
Abstract
During the Holocene, multiple thermal changes commonly occurred in the northern hemisphere. They are well-recorded in lakes with minimum human impact from the Arctic Circle area. The development of these lakes reflects ecological and climatic changes occurring from the formation of the lakes until present-day times. All environmental fluctuations affect biodiversity and are reflected in the number and composition of species. The goals of this study were to detect the ecological changes in a small Finnish lake using pollen, Cladocera and geochemical analyses. The research area is located within the northern zone of boreal coniferous forest and is the most sparsely populated region of Finland. The lake is located in Kuusamo uplands, E Finland, near the polar circle and over 20 km from the Russian border. Indicators of cold water were found only during the initial stage, after the 8.2 ka event and then the temperature was higher. Trophy was high at the beginning of the lake development and then a significant increase in trophy was found after 2600 BP. The impact of human activity is hardly traceable in Arctic Circle Finland throughout the Holocene Thermal Maximum. During the late-Holocene (after 4200 yr cal. BP), this impact is still weak and, even as late as the 20th century, only a few traces of human activity are recorded. General conclusion is that long-term climatic shift has been the most important factor driving changes in the limnology of Lake Talvilampi.
Introduction
During the Holocene, multiple thermal changes commonly occurred in the northern hemisphere (Janbu et al., 2011). In particular, these are well-recorded in lakes with minimum human impact from the Arctic Circle area (Anderson et al., 1999; Brodersen and Anderson, 2000; Eddudóttir et al., 2015; Smol et al., 2005). The development of these lakes reflects ecological and climatic changes occurring from the formation of the lakes until present-day times (Sarmaja-Korjonen et al., 2006; Szeroczyńska et al., 2007). The accumulated sediments commonly consist of organic matter (OM) and mineral deposits in different proportions, which, using palaeoecological and physicochemical methods, enables environmental conclusions to be drawn (e.g. Berglund et al., 1986; Birks and Birks, 2006; Tobolski, 2000).
All environmental fluctuations affect biodiversity and are reflected in the number and composition of species inhabiting water and terrestrial habitats (Ellenberg et al., 1991; Kjellman et al., 2018; Szeroczyńska and Zawisza, 2011b). These changes are visible in plant communities (Hicks, 1992; Iversen, 1964a; Tobolski, 2010) as well in the limnic environment, and, in the vicinity of lakes, influence zooplankton (Korhola and Rautio, 2001; Smol et al., 2005). Cladocera, one of the components of this plankton which is preserved as shells in lake sediments, is a group with good bioindicative value. Plants and Cladocera allow for the reconstruction of not only the development of lakes but also changes of regional nature (Kamenik et al., 2007; Sarmaja-Korjonen and Seppä, 2007; Smol and Douglas, 2007; Sorvari et al., 2000).
In lake sediments, OM is composed of detritus from higher terrestrial plants occupying the drainage basin, aquatic higher plants growing in and around lakes and algae in the lake (Tobolski, 2000). Lake morphology, catchment topography and the relative abundance of lake and catchment plants strongly influence the relative contributions from terrestrial and aquatic sources to lake sediments (Meyers and Ishiwatari, 1993). The δ15N and δ13C ratios of bulk OM in lake sediments can be used to infer changes in OM (aquatic or terrestrial sources). Changes in δ13C and δ15N can also provide information on nutrient cycling, lake mixing regimes and water column stability (e.g. Meyers and Ishiwatari, 1993; Meyers and Lallier-Vergès, 1999). Pollen studies enable the reconstruction of vegetation by regional plant communities and macrophytic species of limnic and telmatic origin. The bioindicative value of stenotopic species is used as well as general knowledge of the climatic zonation and interglacial succession of plant communities (Berglund, 1986; Smol et al., 2001; Tobolski, 1976). Cladocera data are frequently used as indicators of past environmental conditions as well as climatic change (Szeroczyńska, 1985).
In the cool, northern zone of Europe (boreal zone), many oligotrophic, soft-water lakes still exist and these are a common type of lakes. Their location is related to the geographical range of low-demanding plants (low content of nutrients and low pH of the water) (Heegaard et al., 2001). During the 20th century, the number of oligotrophic lakes in temperate zones in Europe decreased significantly because of agricultural activity and common eutrophication of soil and water (Murphy, 2002; Roelofs, 1983; Szmeja, 2001).
The goals of this study were to detect the ecological changes in a small Finnish lake – Lake Talvilampi – during the last several thousand years and also try to identify some traces of human activity in this region of the Arctic Circle. The results were achieved using pollen, Cladocera and geochemical analyses focused on: (1) changes in the content and composition of OM in the sediments of Lake Talvilampi during the studied period, (2) reconstruction of vegetation in the last 8000 years in relation to climate changes, (3) changes in the quantity and diversity of the Cladocera species and (4) estimation of the lake conditions, such as water temperature and level, and a trophy state.
Study site
The bedrock of Finland belongs to the Precambrian East European craton of northern and eastern Europe and northwestern Russia, and it was eroded during the Pleistocene continental glaciation (Vaasjoki et al., 2005). The central area of Finland was deglaciated at the turn of the Late Glacial and the Holocene (Lundqvist, 1986). According to Stroeven et al. (2016), the glacial retreat in the research area took place 10,200 yr BP.
The research area is located within the northern zone of boreal coniferous forest and is the most sparsely populated region of Finland. Talvilampi is a lake located in Oulanka National Park (Kuusamo uplands, E Finland), near the polar circle and over 20 km from the Russian border 66°22′42.28″N, 29°17′5.41″E (Figure 1). The climate is cold and snowy with permanent snow cover between November and May, reaching about 70 cm at its peak in March. Mean annual temperature is about 0°C, ranging between − 12.4°C in January and 14.7°C in July (Wyszkowski, 1987). Mean annual precipitation exceeds 500 mm.
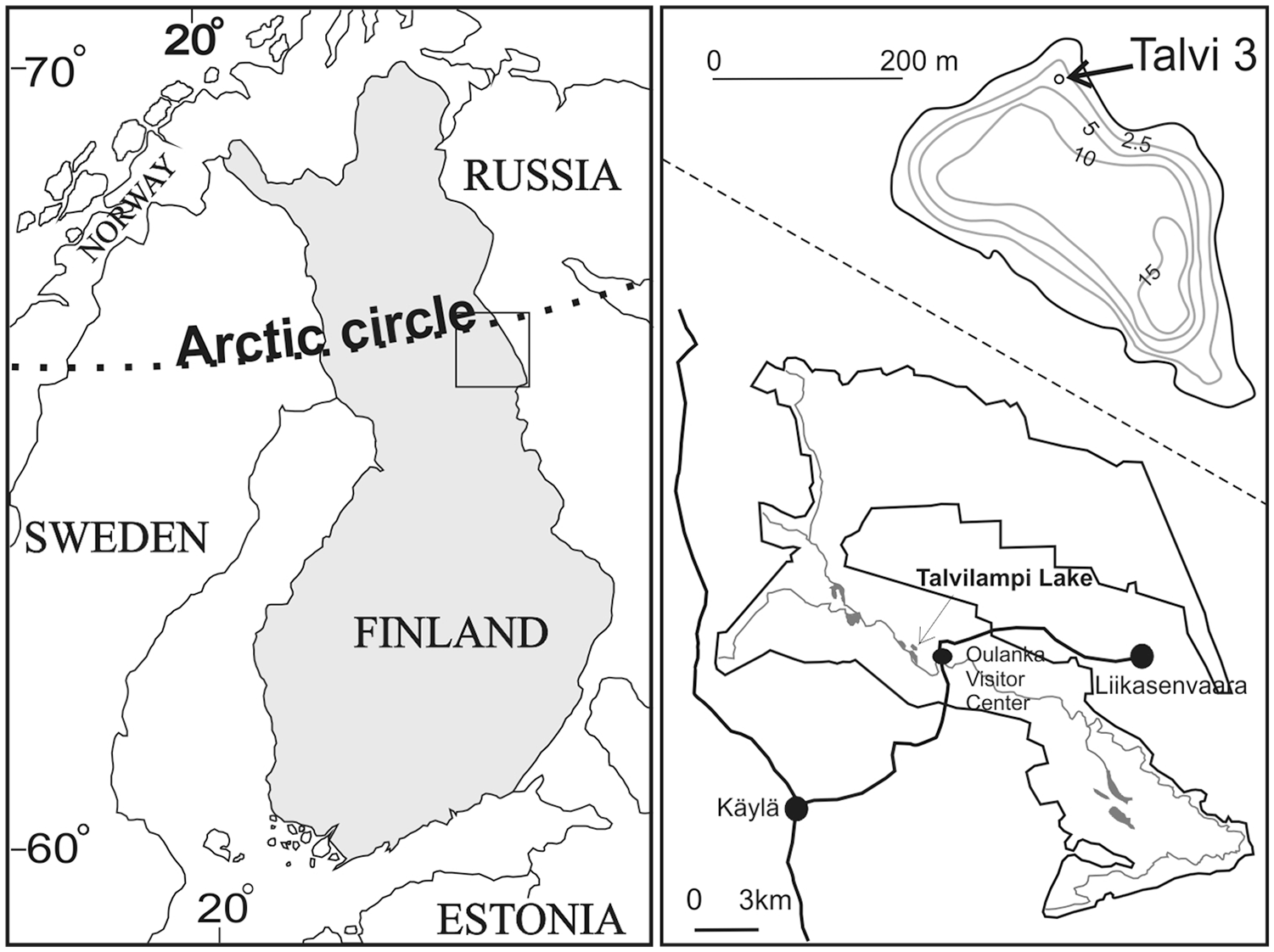
Location of the research area and Talvilampi Lake in Finland.
The lake is located on the Oulanka River terrace and is directly connected with the river by a narrow canal. Its water-level regime is thus strongly influenced by river flow. The maximum depth reaches 15.9 m. The slope of the lake basin in the north-eastern part of the lake, from where the core was taken, reflects the slope of the lake shore area, that is, the slopes of the Oulanka River Valley.
The region is covered by boreal forest with Pinus sylvestris as a dominant plant. Talvilampi is an oligotrophic lake with Isoëtes lacustris partly covering the bottom. The surrounding of the lake is covered with P. sylvestris, Betula pubescens, Sorbus aucuparia, Picea abies and Juniperus communis. In the bottom layer, Ledum palustre, Vaccinium vitis-idea, V. uliginosum and V. myrtillus were found. Trientalis europea, Linnea borealis, Galium boreale, Viola epipsila, Ranunculus acris and Geranium sylvaticum belong to the herbal vegetation. There is also a consistent layer of moss species.
Methods
Field work
Sediment cores were drilled in 2008 as a part of the LAPBIAT (Lapland Atmosphere-Biosphere Facility) project concerning Lobelia lakes in Finland called ‘Loblakes’. A thick (about a half metre) ice cover on the water surface enabled safe drilling. Three cores were taken in transect from littoral to the deeper part of the lake. Only one core was analysed and described in this article. The Talvilampi 3 core (Tal3), 153 cm long, was drilled from the depth of 4.30 m with a piston corer with an internal tube diameter of 45 mm. It was located 36.5 m from the north-eastern shore of the lake. The excavation in this part of the lake followed suggestions of local inhabitants who had observed the existence of an Isoëtes population there.
Chronology
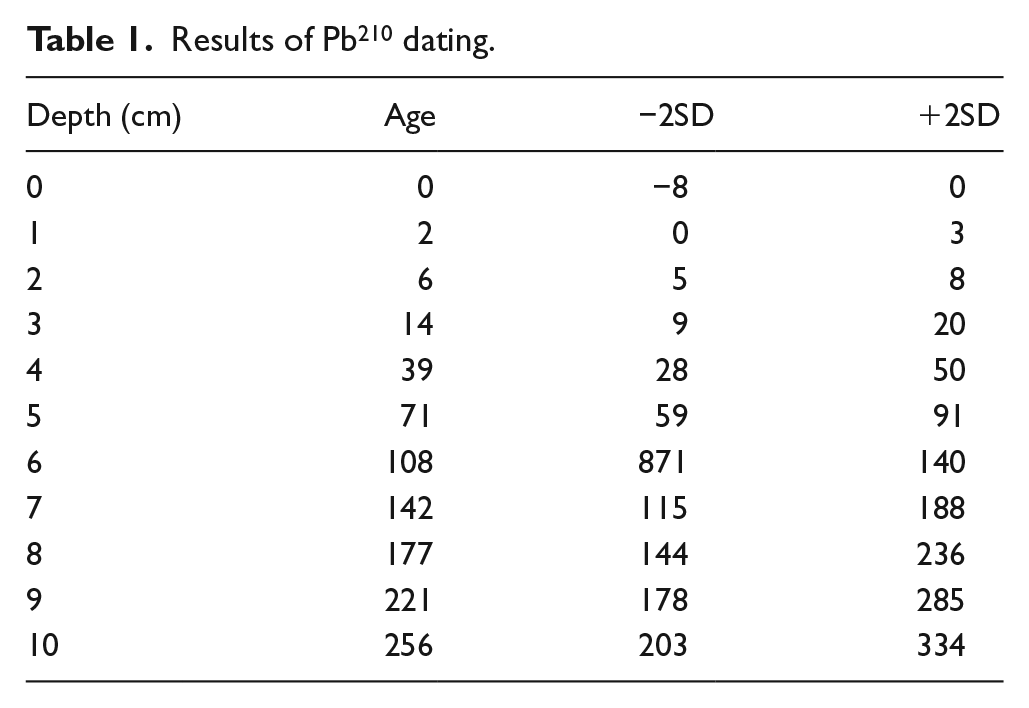
Ten Pb210 dates were obtained for the uppermost 10 cm of sediments in the Isotope Dating and Environment Research Laboratory at the Institute of Geological Sciences, Polish Academy of Sciences. Four accelerator mass spectrometry (AMS) dates were determined in the Poznań Radiocarbon Laboratory and the next three at Beta Analytic – Radiocarbon Dating, Miami, Florida, US. Terrestrial macrofossils (in Poznań) and pollen concentrate (in Florida) were the material for dating. Macrofossils for the AMS 14C dates were selected from the studied sediments but only two were obtained (Table 2) because of the extremely small amount of selected fossils. Pinus sylvestris bud scales and/or bark, and needles were selected from the depths of 17 and 90 cm; however, no more than 0.02 and 0.07 mg C, respectively, was obtained so the remains were too small for dating. Two dates from the depth of 48 and 145 cm were obtained and used in the age-depth model (Figure 2). Two samples of macrofossils (from the depths 17 and 90 cm), AMS dated in Poznań, were omitted because of insufficient content of separated carbon: 0.02–0.07 mg C. A further three were obtained at Beta Analytic – Radiocarbon Dating, Miami, Florida, US based on pollen concentrate from bulk gyttja. The date from the depth 32/33 cm Beta 38388 was too old and not in chronological order and it was omitted. According to the dates, all the diagrams are plotted with the yr cal. BP and depth scale in Tilia and TGV Programmes.
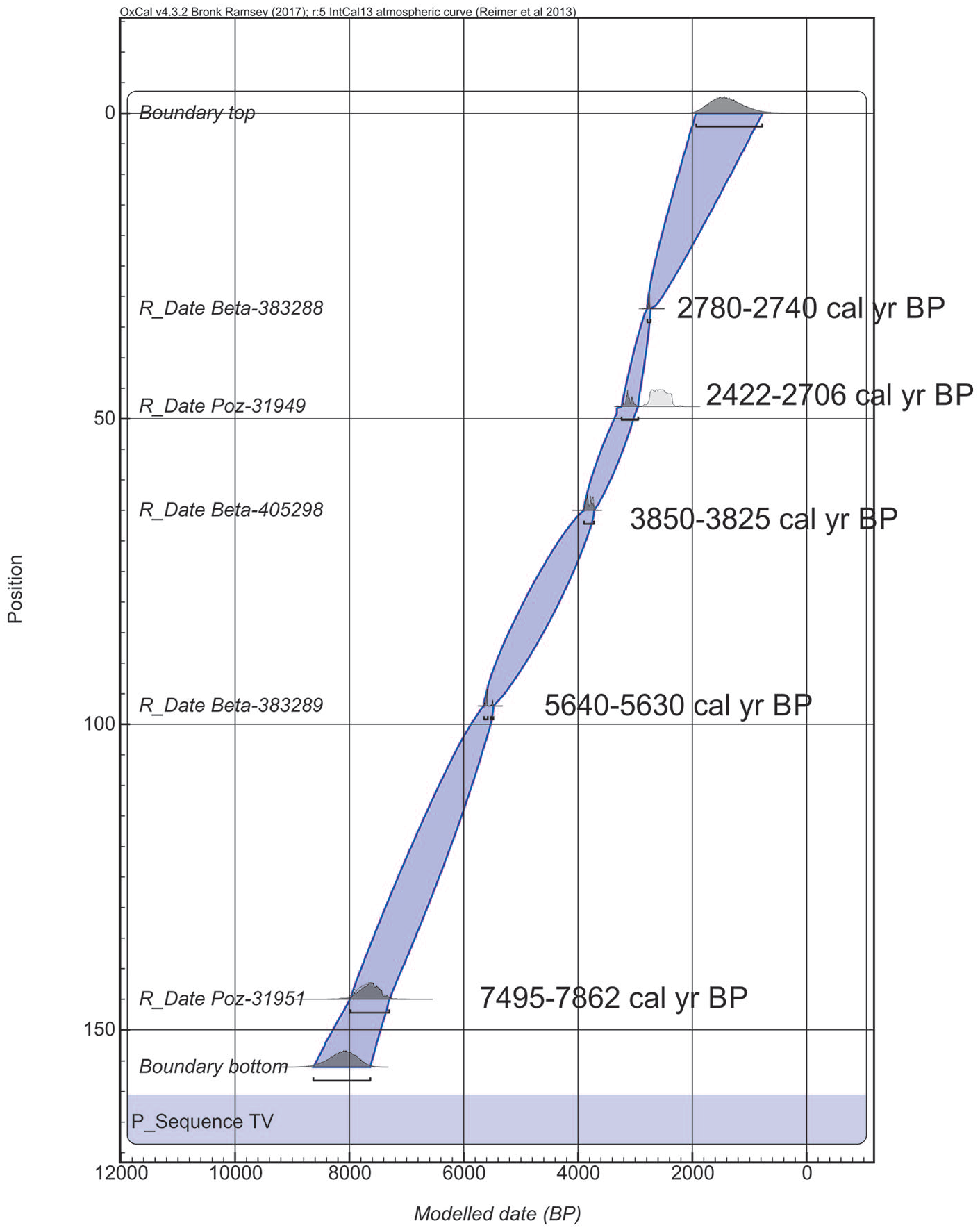
Depth-age model based on 14C dates, OxCal 4.3.2.
Isotopic and geochemical analyses
For stable isotope (carbon and nitrogen) and selected geochemical analyses, OM samples were collected every 1–2 cm from the depth of 3–150 cm (90 samples). The sediments were dried at 60°C and ground. The carbonate fraction was removed with hydrochloric acid. The carbon and nitrogen isotope compositions were analysed using a Flash Elemental Analyser 1112 and Thermo MAT 253 mass spectrometer, which were calibrated based on an internal nicotinamide standard and reported as per mill (‰) deviations versus atmospheric N2 (δ15N) and Vienna Pee Dee Belemnite (δ13C). The analytical errors (l SD) for the δ13C and δ15N measurements were 0.17‰ and 0.24‰, respectively.
The carbon and nitrogen stable isotopes were used to define the changes in the quantity and source of OM transported into the paleolake. Lake morphology, catchment topography and the relative abundances of lake and catchment plants significantly influence the relative contributions to lake sediments from terrestrial and aquatic sources (Meyers and Ishiwatari, 1993).
The organic carbon and nitrogen concentrations were analysed using a Vario Micro Cube Elemental Analyser. The analyses were performed in the Laboratory for Isotope Dating and Environmental Studies at the Institute of Geological Sciences of the Polish Academy of Sciences in Warsaw, Poland. The samples were collected from the depth of 3–150 cm (37 samples; Figure 3).

Geochemical-isotopic results (Lake Talvilampi, Arctic Circle, Finland, core Talvilampi 3).
The combination of selected geochemical (total organic carbon – TOC, total nitrogen – TN, TOC/TN) and isotopic data (δ15N, δ13C) obtained from OM with botanical proxies is used for paleoenvironmental reconstructions (e.g. Janbu et al., 2011; Kołaczek et al., 2015; Lauterbach et al., 2011; Niska and Mirosław-Grabowska, 2015).
Pollen analysis
For pollen analysis, samples of 1 cm3 in volume were taken every 1 cm. These were prepared using laboratory technics according to a standard procedure (Berglund and Ralska-Jasiewiczowa, 1986). Acetolysis was maintained for 3 min, and then samples were washed in acetic acid and covered with glycerine. Two tablets of Lycopodium spores were added. Just before preparation of each slide for microscope analysis, samples were stained with safranine. Every sample was counted to at least 500 pollen grains of trees and shrubs; however, the bottommost samples did not exceed more than 300 pollen grains because of the low frequency. Percentages were counted according to the equation AP + NAP = 100% (AP is the sum of trees and shrubs, and NAP is the sum of terrestrial herbs). Percentage pollen diagrams were prepared in Tilia and Tiliagraph software with chronology in yr cal. BP as a primary scale (Figures 4 and 5).

Percentage pollen diagram of selected terrestrial plants (in black) (Lake Talvilampi, Arctic Circle, Finland, core Talvilampi 3). Exaggeration ×10 are given for improved reading of rare taxa.

Percentage pollen diagram of selected plants of local environment (Lake Talvilampi, Arctic Circle, Finland, core Talvilampi 3). Exaggeration ×10 are given for improved reading of rare taxa.
Macroremain analysis
Cores for macrofossil analysis were sampled irregularly to assess the richness of findings and to take fossils for radiocarbon dating. Samples (10–15 cm3 each) were prepared using standard methods for macrofossil analysis (Birks, 2007; Tobolski, 2000). The material was sieved on 0.125 mm mesh sieves. The residue was sorted under a stereoscopic microscope at 10–200× magnifications. Selected terrestrial plant macrofossils were used for dating.
Cladocera analysis
Subfossil Cladocera analysis was conducted on 101 sediment subsamples (1 cm3) that were processed according standard procedures (Frey, 1986). Each sample was heated and stirred in a 10% solution of KOH for 20 min in order to remove humic matter and next washed through a 38 μm mesh. The final residue was dissolved in 10 mL distilled water then 0.1 mL of the final solution was mounted on microscope slides and two to four slides (minimum of 200 remains) were counted from each sample. The extracted remains were identified using an OLYMPUS BX40 transmitted-light microscope at magnifications of 100×, 200× and 400×. All cladoceran remains were counted (head shields, shells, ephippia, postabdomens). The identification of Cladocera remains was based on Szeroczyńska and Sarmaja-Korjonen (2007) and Flössner (2000). The results were plotted as a relative abundance diagram (Figure 5), using C2 software (Juggins, 2007). Cladocera Assemblages Zones (CAZ) were distinguished based on significant changes in the relative abundance of Cladocera, total Cladocera sum and the composition of species. The total number of Cladocera indicated in the diagram (Figure 6) were standardised to concentration (Cladocera individuals per gram of dry sediment).
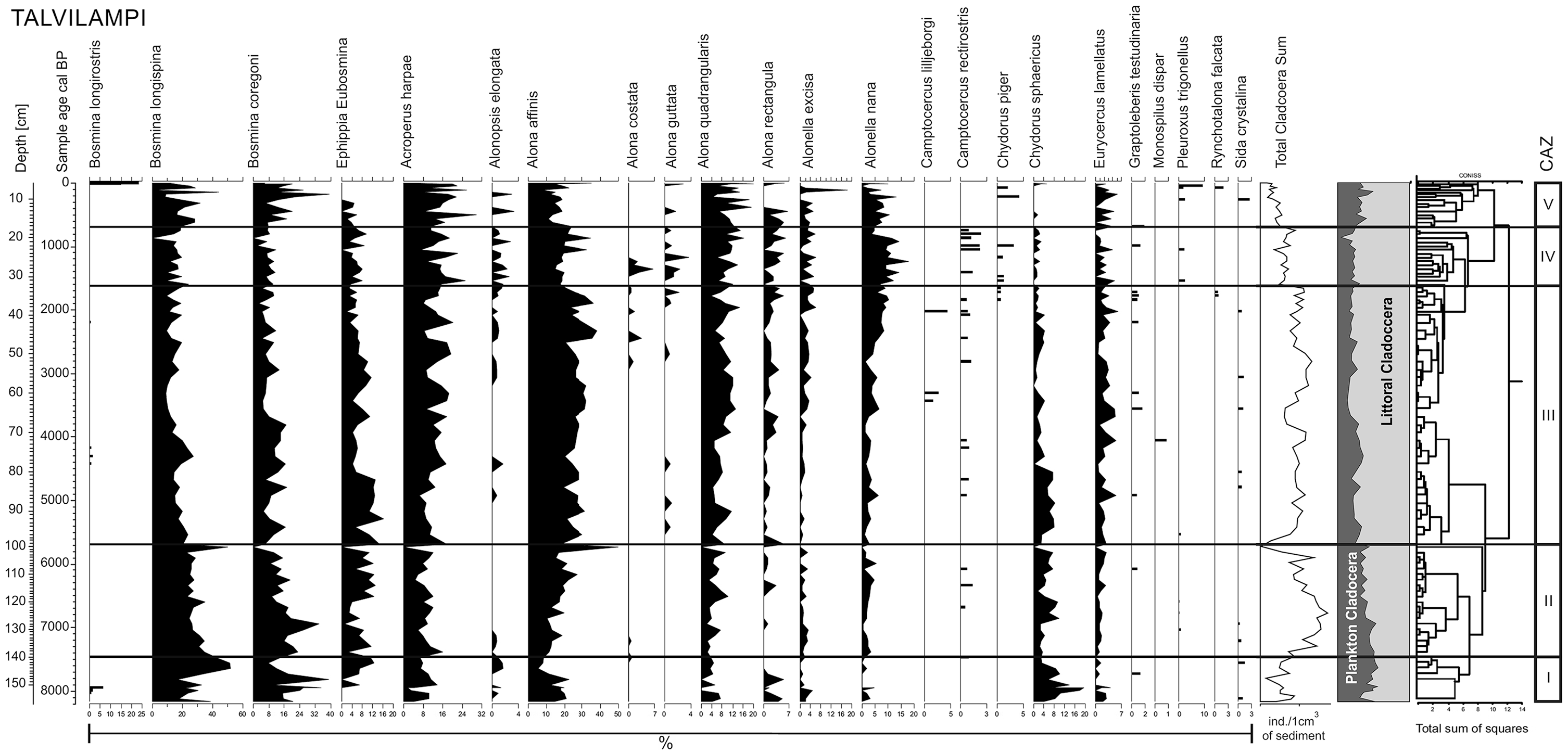
Cladocera diagram (Lake Talvilampi, Arctic Circle, Finland, core Talvilampi 3).
Results
Chronology
Ten Pb210dates give the age for the youngest layers of sediments. They revealed the accumulation of l cm of sediment over more than 30 years on average, except for the uppermost 3 cm layer (Table 1).
Results of Pb210 dating.
Two AMS 14C dates on terrestrial plant macrofossils were obtained from the studied sediments. Three others were done on pollen concentrates and pollen extractions generally followed Brown et al. (1989) (Table 2). Pinus sylvestris bud scales and bark were selected from the depth of 17 cm; however, no more than 0.02 mg C was obtained, so the sample was too small for dating. The date from the depth 32/33 cm Beta 38388 was too old and not in chronological order and so it was omitted. The calibrated ages of the other four samples are in stratigraphical order. Based on these data, the age of the sediments from the Tal3 succession was determined. According to the age-depth model (Figure 2) the accumulation of bottom sediments of core Tal3 started over 8000 cal. BP.
Results of C14 dating.

AMS: accelerator mass spectrometry.
Sediments
First description of sediments was made during the field work. Then the observation during laboratory preparation of samples and geochemical results made us modify this classification. Detritus gyttja with fine mineral matter, dark grey in colour, was the only component of the core Talvilampi 3. The description is: 156–134 cm – Ld1, Ag3; 134–1 cm – Ld2, Ag2 (Figures 4 and 7).
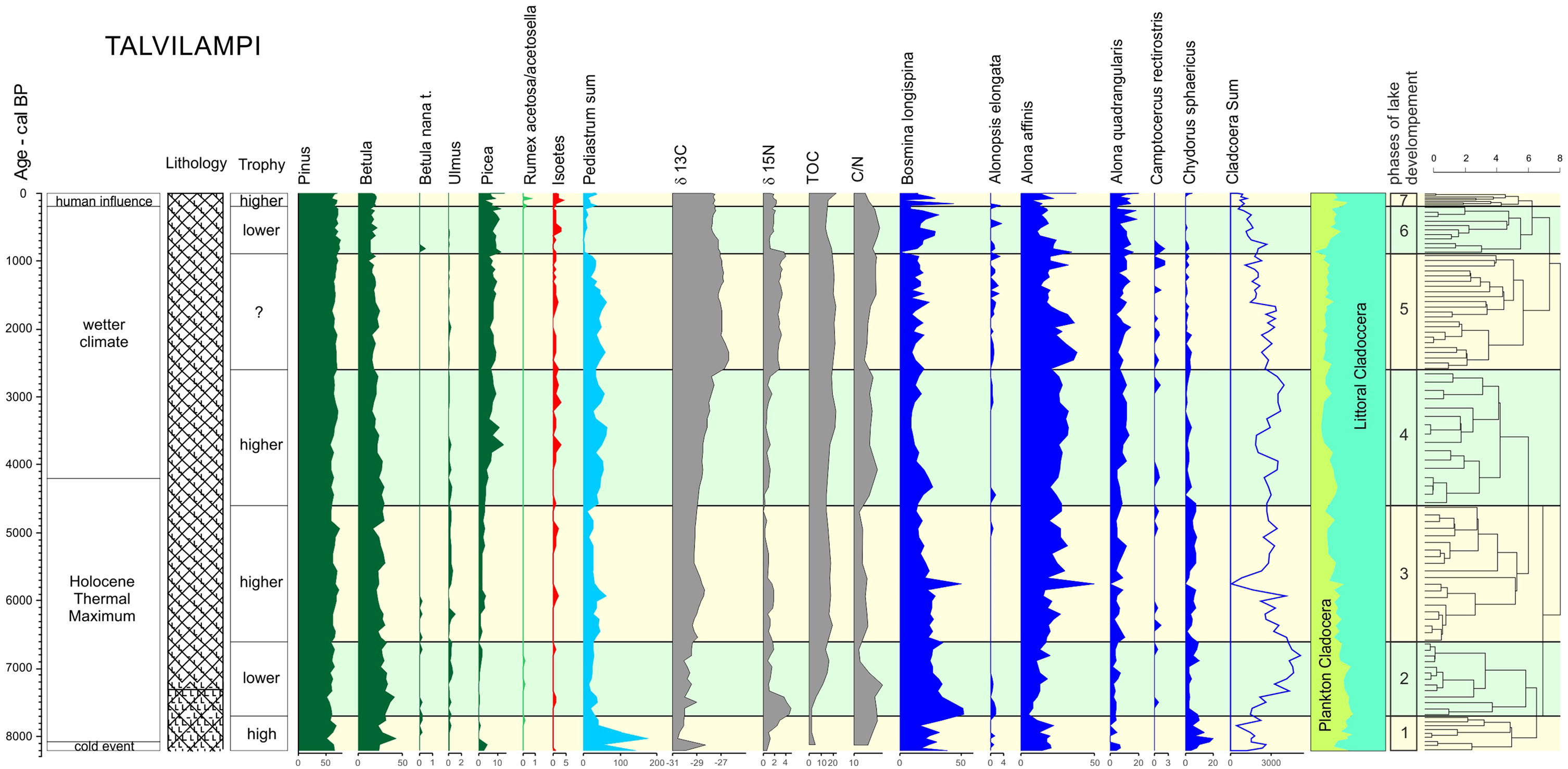
Changes of local and regional environment in the eastern part of Arctic Circle Finland based on analyses of Talvilampi 3 sediments.
Isotopic and geochemical data
Carbon to nitrogen ratio (TOC/TN) is an indicator for the nitrogen limitation of plants and other organisms. TOC/TN ratio indicates the primary source of OM in a lake, for example, TOC/TN ratios in the range 4–10 are usually from marine sources, whereas higher ratios are likely to come from a terrestrial source. The TOC/TN ratios less than 10–12 are typical for a mainly algal source and those greater than 20 are suggestive of a terrestrial plant origin. Therefore, TOC/TN ratios of sediments have frequently been used to infer the relative proportions of organic detritus derived from algae and terrestrial plants (Meyers and Lallier-Vergès, 1999; Smeltzer and Swain, 1985; Tyson, 1995).
In the studied sediments, the amounts of TOC and TN rose from 2% to 22% and from 0.2% to 1.8%, respectively. The TOC/TN ratio fluctuated between 11 and 16. The carbon isotope ratio varied between −30.6‰ and −26.3‰, while the nitrogen isotope ratio changed between 0‰ and 4.9‰. From the results of geochemical and stable isotope analyses, six geochemical-isotopic zones (GI-1–6) were defined (Table 3).
Geochemical-isotopic zones.

TOC: total organic carbon; TN: total nitrogen.
Pollen
Results of the pollen analysis let us define four pollen assemblage zones (Table 4). They show dominant taxa in each stage of environmental changes. Pinus sylvestris create the highest curve and fluctuate mostly between 65% and 75%. Betula is the second important taxon. Alnus and Juniperus have lower percentages. Very small curves of mesophilous trees (Quercus, Ulmus, Tilia, and Fraxinus) are not continuous and do not exceed 0.5%, except for a few samples of Ulmus. In PAZs III and IV, the clear presence of Picea is noted. Some more details are shown in Table 4. All the results are shown in two diagrams: trees, shrubs and terrestrial herbs (Figure 4), and telmatophytes, aquatics and algae (Figure 5). Certain rare pollen types of no significant importance for the interpretation are not included in the diagrams.
Description of pollen assemblage zones.

Macroremains
The quantities of macrofossils were very low, leading the interpretation to be poor and problematic. Thus, completely investigated were only samples selected for dating, and remains were described only qualitatively. Even fossils designed for dating were extremely rare and small.
List of macrofossils determined when selecting remains for radiocarbon dates in the Tal3 core:
Tal 17 cm: Pinus sylvestris bud scales and bark (for dating), Sphagnum leaves, Sialis mandibles and tergum, Trichoptera case, Cristatella mucedo statoblasts, Chironomidae head capsules.
Tal 48 cm: Pinus sylvestris bud scales and needles (for dating), Sialis sp. mandibles, labrum and tergum, Trichoptera case, Cristatella mucedo statoblasts, Chironomidae head capsules, Oribatida, Ephemeroptera mandible, single sand grains.
Tal 90 cm: Pinus sylvestris bud scales and needles (for dating), Betula sp. fruit and bark, Bryales stems (leafless), Nymphaeaceae sclereids, Sialis sp. mandibles and labrum, Chironomidae head capsules.
Tal 145 cm: Pinus sylvestris bud scales and needles, Betula sp. fruit (for dating), Bryales stems (leafless), Chara oospore, Sialis sp. mandibles and tergum, Chironomidae head capsules, Oribatida, Daphnia ephippium, Cristatella mucedo statoblasts, a lot of sand grains.
Cladocera
A total of 22 Cladocera species belonging to three families were identified in the sedimentary sequence of Lake Talvilampi. Planktonic species were represented only by one family – Bosminidae, whereas littoral species by two: Chydoridae and Sididae. Littoral species represented the major (~70%) component of the Cladocera assemblages, especially through the last 5600 years (Figure 6). Cladocera assemblage composition and structure allowed the identification of five Cladocera assemblage zones (CAZs) that summarise the main stages of Cladocera development in Lake Talvilampi (Table 5, Figure 6).
Cladocera assemblage zones (CAZs) distinguished in sediments of Lake Talvilampi.

Vegetation and lake development phases
Phase 1: 8200–7700 yr cal. BP
The bottommost sediments were accumulated during the turn of the early Holocene and the climatic optimum (acc. to Janbu et al., 2011). The area was covered with boreal vegetation, with Pinus sylvestris as a dominant tree and Betula and Juniperus admixtures. Small amounts of Corylus and Ulmus pollen reveal the near limit of the range of these mesophilous trees typical for Central Europe and broad-leaved forests; however, they were not confirmed as being present at the site, as the percentages are too small (Huntley and Birks, 1983; Ralska-Jasiewiczowa et al., 2004). Dwarf shrubs and herbal plants (below 1% of calculation sum except Poaceae) appeared in the forest bottom (Calluna, Vaccinium, Pteridium aquilinum) or on wet areas related to the lake (Filipendula, Viola palustris, Carex sp.).
In this phase, very low values of TOC as well as TN (ca. 2% and 0.2%, respectively), and a TOC/TN ratio of 14–15 were recorded (GI-1, Figure 3). The values of δ13C were the lowest in the profile (below −30‰). The rapid increase in δ15N suggests lower nitrogen availability in the surface waters (Talbot and Laerdal, 2000). Such geochemical data suggest a low amount of OM (low values of TOC and TN) derived from land (higher values of TOC/TN). Cladocera fauna and pollen indicate the presence of both groups of species: medium and cold tolerant. In the bottommost sediments, the presence of Alonopsis elongate (till 7200 BP), which is a cold-tolerant species typical for northern European lakes (Szeroczyńska and Zawisza, 2011a), was noted. Cladocera species composition (e.g. maximum occurrence of Chydorus sphericus) also indicates that at that time water fertility was quite high. This observation is confirmed by palynological analysis, especially the presence of high amounts of Pediastrum boryanum and Scenedesmus (over 150% and 250% of the calculated sum, respectively). Both pollen and Cladocera analyses indicate that lake waters at that time were relatively cold but with quite a high availability of nutrients.
It is highly probable that the bottom sediment sequence (154–156 cm) and the beginning of this lake development was concurrent with the 8200 ky cold event or followed it swiftly.
Such conditions suggest that phase 1 was the initial phase for the lake. According to Stroeven et al. (2016), ice-cover recession in this area took place 10,200 BP, so melting of Lake Talvilampi is suggested to be nearly 2000 years after the deglaciation. Geomorphological conditions of the lake indicate such a timing to be possible because of its location in a river valley with continuous drainage of water. Moreover, dead ice was probably covered with a thick layer of mineral sediments from the slopes closely surrounding it. Both drainage of the water and the significant sediment cover favoured the maintenance of pieces of dead ice, postponing melting of the lake (Błaszkiewicz, 2007; Homann et al., 2002).
Phase 2: 7700–6600 yr cal. BP
Regional vegetation succession reached its final, climax phase during the Holocene thermic optimum (Iversen, 1964b; Tobolski, 1976). Pollen grains of Tilia and Fraxinus and a slightly higher curve of Ulmus is a result of the line of the maximum northern range of these broad-leaved tree species which runs nearby (Huntley and Birks, 1983). Values below 1% of the calculated sum in the diagram indicate an absence of elm in the research area (Huntley and Birks, 1983; Zachowicz et al., 2004), and that during all the Holocene it was outside the range of these broad-leaved tree species. Growing Picea curve records the slow spread of this tree to Scandinavia reaching the southeastern area of Finland during phase 2 (Moen, 1999).
The amount of OM in the lake rose rapidly (TOC and TN increased from 7 to 15 and from 0.5 to 1.3, respectively, GI-2, Figure 3). The higher value of the TOC/TN ratio (16) in the bottom part of the phase suggests the continuing supply of OM of terrestrial origin into the lake (Meyers and Lallier-Vergès, 1999). Next, a drop in TOC/TN ratio to the lowest values of 11 (minimum values throughout the core), the low δ13C values (around −30‰) and a rapid drop of δ15N to around 1‰ reflect a prominent shift in the lake ecosystem and the increasing share of the freshwater algae component in the OM deposited at the lake bottom (Leng et al., 2005). The geochemical data suggest an enhancement of primary productivity (Ji et al., 2005) caused by an amelioration of environmental conditions, for example, climate warming.
The percentage of planktonic Cladocera species, particularly from the Eubosmina group, was highest of all the studied sequences. The dominance of planktonic species confirms the occurrence of an open water zone and probably indicates a high water level in the lake, which corresponds closely with the result of geochemical analyses. The availability of nutrients in Lake Talvilampi is reflected in Cladocera species composition (Figure 6), especially the significant increase of Cladocera individuals (up to ca. 5000 individuals in 1 g of dry sediment), both planktonic (Eubosmina) and littoral species (e.g. Alona affinis, Alona quadrangularis, Acroperus harpae). The increase of Cladocera individuals suggests that during phase 2, good environmental conditions for zooplankton development existed in the lake. An increase in trophy and better climatic conditions are not confirmed by Pediastrum (30% of calculation sum) and Scenedesmus content, because they fall down below 5% of their values observed in phase 1 (Figure 5). Rises in water level, increases in water volume, the exhaustion of nutrient resources following the initial phase and zooplankton pressures were the probable reasons for such a fall in the algae record.
Phase 3: 6600–4600 yr cal. BP
Pine forest was still the dominant plant community in Arctic Circle Finland. However, a change was observed with the spread of the Picea range and its admixture on wet areas of the region. Concurrently, Betula nana disappeared 5900 BP. It is not identified in pollen spectra up to the youngest layers of sediments, except for a single grain found in the sample dated for 800 yr BP (Figure 4).
The content of TOC and TN were relatively stable (about 17% and 1.4%, respectively; Figure 3). The δ13C values were also stable. The δ15N dropped from 1.8‰ to 0.1‰ (Figure 2). The TOC/TN values oscillated around 11–12. Such results show stable environmental conditions in the lake ecosystem and its sustainable development.
During this time, the total sum of Cladocera fell from about 3000 to ca. 50 individuals in 1 g of dry sediment and rose again to ca. 3000. However, the number of Cladocera taxa (21) was still quite high. The total sum of Cladocera indicates good conditions for fauna development. The increase in the littoral species (especially the subfamily Aloninae) living in association with water plants suggests – like isotopic data – development of macrophytes in the lake shore. This is in agreement with the pollen diagram revealing the presence of many plants living in the water or surrounding wetlands: Cyperaceae (Carex t., Schoenus t.), Filipendula, Potamogeton and some others (Figure 5). Most of these existed in various, relatively rich plant communities in the area along the lake shore. In deeper parts of the lake, Isoëtes communities developed (I. lacustris and I. echinospora) concurrently with the fall back of algae content (Pediastrum, Scenedesmus). This suggests lower trophy of the water with low conductivity (soft-water lake).
Phase 4: 4600–2600 yr cal. BP
Pine forest still dominated; however, Picea patches developed. This species spread in northern and western directions from the southern part of Finland from 5000 yr BP (Moen, 1999). Generally, development of the lake was still stable but some changes are observed. The amount of TOC and TN increased ca. 5% and 1%, respectively, and later slightly dropped. The TOC/TN ratio rapidly grows to 15 and later stabilised at a 13–14 ratio. The δ13C and δ15N regularly rose from below −29‰ to −27‰ and from 0.02‰ to 2.5‰, respectively, in the uppermost samples particularly rapidly. The increase in TOC, TN, δ13C, and δ15N suggests a further intensification of OM accumulation, partly supplied by OM of terrestrial and macrophyte origin (GI-3, Figure 3). The increasing values of δ13C and δ15N may reflect the start of progressing eutrophication. Stable, but higher than in the previous phase, curves of algae and littoral Cladocera could also indicate rising trophy. Littoral taxa Alona affinis grows at the expense of Chydorus sphaericus (Figure 6). Isoëtes population disappeared for ca. 500 years and then developed again enriched in I. Echinospora (Figure 5). This reveals rather poorer trophy conditions because of the higher oligotrophic preferences of the latter species. Decreasing values of TOC from 3200 BP favoured the development of Isoëtes sp. population. These have the ability to exploit various sources of C, even at night because of their crassulacean acid metabolism so they are competitive for other species of macrophytic aquatics in nutrient sufficiency (Boston and Adams, 1987).
Phase 5: 2600–900 yr cal. BP
The regional vegetation did not change: Pinus forest existed with developing Picea patches dependent on soil and local conditions.
The amount of organic carbon and nitrogen oscillated slightly and remained at a comparable level (GI-4, Figure 3). The TOC/TN ratio rose from 12 to 15. The δ13C slightly dropped and δ15N remained at a high level (3.5–4‰). The high δ13C and δ15N values suggest an enhancement of aquatic vegetation and further eutrophication. The higher values of δ13C can reflect the possibility of the occurrence of a strongly stratified water column with anoxic bottom waters. During the period of higher rates of photosynthesis, transfer of OM to the hypolimnion and burial of OM in sediments, the photic zone may be depleted of light isotopes. Especially high rates of primary productivity may deplete the water column of free aqueous CO2 in a relatively shallow lake (Brenner et al., 1999; Talbot and Laerdal, 2000). The geochemical data suggest the lowest water level and the highest trophy about 900 yr cal. BP. Among Cladocera re-development of the population of Alonopsis elongate indicates fewer nutrients in the lake and probably colder waters than in the previous phase. Such conditions favoured the Isoëtes population, its growth and the lack of more competitive species. Isoëtes occurs in poor water (Seddon, 1972) and carbon necessary for photosynthesis is additionally taken by roots from the sediment (Boston and Adams, 1987). The lack of light does not limit the population of Isoëtes because it mostly develops at the depth of 2–4 m (Szmeja, 2001); however, it has been observed even at the depth of 10 m (Podbielkowski and Tomaszewicz, 1996). According to Birks (2000), Isoëtes in Norway is often a dominant species below 3 m of water depth. Thus, Isoëtes could develop a population independent of depth and light supply.
At the end of this phase, a low water level, change of trophy and limited availability of C in deeper parts of the lake were recorded.
Phase 6: 900–200 yr cal. BP
From 900 yr cal. BP, the significance of spruce increased, while that of Juniperus decreased. The reason for this is probably related to the light conditions and/or wetness. The increased number of Picea trees depleted light availability for the light-demanding juniper (Zarzycki et al., 2002). On the contrary, the wetter climate of the late-Holocene favoured Picea and not the drought-resistant Juniperus; however, light demands have a greater influence on its growth than the quality of the soil (Tomanek, 1994).
This phase is characterised by a drop in TOC and TN levels (of 6% and 0.5%, respectively; GI-5, Figure 3) as well as in δ13C and a rapid decrease in δ15N – from 4‰ to 1‰. In contrast to this, TOC/TN ratio increased to 15. The decrease in OM level and the slight higher value of TOC/TN ratio suggest an initial small reduction of primary production and that a higher proportion of OM was derived from the land. Clearly decreasing Pediastrum and Scenedesmus confirm such a trend (Figure 5). Also, total Cladocera sum successively decreases. Many of the littoral species disappeared (e.g. Chydorus sphaericus, Camptocercus rectirostris) or significantly diminished (e.g. from Alona and Alonella genus) (Figure 6). It is highly probable that this is as a result of reduction of nutrients level in water. The decrease in OM amount, less algae and Cladocera and higher proportion of terrestrial OM suggest a short-term deterioration of environmental conditions in the lake.
Phase 7: 200–0 yr cal. BP
In the last 200 yr cal. BP, two significant changes in regional vegetation can be observed. First, forest clearance as indicated by higher Betula, Juniperus and Poaceae contents. Second, pollen grains of broad-leaved trees such as Quercus, Tilia, Ulmus and Carpinus completely disappeared. Pollen grains of Rumex, Plantago lanceolata, Urtica and Chenopodiaceae show the development of some open anthropogenic areas as a result of sparse settlement and animal breeding. The amount of OM rose sharply, but TOC/TN clearly dropped (GI-6, Figure 3) suggesting a lower proportion of OM of terrestrial plant origin and a higher share of algae. Growth of both photosynthetic green algae (Pediastrum, Scenedesmus) and Cladocera species which have higher nutrient needs, such as Bosmina longirostris and Chydorus sphaericus, and a high proportion of planktonic Cladocera support the thesis of a high water level and an increase in trophic status during the last century.
Discussion
The development of the regional vegetation of Arctic Circle Finland based on the sediments of Talvilampi reveals changes during the last 8000 years, that is, the middle and late-Holocene. Throughout all Northern Europe, similar general climate changes and related vegetation development are observed (Rantala et al., 2015). Pollen data from Scandinavia have, however, revealed regional differences (Birks, 1986; Heikkilä and Seppä, 2003; Miller et al., 2008) which are apparently seen in the S-N transect of Finland. The climate conditions during the middle Holocene (8200–4200 BP, Walker et al., 2012) were relatively stable, warm and of low humidity (Korhola et al., 2000; Luoto et al., 2014). The bottommost samples up to 7000 yr cal. BP revealed the stage of well developed regional vegetation – pine forest. The few pollen grains of Ulmus and Quercus proved the nearby N limit of broad-leaved species typical for temperate zones of Europe (Huntley and Birks, 1983; Moen, 1999; Tutin et al., 1964). However up to 6000 BP, there is evidence of pollen grains of Betula nana type, a species characteristic of cold and wet conditions (Moen, 1999). Dwarf birch probably still benefitted development of wetlands during the early Holocene (Rantala et al., 2015) and suggested acidification during the middle Holocene (Bigler et al., 2002). Next, Betula nana grains appeared as late as ca. 400 BP during the ‘Little Ice Age’ (Figures 4 and 7). Dense plant cover developing from the beginning of the middle Holocene, gradually enriched with Sorbus aucuparia, Frangula, Rubus idaeus, Cornus mas and many herbs, inhibited the OM run-off to the lake, which restricted the availability of nutrients to algae and directly to the Cladocera. The magnification factor was the high water volume.
Advancing HTM caused the disappearance of cold-tolerant species of Cladocera, such as Alonopsis elongate (Szeroczyńska and Zawisza, 2011a) after 7200 BP. There a short-lived increase in OM supply from the land (7200 BP) and this was followed by a record of higher productivity in the lake. Warming and drying of the climate during the middle Holocene (Walker et al., 2012; Yu and Harrison, 1995) resulted in decreasing water volume and growth of trophy. According to Korhola and Weckström (2004), OM in lake sediments is controlled by the climate through such factors as soil erosion and transport of terrestrial OM. The abrupt decrease in TOC/TN after 7200 BP suggests a lower proportion of terrestrial and rapid growth of autochthonous OM (Meyers and Teranes, 2001; Rantala et al., 2015). Lower water levels, because of higher temperature and drier climate (Yu and Harrison, 1995), resulted in the change of planktonic/littoral Cladocera relationships and development of the lake-shore vegetation. High temperature and an increase in trophy to 6200 BP is indicated by the quality and quantity of Cladocera (high number of individuals and species with higher nutrient needs) (Bos and Cumming, 2003; Chen et al., 2010). Gradual enrichment of water bodies with nutrients during the middle Holocene is indicated throughout northern European areas. Janbu et al. (2011) revealed that 8000 BP was the beginning of stable conditions and only a slight increase in TOC and TOC/TN was observed then, but the authors indicated the warmer climate as the main driver of trophic changes. As shown in the Talvilampi ecosystem, the climatic factor is also significant; however, changes in OM from the catchment also play an important role and development of the lake is not linear towards eutrophy.
After the HTM up to 2600 BP, the environmental conditions were relatively stable; however, some parameters ranged to the low degree (Figure 7). The different (changing) source of OM in the lake because of development of telmatophytic plant communities could be the reason for this. A clear decrease in Cladocera individuals and an increased proportion of low-demanding Isoëtes could suggest lower trophy (Smolders et al., 2002). However, the high number of Cladocera species and macrophytes confirm the diversity of habitats within the lake ecosystem. The increasing values of δ13C and δ15N also reflect slow, but progressing eutrophication. After 2600 BP, water levels slowly decreased and primary productivity was high. Limited availability of C in deeper parts of the lake resulted in ongoing development of the Isoëtes population in Talvilampi. From 800 yr BP primary production in the lake lowered, which is manifested in an algae decrease, total Cladocera decrease and reduction of littoral Cladocera species. This is consistent with the deterioration of climate during the ‘Little Ice Age’ (Walker et al., 2012). Väliranta (2006) imply that changes occurring in the last few centuries are the result of shorter ice-free periods in the year.
Human disturbances are discussed mainly from 4000 BP or later and mostly in southern parts of the country (Alenius et al., 2017; Huttunen and Tolonen, 1975; Kuosmanen et al., 2016, 2018). In the vicinity of Talvilampi, human activity is barely noticeable and even then only in the youngest layers of sediments, when some species of ruderals (Artemisia, Urtica, Chenopodiaceae), pasture communities (Plantago lanceolata) or weeds (Centaurea cyanus) are found during phase 7. Concurrently, Pinus and Picea decrease and light-demanding Betula increases, showing a higher proportion of open plant communities. Therefore, except for the most recent time period marked by human influences, it should be accepted that in the research area, climate is the main direct and indirect reason for environmental changes. According to Kuosmanen et al. (2016, 2018), even in the south of Finland, climate changes are the main drivers of the environmental development.
Conclusion
The study of sediments from a small Finnish lake – Lake Talvilampi – allowed ecological changes to be followed over the last 8000 years. The study is also an attempt to identify traces of human activity in the region of the Arctic Circle. The comparison of pollen, Cladocera and geochemical data enabled the estimation of environmental conditions, such as water temperature and level, and the trophy state of this lake. The Cladocera and geochemical data show the environmental conditions that occurred in this lake and confirm the climatic changes postulated by pollen data.
Indicators of cold water were found only during the initial stage of Lake Talvilampi as a result of climatic change (i.e. the 8.2 ka event) and geomorphological conditions (probably the prolonged melting of dead ice pieces). After that time, temperature was higher and cold-demanding species disappeared.
Trophy was high at the beginning of the lake development. Flow of nutrients from the slopes surrounding the forming lake was the reason for the high trophy during the initial stage of its functioning. Then, trophy increased after the oligotrophic stage during the older part of the middle Holocene. A significant increase in trophy was found after 2600 BP. The impact of human activity is hardly traceable in Arctic Circle Finland throughout the Holocene Thermal Maximum. During the late-Holocene (after 4200 yr cal. BP), this impact is still weak and, even as late as the 20th century, only a few traces of human activity are recorded.
Isoëtes lacustris populations developed after the climatic optimum and during the period of increasing trophy of the lake. This is not in agreement with the ecological demands of this species, which is a typical component of oligotrophic lakes. We conclude, however, that this is because of the lack of human influence and the low concurrency of other aquatic macrophytes.
The above points lead us to the conclusion that long-term climatic shift has been the most important factor driving changes in the limnology of Lake Talvilampi. Local factors related to climate, such as changes of water level (and water volume), plant communities and supply of nutrients from the catchment, were the next drivers of trophy changes. An exception is the last, very short phase 7, where the presence of pollen from ruderal plants and weeds reveals slight human pressure on vegetation and the environment.
Footnotes
Acknowledgements
We thank Dr Julian Wiethold for careful revision and valuable comments on the manuscript.
Authors’ note
Field work of this study was founded by the government in Finland as LAPBIAT Project.
Funding
The author(s) received no financial support for the research, authorship and/or publication of this article.