calculated with a sum excluding Alnus. Spores were excluded from the pollen sum but are presented in the diagram. We also paid attention to occurrence of non-pollen palynomorphs as described in Mudie et al. (2010), but surprisingly they were absent. The biomisation approach applied in this study follows a standard procedure described in Prentice et al. (1996). All terrestrial pollen taxa found in the Ak-2575 record were assigned to the respective biomes following Shumilovskikh et al. (2012). Biome affinity scores were calculated for each sample following the equation published in Prentice et al. (1996).
Examining all biome affinity scores (and not only deriving the dominant biome name) throughout a pollen record for more objective interpretation of reconstruction results has been presented in various papers published elsewhere (i.e. Marchant et al., 2006; Rudaya et al., 2009; Tarasov et al., 2005). For the purpose of the current study, we selected three biomes for further discussion. Temperate deciduous (TEDE) forest biome reveals the highest affinity score through the entire pollen record and is well represented in the natural vegetation of the region (Shumilovskikh et al., 2012). Taiga (TAIG) biome includes the following PFTs, including boreal evergreen conifers (e.g. Picea, Abies), eurythermic conifers (Juniperus, Pinus s/g Diploxylon), boreal summergreen trees/shrubs (e.g. Alnus, Betula, Larix) and shrubs of the heath family (Prentice et al., 1996). This biome is predicted by the BIOME model at high elevations in the central European mountains, where winters are cold and precipitation meets >75% of demand (Prentice et al., 1992). Steppe (STEP) biome includes grass/shrub communities and is limited (under natural conditions) to the lower elevations where climate is too dry and cannot support forest vegetation. Thus, reconstructed changes in the TAIG and STEP biome scores have potential to represent the climatic/altitudinal extremes in the study region.
Results
Chronology
The 14C AMS ages (Table 1) show that site Ak-2575 recovered a section of Holocene sediments with a basal age of ~9800 cal. yr BP. The age-depth model shows four phases of sedimentation history (Figure 4) constrained by three hiatuses The earliest phase of sedimentation (9800–8950 cal. yr BP) suggests a relatively slow accumulation rate in the earliest part of our record. The oldest hiatus (A) occurs between 8950 and 7950 cal. yr BP (164 cm). The second phase of the sediment record occurs between 7950 and 7770 cal. yr BP. At 156 cm, there is an assumed very short break in the record (<100 years, hiatus B) before a third phase of the record between 7770 and 6020 cal. yr BP. The sediment record shows that hiatus B separates Dreissena coquina facies from a mixed mollusc fauna coquina (Ivanova et al., 2015). The newly obtained date at 153 cm supports this hypothesis of a very short break in the record as well as the conclusion of Ivanova et al. (2015) that the dated Dreissena sp. shell with raw age of 8215 ± 30 14C (9030–8430 cal. yr BP) is reworked.
The youngest break in the sediment record occurs between 6020 and 4130 cal. yr BP (42 cm) and is highlighted in Figures 5–7 as hiatus C. From ~4130 cal. yr BP to present, the sedimentation rate slows and the modelled age errors increase. The dated ostracod level at 32 cm (4820–4440 cal. yr BP) is older than the dated bivalve level at 37 cm (3830–3390 cal. yr BP). A similar situation repeats in the youngest section of the core where the dated ostracod level at 9 cm (3390–3240 cal. yr BP) is older than the dated bivalve level at 19 cm (2800–2360 cal. yr BP). The model assumes that core top is AD 2007. This suggests a very slow sediment accumulation rate in the top 9 cm of this core and interpretation of results from this section should be viewed with caution.
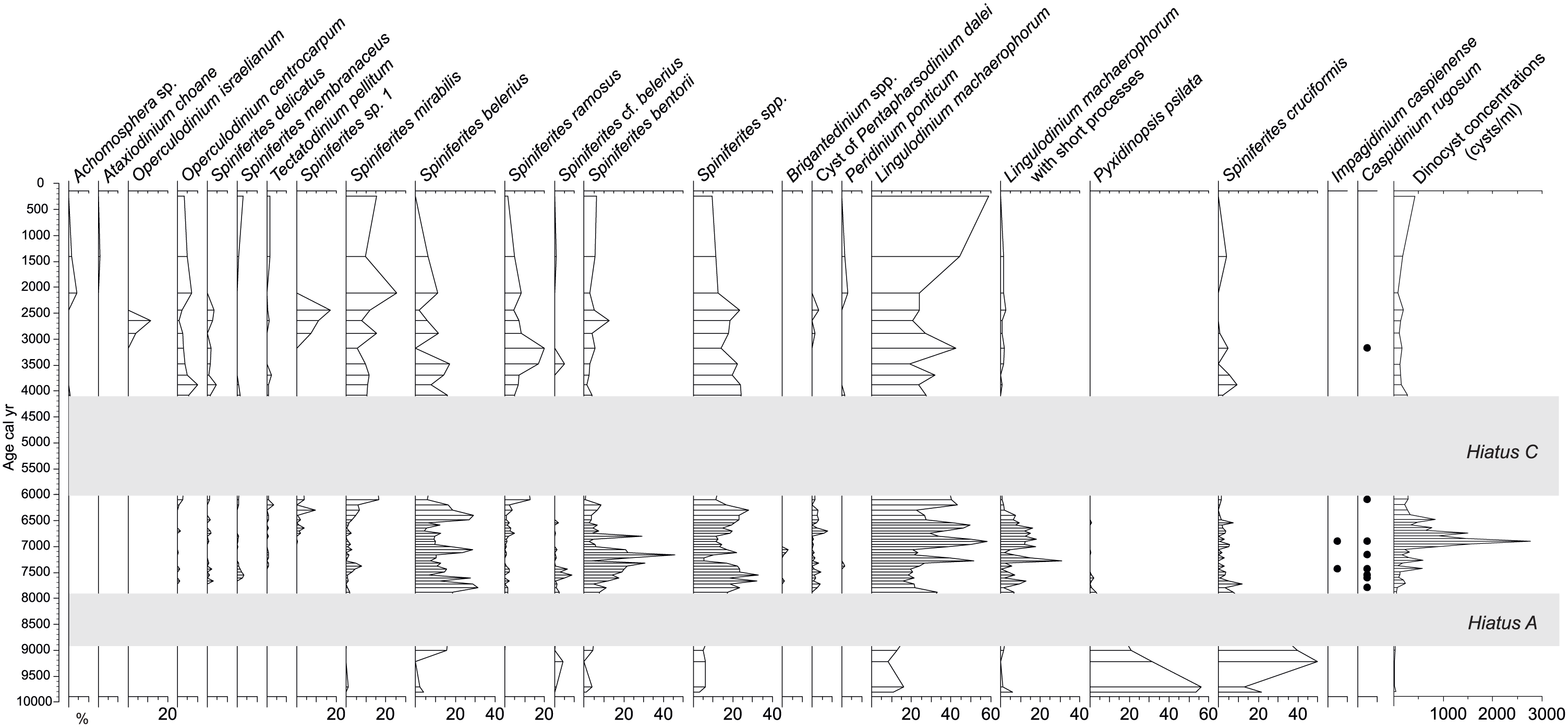
Percentage diagram of dinocyst taxa based on a sum comprising all dinocyst species. Concentration (cysts/mL) curve is depicted on the right side of the diagram. Hiatuses A and C are highlighted with grey bands. Hiatus B is not shown as it is very short-lived (less than 200 years).

Percentage diagram of pollen taxa. Percentages were calculated on a pollen sum that excluded Alnus due to its over-representation in the spectra. Alnus percentages are calculated on a total pollen sum. Pollen taxa are sorted according to vegetation types (trees in green and herbs in yellow). Hiatuses A and C are highlighted with grey bands. Hiatus B is not shown as it is very short-lived (less than 200 years).

Summary pollen and dinocyst diagram. Lowland taxa (Alnus and Corylus), Pinus, arboreal vegetation types (as grouped in Figure 6), numerical scores of selected biomes (steppe, taiga and temperate deciduous forest) and selected dinocyst species indicator of water conditions are presented. Thermophilic group is represented by Operculodinium israelianum; Mediterranean taxa include Operculodinium centrocarpum, Spiniferites delicatus, Spiniferites membranaceus, Spiniferites mirabilis, Spiniferites ramosus, Achomosphaera sp., Ataxiodinium choane; Euryhaline taxa include Lingulodinium machaerophorum and its morphotypes, Spiniferites belerius, Spiniferites bentorii; Caspian/brackish taxa consist of Spiniferites cruciformis, Pyxidinopsis psilata, Impagidinium caspienense and Caspidinium rugosum.
Dinoflagellate cyst assemblages
Dinocyst assemblages of core Ak-2575 are relatively diverse, with a total of 20 taxa, dominated by L. machaerophorum and Spiniferites species (Figure 5). Concentrations (cysts/ml) are overall relatively low, with an average 4185 cysts/ml, ranging from 140 to 27,599 cysts/ml. Three main assemblages can be observed between the two main hiatuses (A and C). Prior to hiatus A, from ~9.8 cal. ka, the assemblages are dominated by two species of Caspian/brackish origin, P. psilata and S. cruciformis. Accompanying taxa are L. machaerophorum, Spiniferites belerius and Spiniferites bentorii. Cyst concentrations are at their lowest during this interval, with fewer than 330 cysts/ml.
Between hiatus A and hiatus C, the species dominance has shifted to L. machaerophorum, in particular its morphotype with short processes. S. belerius and S. bentorii are second. Species of Mediterranean origin are occurring in low frequencies, such as Operculodinium centrocarpum, Spiniferites delicatus, Spiniferites mirabilis and Tectatodinium pellitum. This interval also sees the disappearance of P. psilata while S. cruciformis is still present, with low percentages (up to 7.5%). Interestingly, this interval contains the only occurrences of Impagidinium caspienense as well as sporadic presence of Caspidinium rugosum. Cysts of Pentapharsodinium dalei appeared here for the first time in the record. Concentrations reached their highest values during this interval, with a maximum (27,599 cysts/ml) around 6.9 cal. ka.
The assemblages above hiatus C show that S. cruciformis still occurs, albeit in low percentages, similarly to the previous interval. L. machaerophorum still dominates this interval, but we note the very low occurrences of its morphotype with short processes. Mediterranean taxa occur in higher percentages compared with the previous interval; Operculodinium israelianum briefly appears between 2.9 and 2.3 cal. ka. An unknown taxon of Spiniferites, Spiniferites sp. 1, also temporarily occurs between 2.9 and 1.7 cal. ka. This cyst is of ovoidal shape, sometimes slightly cruciform, with a paratabulation expressed with very developed sutural septa between all processes. The body size is around 40 by 32 microns and process length ~12 microns (Plate 1). The last 2000 years sees the occurrence of euryhaline taxa, Achomosphaera sp. and Ataxiodinium choane. Concentrations are lower than the previous interval, averaging 1834 cysts/ml.

Specimens of Spiniferites sp. 1 from core Ak-2575: depth 12 cm (1–2, 4), depth 16 cm (3) and depth 44 cm (5–6). Scale bar represents 10 microns.
Pollen assemblages and reconstructed biome scores
A total of 44 pollen taxa were identified and are overall dominated by tree species, in particular by Alnus and Pinus, followed by Quercus, Fagus and Corylus (Figure 6). Concentrations are higher compared with those of dinocysts, ranging from a minimum of 862 grains/ml to a maximum of 56,413 grains/ml, and averaging 15,770 grains/ml. Similarly to the dinocyst record, three types of assemblages can be observed between the two main hiatuses A and C. From the bottom of the record to hiatus A, Pinus and Quercus dominate pollen spectra and are accompanied with low occurrences of Carpinus, Fagus and Corylus. Concentrations are the lowest of the whole record, with a maximum of 9704 grains/ml. Herb taxa are present, with a low diversity (13 species of a total of 21 herb taxa) and mainly comprised of Artemisia, Poaceae and Amaranthaceae.
Between hiatus A and hiatus C, Fagus and Corylus dominate the tree taxa, whereas Quercus occurs similarly as in the previous interval. Carpinus and Alnus have their highest occurrence of the whole record. Picea is recorded only between ~7 and 6 cal. ka. Ulmus and Abies record their highest percentages from the whole spectra from 8 to ~7 cal. ka BP. Ostrya is observed only in this interval. Fraxinus is present throughout this interval up to ~6.5 cal. ka BP, always in low percentages. Herb taxa diversity is at its highest and is still dominated by Artemisia, followed by Poaceae then by Amaranthaceae.
From hiatus C to the top of the core, tree pollen is dominated by Pinus, and secondarily by Alnus and Quercus. There are rare occurrences of Picea around 3 and 2 cal. ka. Ulmus and Carpinus have their lowest relative abundance compared with the two previous intervals. Significant percentages of Taxus are observed from 2 cal. ka upwards.
The biome score record (Figure 7) demonstrates that pollen taxa representing the low-elevation TEDE forest have a maximum contribution to the pollen assemblages of the analysed core. A progressive decrease in the TEDE biome scores is recorded since 4 cal. ka BP, accompanied by an increase in the STEP biome scores, suggesting an opening of the landscape during this period forced by either climatic factors and/or human activities. The TAIG scores reflecting pollen contribution from the boreal evergreen and deciduous forest taxa show rather minor fluctuations, suggesting greater stability of the higher elevation forest belt.
Discussion
Contemporaneity of environmental change across the Black Sea basin during the Holocene?
Two of the main debates surrounding environmental change in the Black Sea during the Holocene period are (1) baseline conditions around 10–9 cal. ka BP and (2) subsequent rates of change in sea level and salinity. This study suggests that on the NE Black Sea shelf, the baseline conditions consisted of brackish water (>7 psu), sea level and salinity began to rise after ~9.6–9.0 cal. ka BP; rates of change were gradual (>1000 years) as shown by the slow decline/disappearance of the Caspian/brackish species (Figure 7). Similar findings were observed with the benthic bivalve community (Ivanova et al., 2007, 2012, 2015; Zenina et al., 2017). The dinocyst species composition from core Ak-2575 is consistent with other recent Holocene records from the Black Sea (Bradley et al., 2012; Marret et al., 2009; Shumilovskikh et al., 2013; Verleye et al., 2009), albeit with a lower diversity compared with the records from the SW. It is interesting to note that our record contains only two heterotrophic taxa, with Brigantedinium spp. and Peridinium ponticum. This raises the possibility of a preservation issue for the heterotrophic species as suggested by Zonneveld et al. (2008); however, the anoxic character of the Black Sea suggests that other parameters may explain this paucity of heterotrophic species. It could be that the prevalence of lower productivity conditions in the eastern region compared with the west would have favoured the dominance of autotrophic species, which is the case at present day (Mudie et al., 2017). S. cruciformis and P. psilata are the main components prior to ~9.8 cal. kyr BP (Figure 5) and highlight the dominance of Caspian/brackish taxa (Figure 7). There is a secondary influence of climate on sea-surface conditions with thermophilic taxon being recorded ~2.5 cal. ka BP. Human impact has limited effect on the sea-surface conditions with no toxic dinocysts such as cysts of Gymnodinium catenatum recorded unlike in studies from other areas of the basin such as the southwestern shelf (Bradley et al., 2012; Marret et al., 2009). These cysts were thought to have been introduced from ship ballasts about 3000 years ago.
Baseline (10–9 cal. ka BP)
Although there is a broad agreement as to when the reconnection took place, around 9.6–9.0 cal. ka BP, a dispute remains with regard to the surface water level of the Black Sea prior to the reconnection. According to Yanchilina et al. (2017), the Black Sea palaeoshore was about 120 mbsl, around 9.3 cal. ka BP, with a rapid infilling of the basin, in less than 40 years, isolating this sea from the global ocean. On the opposite side, the recent study of Aksu et al. (2016) demonstrated that there was a strong Black Sea outflow in the Marmara Sea in the early-Holocene, suggesting that the sea level was either at the Bosphorus sill level (currently at 35 mbsl) or slightly higher. Both sets of evidence rely on geophysical, geochemical and micropalaeontological data as well as radiocarbon dated material. The sedimentation record (see Ivanova et al., 2015) from our site shows that the NE shelf was water covered from at least 10 cal. ka BP (Figure 4). There is no evidence of reworked palynomorphs and ostracods (Zenina et al., 2017) at this site which further supports that the area was inundated prior to 10 cal. ka BP.
Another disagreement is the salinity condition of the Black Sea prior to its reconnection, with Ryan and co-workers (e.g. Ryan, 2007; Ryan et al., 2003) supporting the idea of a freshwater environment, whereas other studies suggest brackish conditions (e.g. Marret et al., 2009; Mertens et al., 2012; Mudie et al., 2001). Ivanova et al. (2015) and Zenina et al. (2017) used mollusc and ostracod data to show that the benthic conditions were brackish in this time period, estimating salinity to be around 7 psu. The dinocyst record, especially the presence of L. machaerophorum and S. mirabilis, supports this conclusion and suggests that the surface salinities were also a minimum of 7 psu prior to 9.6–9.0 cal. ka BP. This is similar to the conclusions from studies examining surface conditions (e.g. Bradley et al., 2012; Bradley and Marret, 2013; Marret et al., 2009; Mertens et al., 2012) and benthic conditions (e.g. Williams et al., 2018) on the southwestern shelf.
Transition to modern-day conditions
Perhaps the most striking finding from the dinoflagellate cyst analysis is the occurrence, albeit in low relative abundance, of Ponto-Caspian species in the mid- to late-Holocene. This is in contrast to the findings of Marret et al. (2009) and Bradley et al. (2012) whose analysis of Holocene sediment on the southwestern shelf of the Black Sea showed the species such as S. cruciformis and P. psilata only sporadically occurred after 6000 cal. yr BP. Verleye et al. (2009) found from core GeoB7625-2 taken from a deeper water site at −1242 m that both species were also present, again in a very low occurrence. The modern distribution of these species shows that they are currently present in the Black Sea and Caspian Sea (Mudie et al., 2017).
This leads to two possible explanations. The first and most likely is that current ecological tolerance of S. cruciformis is broader than previously assumed. Mudie et al. (2017) using 45 sites show that the current distribution extends across the Marmara Sea, Black Sea, Caspian Sea and Aral Sea. Previously, this species was thought only to be present in low salinity (4–15 psu) due to its modern occurrence in the Caspian Sea. The finding of the cyst in the freshwater Lake Sapanca (Leroy and Albay, 2010) showed it to be tolerant to freshwater/brackish conditions, and thus meant it was unable to be used in the freshwater versus brackish debate of early-Holocene Black Sea salinities (Hiscott et al., 2007; Soulet et al., 2010). P. psilata has been identified in recent sediments in low percentages (<4%) mostly in the Black Sea, and some rare presence in the Marmara Sea and Caspian Sea. However, it has been observed in abundance during the late-Pleistocene to mid-Holocene period in the Caspian Sea (Leroy et al., 2007, 2013, 2014), where conditions were more saline than present-day conditions.
A second explanation may be linked to SSTs. Besides SSS, SST may play a role in the occurrence of S. cruciformis during the mid- to late-Holocene in the NE. Its current modern distribution indicates that it can tolerate a relatively wide range of annual SST (12–20.5°C), with peak abundance in the warmest waters. Warmer conditions are also suggested with relatively high occurrences of the thermophilic species S. mirabilis from 6.1 cal. ka BP onwards. A possible maximum of warm conditions may have occurred between 3.0 and 2.5 cal. ka BP, as highlighted by the occurrence of O. israelianum. This species has not been seen in modern sediments from the Black Sea nor the Caspian Sea and mainly occurs in waters where winter SSTs are above 14.3°C and summer SSTs are more than 24.2°C, and can tolerate a salinity range between 30.3 and 39.4 psu (Zonneveld et al., 2013). Mudie et al. (2017) have observed modern distribution of O. israelianum only in the Marmara Sea, where SSTs are close to the range that this species lives in but with SSS 10 lower.
This warm episode is in agreement with the warm climatic conditions suggested by pollen records in the Taman Peninsula between 2.9 and 1.5 cal. ka BP (Bolikhovskaya et al., 2018) but has not been recorded in the west by the dinocyst record.
Another evidence of contemporaneity across the Black Sea basin is a significant peak of the thermophilic species S. mirabilis at ~2.0 cal. ka BP, observed in our core as well as in cores MAR05-13 (Bradley et al., 2012) and 22-GC3 (Shumilovskikh et al., 2013). Subtle peaks of this species are also observed in cores MAR02-45 (Marret et al., 2009) and GeoB7625-2 (Verleye et al., 2009). Pollen records in the Taman Peninsula do suggest warm and humid conditions around this date (Bolikhovskaya et al., 2018).
Establishment of present-day conditions may have happened within the last 1500 years, but the low-resolution sampling at the top of the core prevents us to exactly pinpoint this change. However, our dinocyst assemblage indicates cooler conditions with the decrease of S. mirabilis.
Changes in vegetation
Core Ak-2575 provides the first Holocene pollen record for the NE of the Black Sea, albeit with the hiatuses, and reveals some subtle but significant changes in the vegetation source. There is no significant river discharge into the NE Black Sea nearby the core location (cf. Figure 1), and therefore all the pollen is assumed to be from wind dispersal and reflects a regional signal (e.g. Cordova et al., 2009). At present, the dominant wind blows from the NE (Zecchetto and De Biasio, 2007), and therefore the pollen is assumed to be from the eastern edge of the Black Sea and the Greater Caucasus mountains. This complex pollen source area means that pollen is coming from both lowland coastal areas and mountainous areas of the Caucasus.
Within the pollen source area at present, the NE region is currently dominated by the Oriental hornbeam (C. orientalis) and mixed oak forests, which is reflected in the pollen record with the relative high occurrence of Quercus but a low representation of Carpinus at the top of the core. The high percentages of Alnus are likely transported from coastal areas. Biome reconstructions show that the TEDE biome dominates the record but overall declines from the bottom of our record.
During the early-Holocene, our pollen record indicates the dominance of Pinus taxa, with some evidence of a mixed oak-hornbeam-beech forest, suggesting relatively cool and dry conditions. At present, no other vegetation records from adjacent regions can be compared with our data. Further south to our study site, pollen data from Georgia (e.g. Connor and Kvavadze, 2009; Joannin et al., 2014) reflect lower humidity. Reconstructed climate conditions based on pollen spectra in Europe (Mauri et al., 2015) suggest overall cooler and drier conditions during the time interval prior to hiatus A in our core.
The period between hiatuses A (~7900 cal. yr BP) and C (~6000 cal. yr BP) shows a steady ratio for the tree/shrub/herb ratio (Figure 6), but the changes in the relative abundance of the tree and herbs (green and yellow curves, respectively, in Figure 6) suggest large-scale changes in vegetation of the eastern Black Sea shore and the Greater Caucasus mountains. Significant increase of Carpinus, Fagus and Corylus accompanied by the decline of Pinus, but also the presence of Ostrya and Ulmus, suggests the development of a thermophilous deciduous forest, indicating an overall warming trend and humid conditions. Such climate conditions were also recorded in the Taman Peninsula (Bolikhovskaya et al., 2018) and further south (Connor and Kvavadze, 2009). Interestingly, this onset of warm/humid conditions coincides with the establishment of a mixed Caspian–Mediterranean benthic fauna observed in our core (Ivanova et al., 2015).
Following hiatus A, from around 4 cal. ka BP, a remarkable dominance of Pinus pollen among the tree taxa, accompanied by higher abundance of herb taxa and a steady increase in the STEP biome scores, suggests an opening in the landscape due to possibly a combination of drier/warmer conditions and human impacts. This landscape opening more or less coincides with the onset of the Meghalayan Age. However, our dinocyst record suggests the onset of warmer water conditions around 6.1 cal. ka BP, prior to hiatus A. It is worth noting that the peak of thermophilic dinocyst taxa (Figure 7) corresponds to a clear decrease of TEDE biome scores, suggesting that, most likely, changes in the vegetation may have been triggered by climatic conditions rather than human activities around 3–2.5 cal. ka BP. The noticeable occurrence of the taxon Taxus, a taxon which belongs to the intermediate temperate conifer PFT and is related to mild winters (i.e. mean temperatures of coldest month between −2°C and +5°C: Prentice et al., 1996), around 2 cal. ka BP is concomitant with the warm water conditions suggested by the peak of S. mirabilis across the basin. The increase in the STEP biome scores during the last 2000 years may reflect a combined effect of the warmer temperatures (i.e. increased evaporation) and intensified human impact (i.e. opening of the landscape and spread of grassland/meadow vegetation). Although humid and warm conditions were reconstructed further north (Bolikhovskaya et al., 2018), their moisture reconstruction may represent local situation in the Kuban River delta rather than the regional trend.
Conclusion
We provide here the first integrated record of climatic conditions and terrestrial environmental changes from the NE region of the Black Sea for the Holocene period. Despite the presence of two hiatuses, the dinocyst record has enabled us to confirm the contemporaneity of the major changes that have occurred prior, during and after the reconnection of the Black Sea with the world ocean. The dominance of the Caspian/brackish species, S. cruciformis and P. psilata, accompanied with the euryhaline species L. machaerophorum and other Spiniferites species, supports the theory that the Black Sea was brackish, with a minimum of surface salinity of 7 psu prior to the reconnection. The gradual change in dinocyst species composition and their abundance further corroborates previous findings from the NE and SW for a gradual infilling of the Black Sea rather than a catastrophic flooding. The relative abundance of thermophilic species highlighted that the eastern region was warmer than the west around 3.0–2.5 cal. ka BP, which reflects the current gradient of SST in the Black Sea. Contrary to the western region, our record does not contain any toxic dinoflagellate species, which suggests a limited impact of human activities in this region.
In parallel, our pollen record also provides the first regional biome reconstruction for the NE Black Sea and shows that during the early-Holocene, the vegetation was responding to the regional climatic conditions, cooler and drier. The onset of warm and humid conditions around 7.9 cal. ka BP is recorded by the establishment of a TEDE forest; this biome is gradually changed towards an increase of open landscape and Pinus after 4 cal. ka BP likely due to a combination of climatic conditions and human activities.
Footnotes
Acknowledgements
We would like to thank Alan Henderson with the preparation of the samples for the palynological analysis. We are grateful for the constructive comments from two anonymous reviewers which helped to improve our manuscript.
Funding
This study was supported by Leverhulme Trust (FM and LRB; project ‘The Black Sea environmental conditions during the Meso- and Neolithic period’, F/00 025/AN), project 0149-2019-0007 by Shirshov Institute of Oceanology RAS (EVI, IOM and MAZ) and by the NERC Radiocarbon Facility NRCF010001 (allocation number 1729.1013; FM).