
Abstract
The study of past environments, their ecology, and detailed changes through time has become an important task of environmental science. Records of paleoenvironment usually vary between regions owing to different influences of local climate, vegetation, relief, and depositional processes. Therefore, studying local sequences thoroughly allows paleoenvironmental and paleoecological reconstructions for particular regions, but it also provides important data that complement the global record. The Żarska Cave is an exceptional site, which owing to its thick and undisturbed Holocene sediments with very rich paleobotanical and paleozoological materials has become the most complete example of the uppermost Upper Pleistocene and Holocene cave deposits in the Polish uplands. The aim of our study was to understand paleoenvironmental changes in southern Poland, from the late Glacial to the late Holocene, which has been targeted by use of a detailed geological analysis accompanied by analysis of a wide range of paleobotanical, paleozoological, and archeological assemblages. All the results obtained have permitted characterization of the paleoenvironmental changes occurring in the area of the Polish Jura during the last >15 ka years. A particularly well-represented sequence covers the Allerød interstadial, which revealed the presence of forests with associated shade-loving mollusks and rodents. The beginning of the Holocene was clearly identified with an increase of shaded forest habitats, which developed in a still relatively cold climate with continental features, and with the first appearance of mesophilous deciduous trees. During the middle Holocene, unusual evidence for maple forests is documented, which developed before the formation of beech forests, typical for the late Holocene. The obtained sequence has great significance not only for the Polish Jura region, but also has wider implications for southern Poland and the vast area of the northern Carpathian foreland.
Introduction
The study of past environments and their detailed changes through time has recently become a key task of environmental science. The better understanding of paleoenvironments and their ecology can help to predict forthcoming changes, including the most important challenges of the modern world, such as climate change, natural habitat loss, decrease of biodiversity, or species extinctions. Paleoenvironmental records usually differ between regions because of local climatic influences, various types of vegetation cover, distinct relief, and depositional processes. Therefore, studying each region in detail allows to reconstruct the paleoenvironment and paleoecology of particular regions, and to complement the global record.
The northern Carpathian foreland is a huge area ranging from Moravia and Silesia, through the uplands of southern Poland to Podolia and Prykarpattia of western Ukraine. Although the large-scale changes of the Pleistocene terrestrial environments and ecology have been well studied for this region (e.g. Komar et al., 2009; Łanczont and Madeyska, 2005; Socha et al., 2016), current knowledge of the late-Glacial and Holocene environments is poor in relation to the neighboring lowlands. This is due to a lack of long, undisturbed sedimentary sequences, which would also be abundant in archeobotanical and paleontological material, and which could provide a record of millennial scale. Such record is well known from the northern lowlands, for example, from lacustrine and peat bog sites (Ralska-Jasiewiczowa et al., 1998; Wacnik et al., 2011), but such sites are sparse in the upland zone. The sites known from this region mostly reflect short-time deposition and provide a limited representation of the environment of the region. These are mostly the fills of karst fissures and tufa deposits (Alexandrowicz, 2000a, 2000b, 2001; Alexandrowicz and Rudzka, 2006). Another common type of sites for this region are caves, which could potentially serve as depositional basins for millennia-scale time intervals. They are especially numerous in the Kraków-Czestochowa Upland (southern Poland, also known as the Polish Jura), the central part of the northern Carpathian foreland. However, Holocene cave sediments are poorly known from the Upland. There are several reasons for this sparsity of data. The most important one is the generally poor preservation of Holocene deposits in caves of the Polish Jura. This is an effect of using cave sediments as fertilizer in agriculture, resulting in a large-scale exploitation of cave deposits during the 19th and the first half of the 20th centuries (Kowalski, 1951). Aside from this, caves might have been intentionally cleaned by people at times, as we suspect, to adapt the chambers for utilitarian purposes (Krajcarz et al., 2014a; Sobol, 1948; Valde-Nowak et al., 2014; Wojenka, 2018). Another factor is the focus of former speleologists on Pleistocene sediments. As a consequence, Holocene sediments have been poorly described in previous publications. Among the few cases of well-recognized late-Glacial and Holocene deposits preserved in caves in the Carpathian region, the best examples are the Býčí Skala cave system (Czech Republic) and the Nad Mosurem Starym Duża Cave (Poland). However, the sediments in these caves cover only a part of the Holocene period (Horáček et al., 2002; Nadachowski et al., 1989).
The well-preserved multilayered and long sequences of Holocene sediments, as the one found in the Żarska Cave, are extremely rare in the Carpathian forelands. The Żarska Cave has been recently excavated by a team led by JW and MW (Wojenka et al., 2016). During fieldwork, exceptionally preserved late-Glacial and Holocene deposits have been revealed. The sediments were extremely rich in subfossil material of mollusks, vertebrates, and plants. This provided an exceptional opportunity of an insight into the detail paleoenvironmental changes that occurred in the region approximately during the past 15,000 years. The aim of this study was to explore the rich material from this site and to use its geological and paleontological record to build up the comprehensive and multiproxy reconstruction of the late-Glacial–Holocene paleoenvironment of the northern Carpathians. Our detailed objectives were to (1) characterize the local changes of fauna and vegetation since the terminus of the Last Glaciation until the last centuries, (2) compare the observed sedimentary record and local paleoenvironmental changes with other relevant sites from Central Europe, and (3) supply the dataset of numerous local records of paleoenvironment known from Central Europe with this new and important sequence. This will provide data for the future assessment of the role of local and regional dynamics of natural processes in the development of the current state of the environment in the European continent.
The site
The fieldwork was the very first archeological investigation of the cave since 1879, when the famous Polish scholar, Gotfryd Ossowski verified its archeological significance with negative results (Wojenka et al., 2011). Since then, the cave in Żary was considered to be perspectiveless for over 130 years, thus the only known cave of this size located at Polish Jura, and not excavated earlier. This opinion lasted no longer than 2008 when A Górny and M Szelerewicz led a survey during which they discovered charcoal and pottery fragments (Szelerewicz and Górny, 2008). These promising results were the basis of our fieldwork undertaken from 2011. During our fieldwork, we have revealed extremely rich and well-preserved late-Glacial and Holocene deposits, which became the subject of multiproxy analyses undertaken by an interdisciplinary team. In this paper, we present the results of analyses of materials from the 2012–2015 excavations carried out in the Żarska Cave (Wojenka et al., 2016), and interpret trends in environmental changes during the late-Glacial and Holocene.
Żarska Cave (in Polish – Jaskinia Żarska) is located in the Żarski Gorge (in Polish – Wąwóz Żarski) ca. 20 km northwest of the city of Kraków. The gully is a dry tributary to the Racławka Valley, which is one of several deeply entrenched valleys draining the south part of the Kraków-Częstochowa Upland (Figure 1). The upland is composed predominantly of Upper Jurassic limestone, and thus the area is traditionally called the Polish Jura. Jurassic limestone hosts numerous though relatively small caves formed in the Neogene. During the cold and dry Last Glacial the whole area was blanketed by loess (Madeyska, 2002), whose thickness locally exceeded 10 m (Łanczont et al., 2015). At the end of the Pleistocene, the loess was subsequently redeposited and washed down the slopes and into the caves (Krajcarz et al., 2016; Pawelec, 2006). Tufa grew at the bottom of the valleys in the early and middle Holocene (Alexandrowicz, 2004; Pazdur et al., 1988), but ceased to grow in the late Holocene, which is associated with deforestation and the next phase of loess removal from the slopes (Gradziński et al., 2017).
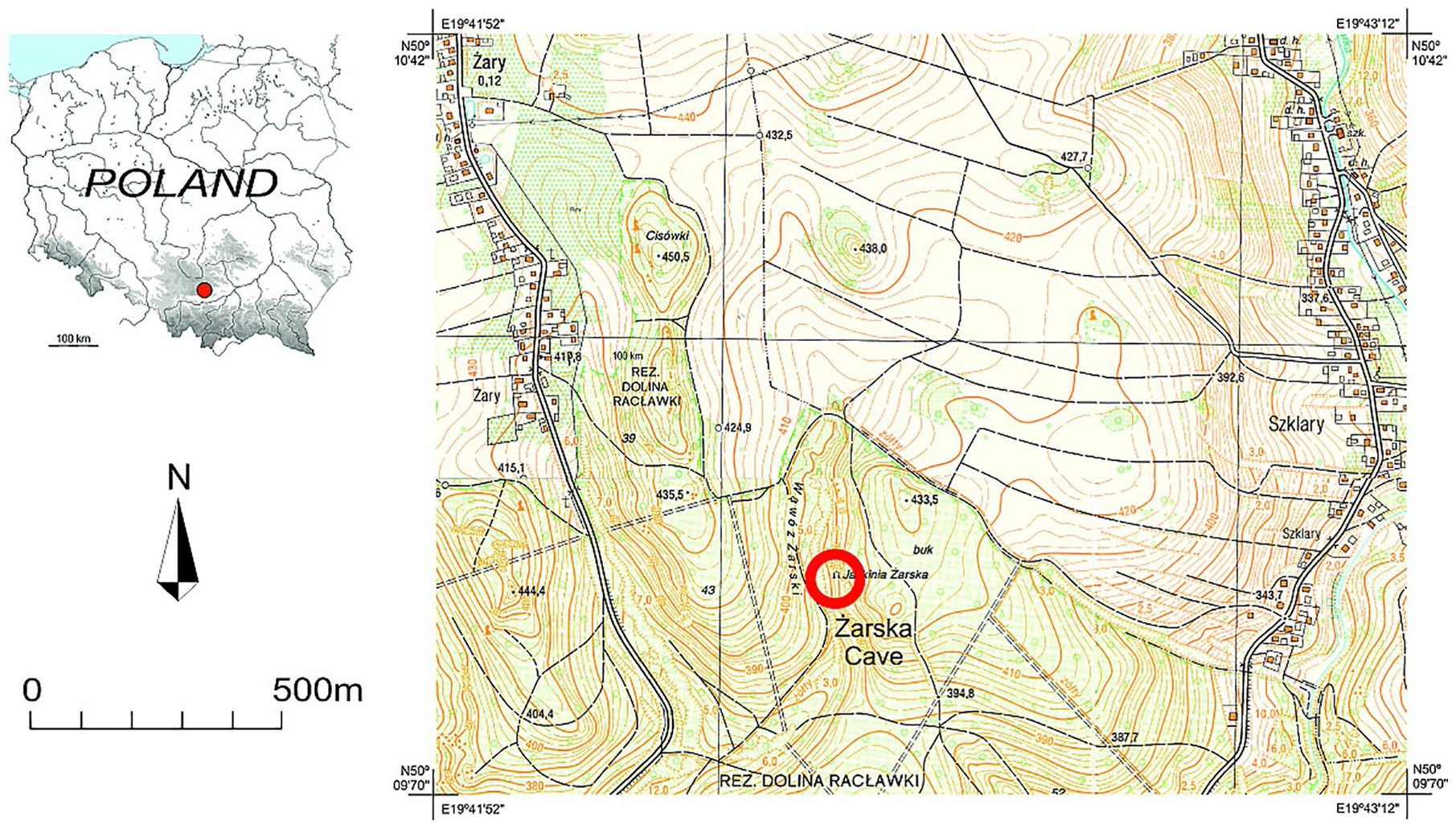
Location of the Żarska Cave.
The Żarska Cave is 143 m long (Szelerewicz and Górny, 2008) and it has a 23 m wide entrance, which leads to a laterally large but low chamber with some short and narrow off-passages. The cave is a remnant of a bigger inactive cave system dissected and partly destroyed by erosion. It represents a relict-type cave. The main chamber shows some traces of modeling by an underground water flow under phreatic conditions. The cave was formed along joint fractures. Some parts of it, including the entrance zone, were modified by tilting of limestone blocks and collapses guided by fractures. The time of the cave origin is unknown; however, a pre-Quaternary age of the Żarska Cave is hypothesized. During the Quaternary, the cave was breached and gradually filled with sediments. Some speleothems, namely, stalagmites, stalactites, and flowstones, occur in the cave as well. Flowstones comprise black laminae which were colored by minute charcoal particles and soot associated with prehistoric human activity (Gradziński et al., 2003).
Materials and methods
Our studies are based on materials discovered during fieldwork in the Main Chamber (trench 2) of the Żarska Cave by JW and MW (Wojenka et al., 2012, 2016). The trench is roughly 4.0 by 3.5 m, situated in the north-western part of the cave, and the area excavated is approximately 12.5 m2. All sediments including modern heaps were excavated by hand. Recovery methods included hand-collection and wet-sieving of all sediments using nesting screens of different mesh sizes. The positions within the excavation area of all archeological finds and animal remains over 2 cm in size were recorded in three dimensions (3D).
Taxonomically identified charcoal fragments were selected for radiocarbon dating, as such objects constitute the most appropriate material to be dated (Moskal-del Hoyo et al., 2017a; Moskal-del Hoyo and Kozłowski, 2009). Also, the selected identified human and animal remains discovered in different layers were dated. AMS radiocarbon measurements were performed in the Radiocarbon Laboratory in Poznań (see Goslar et al., 2004 for details). Calibrated radiocarbon ages (cal. AD/BC) were obtained based on the IntCal 13 radiocarbon calibration curve (Reimer et al., 2013) and OxCal 4.3.2 calibration software (Bronk Ramsey, 2009, 2017).
Macroscopic sedimentary structures were observed in the field on the vertical walls of the archeological pit, documented by drawings and photographs, while the inclinations of structures were measured with a geological compass. The sedimentary sequence was divided into ‘layers’ numbered from 1 to 19 (from top down), which were distinguished during the excavation. Samples for the micromorphological analysis of structures and textures were also taken and will be presented in a separate paper. Samples of sediment weighing approximately 5 kg were taken from each layer. The samples were sieved using 2, 4, 10, 20, 40, and 80 mm meshes to calculate the percentage of each fraction. The fraction below 2 mm was subjected to laser grain size analysis using a diffractometer Fritsch Laser Particle Sizer ‘Analysette 22’ in the Polish Geological Institute, Warsaw. The measuring range was 0.1–1884.48 μm. The programmed limits of fractions were 2 and 63 μm, corresponding to the boundaries of clay/silt and silt/sand fractions, respectively. The fraction above 20 mm was subjected to optical morphological analyses, following the methodology of Krajcarz and Madeyska (2010). Limestone clasts in each sample were divided into four morphological classes (A, Ba, Bb, C) according to the increasing roundness, and classes weighted to calculate the class proportion.
For geochemical analyses, a 2 g sample of homogenized <2 mm fraction was powdered, dried at 105°C, weighed, and decalcified with 10% hydrochloric acid (HCl) for 24 h on a shaking plate. The samples still bubbling were supplied with additional HCl and stored for the next 24 h. The samples were then centrifuged, rinsed three times with distilled water, oven dried, and weighed. The difference in weight was considered to be the calcium carbonate (CaCO3) content. Around 5 mg aliquots of sediments, previously decalcified and homogenized, were analyzed in a Vario MicroCUBE CHNS elemental analyzer to determine the content of organic carbon. Sulfanilic acid was used as a standard. The measuring accuracy is represented by the standard deviation ±0.6%.
For anthracological, malacological, and zooarcheological analysis, sediment samples of 5–8 L in volume were taken from various stratigraphic layers during the excavations. They were processed with the use of 0.5 and 1 mm meshes and sorted with stereomicroscopes.
Wood charcoal remains were studied at the W. Szafer Institute of Botany of the Polish Academy of Sciences with the help of a modern reference collection of wood and charcoals, and atlases of wood anatomy (Schweingruber, 1990a, 1990b). The anatomical sections of the wood of the charcoal fragments were observed using a reflected light microscope of up to 500× magnifications (Zeiss, Axio Lab. A1). The taxonomic identification is mostly limited to genus level, and species level is indicated when only one species grows in the present-day vegetation of the region (Lityńska-Zając and Wasylikowa, 2005). Analysis of ring curvature was performed (Marguerie and Hunot, 2007) and a presence of decayed wood was indicated (Moskal-del Hoyo et al., 2010). Also, a Hitachi S-4700 scanning electron microscope (SEM) was used for detailed analysis at the Laboratory of Field Emission Scanning Electron Microscopy and Microanalysis of the Institute of Geological Sciences of the Jagiellonian University (Kraków, Poland). The quantification method is based on fragment counts of charcoal remains (Chabal, 1988, 1997). The anthracological diagram, in the form of histograms, was plotted with the POLPAL software (Nalepka and Walanus, 2003). This diagram and the relative frequencies of taxa were prepared only on the basis of charcoal fragments identified with species and genus levels. They represent scattered material (dispersed charcoals), therefore they were used for plotting the anthracological diagram. The remains of a hearth from layer 3 (concentrated charcoals, Table 1 Appendix, available online) were excluded from this diagram.
Charcoal fragments were preserved in 126 sediment samples. In the samples from layers 7 to 11, the charcoal fragments were frequently small and scarce (<2 mm3), while from the upper layers they were frequently bigger (>2 mm3). The majority of the samples represent scattered materials (dispersed charcoals) that can be interpreted in terms of the paleoenvironment (Chabal, 1988, 1997; Moskal-del Hoyo, 2013; Ntinou, 2002; Théry-Parisot et al., 2010). The higher number of samples with charcoal fragments and therefore the largest assemblage came from layer 6, which was related with the human occupation of the cave. Charcoal materials from lower layers (7–11) probably originated from natural processes or short-term visits of people to the cave, whereas the charcoals found in the upper layers (1–5) were related to only sporadic human presence in the cave. In the case of layer 3, the remains of a hearth with a large amount of ashes were preserved.
The malacological analysis was performed in samples from the sediments filling the cave in close connection with the lithological types distinguished there. In total, the basis of the analysis was provided by more than 200 samples, representing 19 layers. In layers 13 and 19, no shells of mollusks were found. Five types of molluskan assemblages can be distinguished based on cluster analysis. The Morisita algorithm was used (Morisita, 1959). All calculations were completed using the PAST statistical software package (Hammer et al., 2001).
After flushing and taking out shell material, faunal remains were identified with the use of identification guides and comparative collections. The numbers of specimens were then determined in particular samples and summed in particular layers. The standard methods of malacological analysis (Alexandrowicz and Alexandrowicz, 2011; Ložek, 1964) were used for the paleoenvironmental analysis and for drawing stratigraphic conclusions.
Vertebrate remains were identified with the help of an osteological collection at the Institute of Systematics and Evolution of Animals, Polish Academy of Sciences in Kraków, and by the use of available identification keys (e.g. Bacher, 1967; Bocheński and Tomek, 2009; Erbersdobler, 1968; Fick, 1974; Gromova, 1950; Kraft, 1972; Pales and Garcia, 1981a, 1981b; Tomek and Bocheński, 2000, 2009; Woelfle, 1967). Taxonomical and anatomical analyses of fish bones were done with the help of reference collections in the Archaeological Research Collection of Tallinn University, Estonia. Latin taxonomic nomenclature for birds follows the work of Dickinson and Remsen (2013) and Dickinson and Christidis (2014). Two quantified calculations were made of the remains: NISP (Number of Identified Specimens) and MNI (Minimal Number of Individual Animals) (Klein and Cruz-Uribe, 1984; Lyman, 1994, 2008).
All the results obtained were used to determine the paleoenvironmental changes occurring in the area of the Polish Jura, and generally southern Poland, during the last several thousand years, with a particularly well-represented sequence covering the period from Allerød to modern times.
Results and interpretations
Basically, five principal phases of human occupation were identified, generally in two well-separated layers. The first human occupation phase is represented in layer 6, where numerous finds representing the Eneolithic Baden culture were found. Also sparse artifacts of the Eneolithic/early Bronze Age Strzyżów culture were found in this layer. The other three horizons were preserved as redeposited in topsoil-like silty to sandy loam of layers 1 and 2. These materials represent the late-Roman Period/early Migrations Period (Przeworsk culture, including two human skeletons), Middle Ages, and the Modern Period (Wojenka et al., 2016).
Radiocarbon dating
The dating results are listed in Table 1. The obtained radiocarbon dates are concentrated in the Holocene (mainly layer 6) with a single date for layer 10, which was determined to be Allerød interstadial (Table 1). Unfortunately, attempts to date the Pleistocene materials in layers 13–15 (Ursus sp. and Bos/Bison remains) were unsuccessful because of low collagen content. The rather large interval covered by the dates obtained for layer 6 is not surprising, and is also evidenced by the presence of diverse archeological material typologically dated to the Eneolithic period and the early Bronze Age. Human remains discovered in the cave can be clearly linked with the Roman Period thanks to radiocarbon dating.
Results of radiocarbon dating of the anthropological, paleobotanical, and paleozoological material from the Żarska Cave.
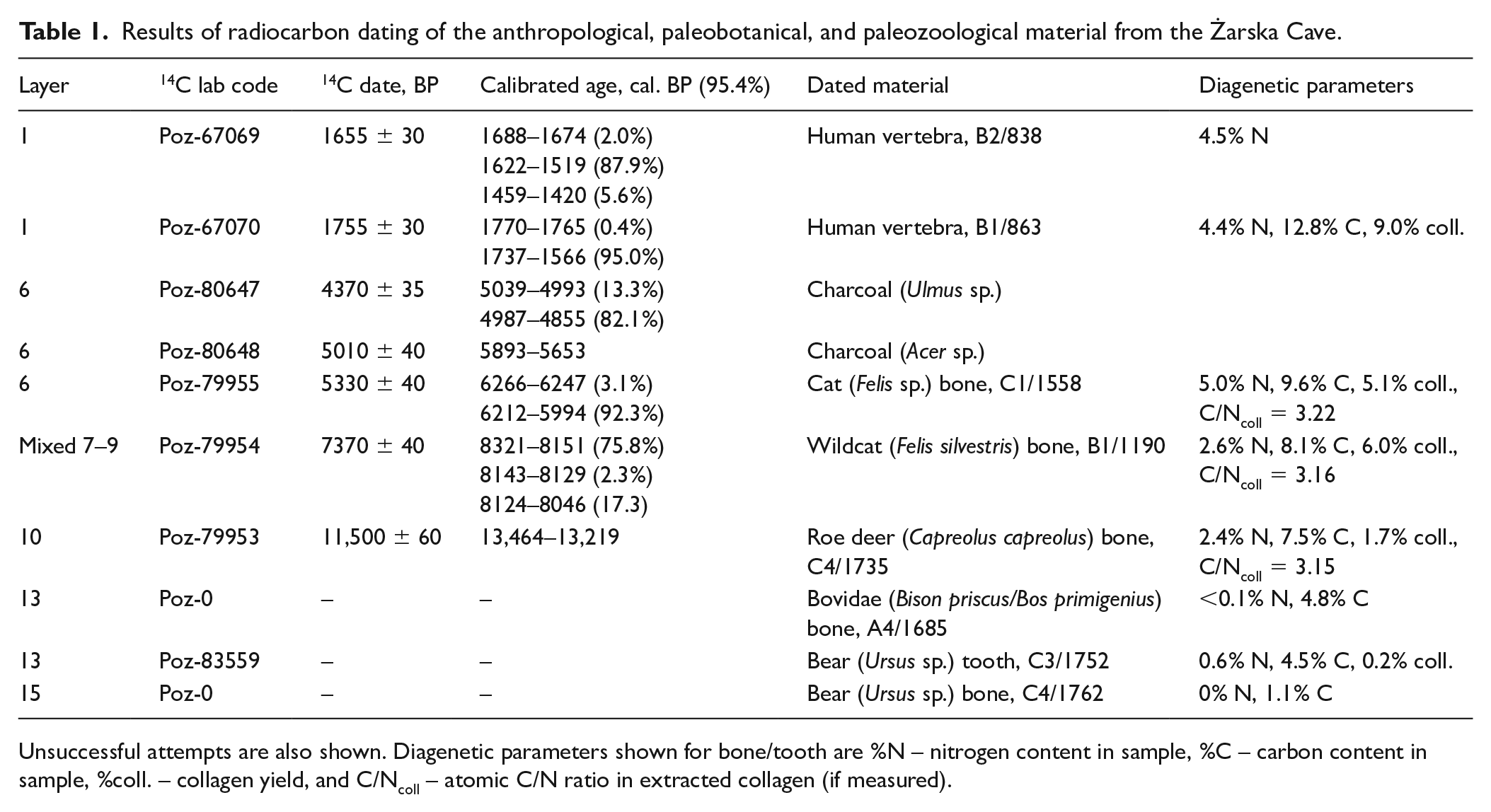
Unsuccessful attempts are also shown. Diagenetic parameters shown for bone/tooth are %N – nitrogen content in sample, %C – carbon content in sample, %coll. – collagen yield, and C/Ncoll – atomic C/N ratio in extracted collagen (if measured).
Geological analysis
The sediments of the Żarska Cave are clay silts or silty clays with variable abundance of limestone clasts (Figures 2 and 3), typical for caves of the Kraków-Częstochowa Upland (Krajcarz et al., 2014b; Madeyska, 1981, 1988; Valde-Nowak et al., 2014).

Lithofacies profile of sediments from the Żarska Cave. Only macroscopically visible sedimentary and post-sedimentary structures were marked.

Basic lithological and geochemical parameters of the sediments of the Żarska Cave.
A characteristic feature of the Żarska Cave is the distinct inclination of layers, marked especially in layers 2–10, in general with NE dips (toward the cave interior). Dip values range from 12° to 20°. The inclination in the whole series probably reflects the inclined cave floor formed after the sedimentation of layer 11. Lower layers are less inclined, except for layer 17 with a dip of approximately 6–7° to N. Several erosional boundaries occur in the section (Figure 2). At least one of them, the lower boundary of layer 6, is of anthropogenic origin. Where this layer occurs, the earlier sediments were partially removed, especially from the area near the east wall of the cave, possibly to enlarge the living space and prepare a waste pit for ashes. Other important erosional boundaries are situated at the bottoms of layers 8, 10, 11, and 16. Other sedimentary structures include shallow erosional channels, intralayer lamination, planar orientation of clasts, and imbrication. Most of these occur in the middle part of the sequence, in layers 8–17 (Figure 2). These structural features are typical for colluvial sediments and indicate redeposition by washing, mud flows, debris flows, and soil creep. Only two examples of post-sedimentary structures are present in the Żarska Cave. These are (1) unstable density stratification, seen at the boundary of layers 12/13, probably connected to the rapid deposition of the upper layer on the waterlogged sediment; and (2) admixed packets of layers 1–4 inside layer 5, connected probably with relatively late bioturbation.
The entire sequence may be subdivided into three lithostratigraphic series. The lowermost one contains layer 19 and is characterized by a lack of sand and gravel fractions and high content of carbonate and organic matter (Figure 3). The middle series contains layers 12–18, which are silty sediments with variable amount of limestone clasts, with traces of colluvial activity, and low content of carbonate and humus. The upper series contains layers 1–11, characterized by increased amount of sand, generally high carbonate and humus content, high amounts of limestone clasts, and smoothness of clasts (Figure 2).
Lithological and geochemical characteristics (Figures 2 and 3) taken together with chronometric data (Table 1) allow to propose the general chronostratigraphic scheme for the sedimentary fill of the Żarska Cave. The stratigraphic attribution is also supported further by the data derived from the remains of paleovegetation and fauna. The upper series represents mostly the Holocene. The sediments of the middle series represent the Pleistocene, most likely Upper Pleistocene on the basis of lithological similarity to sediments known from other caves in the region (Madeyska, 1988). The lower series may be much older, as analogous bottom clayey deposits are known from the other caves of the region, such as the Biśnik Cave or the Żabia Cave, which are dated to the early Pleistocene or the pre-Pleistocene (Bosák et al., 1982; Krajcarz et al., 2014b).
Anthracological analysis
More than 1800 charcoal fragments were analyzed from the Żarska Cave. A total of 19 taxa identified to species and genus were found: Abies alba, Cornus sanguinea, Carpinus betulus, Corylus avellana, Fagus sylvatica, Fraxinus excelsior, Juniperus communis, Picea abies or Larix decidua, Pinus sylvestris, Taxus baccata, Viscum album, Acer sp., Betula sp., Prunus sp., Quercus sp., Salix sp. or Populus sp., Sambucus sp., Tilia sp. Ulmus sp. (Table 1 Appendix, available online).
The charcoal assemblages from all layers were characterized by the presence of branchwood and fragments infected by fungi. These features were especially detected in charcoals found in layers 4 and 6 and in the hearth from layer 3. In layer 6, a relatively high number of charcoals of Fraxinus excelsior had very narrow rings.
The results of charcoal analysis based on fragments coming from various layers are summarized in a diagram that is divided into five local charcoal assemblage zones (Figure 4). The limits of the zones were determined by the appearance, disappearance, increase, or decrease in numbers of taxa that are quantitatively significant. The first appearance of charcoal fragments was noted in layer 11 and the first local zone was distinguished, with charcoal assemblages coming from layers 10 and 11. This local zone is characterized by a dominance of Pinus sylvestris (Figure 5a and b), followed by Betula sp. and Picea abies or Larix decidua. A few fragments of Salix sp. or Populus sp. were also found. The second local zone (layers 8 and 9) is also dominated by P. sylvestris, followed by Betula sp. and Picea abies or Larix decidua (Figure 5c), but also with the first appearance of mesophilous deciduous trees and shrubs such as Ulmus sp., Corylus avellana, and Acer sp. The third local zone (layers 6 and 7) differs substantially from the taxonomic composition of previous layers as it is characterized by a decrease of P. sylvestris and a dominance of Acer sp. (Figure 5f and g), Ulmus sp., Fraxinus excelsior (Figure 5d and e), and C. avellana. The highest taxonomic richness was observed in layer 6. In the overlying local zone (layers 4 and 5) the percentage of P. sylvestris and Quercus sp. charcoals increased. The second highest taxonomic diversity was in layer 4. The most recent local zone (layers 1 and 2) is characterized by a very low incidence of charcoal remains, but with the highest share of Fagus sylvatica (Figure 5h and i).

Anthracological diagram from charcoal assemblages from the Żarska Cave.

Selected taxa from the Żarska Cave. (a, b) Pinus sylvestris, transverse section (TS) and longitudinal radial section (RLS), scale bars: 500 and 100 µm. (c) Picea abies or Larix decidua, RLS, scale bar: 50 µm. (d, e) Fraxinus excelsior, TS, scale bars: 1.00 mm and 200 µm. (f, g) Acer sp., TS and longitudinal tangential section (TLS), scale bars: 400 and 200 µm. (h, i) Fagus sylvatica, TS and TLS, scale bars: 500 and 300 µm. Micrographs: M. Moskal-del Hoyo.
Animal remains
The malacofauna is greatly diversified within the section, seen both in species composition and in the structure of assemblages, corresponding to the changes in environmental conditions generated chiefly by climatic factors (Figure 6). It included more than 45,000 specimens of 71 species of mollusks (Table 2 Appendix, available online). The number of specimens varied from 102 (layer 15) to 21,509 (layer 6), while the number of species ranged from 6 (layers 14 and 17) to 66 (layer 6).
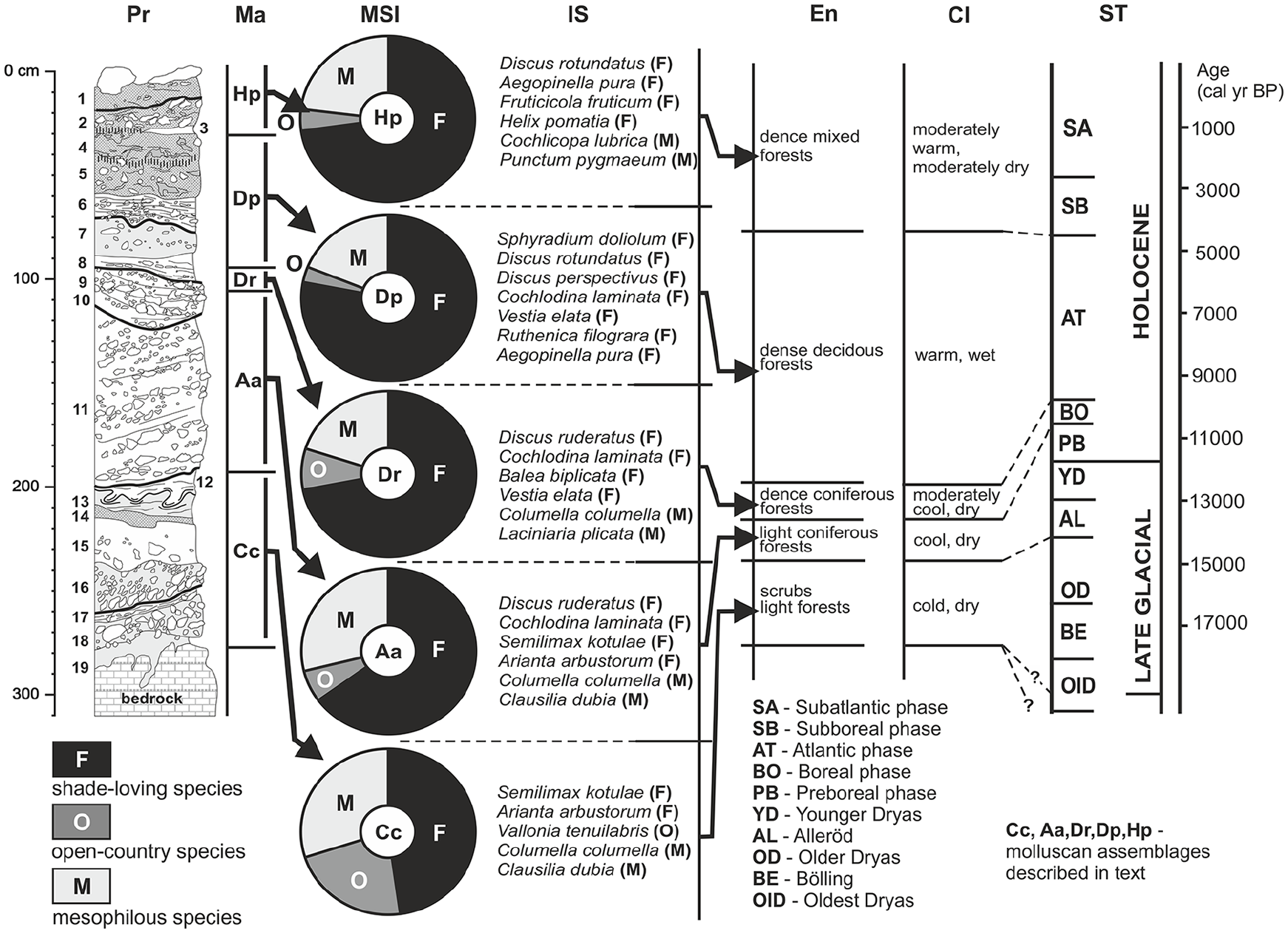
Malacofauna of the late-Glacial and Holocene deposits from the Żarska Cave.
The lower part of the sequence, including layers 12–18, contains a poor fauna (assemblage with Columella columella – Cc). Only species with wide thermal tolerance adapted to cold climatic conditions were identified there. The characteristic feature in this section is the abundant occurrence of Vallonia tenuilabris. It is a species typical for loess deposits, which withdrew from Central Europe at the beginning of the late-Glacial (Alexandrowicz et al., 2014a; Ložek, 2001; Sümegi, 2005). Besides Vallonia tenuilabris, there are numerous shells of Columella columella, as well as species preferring more shaded habitats: Arianta arbustorum and Semilimax kotulae.
The sediments situated above (layers 11 and 12) contain more diversified fauna (assemblage with Arianta arbustorum – Aa). Shadow-loving species appear in higher numbers, particularly Arianta arbustorum. The taxa tolerating cold climate (Columella columella, Semilimax kotulae, Discus ruderatus) play an essential role in the described fauna. However, Vallonia tenuilabris, which is typical to the Pleistocene, disappears.
The malacoenosis identified in layer 9 is characterized by increased species diversity, particularly with reference to shadow-loving taxa (assemblage with Discus ruderatus – Dr). The most numerous is Discus ruderatus. Besides it, some other species of shaded habitats appear, including taxa having relatively narrow habitat requirements (Vestia elata, Ruthenica filograna), whereas species typical of cold climate (Columella columella, Semilimax kotulae) gradually disappear (Figure 6).
Numerous molluskan shells are present in layers 5–8, situated above layer 9. The identified fauna demonstrates very high species diversity (assemblage with Discus perspectivus – Dp). The most important components are typical forest species with narrow ecological requirements: Sphyradium doliolum, Discus rotundatus, Discus perspectivus, Cochlodina laminata, Ruthenica filograna, Vestia elata, and many others. In this interval, the frequency of cold-tolerant species (e.g. Discus ruderatus) significantly decreases and the cryophilic taxa completely disappear.
The uppermost part of the sediments (layers 1–4) contains a markedly poorer faunal assemblage. The predominance of shade-loving taxa is maintained, but the change in species composition is evident (assemblage with Helix pomatia – Hp). The proportion of thermophilous taxa (Discus perspectivus, Vestia elata, Ruthenica filograna, Sphyradium doliolum) decreases at the expense of snails with broader environmental tolerance: Balea biplicata, Aegopinella pura, Acanthinula aculeata, and many others. The occurrence of Helix pomatia is the characteristic feature of this fauna.
The presence in layers 3 and 4 of the shell of freshwater bivalve Unio pictorum is also noteworthy. It is a foreign element whose presence in the sediments can be associated with human activities. The shells of this species were often used for decorative or economic purposes (Ożgo et al., 2012).
Among the other faunal finds, fish bones are not numerous. They were mostly found in sediments of layers 11–18, and are interpreted as non-anthropogenic accumulation being remnants of prey collected by birds and mammals. Two fish bones are also associated with the Eneolithic sediments, but the human role in this small fish deposit is also not clear. A total of 79 fish bones were taxonomically and anatomically analyzed (Table 3 – Appendix, available online). The burbot (Lota lota) predominates in the materials of both late Pleistocene and early Holocene layers, followed by some whitefish (Coregonus) species (most probably vendace, Coregonus albula). Other fish were represented by only few bones: pike (Esox lucius) by three vertebrae, bullhead (Cottus sp.), some cyprinid, probable trout (Salmo sp.), and grayling (Thymallus thymallus) by one vertebra each. From the Eneolithic layer only two fish vertebrae were found: one from vendace and the other from a cyprinid. All identified fish species are those of cold adapted, which means that they can live in cold as well as temperate waters and are therefore quite useless in late-Pleistocene/early Holocene climate reconstructions.
The whole assemblage yielded 1173 avian bones from at least 138 individuals of at least 46 taxa (Table 4 – Appendix, available online). Galliformes and Passeriformes are the best represented groups, and only a few well-separated sediments (e.g. layers 1, 6, and 15) yielded sizable numbers of identifiable remains. Layers 13–18 are dominated by two galliform species, willow grouse and black grouse. Another identified species from the same layers, song thrush, is typical of mixed broad-leaved and coniferous forests or forest edges, and mistle thrush from the layers is a forest species. Two other forest species (capercaillie and wryneck) were identified in mixed Pleistocene layers, as well as one species (corncrake) that occupies open environments. Layers 8–11 yielded small numbers of bird remains. Capercaillie and black woodpecker in the assemblage suggest the presence of heavy, thick forest near the cave, but other taxa (common starling and tawny owl) indicate the occurrence of more lightly wooded areas.
Sediments from layer 7 yielded single remains of at least 14 bird taxa (e.g. hazel grouse, tawny owl, wryneck, Eurasian wren, nuthatch, mistle thrush, and Eurasian blackbird). White wagtail from the layer inhabits rather open and grassy areas along rivers, albeit often by the edges of forests. Identified Eurasian scops owl occupies arid and lightly forested region, but not boreal forest or swampy forests. Eneolithic layer 6 contained numerous bones of hazel grouse, tawny owl, nuthatch, and thrushes. All these species are forest-dwelling. Less numerous are bones of a few species that occupy open environments (common quail, gray partridge, and northern goshawk). Younger sediments (layers 4 and 5) yielded remains of forest species (hazel grouse and thrushes) but also of those that occupy swampy/aquatic environments (short-eared owl) or open areas (corncrake). In the youngest layers (1–3), bones of domestic chicken dominate the bird assemblage. Remains of goose and duck that might have been bred by humans are also present. Other birds identified are forest-living species (e.g. hazel grouse, hawfinch, tawny owl, green woodpecker, and thrushes) or birds that occupy open areas (common quail, gray partridge, corncrake, and northern goshawk).
Rodents and lagomorphs are the most numerous groups of vertebrate remains in the cave sediments, numbering 6190 identified specimens belonging to 22 species (Table 5 – Appendix, available online). At the deepest layers, 14–18, the most abundant species is Microtus gregalis, an inhabitant of tundra and taiga-tundra environments (Figure 7). With it are two lemmings (Dicrostonyx gulielmi and Lemmus lemmus) which inhabit steppe-tundra. Species with a broad environmental tolerance range are also common: Microtus arvalis/agrestis (17–26%). The dominant species in this series of layers are supplemented by species indicating habitats abounding in water, Microtus oeconomus and Arvicola terrestris, and by Clethrionomys glareolus, a typical forest species tolerating cold climate at the end of the Last Glacial (Nadachowski, 1982). In layers 12 and 13 the steppe-tundra rodent species are less frequent (40–42%), while eurytopic Microtus arvalis/agrestis (up to 26–30%) and species connected with water increase in frequency (up to 23–25%). There also appears incidentally typical steppe species Cricetus cricetus and Cricetulus migratorius.
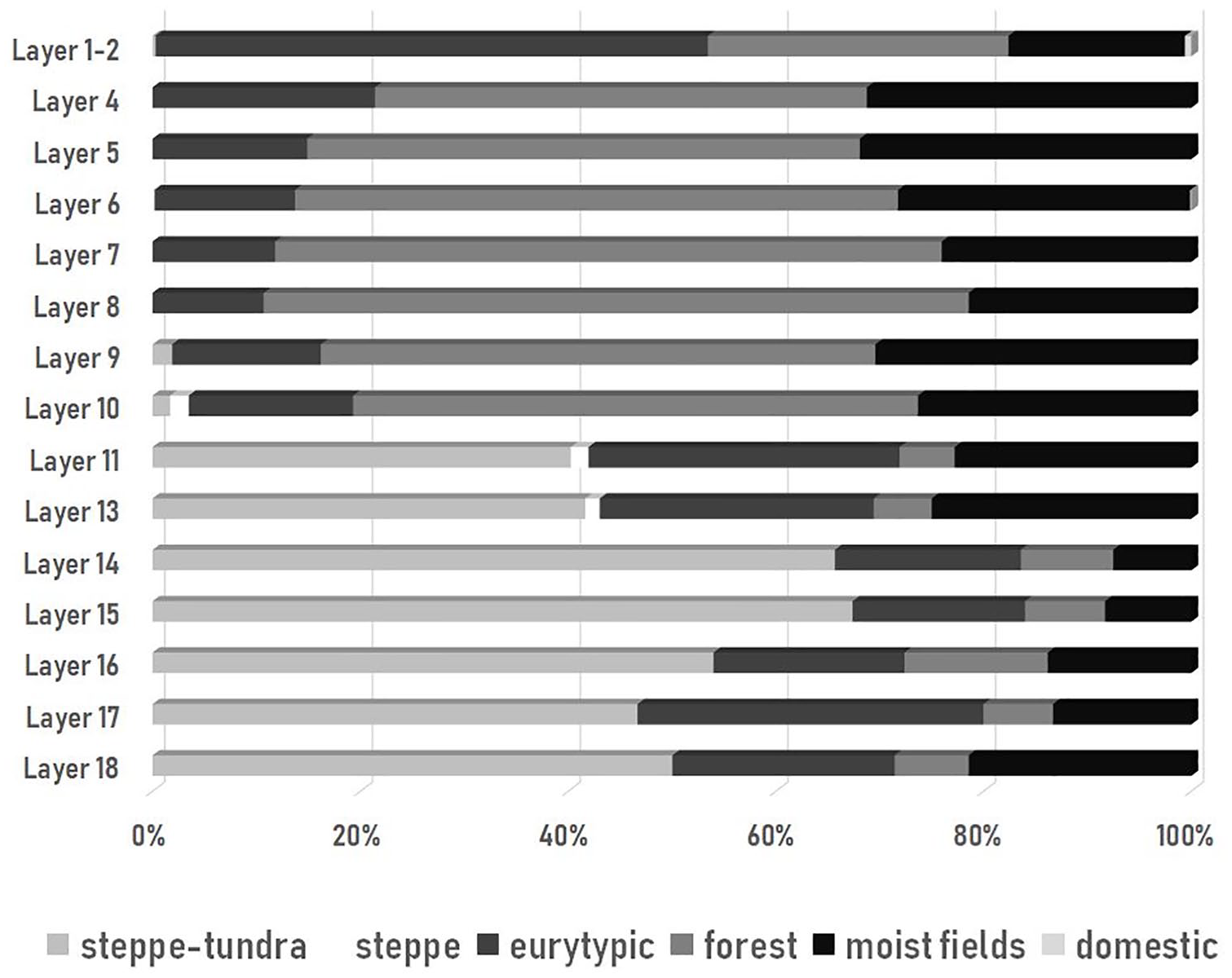
Percentage representation of habitat categories for rodent species in successive layers of the Żarska Cave according to MNI counts: steppe-tundra (Dicrostonyx gulielmi, Lemmus lemmus, Microtus gregalis, Ochotona sp.), steppe/open fields (Cricetus cricetus, Cricetulus migratorius, Apodemus microps, Lepus sp.), eurytopic (Microtus arvalis/agrestis), forest (Clethrionomys sp., Apodemus sylvaticus/flavicollis, Sicista betulina, Sciurus vulgaris, Gliridae), moist fields (Microtus subterraneus, M. oeconomus, Arvicola terrestris, Micromys minutus), and domestic species (Mus musculus, Rattus rattus).
In layer 10, forest species such as Apodemus and Clethrionomys increase significantly (up to 54%) as do species from open moist fields, such as Microtus subterraneus (to 26%). The proportions of rodent species in layers 7–9 indicate further climatic warming: in these layers there is an increase in the frequency of forest species such as Apodemus sylvaticus/flavicollis and Sciurus vulgaris (up to 53–68%) and a decrease of eurytopic species such as Microtus arvalis/agrestis to 11–14%. In these layers, steppe inhabitants (Cricetus and Cricetulus) disappear. The largest share of species preferring forests was recorded in layers 7 and 8 (64–68%).
A gradual change in rodent composition is observed in layers 4–6. The forest inhabitants such as Microtus subterraneus or Apodemus sylvaticus/flavicollis decrease in percentage (58–47%) while the species preferring moist fields or banks of water reservoirs such as Clethrionomys glareolus increase in frequency to 31%, similar to the trend in eurytopic rodent species (up to 21%). In layer 6, the first appearance occurs of synanthropic species associated with humans, Mus musculus and Rattus rattus. In layers 1 and 2 there is a significant reduction in the number of the forest dwellers (goes down to 28%) and is associated with moist fields (to 16%), and an increase is seen in the number of species of open areas (to 52%) such as Microtus arvalis/agrestis.
Chiropterofauna from the Żarska Cave occur throughout the stratigraphic sequence, exhibiting low abundance of remains in the Pleistocene sediments, followed by an increasing trend in the Holocene, although fluctuating with some episodes of decline. In layers 12–18 the number of chiroptera remains is low (Tab. 6 – Appendix, available online). The situation changes in layers 10 and 11, where an increase is observed in the number of taxa (represented by Barbastella barbastellus, Myotis daubentonii, and Plecotus auritus) as well as the quantity of remains.
A significant increase in the number of bat species occurs in layers 8 and 9, where such taxa as Eptesicus serotinus and Myotis emarginatus indicate a more thermophilous assemblage than those identified in the lower layers. In the middle-Holocene Optimum (layer 7), the trend continues of rising number of temperate forest species (Myotis bechsteinii, Plecotus auritus, Barbastella barbastellus). Surprisingly in layer 6, where a distinct record of Eneolithic human occupation exists, chiroptera faunal elements are well represented. In that layer, the remains of Myotis bechsteinii, Myotis daubentonii, Plecotus auritus, and Barbastella barbastellus are prevalent. In the most recent layers, there is a slight decrease of remains in layer 5 and a temporary disappearance in layers 1–3, when the record of human occupation is well documented.
Large herbivores (only Artiodactyls; Perissodactyls do not occur in this sequence) are represented by both domestic and wild animals. The numbers of remains vary in the individual layers, and while in the upper layers herbivore remains are rather numerous, in the lower ones only single identifiable bones and teeth are present (Table 7 – Appendix, available online).
Within layers 12–18, only layers 13 and 17 contained single identified bones of aurochs/bison, including second phalange (layer 17) and whole mandible (layer 13). In the younger Pleistocene layers (10 and 11) dated to the Allerød phase, only single bones of roe deer were found. The same situation is repeated in layers 8 and 9, as well as layer 7, where besides roe deer, additionally a single bone of red deer was found. The most abundant material of large herbivores was discovered in layer 6, where numerous Eneolithic ceramics and lithic artifacts were found. In the bone assemblage of this layer (N = 206), bones and teeth of goat or sheep dominate, constituting more than half of the material, and nearly 90% of the identified bones (N = 116). The remains belong to a minimum of seven individuals, most of them young subadult specimens. Other taxa such as cattle (N = 5), pig (N = 1), red deer (N = 1), and roe deer (N = 7) are represented only by single bones.
In the uppermost layers, 1 and 2, dated from the Roman Period up to the Post-Medieval period, domesticated animals prevail. Among them, pig remains are the most numerous. Other specimens such as cattle and goat/sheep were represented by only single bones. Because domestic animal remains were discovered at the whole surface of the trench and without any relation to human burials dated to the Roman Period, it can be presumed that they are predominantly associated with other periods of human occupations, that is, with early Medieval and post-Medieval occupation.
In the sediments of the Żarska Cave, at least 11 carnivore taxa were found from four taxonomic families (Mustelidae, Felidae, Canidae, and Ursidae). The carnivore material was represented by disarticulated skeletal elements, with the majority being parts of cranial bones, isolated teeth, long bones, metacarpals, and metatarsals (Table 8 – Appendix, available online).
Two faunal sub-assemblages were identified in the carnivore material. The older one, found in layers 12–18, included scarce material of two ursid taxa (cave bear, Ursus ex gr. spelaeus, and brown bear, Ursus arctos), the postcranial material of pine marten, Martes martes (all from layer 13, and all probably from one individual), and single findings of two smallest mustelids, Mustela cf. erminea and Mustela nivalis. The young ursids in layers 15–18 and 13 indicate that the cave was used by bears as a place for hibernating and raising cubs.
The younger sub-assemblage in layers 1–11 contains a relatively rich assortment of nine species, dominated by some mustelids, Meles meles, Martes martes, and a small wildcat (Felis silvestris). Noteworthy is the presence of domestic Carnivora such as domestic cat Felis catus in layers 1–2 and dog Canis lupus familiaris in layers 1–2, and 6. Metrically and morphologically, the animals from these sediments correspond to the modern European forms.
Discussion
The thick and undisturbed Holocene sediments discovered in the Żarska Cave are unusual for the Kraków-Częstochowa Upland. Until recently, the Nad Mosurem Starym Cave in Ojców National Park was thought to contain the greatest thickness of Holocene sediments in the Polish Upland (Nadachowski et al., 1989). However, the sediments in this cave span only the upper part of the Holocene, from the Epi-Atlantic until recent times. The Żarska Cave provides a record of a much longer time range and a greater richness of paleobotanical and paleontological material; thus, it is the more complete example of late-Pleistocene–Holocene cave deposits in the Upland. In fact, it provides a unique record of paleoenvironmental changes for the whole north-Carpathians region. Its organogenic materials allow for a detailed tracking of paleoenvironmental changes.
The summarized results of the interdisciplinary studies are shown in Figure 8, which illustrates the changes that took place in the nearby cave area from the late-Glacial period, along with a full record of changes covering the entire Holocene, with well-defined early and middle phases.

The changes in biotic and abiotic environment and human cultures around the Żarska Cave based on the results of multidisciplinary studies of the cave section.
Among recently excavated caves, the most similar from a lithostratigraphic point of view are the Shelter in Smoleń III (Sudoł and Krajcarz, 2014), the Cave above the Słupska Gate (Szymanek et al., 2016), and the Perspektywiczna Cave (Krajcarz et al., 2016). The closest analogy may be found in Shelter in Smoleń III, situated ca. 40 km to the north. However, the sedimentary fills in these cave sites are much poorer with regard to both the thickness of the sediments, and especially the size and variety of the obtained fossil material, such as the mammal remains represented only by isolated teeth and fragments of bones at the Cave above the Słupska Gate (Szymanek et al., 2016).
The geographic areas near the Żarska Cave are still poorly known in terms of the history of late-Glacial and postglacial vegetational changes. This is an issue relevant to the entire loess upland zone of southern Poland (Ralska-Jasiewiczowa, 1989; Ralska-Jasiewiczowa and Latałowa, 1996), because of a low number of easily detectable water reservoirs suitable for paleoecological and especially palynological analysis (Żurek, 1987). The closest pollen sequences are available at a distance of 20–45 km from the Żarska microregion, in the south-eastern Silesian-Kraków Upland and Częstochowa Upland, but they are located in different geobotanical regions and contain a sequence covering late-Glacial (Okupny et al., 2016), late-Glacial and only early Holocene (Szczepanek and Stachowicz-Rybka, 2004), late-Glacial and parts of the Holocene (Latałowa, 1976; Latałowa and Nalepka, 1987; Żurek et al., 2011), and the Holocene (Madeyska and Obidowicz, 2001).
Early late Glacial (layers 12–18)
The age of layer 18 is unknown, but may be much earlier than late-Glacial, as evidenced by calcite crusts around the limestone clasts, typical for Lower Weichselian (MIS 5a-d) deposits in caves of the region (Madeyska, 1981, 1988). The remains of fauna and flora from this layer are of limited usefulness for ecological reconstructions and biostratigraphic dating.
The overlying part of the section (layers 12–17) consists of colluvial sediments. The source material for some layers of this series was a typical cave deposit, that is, humiferous gray-brown silts with high amount of limestone clasts, while the source materials for other layers were typical open-air deposits, including loess and paleosols. The sediments of this series were redeposited into the cave or reworked within the cave and may contain mixed materials of different original position and chronology. Radiocarbon dating is not useful in determining the chronology of the series, because of poor preservation of collagen in bones (Table 1), but the taxonomic composition of fossil flora and fauna strongly supports the late-Pleistocene age of the material. The age of the redepositional processes may be much younger. The presence of unstable density stratification in layers 12 and 13 indicates a temporal water saturation of sediments. This phenomenon may be connected with melting of permafrost during a period of climatic warming. The general stratigraphic position indicates a link between the water saturation and interstadials of late-Glacial time or the beginning of the Holocene.
Cold climatic conditions that produced open habitats are well recorded in the faunal assemblage, especially the data from mollusks and rodents. The molluskan taxa present in the Pleistocene layers (12–18) are often found in loess formations (e.g. Vallonia tenuilabris), as well as in sediments of the cool periods of the late-Glacial. The malacoenosis indicates the occurrence of open habitats with patches of open-canopy forests and bushes (Arianta arbustorum and Semilimax kotulae), developing in a cold continental climate, corresponding to the older part of the late-Glacial. Faunas of similar composition and age were noted in several sites distributed all over Central Europe (Alexandrowicz, 2014; Alexandrowicz and Dmytruk, 2007; Alexandrowicz et al., 2014a; Ložek, 2001; Moine, 2008; Sümegi, 2005).
This sequence of layers (13–18) contains relatively abundant remains of willow ptarmigan, an Arctic–boreal bird commonly found in the European Pleistocene (Tyrberg, 1998). The predominance of this species suggests a semi-open environment of marshy heathlands, shrub tundra, and areas with a light growth of birch and pine, possibly located next to coniferous forest.
Cold climatic conditions are well recorded in the rodent assemblage. The deepest Pleistocene layers (14–18) are characterized by a predominance of tundra species (ca. 47–67% of the total fauna). The most severe environmental conditions occurred during the formation of layers 14 and 15, in which the proportion of Microtus gregalis and two lemming species is the largest (65–67%). The upper layers 11 and 13 show a slight change in the proportion of species and the start of climatic warming. The large diversity of rodent fauna in these layers reflects the mosaic environment around the Żarska Cave. Harsh climatic conditions are also confirmed for that sequence by the exiguous presence of a faunal assemblage of chiroptera (Table 2 Appendix, available online), composed of species tolerating cold hibernacula having average temperatures between 2°C and 5°C. Assuming that the cave microclimate reflects the external mean annual temperature, this suggests that the outside air paleotemperatures were much lower than recent temperatures in the region, which are ~6–7°C. Generally, in layers 12–18 only single large animal remains were discovered. Relatively numerous are the remains of bears, which used the cave as a place for hibernating and to raise cubs. Bison/aurochs remains in layer 13 may indicate warmer and more humid climate during deposition of that layer. Noteworthy is the presence of a large pine marten and two smallest must lids, Mustela cf. erminea and Mustela nivalis, whose small sizes were characteristic of late-Pleistocene cold phases.
Allerød (layers 10 and 11)
Layers 10 and 11 are colluvial sediments, redeposited in ways similar to the origin of the lower series. An abrupt increase of humus content in layer 11 (Figure 3) indicates an important change in climate and vegetation in relation to the lower layers, toward warmer and moister conditions. This is also confirmed by the relatively high amount of smoothed limestone clasts, noted already in layer 12. Layer 10 shows an increase of angular clasts, which may indicate climatic cooling.
Palynology-based histories of vegetation in the uplands of southern Poland have identified open forests with boreal trees such as Pinus sylvestris and Betula along with Larix and Picea during the warmer intervals of the late-Glacial, especially in the Allerød (Latałowa, 1976; Latałowa and Nalepka, 1987). Charcoal assemblages from the Żarska Cave corresponding to layer 11 and probably layer 10 can be related to this phase, which most likely represents pine forest and/or birch-pine woodland, since the charcoals are characterized by a dominance of Pinus sylvestris with a high share of Betula sp. Among charcoals, a taxon referable to two species (Picea abies or Larix decidua) was also found, but unfortunately the two species cannot be easily distinguished based on wood anatomy (i.e. Anagnost et al., 1994; Schweingruber, 1990b), especially that some features, such as a transition from earlywood to latewood or the appearance of biseriate bordered pits, cannot always be used as distinctive ones. Both Larix and Picea appeared in pollen sequences from Wolbrom dated to the late-Glacial, although Larix reached a higher share (Latałowa, 1976, 2003).
The malacoenosis from layers 10 to 11 is characteristic of the younger part of the late Glacial, and relatively numerous occurrences of the shadow-loving snails (i.e. Arianta arbustorum) suggest its connection with the Allerød interphase. Similar assemblages were described from several other sites (Alexandrowicz, 1997; Ložek, 1991).
Remains of the identified birds (e.g. Capercaillie) and mollusk from layers 10 to 11 indicate the occurrence of a wooded area in the cave vicinity. It is noteworthy that no single bone of willow grouse has been identified in layers 10 and 11 (nor in upper layers 1–9), despite their abundance in the deeper layers. This change corresponds well with the altered distribution of the bird because of climatic warming (see Lagerholm et al., 2016).
The development of open forests with boreal trees is well reflected in the rodent assemblage from layers 10 and 11, in which the proportion of tundra species drops dramatically to 2%, in favor of forest species such as Apodemus and Clethrionomys (up to 54%) and species preferring open moist fields (up to 26%). These changes indicate rapid warming of the climate and the emergence of boreal forests. Such a global warming and forest expansion is also indicated in layers 10 and 11 by the increasing number of chiroptera remains. However, it is also likely that layer 10 represents a cold phase between the Allerød and the early Holocene, given the detection of Barbastella barbastellus as Plecotus sp., a species capable of tolerating and even preferring low temperatures during hibernation (Lesiński, 1986; Pocora and Pocora, 2011). Its continuous persistence together with that of Myotis bechsteinii suggests the presence of an old-growth forest (Dietz and Pir, 2009; Russo et al., 2004) with old rotting trees, having a higher proportion of open woodland areas in the Pleistocene than in the Holocene, although the latter species also shows the presence of small glades. Placid regime watercourses (i.e. not torrential) bordered these surroundings, representing the preferred hunting environment for the bats M. daubentonii and M. dasycneme (Britton et al., 1997; Parson and Jones, 2006).
Although herbivore remains are not as useful for reconstructing paleoenvironmental changes in comparison to malacofauna or rodents, some general tendencies are apparent. The presence of single remains of medium mammal specimens in layer 11, such as roe deer, is notable. This species prefers open areas typical of forest margins. One bone from layer 10 was radiocarbon dated to 11,500 ± 60 BP, confirming the Allerød age (Table 1), but the presence of cold adapted specimens in that layer may be the result of partially mixing with sediment from the Dryas stadial.
Early Holocene (layers 8 and 9)
Layers 8 and 9 exhibit geological characteristics of deposition during the warm and humid climate. This is confirmed by the chemical weathering of limestone clasts recorded as smoothed or nearly rounded pebbles, and high carbonate and humus content in sediments (Figure 3). Both layers bear erosional structures (Figure 2), indicating a temporal inflow of water. The sediments were redeposited by colluvial activity.
At the beginning of the Holocene, an expansion of trees with greater ecological requirements such as Ulmus, Quercus, Tilia, Fraxinus, Alnus, and Corylus has been observed in various regions of southern Poland, based on pollen data (Latałowa, 1976; Latałowa and Nalepka, 1987; Madeyska and Obidowicz, 2001; Ralska-Jasiewiczowa et al., 2004; Szczepanek and Stachowicz-Rybka, 2004a). Similarly, in the Żarska Cave the beginning of the Holocene can be seen in the charcoal assemblage with the first appearance of mesophilous taxa such as Corylus avellana and Ulmus sp.
The malacofauna from layers 8 and 9 demonstrates the features of Ruderatus-fauna, characteristic of the early Holocene (e.g. Alexandrowicz, 2004, 2013; Alexandrowicz et al., 2014b, 2016; Dehm, 1967; Gedda, 2001; Horáčková et al., 2015; Juřičková et al., 2014; Limondin-Lozouet, 2011; Limondin-Lozouet and Preece, 2014; Meyrick, 2002; Žak et al., 2002). The faunal characteristics indicate a predominance of shaded habitats overgrown by compact forests, and in a relatively cold climate with continental features. Since warm climatic conditions are expected during the early Holocene, it is possible that some redeposition took place. This means that some older material has been incorporated into layers 8 and 9 as an effect of erosion of older layers and colluvial deposition. The presence of forests is also confirmed by bird species in the layers (e.g. black woodpecker and tawny owl).
The proportions of rodent species in layers 8 and 9 indicate further climatic warming, based on the increased frequency of forest species (up to 53–64%) reflecting the development of large forest complexes and the decrease of eurytopic species. Abundant bat remains belonging to numerous species, such as Myotis emarginatus and Eptesicus serotinus (Lanza, 2012), which are more thermophilous than those in the lower layers, may be related to the temperature increase that occurred in the pre-Boreal period.
Middle Holocene (layers 6 and 7)
Layer 7 is the most distinct one in the section because of its bright rusty color. This layer was deposited during the climatic optimum, evidenced by a high clay content and rounded limestone clasts (Figure 3), typical features of sediments deposited during warm climatic phases (Krajcarz and Madeyska, 2010; Madeyska, 1981). Layer 6 is an anthropogenic sediment, composed mostly of ash and redeposited material of lower layers. Starting from layer 8 upward, the sediments contain low amounts of humus (Figure 3), which may be connected with reduced input of organic matter from the external environment such as during partial closing of the cave entrance, or to more advanced decay of organic remains because of favorable climatic and hydrogeological conditions.
The oldest artifacts in sediments of the Żarska Cave are dated to the Eneolithic (Wojenka et al., 2016). The most numerous artifacts with determinable ages are dated to the Baden culture. Most materials can only be imprecisely assigned to the Eneolithic; several ceramic vessel fragments are linked to the turn of the Eneolithic to the Bronze Age, probably belonging to the Strzyżów culture. The age of layer 6 is also confirmed by the radiocarbon dating of bones and charcoals: 5330 ± 40, 5010 ± 40, and 4370 ± 35 BP (Table 1).
The first decrease of P. sylvestris was detected in layer 7, which likely represents the Atlantic phase, while layer 6 might correspond to the final stages of this phase. During the Atlantic, southern Poland saw the greatest development of deciduous woodland in the Holocene Climatic Optimum (Ralska-Jasiewiczowa et al., 2004a). A dominance of deciduous trees and shrubs is also evident in the Żarska Cave based on the anthracological analysis, but locally a maple forest could have predominated on the slopes near the cave. Nowadays, this kind of forest (Phyllitido-Aceretum) is typical for slopes surrounding valleys of the Ojców region, the nearest to the cave (Maciejowski et al., 2013), and in general also the Kraków-Częstochowa Upland (Matuszkiewicz, 2005; Medwecka-Kornaś et al., 1972). Usually this forest is rather dense, but in the Ojców region such a maple forest can also have a broken canopy because of a high proportion of Corylus avellana and Tilia platyphyllos (Bodziarczyk, 2002). However, the presence of narrow rings in the charcoals of Fraxinus excelsior might suggest that these trees developed under unfavorable habitat conditions, for instance, in dense woodlands or on shallow soils. Although Acer trees were probably important constituents of deciduous woodlands in the Holocene, they are underrepresented in pollen diagrams (Noryśkiewicz et al., 2004), because they are insect pollinated. Charcoal fragments of Ulmus sp., F. excelsior, and Acer sp. were usually found in the assemblages from Neolithic archeological sites from southern Poland (Burchard and Lityńska-Zając, 2002; Lityńska-Zając, 1997; Lityńska-Zając et al., 2017; Moskal-del Hoyo, 2016; Moskal-del Hoyo et al., 2017b, 2018; Wasylikowa et al., 1992). However, these taxa did not reach such high proportions in archeological charcoal assemblages and therefore, a unique charcoal Acer-Fraxinus-Ulmus-Corylus-dominated assemblage might be interpreted as a past forest community similar to a present-day Phyllitido-Aceretum association. It is likely that in the Atlantic phase, this kind of deciduous forest could have been widespread in habitats of wet and shady slopes and might represent a relict forest community that developed before a spread of Fagus sylvatica (Leuschner and Ellenberg, 2017), and which now persisted only in a few locations on steep limestone slopes and rock shelves in the Kraków-Częstochowa Upland among predominated beech forests (Baran and Bodziarczyk, 2019; Bodziarczyk, 2002, Figure 1).
Conversely, a high frequency of hazel and oak in layer 6 may indicate the presence of open woodland dominated by Quercus sp. with C. avellana in the understory, but these forest communities likely would have developed in areas above the cave covered by loess. Relatively frequent charcoal assemblages are found in the nearby archeological sites dated to the Atlantic and the early Sub-Boreal phases from southern Poland. The evidence from these sites indicates the main forest communities growing on the loess-mantled areas were dominated by Quercus sp. and Pinus sylvestris (Burchard and Lityńska-Zając, 2002; Lityńska-Zając, 1997; Lityńska-Zając et al., 2017; Moskal-del Hoyo et al., 2017b, 2018).
The fauna in layers 6 and 7 also indicates the presence of compact mixed and deciduous forests, as well as a warm and relatively humid climate with marked effect of Atlantic air masses. The malacofauna assemblage from these layers is similar to the ‘Perspectivus-fauna’, considered to be an indicator malacocoenosis for the middle Holocene (Alexandrowicz, 2004; Alexandrowicz et al., 2014b, 2016; Dehm, 1987; Horáčková et al., 2015; Juřičková et al., 2014; Limondin-Lozouet, 2011; Limondin-Lozouet and Preece, 2014; Meyrick, 2002; Žak et al., 2002). Most of the identified bird species in layers 6 and 7 would have occupied denser or lighter forest environments, although in these layers bones are present of a few species occupying open environments. A gradual change in the rodent composition is observed in layers 6 and 7. Forest inhabitants increase to 68% (the maximum in the Żarska Cave), followed by a gradual decrease of such species to 58%. This process continued in the younger layers.
Relatively rich mammal remains were discovered in layer 6, where signs of human occupation provide important information about Eneolithic diets and husbandry. In this layer, domesticated animals from the tribe Caprini played the most important role, which can be related to the specific environmental conditions of the limestone Uplands, favorable for grazing taxa such as goat/sheep. Traces of Eneolithic settlements discovered in this cave are considered to be elements of seasonal pastoral camps. The hypothesis is that humans and bats alternated seasonally in their occupations of the cave: people used the cave during summers when caprine flocks were grazing in the surrounding area, and bats used the cave to hibernate in winters, supported by numerous remains of bats.
Layers 6 and 7 contain abundant remains of carnivores involving both wild (badger, red fox, pine marten, and wildcat) and domestic species (dog). Most of them are recognized as eurytopic species. The only exception is pine marten, a species characteristically associated with forested habitats. This species prefers old-growth deciduous forest, but also inhabits mixed, coniferous, and taiga-type forests (Anderson, 1970; Hernández-Fernández, 2001; Marciszak, 2012; Overskaug et al., 1994). The other two highly adaptable species which can live in a wide range of habitats, Meles meles and Felis silvestris, are usually forest-dwellers in Central and Eastern Europe (Lozano, 2010; Proulx et al., 2016; Yamaguchi et al., 2015).
Late Holocene (layers 1–5)
The uppermost layers differ from the lower part of the section by having a dark color and high content of organic matter (Figure 3). These sediments resemble topsoil. The large amount of organic matter may be explained in different ways: (1) a change of vegetation around the cave, (2) a higher rate of organic matter input to the cave, or (3) a less intense decaying process because of the sediment’s young age or to unfavorable climatic conditions.
In the local zone recorded in layers 4 and 5, the percentage of the Pinus sylvestris charcoals increased, followed by Quercus sp. and Corylus avellana. It is likely that this zone corresponds to mixed forest (pine–oak forest), perhaps related with loess habitats. The percentage of Fagus sylvatica and Abies alba began to increase in this local zone. Also, a presence of Carpinus betulus was noted, but only in layer 4. All of these tree species are late migrators that became more important constituents of forest communities during the Sub-Boreal phase (Ralska-Jasiewiczowa et al., 2004b).
The percentage of Fagus sylvatica is the highest in the last local zone (layers 1 and 2). Although this zone is characterized by a very low number of charcoal fragments, a present-day dominance of various beech forests around the Żarska Cave (Maciejowski et al., 2013) might suggest that during these periods the beginning of a transformation into the beech forests took place. Also, beech is typically found in pollen diagrams from the Sub-Atlantic phase (e.g. Latałowa and Nalepka, 1987). Layers 1 and 2 probably can be linked with Medieval or post-Medieval chronology since layer 3 contained a hearth dated to the Middle Ages.
Animal remains discovered in late-Holocene layers are dispersed very unevenly; some layers have no representatives of some groups or entire orders. Moreover, these uppermost layers have been partly destroyed because of post-medieval human activities, and any detailed stratigraphic studies are limited.
The malacofauna is poorly represented in layers 4 and 5. The malacoenosis from layers 1–3 is characteristic of dense forest and a climate cooler than what was present when the underlying layers were deposited. This fauna could be linked to the late-Holocene, a stratigraphic interpretation suggested by both the features of the molluskan assemblage (Alexandrowicz, 2004) as well as the archeological data and radiocarbon dating (1755 ± 30 and 1655 ± 30) (layer 2). The presence of a freshwater bivalve Unio pictorum in layers 3 and 4 probably indicates the use of the cave by humans.
The number of bat remains decreased in layer 5, and even a temporary disappearance is marked in layer 3. This can be explained by the human exploitation of the cave, possibly much more intense than during Eneolithic, when the cave was probably used only as a seasonal shepherds’ shelter.
In layers 4 and 5, rodent species preferring moist fields or banks of water reservoirs increase in frequency to 31%, similar to eurytopic rodent species (up to 21%). In turn, data from the youngest layers 1 and 2 show a rapid and noticeable change in the proportion of rodent species and a change in the quality of the environment. Some data from layers 1 and 2 are in contrast to anthracological and malacological data indicating developing forests, in that they show a significant reduction in the number of the forest dwellers (up to 28%) and taxa associated with moist fields (to 16%), and an increase in the number of species of open areas (to 52%). These trends suggest deforestation and drying of the Żarska Cave surroundings. The data might reflect a possible existence of mosaic-like vegetation, with a development of denser forests (beech woods) on the slopes surrounding the cave and possible deforestation of an upper zone located above the cave where oak–pine forests could have been initially developed on loess substratum and could have been partially changed into fields by medieval farmers. The remains of oak and pine were still important components of anthracological assemblages from layers 1 to 5 (Figure 4), while evidence of an open landscape was also indicated by the presence of charcoals of Juniperus communis, especially in layer 3 with the remains of a hearth. It should be emphasized that rodent remains deposited in the cave would have been mainly residues of pellets of undigested material egested by raptors, notably owls, whose hunting territory can cover open fields and farmlands located in the cave vicinity (O’Connor, 2000).
Our analysis of mammalian bones also indicates the presence of forests near the cave, since remains of badger, pine marten, and wildcat appeared in faunal assemblages from the youngest layers. However, the most characteristic feature of these assemblages is a predominance of remains of pigs, which demonstrates that the cave was used extensively by people, as the abundant pig remains were associated with numerous Medieval (mid-13th–mid-14th century) and post-Medieval period artifacts. These data provide information about the subsistence behavior of humans living near the cave in these periods. This may be described as focusing on quick returns with minimal use of resources. Such conclusion is based on the fact that pigs as typical omnivores eat a very wide range of foods and are highly efficient producers of energy (Harris, 1985; Marciniak, 2005). However, the dominance of pig in this assemblage differs from the data obtained in Medieval and post-Medieval sites of Kraków, where cattle prevailed (Wilczyński et al., 2015; Wojtal et al., 2010). This may indicate different environmental conditions such as the presence of forests with beech and oak near the Żarska Cave, which would have been favorable to the pig-keeping, supported by paleobotanical analysis. In addition, in layers 1–3, abundant remains of domesticated birds appeared, which is consistent with the use of the cave more intensively by people. In addition, many chicken bones and a few remains of goose bear mammal tooth marks were recovered. A few of the identified species are more or less synanthropic and can be observed today at human settlements (common kestrel, magpie, and carrion crow).
Conclusion
The Żarska Cave provides an extraordinary sequence of Upper Pleistocene–Holocene cave deposits, containing both paleobotanical and paleontological data. Because of its richness and diversity, it contains one of the fullest records of paleoenvironmental changes known from southern Poland, and generally for the north-Carpathian region. The well-preserved cave deposits are a continuous archive of temporal changes in geological, paleobotanical, and faunal data, which is especially relevant since cave sediments containing paleontological evidence are commonly destroyed by human activities in this region.
The data allow a clear distinction between the fauna and flora of late-Glacial cold and dry environments. Especially relevant assemblages were dated to the Allerød phase, when shade-loving mollusks and rodents of forests appeared. The beginning of the Holocene was clearly identified with an increase of shaded habitats in forests, which developed in a still relatively cold climate with continental features, with a significant annual variation in temperature, and with the first appearance of mesophilous deciduous trees. The middle-Holocene fauna and flora developed under a warm and relatively humid climate, as indicated by the presence of mixed and deciduous forests. Within the middle-Holocene sequence, layer 6 is unique because of the use of the cave by Eneolithic people as a shelter during the summer grazing of sheep and goats, giving insight into animal husbandry practices. This layer also revealed an exceptional evidence of maple forests that developed before the formation of beech forests. The floral and faunal materials in the late-Holocene layers offer evidence of cooler conditions relative to the preceding middle Holocene. During the late-Holocene, the cave was more intensively used by people, which is associated with important changes in local fauna (i.e. disappearance of bats) and vegetational cover. Local deforestation took place, likely in areas of rich soils developed on loess substratum, resulting in the formation of the present-day dominant forest communities with beech, fir, and hornbeam. In the Middle Ages, a selected use of the cave’s surrounding landscape for pig breeding is suggested.
The presented sequence of the Żarska Cave can be regarded as one of the local records of past environment from the northern Carpathian zone. As a result, together with other sites it can be used for the wider paleoenvironmental reconstructions, being of regional or continent scale. Moreover, it is also one of the most complete sequences, covering the entire Holocene period. The long record of multiple proxies (i.e.: fossil mollusks, fish, birds, rodents, chiropterans, large mammals, and charcoals, all within the stratigraphic and chronological outline) makes it one of the most important sites in the northern part of Europe.
Supplemental Material
Zarska_Table_1_Appendix_-_antraco_REVISION – Supplemental material for Late Glacial and Holocene paleoecology and paleoenvironmental changes in the northern Carpathians foreland: The Żarska Cave (southern Poland) case study
Supplemental material, Zarska_Table_1_Appendix_-_antraco_REVISION for Late Glacial and Holocene paleoecology and paleoenvironmental changes in the northern Carpathians foreland: The Żarska Cave (southern Poland) case study by Jarosław Wilczyński, Maciej T Krajcarz, Magdalena Moskal-del Hoyo, Witold Paweł Alexandrowicz, Barbara Miękina, Andrea Pereswiet-Soltan, Krzysztof Wertz, Grzegorz Lipecki, Adrian Marciszak, Lembi Lõugas, Michał Gradziński, Anita Szczepanek, Albert Zastawny and Michał Wojenka in The Holocene
Supplemental Material
Zarska_Table_2_Appendix_-_malaco_-_added_REVISION – Supplemental material for Late Glacial and Holocene paleoecology and paleoenvironmental changes in the northern Carpathians foreland: The Żarska Cave (southern Poland) case study
Supplemental material, Zarska_Table_2_Appendix_-_malaco_-_added_REVISION for Late Glacial and Holocene paleoecology and paleoenvironmental changes in the northern Carpathians foreland: The Żarska Cave (southern Poland) case study by Jarosław Wilczyński, Maciej T Krajcarz, Magdalena Moskal-del Hoyo, Witold Paweł Alexandrowicz, Barbara Miękina, Andrea Pereswiet-Soltan, Krzysztof Wertz, Grzegorz Lipecki, Adrian Marciszak, Lembi Lõugas, Michał Gradziński, Anita Szczepanek, Albert Zastawny and Michał Wojenka in The Holocene
Supplemental Material
Zarska_Table_3_Appendix_-_fish_REVISION – Supplemental material for Late Glacial and Holocene paleoecology and paleoenvironmental changes in the northern Carpathians foreland: The Żarska Cave (southern Poland) case study
Supplemental material, Zarska_Table_3_Appendix_-_fish_REVISION for Late Glacial and Holocene paleoecology and paleoenvironmental changes in the northern Carpathians foreland: The Żarska Cave (southern Poland) case study by Jarosław Wilczyński, Maciej T Krajcarz, Magdalena Moskal-del Hoyo, Witold Paweł Alexandrowicz, Barbara Miękina, Andrea Pereswiet-Soltan, Krzysztof Wertz, Grzegorz Lipecki, Adrian Marciszak, Lembi Lõugas, Michał Gradziński, Anita Szczepanek, Albert Zastawny and Michał Wojenka in The Holocene
Supplemental Material
Zarska_Table_4_Appendix_-_aves_REVISION – Supplemental material for Late Glacial and Holocene paleoecology and paleoenvironmental changes in the northern Carpathians foreland: The Żarska Cave (southern Poland) case study
Supplemental material, Zarska_Table_4_Appendix_-_aves_REVISION for Late Glacial and Holocene paleoecology and paleoenvironmental changes in the northern Carpathians foreland: The Żarska Cave (southern Poland) case study by Jarosław Wilczyński, Maciej T Krajcarz, Magdalena Moskal-del Hoyo, Witold Paweł Alexandrowicz, Barbara Miękina, Andrea Pereswiet-Soltan, Krzysztof Wertz, Grzegorz Lipecki, Adrian Marciszak, Lembi Lõugas, Michał Gradziński, Anita Szczepanek, Albert Zastawny and Michał Wojenka in The Holocene
Supplemental Material
Zarska_Table_5_Appendix_-_rodents_REVISION – Supplemental material for Late Glacial and Holocene paleoecology and paleoenvironmental changes in the northern Carpathians foreland: The Żarska Cave (southern Poland) case study
Supplemental material, Zarska_Table_5_Appendix_-_rodents_REVISION for Late Glacial and Holocene paleoecology and paleoenvironmental changes in the northern Carpathians foreland: The Żarska Cave (southern Poland) case study by Jarosław Wilczyński, Maciej T Krajcarz, Magdalena Moskal-del Hoyo, Witold Paweł Alexandrowicz, Barbara Miękina, Andrea Pereswiet-Soltan, Krzysztof Wertz, Grzegorz Lipecki, Adrian Marciszak, Lembi Lõugas, Michał Gradziński, Anita Szczepanek, Albert Zastawny and Michał Wojenka in The Holocene
Supplemental Material
Zarska_Table_6_Appendix_-_bats_REVISION – Supplemental material for Late Glacial and Holocene paleoecology and paleoenvironmental changes in the northern Carpathians foreland: The Żarska Cave (southern Poland) case study
Supplemental material, Zarska_Table_6_Appendix_-_bats_REVISION for Late Glacial and Holocene paleoecology and paleoenvironmental changes in the northern Carpathians foreland: The Żarska Cave (southern Poland) case study by Jarosław Wilczyński, Maciej T Krajcarz, Magdalena Moskal-del Hoyo, Witold Paweł Alexandrowicz, Barbara Miękina, Andrea Pereswiet-Soltan, Krzysztof Wertz, Grzegorz Lipecki, Adrian Marciszak, Lembi Lõugas, Michał Gradziński, Anita Szczepanek, Albert Zastawny and Michał Wojenka in The Holocene
Supplemental Material
Zarska_Table_7_Appendix_-_artio_REVISION – Supplemental material for Late Glacial and Holocene paleoecology and paleoenvironmental changes in the northern Carpathians foreland: The Żarska Cave (southern Poland) case study
Supplemental material, Zarska_Table_7_Appendix_-_artio_REVISION for Late Glacial and Holocene paleoecology and paleoenvironmental changes in the northern Carpathians foreland: The Żarska Cave (southern Poland) case study by Jarosław Wilczyński, Maciej T Krajcarz, Magdalena Moskal-del Hoyo, Witold Paweł Alexandrowicz, Barbara Miękina, Andrea Pereswiet-Soltan, Krzysztof Wertz, Grzegorz Lipecki, Adrian Marciszak, Lembi Lõugas, Michał Gradziński, Anita Szczepanek, Albert Zastawny and Michał Wojenka in The Holocene
Supplemental Material
Zarska_Table_8_Appendix_-_carniv_REVISION – Supplemental material for Late Glacial and Holocene paleoecology and paleoenvironmental changes in the northern Carpathians foreland: The Żarska Cave (southern Poland) case study
Supplemental material, Zarska_Table_8_Appendix_-_carniv_REVISION for Late Glacial and Holocene paleoecology and paleoenvironmental changes in the northern Carpathians foreland: The Żarska Cave (southern Poland) case study by Jarosław Wilczyński, Maciej T Krajcarz, Magdalena Moskal-del Hoyo, Witold Paweł Alexandrowicz, Barbara Miękina, Andrea Pereswiet-Soltan, Krzysztof Wertz, Grzegorz Lipecki, Adrian Marciszak, Lembi Lõugas, Michał Gradziński, Anita Szczepanek, Albert Zastawny and Michał Wojenka in The Holocene
Footnotes
Acknowledgements
The authors express sincere thanks to G Haynes and JM del Hoyo-Meléndez for proofreading the final manuscript.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The study of the Żarska Cave is part of a larger ongoing research program supported and financed by the National Science Center, Poland (Grant No. 2013/11/D/HS3/01877 awarded to MW). Radiocarbon dating was partially financed by the National Science Center, Poland (Grant No. 2013/14/D/HS3/03842 awarded to MTK). The anthracological analysis was also partially financed by the Statutory Research Tasks of the W. Szafer Institute of Botany of PAS, while the chiropterological study was supported by Grant No. 2016/20/S/ST10/00163 from the National Science Center, Poland.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.