
Abstract
Silt-rich and clay-rich vesicular horizons (V horizons) that develop beneath desert pavements limit infiltration and plant-available soil moisture. A radiocarbon date from buried charcoal constrains the timing of emplacement of a Sonoran Desert alluvial fan deposit to the very end of the Pleistocene (approximately 12.7 ka). The surface of the fan is dotted with 1.5–3 m light-colored plant scars that mark places once occupied by larger perennial plants. A few very large, ring-like clones of Larrea tridentata have persisted on the surface and represent plants that may have initially established as long ago as the early Holocene. The gravelly fan deposit initially would have possessed high infiltration capacities and fostered the establishment of vegetation. Two principal sets of factors have driven vegetation change over the course of the Holocene. During the first part of the Holocene, differences in life history attributes of various species (dispersal and colonization vs. competitive abilities) would have contributed to a succession of perennial plant occupants, culminating with the predominance of the slow-growing, long-lived, but competitively dominant L. tridentata. During the latter part of the Holocene and continuing to the present, continued development of the pavement and associated V horizon has increasingly diminished the capacity of the surface to absorb precipitation and is responsible for the more recent phase involving progressive elimination of vegetation. Development of soil features that markedly influence soil hydrology is a major component of vegetation change during the late Quaternary in the more arid portions of the Sonoran and Mojave deserts.
Introduction
During the last 50 years, investigations of plant macrofossils preserved in ancient woodrat middens have generated a detailed picture of late-Quaternary vegetation changes in what now are the hot deserts of the American Southwest (Betancourt et al., 1990). In response to major climate change of the Pleistocene–Holocene transition, lower elevation occurrences of pinyon pine, junipers, and other relatively mesic woodland species shifted upward approximately 1000 m and desertscrub species occupied the landscape.
In addition to its direct influences on plant distribution, climate also influences soil-forming processes. Precipitation and temperature regimes strongly control weathering, downward translocation of materials, and soil horizon formation. Climate also exerts complex, indirect effects on soils by influencing the kinds of organisms that occur and their effects on soils (e.g. quantity and type of organic matter additions, bioturbation). Climate can also influence parent material additions. For example, deposition of aeolian sediments (loess) in response to climate changes contributes significantly to soil features (McFadden, 2013; Simonson, 1995; Wells et al., 1987).
Certain soil characteristics that have developed in the Sonoran and Mojave deserts under the more arid conditions of the Holocene strongly influence soil hydrological behavior and, therefore, plant and vegetation responses. The purpose of this paper is to present information on soil horizon and pavement development during the Holocene in gravelly alluvium deposited at the very end of the Pleistocene. A radiocarbon date from charcoal buried by this alluvium provides rare numerical age control for determining the pace at which soil conditions develop that have contributed significantly to late-Quaternary vegetation changes in the region.
Study area and methods
Site location
The study area is centrally located in the Sonoran Desert, 28 km southwest of Ajo, Arizona in the northwestern corner of Organ Pipe Cactus National Monument, along Growler Wash, a major ephemeral stream (Figure 1). The upper section of Growler Wash and a similar-sized tributary, the Cuerda de Leña drain the wide Valley of the Ajo located between the Ajo Range on the east and the Bates and Growler Mountains to the west. Those two major trunk streams join at the northern end of the Bates Mountains and pass through Growler Canyon (Figure 1). After exiting the canyon, Growler Wash passes in a southwesterly direction through an area of low rhyolitic hills before debouching on the wide basin floor of Growler Valley. The study area is located at 370 m elevation in the eastern end of the Growler Valley, approximately 7–10 km downstream from the outflow (western) mouth of Growler Canyon and 2 km downstream from the area of low-relief rhyolitic hills. The area of the Growler Wash drainage basin above the study area is 921 km2 (Figure 1).
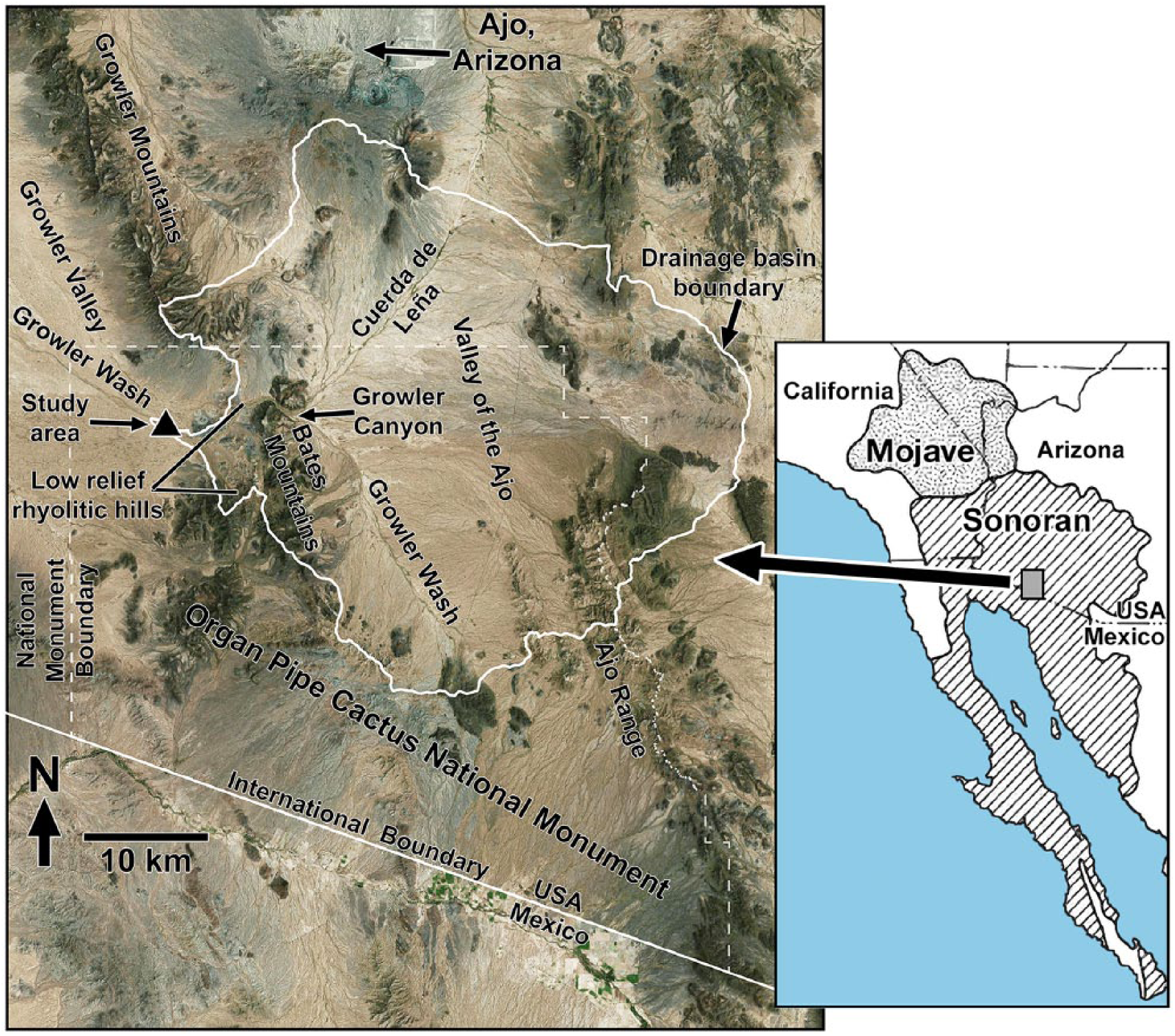
Location of the Grower Wash study area in southwestern Arizona. Detailed map on left includes place names used in text. The study area is shown as a solid black triangle in the left center, and boundary of the Growler Wash drainage basin above the study area is mapped with a white line.
Average annual precipitation at the site from 1982 to 2017 was 202 mm, recorded by a gauge located 0.75 km from the study area (no records for 1999 and 2012). During the first half of this period (1982–1998), the annual average was 249 mm, reflecting the frequent occurrence of moderate-to-large ENSO events during that interval. That amount contrasts with an annual average of 155 mm from 2000 to 2017.
Methods
Fluvial geomorphic surfaces within the 2.5 × 1 km study area (Figure 2a) were identified on aerial imagery based on vegetation characteristics, surface attributes (e.g. the presence of desert pavements), and readily recognized, sharp boundaries. The fraction of the study area occupied by relatively barren desert pavements was measured by overlaying a grid of 9216 points on aerial imagery and determining the fraction of points that occurred within pavement-covered areas. Field verification of designations of geomorphic surfaces was conducted in May, 2015 and elevation differences of adjacent surfaces were measured with a vertical measuring staff and sighting level. Size distribution of clasts on the pavement surface was determined by placing a 2-m measuring stick transversely in five locations, spaced 10 paces apart, and recording the diameter of the clast (major axis) below each 10-cm interval on the stick, for a total of 100 clast measurements.
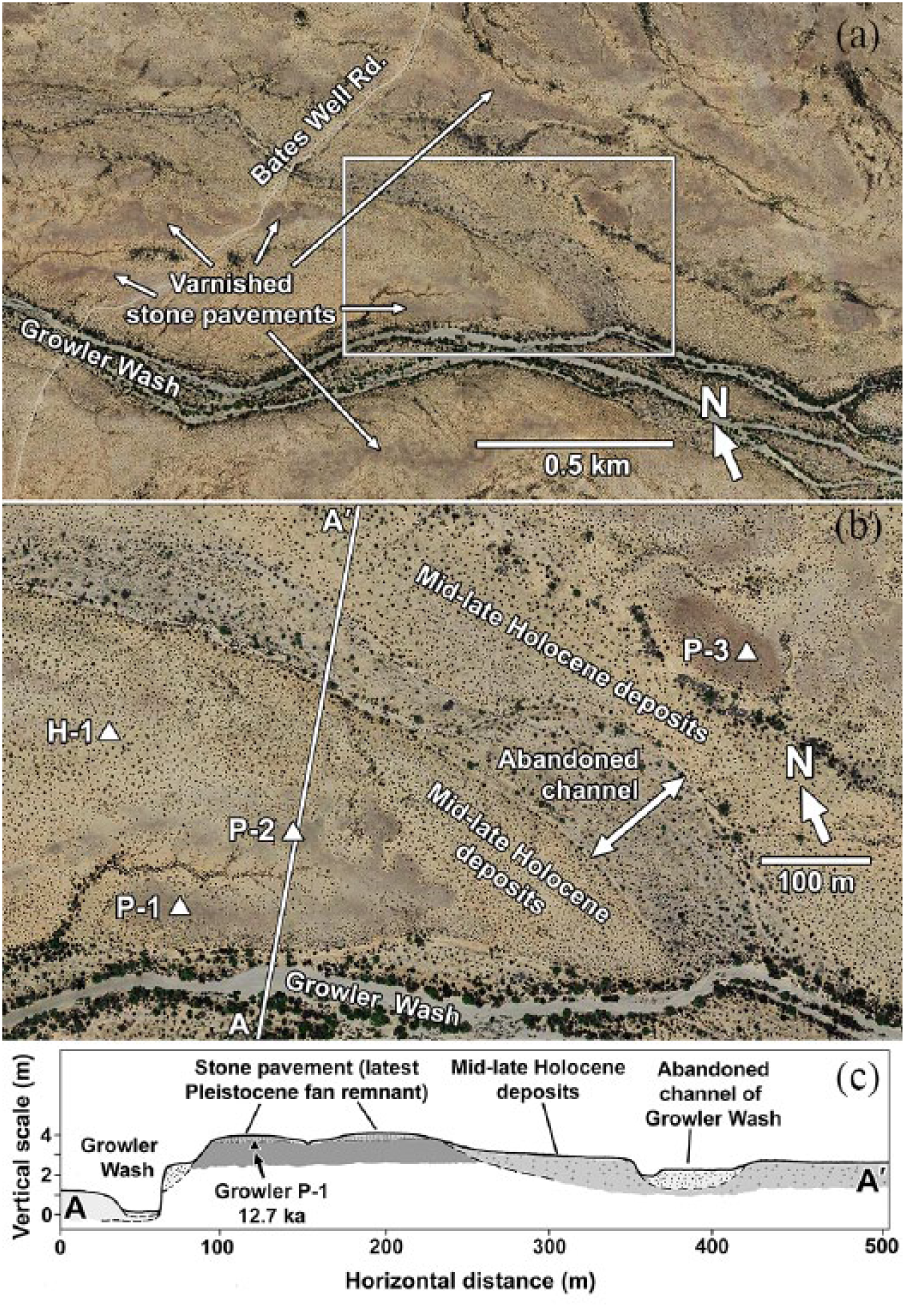
(a) Overview of the study area in Growler Valley. The area enclosed in the white rectangle is enlarged in (b) (Google Earth™ imagery). (b) Enlarged view showing different alluvial surfaces and locations of soil profile descriptions from excavated pits (solid triangles). Geographic coordinates of the pits are included in Table 1. The line A-A’ shows the location of the cross-section diagrammed in (c) (Google Earth™ imagery). (c) Cross section of A-A’ shown in (b).
Soil profile descriptions, Growler Wash study area. Geographic coordinates are locations of soil pits.

Structure – 1: weak; 2: moderate; f: fine; m: medium; c: coarse; sbk: subangular blocky; pl: platy; gr: granular; (ve): vesicular; m: massive.
Dry consistence – so: soft; sh: slightly hard; h: hard.
Wet consistence – ns: non-sticky; ss: slightly sticky; np: non-plastic, sp: slightly plastic.
Textural class – LS: loamy sand; SL: sandy loam; L: loam; SCL: sandy clay loam; CL: clay loam; SiL: silt loam.
Rock + gravel (visual estimate).
HCl reaction (effervescence) – ne: non-effervescent; e: slightly effervescent; se: strongly effervescent; ve: violently effervescent.
Carbonate accumulation stage (Gile et al., 1966) – 0 = absent; I–IV = Stages I–IV.
Horizon lower boundary – a: abrupt; c: clear; g: gradual; s: smooth.
Perennial plant species on various surfaces was recorded in May 2015 and canopy cover of dominant species (% of total area) was estimated visually. Large, ring-like clones of Larrea tridentata were identified on Google Earth™ imagery and subsequently located and examined in the field. Maximum diameters of clonal rings were measured with the Google Earth™ measuring tool to the nearest 0.1 m. Diameters of light-colored plant scars on pavement surfaces (McAuliffe and McDonald, 2006) were similarly measured.
Soils were described in February 2003. Soil pits were dug by hand and profiles were described according to standard methods and nomenclature of Soil Science Division Staff (2017). Bulk samples were collected from each horizon for laboratory analyses. After air drying, the samples were oven dried for approximately 8 h at 50°C. Dried samples were gently crushed with a mortar and pestle and passed through a 2-mm sieve. The combined silt + clay content of the fine-earth (<2 mm) fraction was measured using the hydrometer method with a reading taken at 40 s (time required for settling of the >0.05-mm sand fraction). Due to limitations of the hydrometer method in accurately separating silt and clay fractions (Gee and Bauder, 1986; Karkanis et al., 1991), the combined silt + clay contents are reported in this paper. Total dissolved salts were measured by making a deionized water extract and measuring the conductivity of the filtrate. Volumetric soil samples (50 mL, tapped lightly to settle the contents) were added to 100 mL deionized water and were periodically shaken and allowed to stand for a minimum of an hour before filtration through filter paper. Electrical conductivity was measured with a LaMotte Multi-range Conductivity Meter (Model DA-1); total dissolved salts in the filtrates were calculated following procedural directions for agricultural soil testing that accompanied the instrument (ppm total dissolved salts = microsiemens/cm × 0.7).
Fragments of wood charcoal were exposed in the face of one soil pit. Stratigraphic positioning of the charcoal with respect to soil horizon boundaries was described, measured, and sketched in the field. Radiocarbon AMS dating was conducted by Beta Analytic Inc. (Beta-410908), and the age reported in this paper is the calibrated calendric date (yr BP), using the calibration database of Reimer et al. (2013). Radiocarbon dates that were originally reported in other papers as radiocarbon years BP were converted to calendric years BP using the online calibration calculator (http://www.calpal-online.de) in order to ensure accordant temporal comparisons. A reference collection of stems of 26 species (22 genera) of shrubs and small trees was gathered from sites ranging in elevation from that of the study area (370 m) to approximately 1370 m in the eastern Mojave Desert of Clark County, Nevada. The samples were ignited using a propane torch and partially combusted to create reference charcoal samples for anatomical comparisons with the buried charcoal.
During the 2003 fieldwork, the rain gauge near the study area recorded a major frontal precipitation event of 52 mm on 12–13 February. On 21 February 2003, vertical depths of soil wetting fronts were measured to the nearest centimeter in soils of different geomorphic surfaces. Measurements were taken from fresh, vertical excavations made with a spade. The boundaries of wetting fronts were very clear (<1 cm width) and reliably observed and measured. Multiple measurements were made on different surfaces, and excavations were made at 20 pace intervals along linear transects.
Results
Topographic and stratigraphic relationships of fluvial surfaces
Four fluvial geomorphic surfaces were identified in the study area. The oldest is represented by a few isolated remnants, each approximately 100 m across (referred to as late-Pleistocene fan remnants; Figure 2b). They appear darker than surrounding areas in aerial images due to thicker, more continuous varnish coatings on pavement clasts. A more areal extensive latest Pleistocene fan surface is inset approximately 1 m below the uppermost surfaces of the isolated late-Pleistocene remnants. Age differentiation of this surface from the somewhat older late-Pleistocene fan remnants is based on a 14C date and soil developmental features discussed in the next sections. The latest Pleistocene fan surface is characterized by nearly vegetation-free stone pavements, and within the area of study pictured in Figure 2a, those pavements occupy 14% of the landscape.
A mid–late Holocene surface is inset from 83 cm to 100 cm below the latest Pleistocene surface. A latest Holocene surface consists of an abandoned major channel of Growler Wash, inset approximately a meter below the mid–late Holocene surface (Figure 2b and c).
The active channel of Growler Wash is entrenched approximately 4 m below the uppermost surfaces of the latest Pleistocene fan remnants (Figure 3a). Scarp exposures of the latest Pleistocene fan surface reveal coarse gravelly to cobbly alluvium in the uppermost portion. Those coarse materials bury a thick (>1.8 m), fine-textured unit that lacks coarse clasts (Figure 3b). A prominent inset terrace with a surface 1.8–2.0 m above the Growler Wash channel is probably coeval with the latest Holocene surface of the abandoned channel discussed previously (Figure 2b and c and Figure 3a).

(a) View toward the north from the bed of the entrenched channel of Growler Wash. The surface of the terrace is 2 m above the channel bed. The latest Pleistocene surface is 2 m above that terrace. (b) Close-up view of an eroded scarp cut into the latest Pleistocene surface. The upper, coarse gravelly to cobbly unit is the deposit on which the pavement has formed. The underlying fine-grained alluvial unit also appears as a buried surface in the lower portions of soil profiles P-1 and P-2 (Table 1).
Age of the latest Pleistocene alluvium
Soil pit P-1 on the latest Pleistocene surface yielded 3.3 g of wood charcoal at 38 cm depth, 3 cm below the upper boundary of a buried soil surface (Figures 2b and 4a). Charcoal fragments up to 13 mm in diameter were confined to an area a 5–10 cm wide in one of the pit faces. This restricted occurrence and absence of macroscopically observable charcoal in the other pit faces suggest the charcoal was derived from a moderately large woody plant that burned in place, and the incompletely combusted, basal portions of the stem were retained and preserved directly below the existing soil surface before burial by the upper, coarse gravelly alluvium. Anatomical characteristics (structure of medullary rays and vessels) observed microscopically in cross-sectional surfaces of the fragments matched those of reference charcoal samples prepared from two species of Lycium (L. pallidum and L. andersonii), but exact species identity of the buried charcoal could not be ascertained. Radiocarbon dating yielded a calibrated age of 12,745 yr BP (95% probability = 12,720–12,820 yr BP; Beta-410908). The positioning of the buried charcoal reasonably constrains the deposition of the overlying, coarse gravelly to cobbly alluvium to within a millenium preceding the Pleistocene–Holocene boundary.

(a) Pit in foreground is the location of soil profile P-1 on the latest Pleistocene surface (23 February 2003). Soil profile diagram showing position of charcoal sample Growler P-1 copied from original field sketch. (b) Vertical view of stone pavement on latest Pleistocene fan surface near soil profile P-1; scale in centimeters.
Characteristics of pavements and soils
Latest Pleistocene surface
The latest Pleistocene surface is covered with a pavement consisting largely of fine to very coarse gravel (4–64 mm clast diameters); 87% of the surface was covered by clasts 4-mm diameter and larger (Figure 4b). Mean and median surface clast diameters were 17.9 and 15.5 mm, respectively. Coatings of rock varnish on pavement clasts are very light and discontinuous, and smaller clasts frequently lack recognizable varnish coats. Maximum varnish development on the largest clasts has a Munsell color of 5YR 3/1(very dark gray). Lower (subaerial), rubified surfaces of those clasts have a maximum reddening of 5YR 6/4 (light reddish brown).
Both soil profiles described from the latest Pleistocene surface (P-1, P-2; Figure 2b) contained a 2 cm-thick, fine-textured, Vk horizon directly beneath the pavement. This nearly clast-free horizon had a higher silt + clay content than underlying horizons, reflecting the formation of this horizon through the accumulation of aeolian dust. The Vk horizons of both profiles contained many, fine pores (⩾50 per dm2; 0.5–2.0 mm diameter). Underlying Btk horizons exhibited slight structural development and CaCO3 accumulation but negligible increase in reddening (Table 1). The upper sequence of horizons (Vk-Btk1-Btk2-Bk) have formed within the most recently deposited, coarse gravelly to cobbly alluvium. The surface of a buried, weakly developed soil is present at 35 cm in both profiles (Bwkb in P-1; Btkb in P-2). The deepest described horizon in both profiles is within another buried soil formed within very fine-textured alluvium, nearly clast-free alluvium, and contrasts strongly with coarser materials and textures of overlying horizons (Table 1). This same fine-textured unit outcrops in exposures along Growler Wash located 50 and 100 m south of the P-1 and P-2 soil profiles (Figure 3b). The sharp transition from this thick (>1.8 m), fine-textured lower unit to the overlying coarser materials indicates a marked transition the types of sediments carried and deposited by the stream near the end of the Pleistocene, particularly an increased contribution of coarse rhyolitic gravels from hillslopes immediately downstream from Growler Canyon (Figure 1).
Late-Pleistocene fan remnant
This isolated remnant exhibits more advanced soil development than that of the latest Pleistocene surface, indicating somewhat greater age (P-3 soil site, Figure 2b). A stone pavement is present on the central, best-preserved portion of the remnant, and the larger pavement clasts possess continuous coatings of rock varnish that are thicker and darker (5YR 2/1–5YR 3/1; black to very dark gray) than those of the latest Pleistocene surface. Likewise, lower surface of the larger clasts exhibit considerably stronger rubification than those of the latest Pleistocene surface (2.5YR 4/4; reddish brown). A 2-cm-thick, clast-free, fine-textured Vk horizon lies directly beneath the pavement. Underlying Btk horizons exhibit substantially greater clay accumulation (sandy clay loam textures) and stronger reddening (5YR 5/4; reddish brown) than those of the latest Pleistocene surface (Table 1).
Mid–late Holocene surface
The fine gravelly alluvium of the mid–late Holocene surface exhibits only slight pedogenic development, in the form of weak structural development in the Bwk horizon and carbonate accumulation limited to thin, discontinuous coatings on larger clasts within Bwk and Bk horizons (H-1 soil site, Figure 2b; Table 1). There is no V horizon, and A, Bwk, and Bk horizons all have loamy sand to sandy loam textures and negligible textural discontinuities between horizons.
Vegetation, vegetation traces, and soil conditions influencing plant function
Holocene surfaces
The two Holocene surfaces support shrub-dominated vegetation, but species compositions and sizes of plants differ on them. L. tridentata and Ambrosia dumosa contribute roughly equivalent amounts of canopy cover on the youngest depositional surface, the abandoned major channel of Growler Wash (Figure 5a). Other species including Prosopis velutina, Parkinsonia [= Cercidium] microphyllum, L. andersonii, and Ambrosia [= Hymenoclea] salsola are also present but contribute only a small fraction of total canopy cover. L. tridentata is a potentially long-lived shrub, reaching ages of thousands of years, during which outward growth of individual plants generates ring-like clones that can reach many meters in diameter (McAuliffe et al., 2007; Vasek, 1980). The small basal diameters of the L. tridentata on this surface (<50 cm) indicate the youth of this surface. In contrast to the diverse mix of species on the youngest surface, the mid–late Holocene surface (site of H-1 soil profile) is overwhelmingly dominated by large L. tridentata with basal diameters up to 1 m and canopy diameters from 2 to 2.5 m, indicating greater age of those plants (Figure 5b). Few, scattered individuals of A. dumosa occur on the surface.

(a) Vegetation on the latest Holocene surface, the abandoned major channel of Growler Wash. The smaller shrubs with light-colored foliage in the foreground are Ambrosia dumosa. The single, tall canopy on the left is an isolated Prosopis velutina (velvet mesquite). The other medium-height shrubs throughout the middle and background are Larrea tridentata. Desiccated ephemeral plants from the previous winter–spring season, primarily Plantago ovata, cover the soil surface (9 May 2015). (b) Vegetation on the late–middle Holocene surface dominated by larger L. tridentata. The few smaller plants in the center and far left are A. dumosa (9 May 2015).
Latest Pleistocene surface
Light-colored plant scars ranging from 1.5 to 3.0 m diameter dot the darker pavements (Figure 6a). Those features mark places formerly inhabited by larger perennial plants, which served as foci for significant bioturbation (McAuliffe and McDonald, 2006). Death and disappearance of those plants yielded circular, light-colored patches where the stone pavement and underlying soil horizons, including the Vk horizon, were originally prevented from forming (Figure 6b and c). The diameters of the plant scars suggest that those places originally were occupied by the larger L. tridentata rather than the much smaller A. dumosa. Scattered individual plants of L. tridentata continue to occupy the areas of pavement, but mortality and loss of those plants continues. In March 2004, McAuliffe and McDonald (2006) documented mortality of remaining L. tridentata on the pavement surface surrounding the P-1 soil pit after 5 years of below-average precipitation. A significantly larger fraction (43% of 21 plants) centrally located within the pavement-covered area had died, whereas only 9% of 21 plants located peripherally beyond the edge of the pavement succumbed to the drought.

(a) Aerial view of pavements on the latest Pleistocene surface south of Growler Wash showing clonal rings of Larrea tridentata (left side) and light-colored plant scars distributed about the surface of the darker pavement surface. ‘B’ and ‘C’ indicated on the right side are the same plant scars shown in ground view in (b) and (c). The white arrows indicate the direction of camera views in the ground photos. The curved lines across the pavement surface are vehicle tracks. Coordinates of the center of this view are 32.15056°N, 113.01888°W (Google Earth™ image). (b) and (c) Ground views of light-colored plant scars, both approximately 2 m in diameter. Lack of a pavement and associated V horizon on these features enables ephemeral plants like those shown here to grow, given ample frontal precipitation during the winter–spring. The single woody stem in (b) is a dead, very young L. tridentata that probably established in the plant scar during the period of above-average precipitation that pre-dated 2000. (d) Aerial view showing different fan surfaces of the alluvial piedmont on the east side of the Soda Mountains, California, Mojave Desert. The dark, well-formed pavement designated 1 is the oldest surface, 2 and 3 are moderately to weakly developed pavements on latest Pleistocene to early Holocene surfaces, and 4 indicates Holocene alluvial deposits. The central square is 100 m on a side and is enlarged in (e). Coordinates of the center of that square are 35.20586°N, 116.21209°W. Parallel dark lines in the lower right are highway I-15 (Google Earth™ imagery). (e) Enlarged view of latest Pleistocene to early Holocene fan surfaces (2 and 3) showing light-colored plant scars distributed over the darker pavement surface. The small white ‘X’ left of center is a marker placed in association with an aerial photography survey (Google Earth™ imagery).
A few of the remaining living plants of L. tridentata on the pavement or along pavement margins are large, ring-like clones or arc-shaped remnants of such clones (Figure 6a). Few clones occur centrally within areas covered by pavements but are more common near pavement edges, where runoff derived from interior portions of the pavements likely contributed to their long-term persistence. Within the study area, the maximum diameters of the largest clones ranged from 4.2 to 7.0 m (mean = 5.6 m, N = 10). Those clones are considerably larger (and much older) than the largest L. tridentata found on the younger surfaces, indicating a duration of occupancy by the individual clones that likely greatly exceeds the age of the mid–late Holocene surfaces.
Soil hydrological behavior and salt accumulation
The fine-textured Vk horizon, although only 2 cm thick, significantly inhibits surface infiltration. This is demonstrated by the difference in wetting fronts in soils of pavements versus those of the mid–late Holocene surface following the 52 mm precipitation event of 12–13 February 2013. A week following that event, average wetting front in soils beneath pavements was approximately only a third of that in soils of the mid–late Holocene surfaces (Figure 7). The prevention of soil moisture recharge to significant depth in soils beneath the pavements negatively affects plants in two ways: (a) directly due to the lack of plant-available soil moisture and (b) indirectly due to the accumulation of extremely high concentrations of soluble salts at shallow depth due to the chronic lack of deep leaching of these materials (Figure 7).
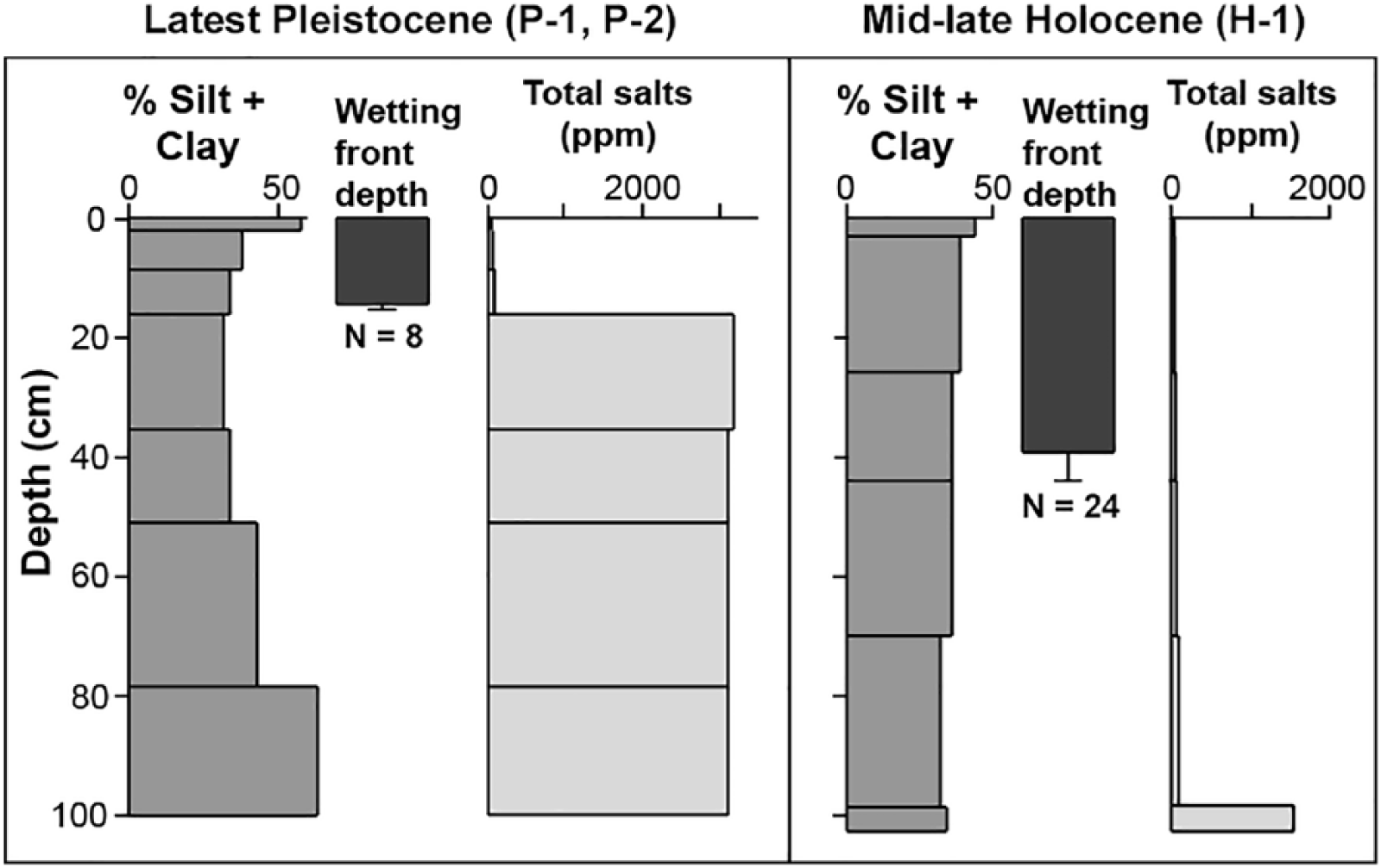
Depth distributions of silt + clay and total salt content of soil horizons sampled on the latest Pleistocene and mid–late Holocene surfaces. The dark solid vertical bar in the center of each panel represents mean wetting front depth; the vertical whisker is the 95% confidence limit. Sample sizes (N) listed below the whisker plots are the number of separate excavations from which measurements were taken.
Discussion
Pavements, V horizons, and soil hydrology
On gravelly or other coarse, rocky parent materials, stone pavements form as aeolian sediments added to the surface are translocated downward beneath surface clasts. As these fine materials accumulate, the developing V horizon lifts and maintains a layer of clasts on the surface, forming the pavement (Anderson et al., 2002; McFadden, 2013; McFadden et al., 1987). The low permeability of this fine-textured horizon greatly impedes infiltration. Multiple studies have shown that infiltration rates and hydraulic conductivity in soils of Pleistocene fan deposits with V horizons in both the Sonoran and Mojave deserts are approximately an order of magnitude lower than those on soils without that horizon (McAuliffe et al., 2007; McDonald et al., 1996; Meadows et al., 2008; Musick, 1975; Shafer et al., 2007; Turk and Graham, 2011; Young et al., 2004). The marked contrast in wetting front depths after a major precipitation event (Figure 7) is a consequence of these different hydrologic responses. Musick (1975) measured similar contrasts in wetting fronts following experimental infiltration. Due to the chronic lack of deep infiltration, high concentrations of salts accumulate at shallow depth beneath pavements (Anderson et al., 2002; Graham et al., 2008; McAuliffe, 1999; McAuliffe et al., 2018; Musick, 1975; Wood et al., 2005). In these upland desert environments, atmospheric deposition rather than groundwater is the source of those salts.
Reduction of infiltration capacity by the V horizon exerts a strong control on plant function and performance. Hamerlynck et al. (2000, 2002) showed that L. tridentata experienced greater water stress in soils of a late-Pleistocene fan surface with a well-developed V horizon than on other surfaces. During prolonged drought, net photosynthesis of those plants dropped below compensation levels, compared with surfaces capable of more readily absorbing precipitation and storing it at depth within the soil. The increasingly hostile soil environment on a developing pavement can lead to plant mortality and a progressive elimination of vegetation. Plant scars on pavement surfaces mark the former locations of larger perennial plants and represent a landscape-scale trace fossil record of the vanished vegetation (McAuliffe and McDonald, 2006). The 14C date obtained from buried wood charcoal constrains the time of emplacement of the latest Pleistocene fan deposit of the Growler Wash site to just before the Pleistocene–Holocene boundary. Consequently, the pavement and associated soil horizons on that surface have developed entirely during the Holocene as has the progressive loss of vegetation.
Tempo and modes of vegetation change during the Holocene
At the elevation of the study area (370 m), L. tridentata and A. dumosa had arrived in the region by the time of the Pleistocene–Holocene transition (11,650 yr BP). Ancient woodrat middens collected at 330–580 m elevation from the Tinajas Atlas Mountains, Arizona, located approximately 100 km west of the study area, contained both L. tridentata and A. dumosa at 18,931 yr BP (originally reported as 15,680 radiocarbon years, Van Devender, 1990). In the Whipple Mountains, California, 260 km north-northwest of the study area, L. tridentata was present nearly continuously in middens collected from 490 to 525 m elevation from 12,329 yr BP (= 10,490 radiocarbon years) to the present (Van Devender, 1990). Both L. tridentata and A. dumosa are more common occupants of alluvial fan environments in basins than of rocky mountainous terrains; consequently, if midden records from rocky uplands indicate these two species were present at the time of the Pleistocene–Holocene transition, they would undoubtedly have also occurred in alluvial basins of comparable elevations like Growler Valley.
Originally, the pedogenically unaltered, gravelly alluvium of the latest Pleistocene fan deposit would have facilitated infiltration and substantial, deep soil moisture storage, thereby readily supporting vegetation. In both the Sonoran and Mojave deserts, newly created alluvial deposits such as this are occupied by a succession of different species. Among the first are short-lived shrubs that rapidly colonize disturbed areas. Most common within this group is A. salsola, which typically predominates on recently abandoned ephemeral stream terraces throughout the region (McAuliffe, 1994; Webb et al., 1988); other species include Encelia spp. and wind-dispersed Stephanomeria spp. The same set of pioneer species rapidly colonizes anthropogenic disturbances where vegetation has been cleared (e.g. excavated transmission line and pipeline corridors, abandoned townsites, etc.; Abella, 2010; Abella et al., 2007; Berry et al., 2016; Bolling and Walker, 2000; Prose et al., 1987; Vasek, 1983; Webb and Wilshire, 1980; Wells, 1961). The short-lived pioneers decline in abundance as longer-lived species including A. dumosa and L. tridentata colonize and eventually predominate. The vegetation of the youngest alluvial surface at the site, the abandoned channel (Figure 5a) represents this subsequent stage of plant succession with A. dumosa and L. tridentata present as co-dominants, but very few A. salsola. Based on the documented relatively slow rates of establishment of L. tridentata in disturbed areas (e.g. Berry et al., 2016; McAuliffe, 1994), the transition from a recently disturbed surface dominated by pioneer species such as A. salsola to one co-dominated by A. dumosa and L. tridentata probably requires nearly a century to perhaps a few hundred years. With time, L. tridentata becomes increasingly dominant and A. dumosa and other shrubs decline in abundance (McAuliffe, 1988), frequently culminating in a near-monoculture of large, widely spaced plants of L. tridentata (e.g. vegetation of the mid–late Holocene surface; Figure 5b). Comparisons of Holocene soil chronosequences in other Sonoran Desert locales demonstrate this phenomenon: a mixture of small, young L. tridentata and other species of shrubs on late-Holocene surfaces, but an overwhelming dominance of large L. tridentata, to the near exclusion of other shrub species on older, middle-Holocene alluvial surfaces (McAuliffe, 1991, 1994). A few thousand years may be required for this exclusion to unfold (McAuliffe, 1994), apparently through chemically mediated root interactions (Mahall and Callaway, 1991) and the ability of L. tridentata to extract soil water held at much higher capillary tensions (i.e. lower water potential) than can other species (Hamerlynck et al., 2002; McAuliffe et al., 2007; Oechel et al., 1972). These below-ground competitive interactions are probably also responsible for the wide spacing between individual, large L. tridentata (e.g. Figure 5b) and the inhibition of recruitment of young plants in those intervening spaces (Fonteyn and Mahall, 1981; Mahall et al., 2018).
The occurrence of large, ring-like clones of L. tridentata on the latest Pleistocene surface is evidence that those plants had originally established well before the pavement and associated V horizon had fully developed. These clones, up to 7 m in diameter, probably represent plants that initially established at least several thousand years ago. Based on radiocarbon dates of wood fragments collected from within ring-like clones of L. tridentata, Vasek (1980) calculated an outward radial growth rate of 0.66 mm/yr. Given comparable growth, the largest clones at the Growler Wash site would exceed 5000 years of age. However, that rate probably significantly overestimates actual growth rates of ring-like clones at the Growler Wash site because of the harsh soil conditions that eventually developed in association with the pavement (Figure 7), compared with the much more favorable soil hydrological conditions documented by McAuliffe, et al. (2007) at the site studied by Vasek (1980). Some of the largest clones that have persisted on the latest Pleistocene surface at the Growler Wash site may represent plants that originally established during the early Holocene.
Process of plant exclusion from developing pavements
Pavements largely devoid of perennial plants are absent in more mesic portions of the Sonoran and Mojave deserts where greater amounts of biological activity associated with denser vegetation prevents the surface stability required for pavement formation (McAuliffe, 1999; Quade, 2001). However, a climatic threshold likely exists beneath which vegetation and associated bioturbation become sufficiently sparse and spatially restricted that the process of pavement formation together with development of the underlying V horizon can proceed. The widespread occurrence of the light-colored plant scars on the pavements of the latest Pleistocene surface indicates that widely spaced, relatively large perennial plants – most likely L. tridentata – once occupied those locations while pavements and underlying V horizons were forming in wide, unoccupied gaps between the shrubs. In these desert environments, zones of bioturbation by burrowing rodents are concentrated directly beneath shrub canopies and rapidly diminish in occurrence with distance from canopy edges (Chew and Whitford, 1992; McAuliffe and McDonald, 2006; Quade, 2001; Shafer et al., 2007). In contrast to that spatially restricted distribution of surface bioturbation, L. tridentata has relatively deep and laterally extensive root systems that extend far beyond edges of canopies (McAuliffe and McDonald, 1995; Schwinning and Hooten, 2009; Stevenson, et al., 2009) and can completely occupy the soil in the wide gaps between plants (Brisson and Reynolds, 1994). Although soil directly beneath a canopy may continue to experience bioturbation that inhibits horizon formation, pavements and associated V horizons can begin to develop away from canopies in areas below some threshold of disruptive biological activity on the surface. Development of the V horizon in the wide spaces between shrubs impedes deeper infiltration in those areas, reducing the supply of soil moisture that the laterally extensive root systems can obtain (Shafer et al., 2007). In addition to this direct effect of reduced soil moisture availability on plants, the progressive, substantial accumulation of soluble salts due to the shallower leaching depth eventually generates a soil environment physiologically unsuitable for occupation by roots (Figure 7). This likely contributes to a positive feedback loop – once inhospitable soil conditions begin to develop in gaps between widely spaced plants, biological activity, and bioturbation becomes further suppressed in those gaps. Consequently, the shrubs become increasingly isolated on shrinking islands of marginally suitable soil conditions, surrounded by soils that are incapable of providing soil moisture resources and eventually become uninhabitable due to shallow accumulation of salts. These stresses lead to progressive stem die-back or death of the entire plant, particularly during extended drought intervals, as previously documented at this site (McAuliffe and McDonald, 2006), ultimately generating barren pavement surfaces dotted with light-colored plant scars. Bioturbation and limited plant growth persists on the scars for some time, but eventually pavements and associated V horizons re-form in those areas as described for surfaces of even older alluvial fan surfaces (McAuliffe and McDonald, 2006).
Summarizing the causes and timing of vegetation change detailed above, during the first half of the Holocene, the latest Pleistocene surface would have been occupied by a sequence of successional stages. A suite of pioneer species would have initially colonized the newly created surface, followed by their replacement by a mix of longer-lived species, including A. dumosa and L. tridentata, and culminating with the eventual predominance of the latter. Those kinds of compositional changes can be attributed mainly to differences in life history attributes of various species (i.e. dispersal and colonization vs competitive abilities). However, during the latter part of the Holocene and continuing to the present, continued development of soil conditions associated with the pavement, particularly the V horizon, has increasingly diminished the capacity of the surface to absorb precipitation and is responsible for the latest phase of vegetation change involving the progressive loss of plants from the surface. Rather than being attributed to directly to the influence of climate change on plants, this near exclusion of perennial vegetation from the pavement surfaces is due to the way climate has influenced soil profile development over the course of the Holocene.
Regional occurrence and overall contribution to late-Quaternary vegetation change
Alluvial fan deposits similar in age to the late-Pleistocene fan deposits described in this paper occur throughout the Sonoran and Mojave deserts. Miller et al. (2010) compiled records of numerically dated alluvial fan deposits in the Mojave Desert and identified a pronounced peak from 9 to 14 ka, indicating region-wide occurrence of alluvial fan aggradation during this interval. Bull (1991) also suggested a similar peak in fan aggradation from 8 to 12 ka in the Sonoran Desert of Arizona. Throughout the lower, more arid elevations of both regions, pavements that have developed on these latest Pleistocene to early Holocene surfaces frequently possess light-colored plant scars like those described in this paper (Figure 6d and e) indicating a similar, progressive loss of vegetation during the Holocene. The existence of somewhat darker, varnished pavements surrounding the plant scars indicates that the plants were present for at least some portion of the time when the process of pavement formation and varnish accumulation was underway. Increased aridity of the Holocene has fostered the development of pavements and associated soil horizons in two ways: (a) an overall reduction of vegetation cover to a level where little bioturbation occurs between widely spaced plant canopies and (b) an increased flux and input of aeolian dust required for formation of the V horizon (Wells et al., 1987). However, the increased aridity of the Holocene is not the factor directly responsible for the progressive elimination of vegetation from these surfaces. Rather, the development of pavements and associated V horizons greatly diminishes infiltration capacity, which prevents the storage of sufficient amounts of soil moisture required to support vegetation, and also leads to substantial accumulations of salts at shallow depth that render the soil environment uninhabitable by most plants. In addition to the substantial changes in species composition and types of vegetation attributed directly to the effects of changing climate following the Pleistocene (e.g. Betancourt, et al., 1990), the progressive development of certain kinds of soil conditions like these during the Holocene is another major, direct contributor to overall vegetation changes during the late Quaternary.
Footnotes
Acknowledgements
National Park Service (NPS) personnel at Organ Pipe Cactus National Monument facilitated fieldwork at the site. NPS Ecologist Susan Rutman invited me to visit the site in 2003 and Tim Tibbits helped secure official permission (NPS Permit #ORPI-2003-SCI-018) to excavate soil pits. Charles Connor and Peter Holm of the NPS as well as Greg McCurdy of the Desert Research Institute provided precipitation records from the Growler rain gauge. Other individuals also contributed to the work; Cherie Wilson assisted in collecting wetting front measurements in 2003, and Veronica Nixon secured satellite GIS imagery for use in identifying geomorphic surfaces and determining the area of the Growler Wash drainage basin. Beth Brand, librarian of the Desert Botanical Garden’s Shilling Library, assisted in obtaining interlibrary loan copies of various publications. Helpful comments on the original manuscript were provided by Les McFadden and two anonymous reviewers.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.