
Abstract
This paper presents an original palaeoparasitological study performed on samples from a sedimentary core in a peat bog (Asi Gonia, White Mountains, Crete, Greece). The aim of the study is to test the preservation and concentration of parasitic remains in peat sediments, to discuss animal presence around the site throughout the record, and to compare the results with other biomarkers of environmental history. In this aim, 22 sediment samples distributed between the Roman period and the present were processed and the residues were observed under light microscopy in search of parasitic markers (microscopic eggs or oocysts). The majority of the samples (86.4%) tested positive for the presence of helminth eggs. Several taxa were observed throughout the studied period, including Trichuris sp., Ascaris sp., Capillaria sp., Fasciola sp., Paramphistomum sp. and Macracanthorhynchus sp. The assemblage of parasite markers changes throughout the record and the variations show close correlations with previously published grazing indicators (fungal spores) and vegetation changes (pollen grains). This study sheds lights on animal associations and changing environments in the watershed over the past 2000 years. Indeed, we detected three main phases consisting of: Roman animal herding with pigs in an evergreen oak forest, then a more irregular but almost constant presence of ruminants during a second Byzantine and modern phase in a heather maquis landscape and, finally, present-day grazing in a phrygana/steppe landscape. These results highlight how palaeoparasitology can contribute to multi-proxy approaches in peat bog sedimentary sequences.
Introduction
Palaeoparasitology is the study of the evolution and the ecological history of parasitic organisms through their recovery in ancient materials, be they archaeological, palaeontological or of medical nature (Araujo et al., 2003; Dittmar et al., 2012). Oviparous species of gastrointestinal parasite worms produce partly chitinous microscopic eggs which are resistant to taphonomical constraints. These preserved eggs indicate the presence of parasites, and can be extracted and identified from various samples. The more commonly studied samples in palaeoparasitology correspond to ancient faeces (also called coprolites), or samples from highly anthropic contexts, such as burials, latrines, pits or sewers. In such archaeological contexts, parasites can provide information on past societies regarding health, sanitary conditions, diet, cooking habits, agronomy or migrations (Le Bailly and Araujo, 2016; Reinhard et al., 2013).
Over the past years, an increasing number of analyses have been performed on non-human materials, opening up the study of ancient animal (non-human) parasitic diversity (Beltrame et al., 2011, 2014, 2015a, 2015b, 2017, 2018; Borba-Nunes et al., 2017; Dentzien-Dias et al., 2013; Dufour et al., 2015; Fugassa et al., 2018; Le Bailly and Bouchet, 2010; Leles et al., 2018; Sianto et al., 2012, 2014; Slepchenko and Reinhard, 2018; Vieira de Souza et al., 2012; Wood et al., 2013). In such contexts, parasites can be used as evidence for animal presence, and shed light on the spread of parasitic species in wild fauna through time.
In her PhD thesis performed on several lakeside settlements in the Alp Mountains between 1998 and 2001, Dommelier-Espejo was the first to conduct palaeoparasitological analyses on samples from sediment cores taken from lake banks (Dommelier-Espejo, 2001; Dommelier-Espejo and Pétrequin, 2016). The initial aim was to grasp the evolution of parasite diversity during the occupation period of the archaeological lakeshore sites. Since then, cores from other archaeological contexts have also been studied in palaeoparasitology (Dufour, 2015; Ledger et al., 2018; Maicher, 2019). However, despite its interest, this methodology had never been applied to landscapes outside archaeological sites.
Gastrointestinal parasite eggs are sometimes retrieved during palynological studies, where they are considered as non-pollen palynomorphs (NPP), among others (Brinkkemper and van Haaster, 2012; Hillbrand et al., 2014; Miras et al., 2015; Tunno and Mensing, 2017; van Geel, 1978; van Geel et al., 2002). Up until now, environmental reconstructions reporting positive results have never been associated with palaeoparasitological analyses, following the standards in this field.
Here, we present an original palaeoparasitological study performed along a core extracted from a peat bog, located at Asi Gonia in the White Mountains of Crete, the largest Greek island in the Aegean Sea. The aim of this paper is to investigate the preservation and concentration of parasitic remains in peat sediments and, consequently, to discuss animal presence around the site throughout the record. Finally, comparisons with other environmental proxies, especially dung fungal spores, may enable us to document and discuss changes in grazing activities in the watershed from the Roman period until today, and in particular the type of husbanded animals. A recent multi-proxy study showed that the landscape around Asi Gonia was mainly of anthropogenic origin, and particularly dependent on grazing activities in the watershed (Jouffroy-Bapicot et al., 2016). Indeed, the analysis showed important accumulations of spores of coprophilous fungi, especially at the onset of bog formation. Dung fungal spores are generally related to the presence of herbivorous animals (e.g. Gill et al., 2013; van Geel and Aptroot, 2006), and have been linked to pastoral pressure in terrestrial ecosystems through palaeoecological studies (e.g. Baker et al., 2016; Cugny et al., 2010). A recent study showed that coprophilous fungi accumulation in moss-pollster was highly dependent on local grazing pressure in the present-day Western mountains of Crete (López-Sáez et al., 2018). Thus, based on the grazing history of this peat bog, we assume that gastrointestinal parasites in the peat bog sediments are associated with the local presence of animals. If this first hypothesis is validated, then results can be analysed in terms of parasite concentration and diversity as a result of grazing type and intensity. Finally, these observations are compared and discussed in the light of other palaeoenvironmental proxies, and in particular correlated with coprophilous fungi, as suggested by Ejarque et al. (2011).
Materials and methods
The peat bog is located on the territory of Asia Gonia, which is a town in the White Mountains of Western Crete (Figure 1), and more specifically, on the eastern edge of the White Mountains (Lefka Ori). The peat bog is a spring-fed formation supplying the River Koularas, at an elevation of 780 m. Despite the prevailing Mediterranean climate in Crete, the region of Asi Gonia is significantly wetter. This peat bog is the only one known in Crete, and was studied by Jouffroy-Bapicot et al. (2016) through a high-resolution multi-proxy analysis of a 6-m-core from the deepest part of the bog and dated from ca. 100 BC (late Hellenistic to early Roman transition) to the present-day, by means of 15 radiocarbon dates. This first study resulted in the analysis of palaeoenvironmental evolution over the past two millennia, with high temporal resolution. The location of the Asi Gonia peat bog at the bottom of a small watershed represents an ideal situation for a very localized collecting effect (Baker et al., 2013). As a part of our study, we analysed 22 sediment samples regularly distributed along the peat core. Each sample consisted of 2 g of wet sediment and was processed following the standard RHM protocol used in the Chrono-environment laboratory to extract parasite markers (Dufour and Le Bailly, 2013). This protocol consists of a three-step process consisting of the rehydration, homogenization and microsieving of the samples. Thus, our samples were placed in a 50 mL solution of 0.5% trisodium phosphate and 50 mL of 5% glycerinated solution for 1 week. A few drops of 10% formalin solution were added in order to avoid algal or fungal development. It was shown in a previous study that this method results in good recovery of parasitic diversity, unlike more aggressive solutions (Dufour and Le Bailly, 2013; Reinhard et al., 1986). The samples were then crushed in a mortar and sonicated in an ultrasonic device for 1 min at 50/60 Hz, in order to separate the parasitic markers from the surrounding sediments. Finally, the samples were strained through a microsieving column of 315, 160, 50 and 25 µm meshes. Only the 50 and 25 µm residues were kept, as they should contain all the observable parasitic markers (i.e. the eggs), ranging from ∼30 to ∼160 µm long. After 24 h of sedimentation, the samples were ready for microscopic analysis. A total of 36 slides (22 mm × 22 mm) were read for each sample, consisting of 18 slides from the 50 µm sieve, and 18 slides from the 25 µm sieve. Thus, a total of 792 slides were observed under the light microscope (Leica DM2000 LED). We added three surface samples consisting of two soil samples from the bog surface, and one modern sheep faeces lying on the ground near the core location. These samples were considered as control samples to test for possible contamination by modern parasite eggs.

Location of the Asi Gonia peat bog in the Mediterranean Sea and nearby environment. Left: the Asi Gonia peat bog is situated in Western Crete, Greece. Right: sheep are reared nearby the peat bog location. Photography: I Jouffroy-Bapicot.
Results
Parasite diversity and possible hosts
Nineteen samples (86.4%) tested positive for the presence of gastrointestinal parasite eggs. In all, 58 eggs were observed, corresponding to 9 morphotypes, identified on the basis of their morphological traits. Five genera of the Nematoda phylum were identified, namely Trichuris sp., Capillariids, Ascaris sp., plus a strongyle-like type and an unidentified morphotype for which some hypotheses can be advanced. Two genera belong to the Trematoda class, namely Fasciola sp. and Paramphistomum sp. Finally, a single morphotype belongs to the Acanthocephalan class, the Macracanthorhynchus genus (Figure 2).

Microscopic eggs observed at Asi Gonia: 1 and 2 – unknown Trichocephalida; 3 and 4 – striated Capillaria sp.; 5 and 6 – punctuated Capillaria sp; 7–10 – Trichuris sp., showing wide size variation; 11 and 12 – Ascaris sp.; 13 and 14 – strongyle-like egg; 15 and 16 – Paramphistomum sp.; 17 and 18 – Fasciola sp.; 19 and 20 – Macracanthorhynchus sp. Scale = 50 μm.
The first parasites of the Nematoda phylum correspond to the whipworm genus Trichuris. They consist of lemon-shaped ova, because of the presence of polar plugs. Their surface is smooth, with no ornamentation (Horton, 2014). Most of the time, mucous plugs and their internal content are missing in palaeoenvironmental contexts. This genus encompasses several species infesting the caecum and colon of mammals and birds. The size of whipworm eggs varies and shows large overlaps, which only allows for partial identification (Dufour, 2015). The size variation of six Trichuris sp. eggs observed in the Asi Gonia peat bog sediment, of approximately 57.8 µm ± 8.6 × 33.2 ± 3.9 (n = 6), may be attributed to the presence of T. trichiura and/or T. suis, respectively, infesting humans and pigs (Figure 2). We cannot completely rule out a human origin, but an animal origin is more likely, considering the Asi Gonia peatbog context.
The second member of the Nematoda phylum was classified in the capillariid group. Capillariid eggs are similar to whipworm eggs. They also display polar plugs, but are more barrel-shaped with a marked angle around the polar aperture, which is not visible in Trichuris sp. Moreover, most of the capillariid eggs show a typically ornamented outer shell. This group comprises a high number of species which are difficult to describe and distinguish (Borba et al., 2019). Capillariids are commonly known to infest the respiratory and alimentary tracts of mammals and birds (Taylor et al., 2016). Three species, with well-described eggs, are known to potentially infect humans, namely Calodium hepaticum (syn. Capillaria hepatica), Paracapillaria philippinensis (syn. Capillaria philippinensis, Aonchotheca philippinensis) and Eucoleus aerophilus (syn. Capillaria aerophila). Several capillariid eggs were observed at Asi Gonia, which can be grouped into two morphotypes. The first morphotype (n = 13) presents striations on the outer eggshell. Striation consists of fine ornamentation tending to organize long protrusions parallel to the greater egg axis. The second morphotype (n = 8) presents a coarsely punctured surface that forms pits with no orientation (Figure 2). These morphotypes are not associated with human-infesting taxa. In these conditions, we can assume that these eggs are of non-human origin, but at the present time, more precise identification is impossible.
One egg attributable to the roundworm genus Ascaris (phylum Nematoda) was observed in the whole core (Figure 2). Ascaris sp. eggs are defined by their oval shape, consisting of an inner, smooth and thick shell, coated with a mamillated outer shell. The only observed egg from Asi Gonia measured 68 µm × 51 µm, which falls within the known variation of this genus (50–75 µm × 40–55 µm, according to Taylor et al., 2016). This genus is often divided into two host-specific species, namely A. lumbricoides and A. suum, respectively, infesting humans (and other primates) and pigs (wild and domestic). Meanwhile, eggs of both species are very similar and it is not possible to distinguish them from each other on the basis of morphometric traits. Moreover, the molecular similarities of both taxa may point toward a single species, namely A. lumbricoides (Leles et al., 2012), though this point is not settled yet (Søe et al., 2016). In archaeological contexts and highly anthropic places, such as ancient latrines or sewers, such a finding could reliably be interpreted as being of human origin. The case of Asi Gonia differs from this typical situation and an animal (non-human) origin of the roundworm eggs can be proposed.
An unknown morphotype was also observed in the Asi Gonia peat bog sediment (Figure 2). On the basis of its morphological criteria, it can be classified in the Nematoda phylum, order of Trichocephalida. In the same way as the previous Trichuris genus and Capillariids, the eggs are lemon-shaped with two polar plugs. The aspect of the plugs, without angulation, points towards a Trichuris-like morphology. However, the presence of a variable ornamentation on the outer eggshell, points to a Capillaria-like morphology. As far as we know, this association of morphological traits does not correspond to any species in the parasitological literature. This morphotype is the most abundant in our study, and was observed in 11 out of 22 samples for 21 out of 58 remains. Interestingly, this unknown morphotype has already been observed in Neolithic sites in Europe, and more recently described in domestic camelids from pre-Hispanic Peru (Le Bailly et al., 2019). All these observations strongly suggest that this taxon parasitizes herbivorous hosts. Regarding its frequency along the Asi Gonia sequence, its phylogenetic identification and comprehension of its life cycle may provide a strong marker of animal presence in the future. Moreover, its presence up to the 19th century may be indicative of a currently undescribed parasite still circulating in present-day herbivorous fauna.
A single liver fluke genus egg Fasciola was identified in the whole core (Figure 2). This genus belongs to the Trematoda class. Fasciola eggs are ovoid, with a smooth surface, and present an operculum. A mucron can be observed at the opposite of the operculated end. Egg size varies approximately between 130 and 150 µm in length, and 70 and 80 µm in width. Even when considering the squashed aspect of our specimen, its approximate size of 133 µm × 70 µm falls within the expected range. Its general shape is preserved and the operculum pushed into the internal volume is clearly visible (Figure 2). A number of genera belonging to the Echinomastida order contain species producing eggs similar to Fasciola sp. As is the case for Echinostoma sp. or Fasciolopsis sp., these similar eggs generally show slightly different morphological criteria or currently known geographical distribution. This allows us to opt for Fasciola sp. The same is true of the different species of the Fasciola genus, such as F. gigantica. Indeed, the only observed egg at Asi Gonia fits well with F. hepatica. Fasciola needs semi-aquatic conditions to maintain its life cycle, and is commonly known as the liver fluke, which infests sheep, cattle and goats, but also horses, deer, rabbits, hares and other mammals, including humans (Taylor et al., 2016).
A second member of the Trematoda class was also observed. Eggs of the rumen fluke genus Paramphistomum sp. were identified in one of our samples (Figure 2). These eggs are very similar to the Fasciola sp. eggs. They also display a granular eggshell and a typically crenellated aperture edge, as noted by Le Bailly (2005). Two specimens were observed at Asi Gonia, with a size of approximately 156.5 µm × 94 µm (for two measured eggs). This semi-aquatic trematode mainly infests the forestomach of wild or domestic herbivores (cattle, sheep, goats, deer …) (Taylor et al., 2016).
Finally, among the Acanthocephala class, eggs attributed to the genus Macracanthorhynchus sp. were observed in two samples (Figure 2). Macracanthorhynchus sp. eggs typically show a thick, brown oval shell of approximately 90–100 µm × 50–56 µm (Mehlhorn, 2001). The shell consists of four layers, which are often barely visible in ancient contexts, with a fine reticle on the surface. M. hirudinaceus is known to infest the small intestine of wild and domestic pigs and requires various dung beetles as intermediate hosts to maintain its life cycle in a given environment (Taylor et al., 2016).
The last observed egg morphotype is classified as strongyle-like and was observed in one sample (Figure 2). This morphotype encompasses a very wide diversity of parasites belonging to the former Strongylida order. The eggs are colourless and oval, although they can be more or less symmetrical depending upon the species, non-operculated, and with a thin and smooth shell. Their size varies greatly with large overlaps between the different species. Identification is generally possible when coprological studies from known hosts are carried out, based on the host species, morphological criteria and the internal content. This content is systematically lost in palaeo/archaeological contexts, and the host is rarely known. Thus, the identification of these remains is highly speculative. Moreover, in the Asi Gonia peat bog sediment, only one specimen was observed, with a size of 114 µm × 59 µm.
Preservation and distribution
The preservation status of the parasite remains is very good (Figure 2). Only for a very small number of specimens, it was not possible to establish a formal diagnosis at the genus level. Thus, distinguishing morphological criteria from purely taphonomical alteration did not present a challenge for our study. Meanwhile, the less resistant mucous polar plugs observable on eggs of the Trichuris or Capillaria genera, were generally no longer present, as is usually the case in palaeoparasitology for specimens attributed to the Trichocephalida order. Thus, for the measurements of these specimens, the length was systematically considered to be the distance between the edge of their polar apertures, and not the distance between the tops of their lost plugs, as would be the case in fresh coprological studies. As previously noted (Dufour, 2015), these taxa logically appear to be slightly smaller than their extant counterparts.
Despite the inherent limitations of the studied samples (i.e. from a single sediment core and the absence of comparison), we assume that the number of remains and the number of taxa per sample vary along the core, as shown in Figure 3. Several morphotypes were very common in all samples, such as Capillaria sp. (41% of the studied samples, 21 out of 58 counted remains), while others were scarcer, such as Fasciola sp, Ascaris sp., or the strongyle-like specimen (only one specimen each) or Paramphistomum sp. (two specimens) (Supplemental Material Table 1, available online). Four main parasite assemblages (PA) can be visually distinguished (Figure 3). Samples from PA1, dated from ca. 100 BCE to 380 CE, contain a relatively high quantity and show a high diversity of remains per sample compared with samples from other phases. Samples from PA1 are particularly marked by the presence of pig-related helminth morphotypes (Trichuris sp., Macracanthorhynchus sp.). During PA2, ranging from ca. 380 to 900 CE, the quantity of helminth eggs and the diversity of morphotypes both decrease sharply, and no remains were observed in the last two samples. Then PA3, lasting from ca. 900–1850 CE, is characterized by an increase in parasite diversity, marked by an increase in ruminant-related helminth eggs (Fasciola sp., Paramphistomum sp.). Finally, the samples from PA4, corresponding to the 20th century, contain very few or no parasitic remains.

Synthesis graph of palaeoparasitological results confronted to fungal remains influx and pollen-inferred vegetation zones. Parasitic eggs diversity and number per sample and corresponding visually established parasite assemblages; total influx of coprophilous and saprophytic fungal remains and zones according to cluster analysis (Supplemental Material Figure 1, available online); vegetation phases according to Jouffroy-Bapicot et al. (2016), and historical periods. Shaded hosts stand for uncertain identifications. Red bars stand for samples without parasite eggs.
Discussion
Comparing local palaeoparasitological biodiversity with zoological and archaeozoological records in Crete
One of the main drivers for the presence of parasites in a given environment is the presence of suitable definitive hosts in this environment. For this reason, on a worldwide scale, the main factor behind parasitic diversity is host diversity. Parasitic diversity can consequently be used as a proxy to test for host diversity in the surroundings of the Asi Gonia peat bog over the past two millennia. Therefore, it is interesting to review archaeozoological data in Crete.
Animal biodiversity through time in the Aegean was not extensively known up until very recently (Masseti, 2012). In Crete, the Pleistocene fauna was characterized by relatively low mammal biodiversity, marked by a few endemic species characterized by dwarfism, as exemplified by Hippopotamus creutzburgi (Masseti, 2012; Vigne, 1999). Mammal diversity during the Holocene is much more extensive, but no Pleistocene fauna appeared to survive into the Holocene, except for the endemic Cretan shrew, Crocidura zimmermanni (Table 1; Vigne, 1999). This broader animal diversity during the Holocene can be explained by the importation of continental species by human settlers from the Neolithic period onwards, be they wild or domestic (Mavridis, 2003; Vigne, 1999). Some of them, like the now endemic Cretan ‘Agrimi’ goat Capra aegagrus, later returned to the wild.
Archaeozoological record in Crete, according to Vigne (1999).
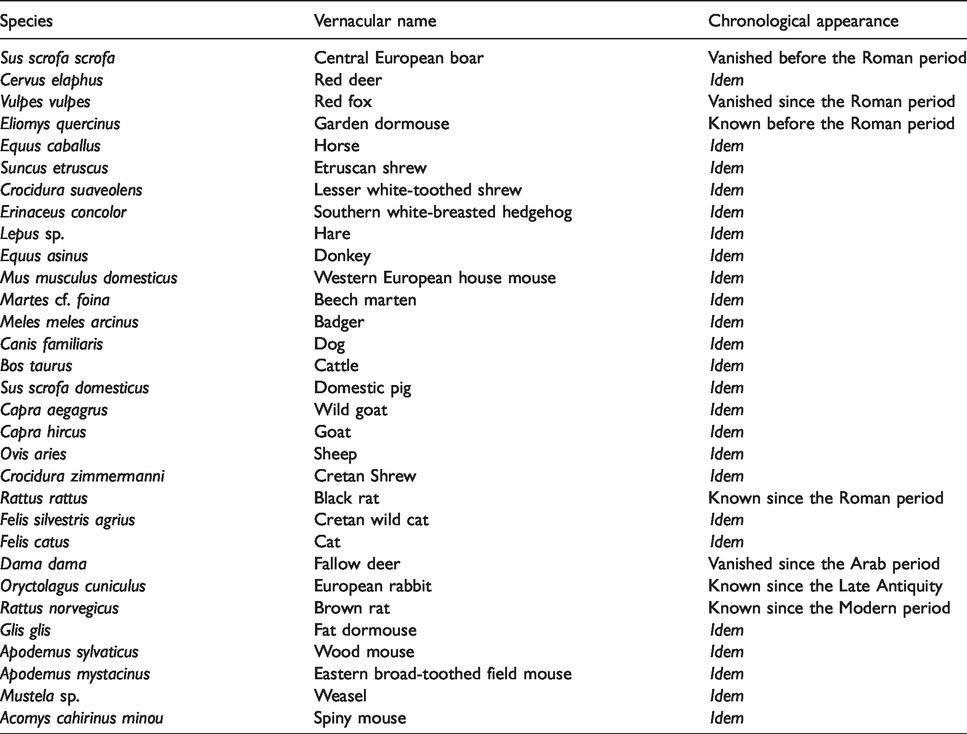
This analysis sheds light on a number of egg morphotypes, mainly related to pigs (such as Trichuris sp., probably T. suis; Ascaris sp., probably A. suum, and Macracanthorhynchus sp., probably M. hirudinaceus) and herbivores, mainly sheep, goats and cattle (Fasciola sp., probable F. hepatica, and Paramphistomum sp.). Birds and rodents could also have participated in this parasitic faunal spectrum, through the dissemination of Capillariid, for example.
Pig-related parasites characterize the oldest phase of the Asi Gonia peat bog record, but no distinction can be made between parasites infesting wild or domestic pigs. It is worth mentioning that the wild boar, Sus scrofa scrofa, is not currently known in Crete (Masseti, 2012). In addition, only the domestic pig, Sus scrofa domesticus, has been identified in the archaeozoological record up until now (Vigne, 1999). Finally, if wild pigs existed during the Minoan period, Nobis (1993) suggested that they could have been large domestic pigs reared during this particularly economically dynamic time. Domestic dogs, Canis familiaris, during the Minoan period (Vigne, 1999), are also known to be occasionally infested by Macracanthorhynchus sp. (Taylor et al., 2016). However, the probable identification of other pig-related parasites, and the complete absence of carnivore-related parasites appear to imply that the swine-related parasitic spectrum at the Asi Gonia peat bog, particularly in PA1 between the 1st century BCE and the 4th century CE, presents strong evidence of local domestic pig rearing.
Herbivores are the second most important host group in our parasitic spectrum, as attested by the presence of Fasciola sp. and Paramphistomum sp., respectively, identified for the period spanning the 10th–16th centuries CE (PA3). Again, wild or domestic hosts cannot be distinguished. As mentioned above, all native herbivores from the Pleistocene in Crete, in this case, the cervid Candiacervus ropalophorus, went extinct before the Holocene (Vigne, 1999). Wild herbivores were later introduced by human settlers during the Holocene, namely the red deer, Cervus elaphus, and the fallow deer, Dama dama (Masseti, 2012), but they disappeared before the period studied at Asi Gonia, or are known to be accidental hosts of the detected parasites, such as Fasciola sp. in the case of the fallow deer (Pybus, 2001). Domestic herbivores are well known in the archaeozoological record from Roman times and later, namely Bos taurus, Capra hircus and Ovis aries. All of them are first-choice hosts for our parasitic spectrum. However, this spectrum cannot distinguish the domestic goat from the feral species, namely Capra aegragus. A few other domestic animals, also known through osteological remains elsewhere in Crete, could have hosted this parasitic diversity, such as the horse and the donkey, Equus caballus and Equus asinus. Hares of the Lepus genus, identified in the archaeozoological record and commonly infested by Fasciola hepatica, may have acted as a reservoir host in this environment. The role of reservoir host seems less likely for the rabbit Oryctolagus cuniculus, as its distribution seems to be restricted to coastal islets since the late-Roman period in Crete (Vigne, 1999). However, even if the Fasciola sp. remains observed in the Asi Gonia peat bog sediments were disseminated by wandering hares, we can suggest that they may also have been part of a transmission cycle of local domestic herbivores. Indeed, Fasciola sp., if it is really F. hepatica, is not known to be easily maintained in wild animal populations in the absence of nearby domestic animals (Pybus, 2001).
The Cretan archaeozoological record shows a wide diversity of small wild mammals, such as the badger, rat, feral endemic Cretan cat, hedgehog or shrew (summarized in Table 1). Occasionally, this diversity of small wild animals may have participated in the parasitic diversity encountered in the Asi Gonia peat bog record, notably in the case of some of the Capillaria sp. specimens.
Finally, comparing the Cretan archaeozoological record with the palaeoparasitological record in the peat bog led to a better understanding of the zoological spectrum responsible for the parasitic signature through time, and highlighted the significant presence of domestic animals in the island over the past 2000 years.
Palaeoparasitology results compared with vegetation history and grazing indicators: Insights into palaeoenvironmental reconstruction and land use history
Multi-proxy analyses previously conducted on the Asi Gonia peat bog sequence (Jouffroy-Bapicot et al., 2016) provided a high-resolution chronological reconstruction of the trajectory of the Mediterranean mountain ecosystem over the past two millennia. Pollen analyses identified three main phases in the landscape history: (1) a first one where the evergreen oak forest dominated land cover from ca. 100 BCE to 850 CE; (2) replaced in the second phase by a heather maquis from ca. 850 to 1870 CE; (3) which developed during the third phase into a very open landscape of phrygana and steppe, which is still extant today (Figure 3). Based on the analysis of fungal remains, pastoralism is assumed to be an important component of local land use history. Indeed, most of the identified fungal remains are related to saprophytic and/or coprophilous fungi growing on herbivorous dung, such as Sporormiella-, Sordaria-, Podospora- or Arnium-type. However, in the previous study (Jouffroy-Bapicot et al., 2016), fungal spores were described and analysed according to pollen zones. In order to make the comparison between the fungal data set easier and more accurate, we performed a stratigraphically constrained cluster analysis on fungal taxa influxes (with CONISS module included in Tilia Graph; Grimm 1991–1993). The results led to the identification of four main phases (Supplemental Material Figure 1, available online), reported in Figure 3. Fungal Zone 1 (FZ1), from ca. 100 BCE to 650 CE, is characterized by high influxes and a high diversity of coprophilous fungi, both decreasing after 500 CE. During FZ2, from ca. 650 to 950 CE, fungal remains are scarce and not diversified. The quantity and diversity of fungal spores rise again during FZ3, from ca. 950 to 1850 CE. Finally, FZ4 from ca. 1850 CE to the present-day is characterized by an increase in coprophilous fungal spores, which start with low values and reach a maximum at the top of the core.
During parasite assemblage 1 (PA1), from 100 BCE to 380 CE (corresponding roughly to the Roman period), the diversity of parasite types per sample and dung-related fungi both reached their highest value. In their paper, Jouffroy-Bapicot et al. (2016) already pinpointed the importance of pastoralism in the Roman Cretan Mountains. The high level of influxes induces high herbivore density, probably related to local livestock rearing (cow, sheep and goat) rather than to wild fauna (Baker et al., 2016; Gill et al., 2013; van Asperen et al., 2019). The palaeoparasitological approach provides new insights into pig herding. The trading of locally produced goods from sheep and goats (milk, wool …) is attested by historical data in Crete under Roman rule (Chaniotis, 1999). However, much less is known about pig rearing in the uplands. Pigs may have been acorn-fed in the Mediterranean oak forest pasture that prevailed at that time according to pollen results (Jouffroy-Bapicot et al., 2016).
PA2 (380–900 CE), which was characterized by a decline in parasitic diversity, is associated with a similar decline in fungal remains after 500 CE (second part of FZ1). Thus, the beginning of PA2 precedes the beginning of FZ2. Meanwhile, both phases (PA2 and FZ2) end ca. 900 CE. Between 400 and 900 CE, fungal remains and parasites tend to show lower animal presence and probably a decrease in pastoral activities in this part of the Cretan mountains during the first Byzantine Empire to the Arab period. One exception is the increase in coprophilous fungal spore influx around 800 CE, which was an important period of change both in terms of the environment (evergreen oak forest to heather maquis) and culture (Arab conquest of the island) (Figure 3).
PA3 (ca. 900–1850 CE) shows a concomitant and sudden rise in parasite eggs and fungal spores. During this phase, the parasite spectrum is composed mainly of herbivore-related species, when coprophilous fungal spore influxes are lower than in FZ1 and show variations. Thus, animal pressure seems to have varied in intensity under the Second Byzantine Empire, Venetian and Ottoman rule, but it is important to mention that all our samples tested positive for parasitic remains during this time lapse. The overall features of the landscape did not change, as heather maquis still prevailed. This last observation suggests that pastoral activities during this period participated in, or at least were not unfavourable to the persistence of this land cover.
The onset of the fourth and last parasite assemblage (PA4), dated to the mid-19th century, shows a new decrease in dung fungal spores in conjunction with the absence of parasite remains. At the same time, vegetation changes from a heather maquis to a phrygana steppe environment. Finally, the top of the core is strongly marked by fungal accumulation but is poor in terms of parasite diversity and remains. Veterinary treatments may have contributed to the disappearance of helminths in the second half of the 20th century when pastoralism was reintroduced to the area. As such, no helminths were observed in the analysis of surface samples from modern sheep faeces and the soil surface.
Interestingly, no pig-related parasites were observed during the Arab and Ottoman periods, while ruminant parasites were detected at the same time. The political and cultural contexts of these periods may have led to specific agro-pastoral activities in the watershed, including a decrease in pig herding. This hypothesis nonetheless requires more data in order to be fully substantiated (Harris, 1987).
Conclusion
This paper presents the results of an exploratory palaeoparasitological study conducted on the Asi Gonia peat bog in the Cretan White Mountains, in Greece. As far as we know, it is the first systematic palaeoparasitological investigation incorporated in a multi-proxy study performed on a sedimentary sequence from natural archives. We identified at least nine helminth morphotypes, for which assemblages along the peat core showed variations correlated with other environmental proxies regarding changes in vegetation (pollen) and grazing activities (coprophilous fungi). Four main phases could be identified throughout time at Asi Gonia, and three are characterized by concomitant fungal and parasite indicators of livestock. The first phase corresponds to Roman animal husbandry with pigs in an evergreen oak forest; the second phase recorded a more irregular but almost constant presence of ruminants during the second Byzantine Empire and modern times in a heather maquis landscape. Finally, modern grazing led to an increase in the amount of coprophilous fungal spores but very few recent parasitic remains were detected as a result of veterinary treatments. A fourth phase, lasting from the mid-4th to the 9th century CE, corresponds to a decrease in grazing pressure in the vicinity of the Asi Gonia peat bog according to our bio-indicators. Tipping points in landscape change are remarkably correlated to samples without parasite remains and are coeval with low amounts of dung fungal spores. These observations may indicate that, from the Roman period to the 20th century, local pastoralism tended to be linked to the equilibrium of ecosystems, rather than acting as a factor of change.
Our pilot study should encourage palaeoparasitologists and other palaeoecologists to undertake systematic palaeoparasitological analyses in peat bog core studies. Such analyses could be improved by sampling several cores in different locations of a same bog, or by increasing the number of samples and taking the sedimentation rate along each core into account, in order to obtain more reliable and statistic interpretations in the future. Such an approach could also be extended to other natural wetland or man-made contexts. However, the present case study contributes to an enhanced knowledge of animal breeding throughout time and its impact on the formation of the landscape.
This approach fits well with the theoretical framework of pathoecology, coined by Martinson et al. (2003), and developed on a number of occasions since then (Arriaza et al., 2010; Gonzalez et al., 2010; Reinhard, 2008; Reinhard and Araujo, 2015). It contributes to the study of land use and pastoral activities throughout time and to inferring variations in the disease ecology of livestock and human-related communities.
Supplemental Material
sj-xlsx-1-hol-10.1177_0959683620919984 - Supplemental material for Ancient parasites from a peat bog: New insights into animal presence and husbandry in Crete over the past 2000 years
Supplemental material, sj-xlsx-1-hol-10.1177_0959683620919984 for Ancient parasites from a peat bog: New insights into animal presence and husbandry in Crete over the past 2000 years by Kévin Roche, Isabelle Jouffroy-Bapicot, Boris Vannière and Matthieu Le Bailly in The Holocene
Supplemental Material
sj-pdf-2-hol-10.1177_0959683620919984 - Supplemental material for Ancient parasites from a peat bog: New insights into animal presence and husbandry in Crete over the past 2000 years
Supplemental material, sj-pdf-2-hol-10.1177_0959683620919984 for Ancient parasites from a peat bog: New insights into animal presence and husbandry in Crete over the past 2000 years by Kévin Roche, Isabelle Jouffroy-Bapicot, Boris Vannière and Matthieu Le Bailly in The Holocene
Footnotes
Acknowledgements
The authors thank The Greek Institute of Geology and Mineral Exploration (IGME) who authorized the fieldwork and core analyses.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship and/or publication of this article: This project benefitted from funds from the Bourgogne Franche-Comte Region through MSHE C.-N. Ledoux, the CNRS PaléoMEx-MISTRALS programme, and the French National Agency for Research through its programme ‘Investissements d’avenir’, referenced as ANR-10-LABX-52.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.