
Abstract
This paper presents the results of multiproxy research (pollen, charcoal, plant macrofossil and testate amoebae) on the biogenic deposits core from Gorodetsky Moch, an ombrotrophic peatland in western Russia (Western Dvina Lakeland). We reconstructed the impact of disturbance on peatland development in the last 300 years by using chronology of the records based on 14C and 210Pb data set. The multiproxy reconstruction was compared with changes in the land cover using historical maps and Corona images, which provides a unique spatial analysis of past ecological and land-use changes. We aimed to determine the effect of local disturbances (drainage) and land-use changes (landscape openness) on the development of the peatland during the last 300 years. Our study suggests that human activity had a crucial impact on the development of the peatland in the last centuries. The analysis of testate amoebae and plant macrofossils revealed a clear disturbed layer in the second half of the 20th century CE. Most probably, the drainage of the peatland triggered changes in the community of testate amoebae and plants, thereby causing a functional shift in Sphagnum peatland ecosystem. The hydrological stress and vegetation composition shift led to the collapse of mixotrophic testate amoebae. However, the peatland showed strong resilience and recovered toward the end of the 20th century CE and the beginning of the 21st century CE, despite the lower water table. Our study shows an example of the peatland ecosystem that experienced a considerable stress but finally sustained the former function.
Keywords
Introduction
Peatlands are particularly sensitive to environmental disturbances resulting from human activities (Dise, 2009). Throughout Europe, many peatlands have been disturbed due to agricultural intensification and forestry practices in the last centuries (Joosten and Clarke, 2002; Tobolski, 2003; Swindles et al., 2019). Loss of wetlands in the 19th and 20th centuries CE was related mostly with mire drainage and peat exploitation (Chambers et al., 2013; Kołaczek et al., 2018b; Swindles et al., 2016). The development of many of these ecosystems has also been disturbed owing to the effects of deforestation (Lamentowicz et al., 2007; Słowiński et al., 2016; Woodward et al., 2014), fire (Florescu et al., 2018; Marcisz et al., 2015; Tinner et al., 2005), and pollution (Fialkiewicz-Koziel et al., 2016). Therefore, there is a need to understand the effects of disturbances caused by human activity to peatland ecosystem. Especially important in this context are high-resolution studies based on several proxies and detailed chronology (Lamentowicz et al., 2009, 2013; van der Knaap et al., 2011) from ombrotrophic peatlands. They are unique repository of high-quality information about environmental changes because receive water and nutrients through a single source (i.e., rainfall).
The simultaneous application of pollen, plant macrofossil, and testate amoeba analyses has been particularly emphasized in the palaeoecology of peatlands in the last two decades. Multiproxy studies have been conducted mostly in Western and Central Europe (Blundell and Barber, 2005; Gałka et al., 2013) and North America (Booth and Jackson, 2003; Booth et al., 2004); however, palaeoecological studies from the European continental peatlands are very rare (Novenko et al., 2017; Payne et al., 2015; Tsyganov et al., 2019). Eastern Europe is affected by the continental climate conditions; therefore, ombrotrophic bogs that possess a flat surface profile and were classified by Kulczyński (1949) as continental-raised bogs in contrast to Atlantic or Baltic raised bogs of the Northern and Western Europe. Past human impact on these ecosystems that cover large areas in this studied region (Lappalainen, 1996) is also insufficiently recognized. To date, multiproxy research on peatlands in the East European Plain was focused primarily on the Holocene climate and vegetation history including long temporal scales. Their relatively low sampling resolution did not allow for a detailed analysis of short-term events. Studies on peatland in the East European Plain (including pollen, macrofossil, testate amoebae, pollen, humification, and isotope analyses) have been performed in Estonia (Sillasoo et al., 2007), European Russia (Novenko et al., 2015, 2016; Payne et al., 2015; Tsyganov et al., 2019), Ukraine (Kołaczek et al., 2018a) and Poland (Gałka et al., 2015, 2017; Karpińska-Kołaczek et al., 2018).
Droughts that we currently observe, and predictions inspire to study disturbances and resilience of ecosystems. The long-term ecosystem monitoring (Bartczak et al., 2019; Lamentowicz et al., 2016; Marcisz et al., 2014; Słowińska et al., 2010) or experiment (Jassey et al., 2018; Rastogi et al., 2019; Reczuga et al., 2018; Samson et al., 2018) provide a short temporal look at a disturbance. In contrast, palaeoecological multiproxy studies provide a long-term perspective (Marcisz et al., 2016; Zawisza et al., 2019). Reconstructions provide information about the ecosystem state before and after disturbances (Fialkiewicz-Koziel et al., 2016; Lamentowicz et al., 2007), but also give reference conditions (Moore et al., 1999). Such knowledge is important for nature conservation (Hamilton, 2018; Marcisz et al., 2018; Słowiński et al., 2019).
The present study aim to determine the effect of local disturbance (drainage), fires and land-use changes (e.g., opening of the landscape and agriculture intensity in the peatland vicinity) on the development of Gorodetsky Moch peatland (western Russia, Western Dvina Lakeland) in the last 300 years. We hypothesized that the drainage of the bog affected taxonomic composition and functional diversity of testate amoeba as well as changes in vegetation structure causing water table deficit in the peatland ecosystem. Furthermore, we aimed to explore a possible self-regeneration process of the bog and its resilience on a long temporal scale (300 years). In this study, we used a several proxies: testate amoebae, which are very sensitive to environmental changes, were used to reconstruct pH and hydrological variability on peatland ecosystem in the past (Charman, 2001; Mitchell et al., 2007a); pollen and plant macrofossils were used for the reconstruction of the local and regional vegetation cover changes, and human impact (Birks and Birks, 1980; Poska et al., 2004); and charcoal was used for the reconstruction of fire history (Conedera et al., 2009). The palaeoecological results were compared with data on land cover changes developed using cartographic materials (analysis of historical maps and Corona images), which provides a unique spatial analysis of past ecological and land-use changes (Ott et al., 2018; Veski et al., 2005). Moreover, a detailed chronology of the records was based on 14C and 210Pb dating.
Materials and methods
Study site
Gorodetsky Moch (local name: Городецкий мох), an ombrotrophic peatland covering an area of 544 ha, is located in western Russia in Pskov Oblast, close to the Smolensk Oblast border, in the basin of the Western Dvina River and around 16 km NE of Velizh city (Figure 1). The coordinates for the central part of the peatland are 55°44’23.2”N and 31°18’02.8”E. The land relief of the peatland neighborhood was formed during the Valdai Glaciation (Weichselian Glaciation in Northern, Western and Central Europe) (Velichko et al., 2011). The average altitude in the peatland vicinity is between 170 and 180 m a.s.l. Presently, the peatland is surrounded mainly by coniferous forests (Pinus sylvestris, Picea abies) with small-leaved forests (Populus tremula, Betula pendula and Alnus incana), fields, and other wetlands. A drainage ditch is situated in its northern part. The Gorodetsky Moch peatland was drained at unknown time for unknown purposes. Currently, despite the presence of a drainage ditch, the peatland is wet and still retains its original characteristic of a raised bog. The present vegetation of the peatland are dominated by hummocks formed by Sphagnum fuscum and Sphagnum magellanicum, and the hollows is dominated by Sphagnum cuspidatum with Eriophorum angustifolium and Eriophorum vaginatum, Andromeda polifolia, Rhynchospora alba, and Carex limosa. The climate of the area is continental with warm summer (Köppen climate classification Dfb) (Kottek et al., 2006) with a mean annual air temperature of 5.4°C; the warmest month is July with a temperature of 17.8°C and the coldest month is February with a temperature of -6.4°C, with annual precipitation of 738 mm (Weather and climate, 2016).
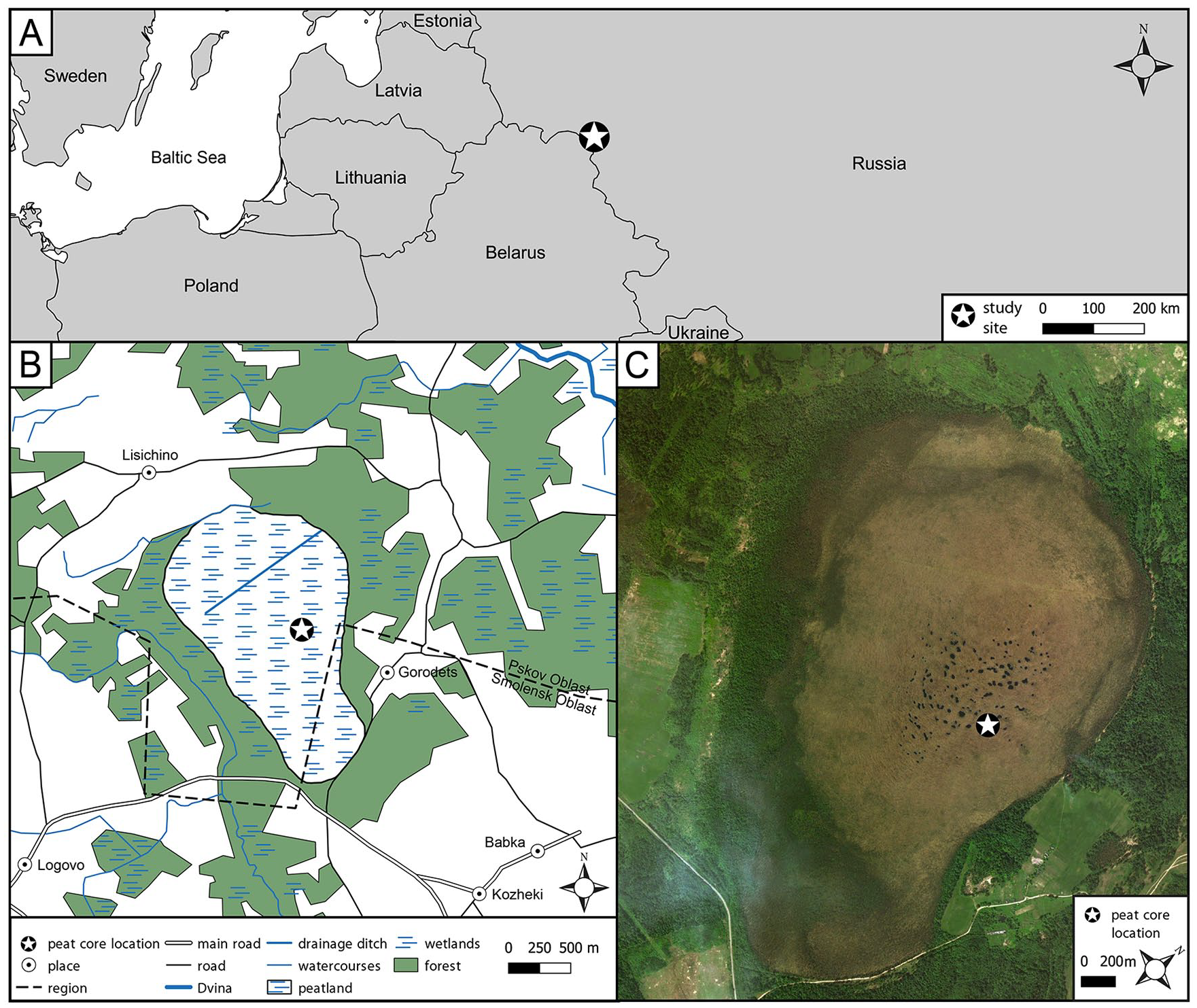
Setting of the study site.
Coring and subsampling
The peat core (dimensions measuring 89 × 10 × 10 cm) was extracted from the central-east part of the peatland in summer 2016. The peat monolith was subsampled every 1 cm (except for the 0- to 2-cm sample) in the laboratory. In total, 47 samples were analyzed for pollen, 44 for testate amoeba, and 89 for plant macrofossils and charcoal.
Chronology
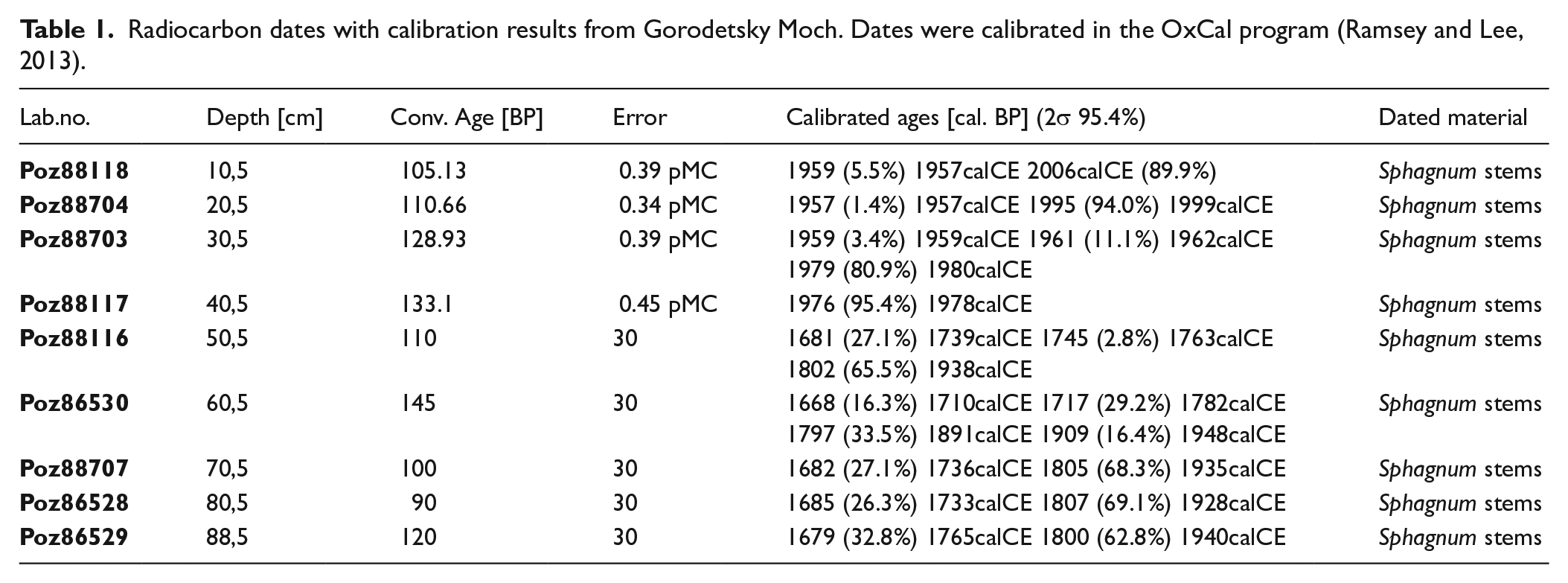
The chronology of the core is based on 14C and 210Pb data set. Nine samples were selected for AMS radiocarbon dating (Table 1). Sphagnum stems of 1-cm thickness and 5 cm3 volume were carefully collected from the samples and treated in Poznań Radiocarbon Laboratory in 2017, and sixty-two 1-cm-thick samples were used for 210Pb measurements at the Institute of Nuclear Physics PAS in Kraków. The activity of 210Pb was determined from the alpha activity of its daughter radionuclide 210Po (half-life 138 days). Secular equilibrium between those radionuclides was assumed because one year had elapsed between the sampling and analyses. Peat samples were spiked with 208Po yield tracer and digested with concentrated acids. Po isotopes were deposited on an Ag disk, and their activity was measured by alpha spectrometry using Ortec silicon detectors. The 210Pb chronologies were established using the constant rate of supply model (CRS model). An age-depth model was calculated using the rbacon in R statistical packages (Blaauw and Christen, 2011) (Figure 2).
Radiocarbon dates with calibration results from Gorodetsky Moch. Dates were calibrated in the OxCal program (Ramsey and Lee, 2013).
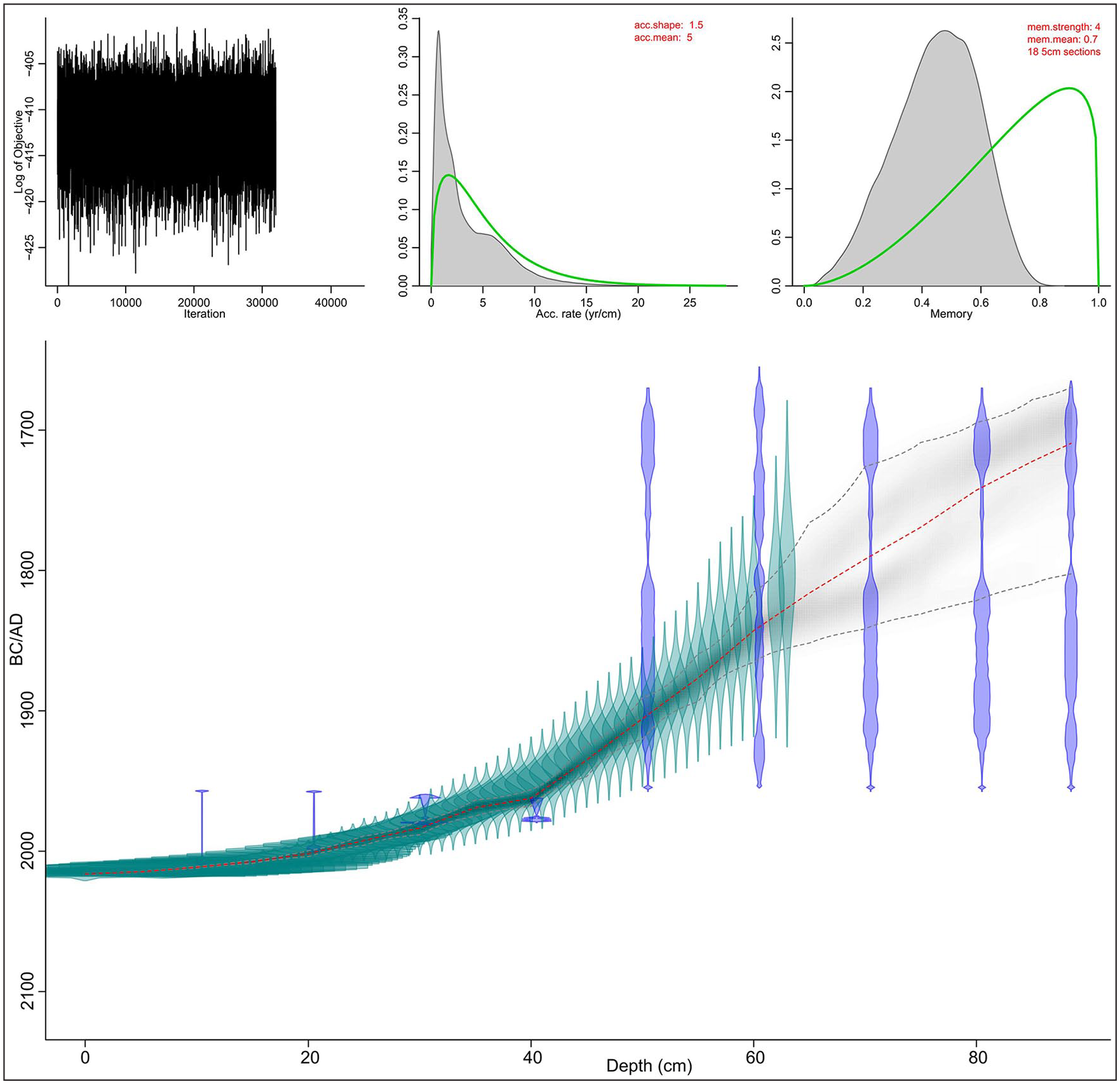
Age-depth model of the peat profile from Gorodetsky Moch.
Pollen analysis
Pollen samples were prepared following the procedure described in a previous study (Berglund and Ralska-Jasiewiczowa, 1986). Every second sample between 2 cm and 46 cm, and the next samples at depths: 48, 49, 50, 52, 54, 56, 58, 60, 62, 64, 66, 67, 68, 70, 72, 74, 76, 77, 78,79, 82, 84, 86, 88, 89 cm (in 1-cm resolution) of volume 1 cm3 were analyzed. The pollen was analyzed using a microscope at 400× and 1000× magnifications. A minimum of 500 arboreal pollens was counted in each sample using a procedure described in the literature (Beug, 2004; Moore et al., 1991). The sums of particular taxa were used to assess the human impact and changes in land-use (Supplementary Table 1) (Behre, 1981; Poska et al., 2004).
Charcoal
Microscopic charcoal (10–100 μm) was analyzed to reconstruct past regional fire activity (Gardner and Whitlock, 2001; Tinner et al., 1998), whereas macroscopic charcoal (>100 μm) was analyzed to reconstruct local fire activity (Clark, 1988; Clark et al., 1998). Microscopic charcoal was counted on the pollen slides until the sum of 200 was reached using a microscope at 400× magnification (Finsinger and Tinner, 2005). Macroscopic charcoal was analyzed in the peat samples of 2 cm3 volume with 1-cm resolution following the procedure described in a previous study (Whitlock and Larsen, 2002), and it was then counted using a stereoscopic microscope at 200× and 400× magnifications in two fractions: 100–500 μm and above 500 μm. Microscopic and macroscopic charcoal influx (particles/cm2/year) was calculated using the charcoal concentration and the peat accumulation rate.
Plant macrofossil analysis
Sediment samples of 5 cm3 volume with 1-cm resolution were washed under running water and sieved through a 0.2-mm-mesh sieve to estimate the values of Sphagnum and vascular plant remains. The macrofossils were identified under a stereoscopic microscope at 200× magnification using identification guides (Grosse-Brauckmann, 1974; Mauquoy and van Geel, 2007; Tobolski, 2000).
Testate amoebae
Testate amoebae samples were analyzed in every second sample (in 1-cm resolution) of volume 2 cm3 following the procedure described in a previous study (Booth et al., 2010). The samples were shaken in distilled water and then washed through a sieve with a mesh of 300 µm. The testate amoebae were analyzed using a microscope at 200× and 400× magnifications. A minimum of 100 tests of testate amoebae were counted in each sample (Payne and Mitchell, 2008). Identification guides were used to recognize testate amoebae taxa (Mazei and Tsyganov, 2006; Siemensma, 2019). On the basis of the construction of tests, the testate amoebae taxa were divided into four categories (idiosomes, idiosomes + organic, protein + calcium, and xenosomes) according to a previous study (Mitchell et al., 2008). Furthermore, three groups were separated: wet indicators (dominant species: Archerella flavum, Hyalosphenia papilio, and Amphitrema wrightianum), dry indicators (dominant species: Cryptodifflugia oviformis, Assulina muscorum, Alabasta militaris, and Difflugia pulex), and indicator of hydrological instability—Arcella discoides (Hendon and Charman, 1997; Lamentowicz and Mitchell, 2005a; Marcisz et al., 2015).
Historical maps and Corona images
Archival maps and Corona images were georeferenced in ArcGIS program. Types of land-use were digitized from the following cartography and photogrammetry archive: (1) Plans of the Russian Empire land survey - Scale: 2 versts in inch (1: 84000) (Period of reflect for Velig region 1750–1780, accessed on 05.2019), (2) Corona images (resolution 2-7 meterdate 1971 July, accessed on 05.2019), (3) Image Landsat ETM (resolution 30 meter, date 2000 May, accessed on 05.2019), (4) Image Sentinel2 (resolution 10 meter, date 2015 May, accessed on 05.2019), (5) Three-verst military topographic map of the Russian Empire - Scale: 3 versts in one English inch (1:126000) (period of reflect 1850–1867, accessed on 05.2019), (6) Topographic military maps of Red Army (RKKA) (the state of the area in 1937–1941, scale 1: 50000 and 1: 100000, accessed on 05.2019). Images were classified using “Iso Cluster Unsupervised Classification” procedure. Types of land-use and forest composition were defined using the data from topographic maps, high-resolution images of Google Earth project, and expert assessment. We distinguished six different land-use categories: (1) arable land-grassland-young cuttings; (2) water bodies; (3) wetlands; (4) open soil-roads-fields after spring fire; (5) settlement; and (6) forests: coniferous (Pinus sylvestris, Picea abies), mixed (Pinus sylvestris, Picea abies, Populus tremula, Betula pendula), and small-leaved forests (forests with Populus tremula, Betula pendula and sometimes Alnus incana domination). The proportions of changes in the land cover during the last 240 years were calculated using ArcGIS program.
Numerical analysis and visualization of the results
Palaeoecological diagrams were plotted using the software Tilia (Grimm, 1992) and C2 (Juggins, 2003). Individual zonation for pollen, charcoal, plant macrofossils, and testate amoebae was based on stratigraphically constrained cluster analysis with the CONISS method (Grimm, 1987) and was plotted using Tilia (Grimm, 1987). Common phases in the peatland development used in the description of results in the article were primarily delimited based on main changes in pollen, macrofossil, and testate amoeba analyses. The testate amoebae were used to reconstruct the hydrological variability (depth to the water table - DWT) and pH with C2 software (Juggins, 2003), using a regional Polish training set (Lamentowicz et al., 2008a). We used the tipping point if DWT=11.7 cm as the reference to define the hydrological stress in the mire (Lamentowicz et al., 2019b). Shannon diversity index (SDI) was calculated to study the community diversity of testate amoebae (Shannon, 1948) using the vegan package (R version 3.5.1) (Oksanen, 2013; Team, 2015). Moreover, six functional traits of the testate amoeba (metabolism – presence or absence of zoochlorella, body size – length of shell, biovolume – the volume of the shell, aperture size – the size of pseudstome, aperture position – location of the pseudostome in a shell, and body range - pseudostome size-body size ratio) were calculated (Fournier et al., 2015; Jassey et al., 2015; Koenig et al., 2018; Lamentowicz et al., 2015; Marcisz et al., 2016) using unpublished trait database using the functional diversity (FD) and vegan packages in R (Laliberté and Legendre, 2010; Oksanen, 2013). TA traits are expressed as the community-weighted mean (CWM) of each functional trait.
Results
Environmental changes in Gorodziecky Moch peatland
Four common phases (Gor-A-D) of the main developmental stages of the peatland were delimited based on pollen (Figure 3), plant macrofossils (Figure 4), testate amoebae (Figure 5), charcoal (Figure 4) and land cover changes (Figure 7).

Pollen percentage diagram for Gorodetsky Moch peatland; 10 times exaggeration is presented as For Peer Review black lines.
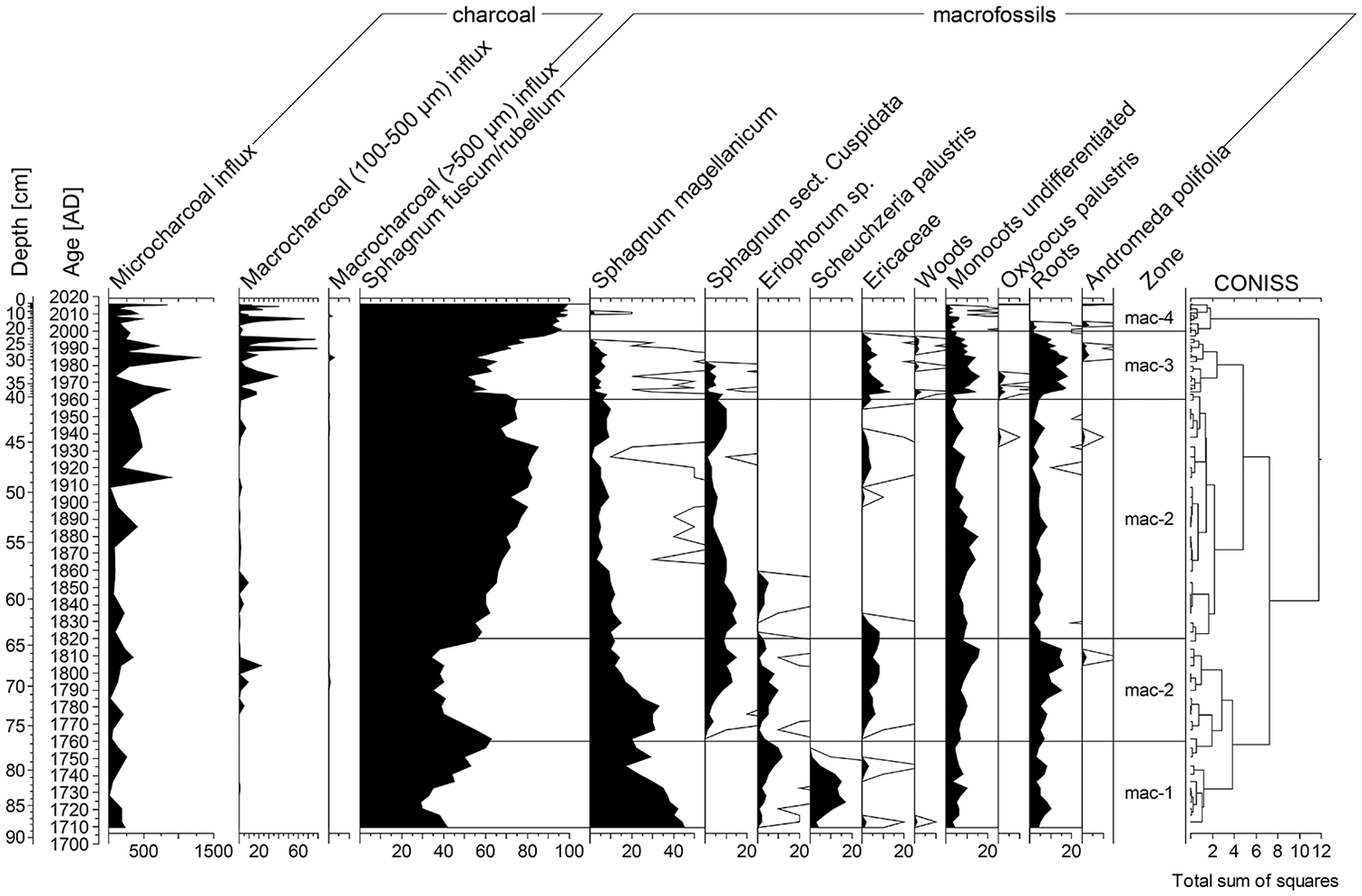
Charcoal diagram presenting fire history and plant macrofossils percentage diagram presenting local vegetation changes in Gorodetsky Moch; 10 times exaggeration is presented as black lines.
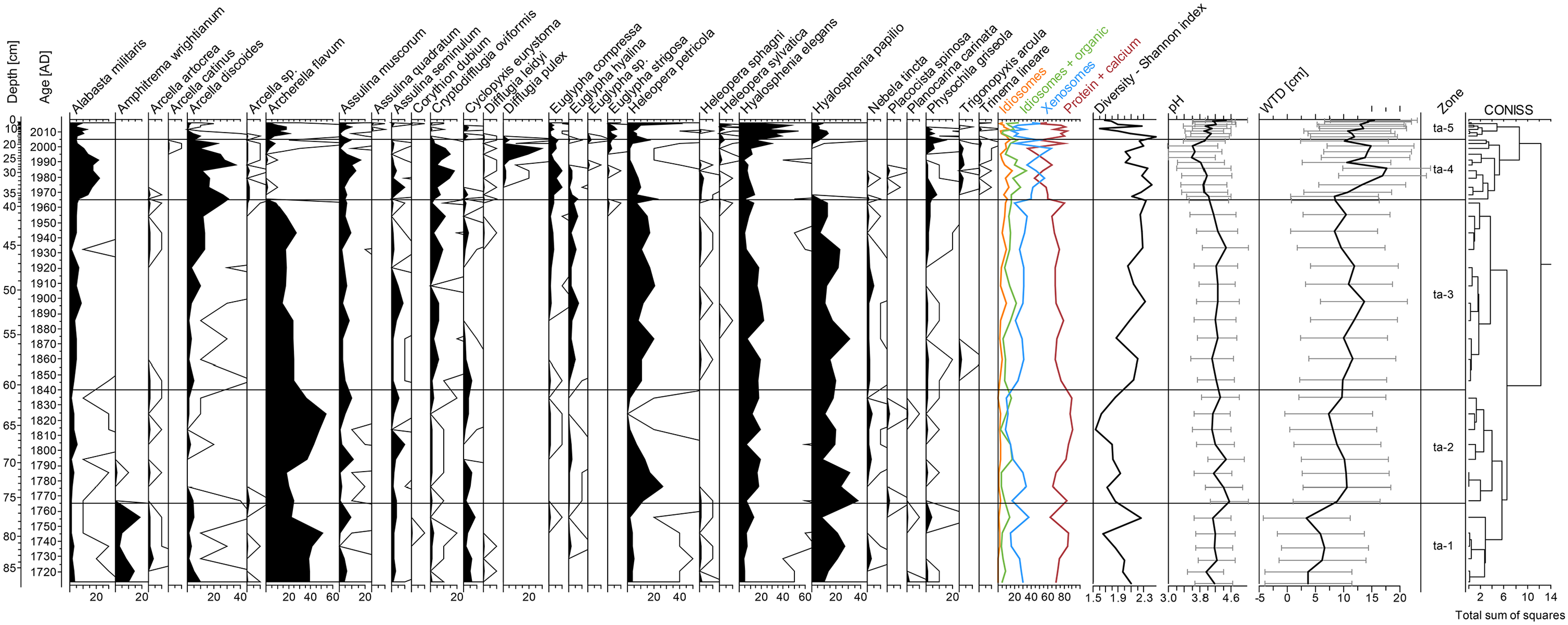
Testate amoeba percentage diagram presenting pH and water table changes in Gorodetsky Moch; 10 For Peer Review times exaggeration is presented as black lines.
Phase Gor—A (1710–1765 CE, 89–74 cm)
In this phase, the tree pollen relative abundance varied from ca 78% to 90%. Here, as well as in the whole section, Betula and Pinus dominated with the participation of Alnus. Picea reached the maximum values in the peat core, and its share increased from ca 6% to 29%. Among herbaceous plants, Poaceae pollen dominated. Microscopic charcoals were remaining at low values (from 6 to 256 particles/cm2/year), whereas macroscopic charcoals practically were absent. In this phase, S. fuscum/rubellum community with Sphagnum magellanicum dominated accompanied with Scheuchzeria palustris. In addition, the phase was characterized by the high share of wet species of testate amoebae and typical species for Sphagnum habitats such as A. flavum, accompanied by H. papilio and Hyalosphenia elegans. We recorded an episode of a higher water table indicated by A. wrightianum. Water table fluctuated in the range of 3.4–7.5 cm, and pH was in the range of 3.95–4.4. Proteinaceous testate amoebae dominated in this phase.
Phase Gor—B (1765–1960 CE, 74–40 cm)
The second phase showed a distinct change in the local vegetation. The relative abundance of AP (arboreal pollen) decreased gradually from ca 83.5% to 53.5%, and the highest decrease in trees pollen occurred in the early 20th century CE. The relative abundance of Picea pollen decreased by few percentages at the beginning of this phase. The relative abundance of Pinus was between 13% and 36.1%, and that of Betula was between 14.4% and 51.7%. Poaceae and pollen human indicators reached the maximum values of 19% and 21%, respectively, in the early 20th century CE. The changes of pollen human indicators were included, among others, the increased to 8% of cultivated plants and also a minor peak of Rumex sum (Rumex acetosa, Rumex acetosella), and Artemisia was recorded at this time. The percentage of Sphagnum spores also increased and remained high during the whole phase. A few macrocharcoal peaks (to 25 particles/cm2/year) were recorded. Microscopic charcoal were remaining at low values to 1870. Since then, we recorded an increase of microscopic charcoal (25–890 particles/cm2/y). S. fuscum/rubellum dominated, with hydrophilous species forming the hummock complex, in this phase also appears Sphagnum sect. Cuspidata. Moreover, plant macrofossils indicated a decrease in Sphagnum and an increase in Ericaceae, monocots, and Carex sp. roots during the first half of the 19th century. The phase was characterized by moderate water table changes, oscillating between 7.4 and 13.6 cm, whereas pH between 4.1 and 4.5. Testate amoebae with proteinaceous shells such as A. flavum, H. papilio, and H. elegans were still dominant. Concurrently, the share of wet indicator species—Heleopera petricola — increased with a distinct peak ca.1775 CE. We also noted a high percentage of A. discoides at the end of the phase.
Phase Gor—C (1960–2000 CE, 40–19 cm)
The proportion of AP pollen varied from around ca. 81% to 93% in this phase. We observed a decrease in Pinus and an increase in Alnus and Betula at the beginning of phases C. Poaceae dominated among the herbaceous plants’ pollen percentage with a minor peak of Vaccinium at the beginning of this phase. The values of microscopic charcoal were still high (from 91 to 1285 particles/cm2/year), whereas macroscopic charcoal (100–500 μm) increased and ranged from 0.5 to 79 particles/cm2/year. The maximum values were reached between 1980 and 1990 CE. The phase was characterized by the decrease in Sphagnum moss and an increase in Ericaceae, monocots, and Carex sp. roots. Sphagnum magellanicum and Sphagnum sect. Cuspidata disappeared at the end of the 20th century. The peatland experienced abrupt water table changes that fluctuated in the range of 8.3–17.7 cm. Testate amoebae species such as: C. oviformis, A. muscorum, and in particular A. militaris and A. discoides reached the maximum growth, whereas A. flavum and H. papilio disappeared. We also observed a rapid increase of D. pulex at the end of the phase. The abundance of proteinaceous testate amoebae decreased in the favor of xenosomic taxa, that built tests from the recycled organic or mineral particles. The pH fluctuated in the range of 3.6 to 4.0.
Phase Gor—D: (2000–2016 CE, 19–0 cm)
The last phase was characterized by a high percentage of AP that increased to 97%. Tree species that were abundant in the peatland vicinity are represented by Betula, Alnus, and Pinus. This phase was also characterized by the high values of microscopic charcoal (from 31 to 848 particles/cm2/y) and macroscopic charcoal (100–500 μm: from 0 to 66 particles/cm2/y; >500 μm: from 0 to 2 particles/cm2/y). Sphagnum increased to above 95%, together with two species of testate amoebae: A. flavum and H. papilio. Concurrently, proteinaceous tests dominated. Testate amoeba biodiversity decreased at the beginning of this phase. The testate amoebae assemblage revealed a decreasing trend of the water level and pH. Water table oscillated between 10.7 and 15.4 cm, whereas the pH was between 3.9 and 4.5.
Functional traits of the testate amoebae
The analysis of functional traits of testate amoebae showed a relatively high variability during the last 300 years (Figure 6). In particular, traits such as body size, biovolume, and aperture size revealed similar changes over time. A visible decreasing trend in metabolism in the peat profile implies the decreasing abundance of mixotrophs. Metabolism CWM values reached minimum between 1965 and 1995 CE, and then started to increase abruptly suggesting higher mixotrophs abundance. Aperture position reached minimum between 1960 and 1970 CE. The maximum fluctuations in the functional diversity started together with the hydrological disturbance during the second half of the 20th century CE. Considering the low water table reconstructed in that time, the all trait CWMs are the most possibly connected with the hydrological fluctuations.

Functional traits of testate amoeba communities calculated from Gorodetsky Moch.
Land-use changes
During 1784–1786 CE, the land covered by forest and used for agriculture occupied 41% and 50% of study area of the Gorodziecky Moch peatland vicinity (Figure 7). The share of wetlands, rivers, lakes, and villages was low. Until CE 1867, forest cover increased up to 60%, while the percentage of agricultural lands decreased to 31%; simultaneously, other land-use types did not change significantly. During 1923–1941 CE, the area of settlements (8%) and agricultural lands (51%) increased, and deforestation was visible (forest area in 1939 CE decreased to 34%). In 1971 CE, many settlements disappeared, and fields were abandoned; therefore, their area decreased (4% and 36%, respectively) and forest cover increased to 54%. The process of depopulation and abandonment of fields continues from 1971 CE to date. Reforestation prevails and forest area increases. The percentage of wetlands and open water reservoirs changed slightly during the last 250 years, it oscillated between 5% and 6.5%.
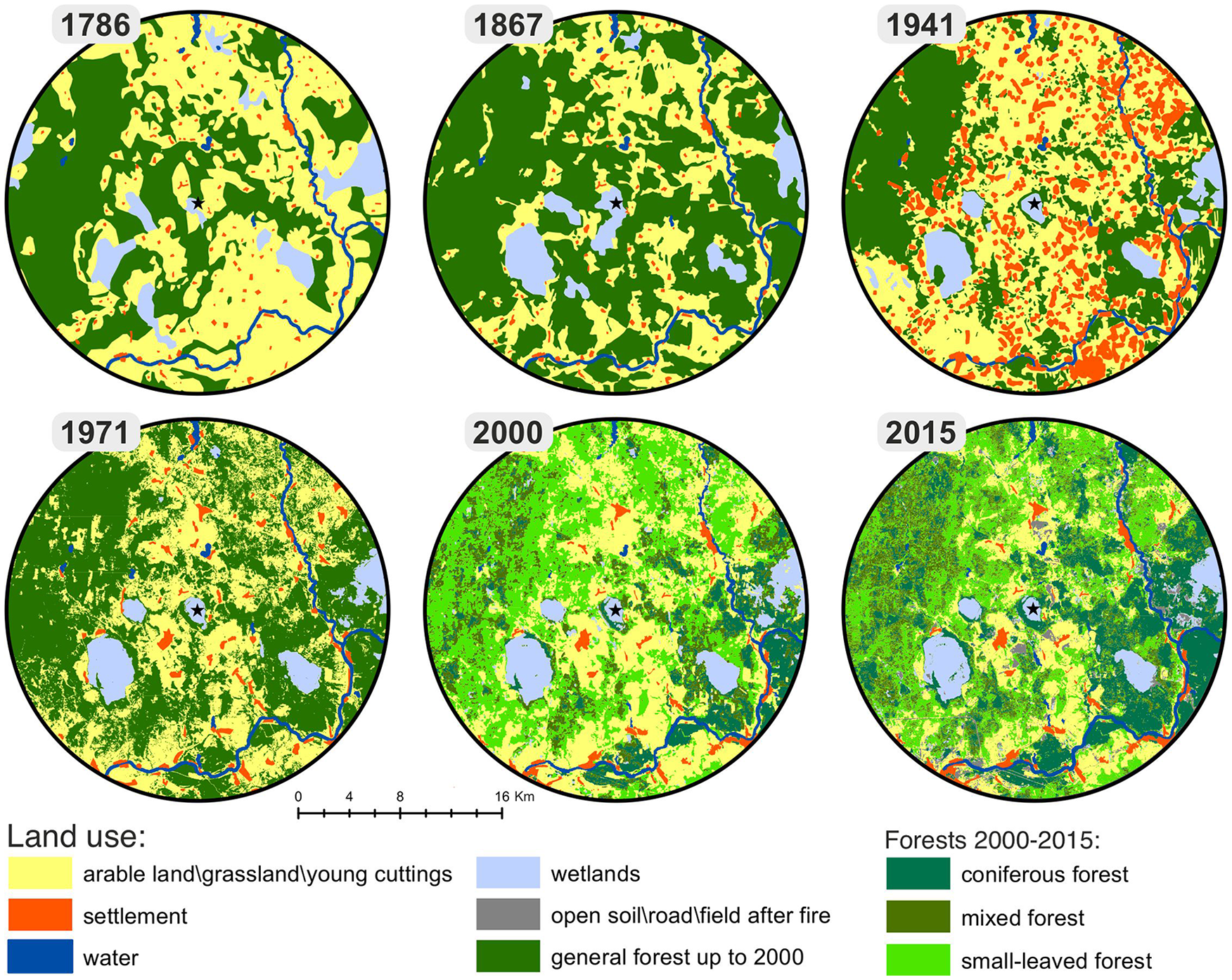
Land cover change map of 1780, 1850, 1939, 1971, 2000, and 2015. The maps distinguished six different land-use categories: (1) arable land-grassland-young cuttings; (2) water bodies; (3) wetlands; (4) open soil-roads-fields after spring fire; (5) settlement; and (6) forests: coniferous (Pinus sylvestris, Picea abies), mixed (Pinus sylvestris, Picea abies, Populus tremula, Betula pendula), and small-leaved forests (forests with Populus tremula, Betula pendula and sometimes Alnus incana domination).
Discussion
The Gorodetsky Moch profile represents a continuous record of peat accumulation during the last 300 years. The multiproxy data with detailed chronology records allowed to identify disturbances to the peatland and their impact on the ecosystem dynamics and palaeohydrology (Figure 8). According to the intensity of human impact on the peatland and local environment, we focused on the changes in the surrounding landscape, dry phase from the second half of the 20th century CE, and peatland recovery after the stress.
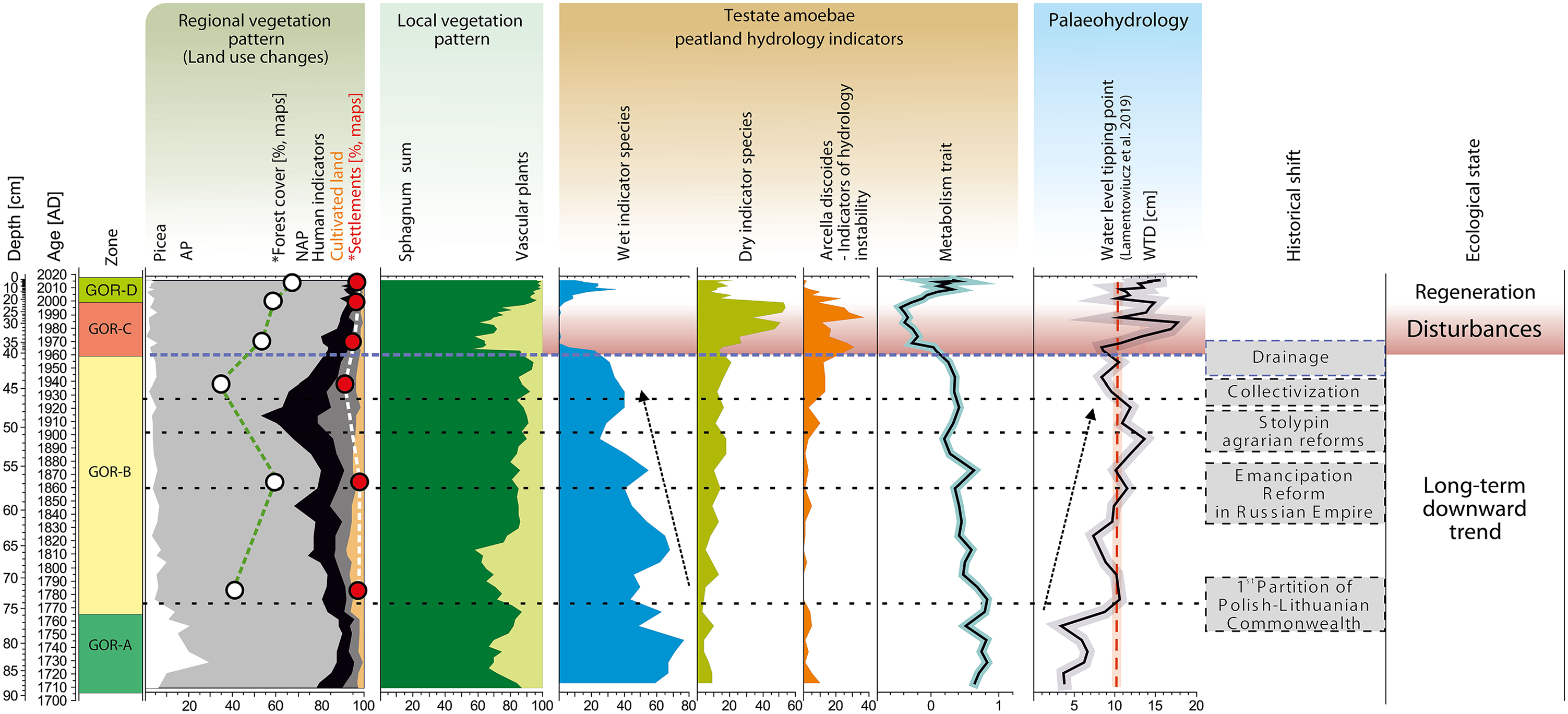
Summary of the most essential trends Gorodetsky Moch peatland. The figure contains: (1) regional vegetation pattern (land-use changes) based on data of pollen, and Historical maps and Corona images; (2) local vegetation pattern based on data of plant macrofossil; (3) testate amoebae peatlands hydrology indicators; (4) hydrology based on testate amoebae; (5) the significant historical shift and (6) ecological state of peatland.
Changes in the surrounding landscape
The important factor that changes the landscape of the peatland vicinity is that at various time periods the region was a border area and an integral part of different states: Grand Duchy of Lithuania (14th–16th century CE), Grand Duchy of Moscow (16th century CE; until 1582 CE), Polish–Lithuanian Commonwealth (16th–18th century CE; 1582–1772 CE), Russian Empire (18th–20th century CE; 1772–1917 CE), Soviet Russia (20th century CE; 1917–1922 CE), Soviet Union (20th century CE; 1922–1991 CE), and Russian Federation (at present, since 1991 CE) (Eremeev, 2010; Shumskaya, 2007). A vulnerable location may have affected the occupation pattern and ancient economy of the region as well as the population density.
Increasing deforestation was observed during the last 300 years (Tarasov et al., 2019). This was also confirmed by the results of research on gully erosion in the lower Serteyka River valley (Piech et al., 2018). In 1714 CE, King Augustus II the Strong (August II Mocny in Polish) allowed for tree cutting in the area of Velizh and privileged the city for producing clapboards, sticks, and logs, among others, resulting in large deforestation and lack of wood in 1773 CE (Eremeev, 2010; Kachulina, 2010). A significant decrease of spruce abundance in the 70s of the 18th century resulted from intense forest exploitation (Figures 3 and 8). However, the forests were not cut out for a longer period until the early 20th century, which confirms Eremeev (2010), as well as the growth of forest cover documented in the pollen analysis results (Figure 3) and on the land cover change maps between 1786 and 1867 CE (Figure 7). Intense felling of forest trees began in the beginning of the early 20th century (Figure 8), as a result of wood industry was developed at that time in the region. A sawmill was built in 1913 CE in Selezni village, and a year later, a shipyard was built in Goriane village for the construction of barges for the army (Eremeev, 2010; Ivanov, 2007). Deforestation is also clearly visible on the map from 1939 (Figure 7). It is highly probable that humans were responsible for deforestation in peatland region during the last 300 years (Figures 3 and 8). However, we did not find significant relationships between the regional vegetation (deforestations) and hydrological changes of the peatland that time (Figure 8). The shift is correlated with decreasing abundance of mixotrophic species and decreasing water table trend since ca 1840 CE. The S. magellanicum to S. fuscum shift is synchronous with the testate amoebae data and might be connected with autogenic succession (Birks and Birks, 1980; Charman, 2002; Zobel, 1988).
The changes in the surrounding landscape of the peatland in the 19th and 20th centuries CE were mostly related to the intense development of human occupation. In that time, the area of agricultural lands and settlements increased significantly (Figure 7). In 1906 CE, 58.6 thousand people lived in the region and only ca 200 people in the Gorodets village in the immediate bog neighborhood in 1912–1913 CE (Eremeev, 2010; Kachulina, 2010; velizh.admin-smolensk.ru). Local human activity was the most intensive in the early 20th century CE (Figure 8). The vicinity of the peatland was used for farming and grazing in that time. This was mostly the result of “the Emancipation Reform in Russian Empire” (from 1861 CE) and of the liberal “Stolypin agrarian reforms” in the early 20th century CE (1906) (Kosonits, 1999). Agriculture, cattle breeding, craftsmanship and fishing developed actively toward the end of the 18th century and the beginning of the 19th century CE in the estate of Selezni village adjoining Serteyka River valley from the west, which was one of the richest properties in the Velizhsky region until 1917 CE (Eremeev, 2010).
During 1929–1932 CE, the forced consolidation of individual peasant households into collective farms called “kolkhozes” was introduced and the private ownership of land (collectivization) was abolished in Soviet Union; moreover, millions of peasant families were dispossessed and deported (dekulakization) (Conquest, 1986). The abundance of cereals in pollen record during that time reached around 2.5% (Figure 3). Depopulation of rural areas of the Soviet Union was observed after 1926 CE (Becker et al., 2012). During that time, the rural population accounted for around 83% of the total population and then dropped to 66% in 1939 CE and to 48% in CE 1952 (Becker et al., 2012). After the German Army occupied the Velizh region, the population decreased, and only ca 600 to 700 people lived in Velizh city in 1943 CE. The higher fire activity during the first half of the 20th century CE (Figure 2) could be partially due to the fire of villages located in the south of the peatland in 1920s CE and because of warfare, as the Velizh region was occupied by the German Army during World War 2. The pollen values of cereals (Figure 3) decreased at the same time as that in the Staroselsky bog, which reflects the decline in agriculture in the area of the forest zone of the former Soviet Union at the end of the 20th century CE (Novenko et al., 2017). Presently, Gorodets village, like many other neighboring villages, was abandoned.
Disturbance and resilience
The results of testate amoeba and macrofossil analyses indicate a layer documenting disturbance (C zone, 1960–2000 CE) in the core starting from the second half of the 20th century CE (Figure 8). After moderate deforestation state and stable wet conditions, a dry phase appeared for about 40 years. It is highly probable that human activity was responsible for the decrease in the water level on the peatland during this time. Gorodetsky Moch was drained, and according to the map from 1989 CE, the location of the ditch is clearly visible, but the exact date of the origin of the melioration works is unknown. However, it is known that the drainage were carried out in Pskov Oblast between 1965 and 1985 CE, in both forest area (148000 ha) and open area (18000 ha) (2009–2010, 2011). This is highly probable that the drainage ditch at Gorodetsky Moch peatland was dug during the 1960s CE.
The disappearance of wet indicator, mixotrophic species of testate amoebae (A. flavum and H. papilio) (Lamentowicz and Mitchell, 2005b) and increased abundance of dry indicators such as C. oviformis, A. muscorum, A. militaris (Lamentowicz and Mitchell, 2005a), and D. pulex indicate dry conditions (Hendon, 1998; Schnitchen et al., 2003). Such composition of testate amoebae, may also suggest a human-induced hydrological disturbances causing peatland habitat instability which has become vulnerable in the periods of drought, what we now observe in results of ecological investigations (Marcisz et al., 2014; Słowińska et al., 2010; Lamentowicz et al., 2020) and in palaeoecological data (Lamentowicz et al., 2008b, 2010; Marcisz et al., 2016). Moreover, A. discoides might be the indicator of hydrological instability (Marcisz et al., 2015; Lamentowicz et al., 2019a) associated with temporal droughts and inundations of peatland surface. This hydrological instability of surface wetness can also be observed in the peatland vegetation cover (Figure 4), namely follows the relationship between species, we note a decrease of Sphagnum fuscum/rubellum and an increase in the abundance of vascular plants. The disturbance phase characterized also by a high diversity of the species composition of testate amoebae, and the SDI was high and fluctuated between 2.0 and 2.5 (Figure 5). We associate the high testate amoebae diversity with drainage, which influenced the seasonal hydrological instability in the ecosystem, and which generated new hydrological conditions with high seasonal fluctuations. This seasonal hydrological instability affects the generation of alternative ecological states, but for a relatively short period causes the possibility of occurrence of a wide range of species. Our research showed that the diversity of testate amoebae was lower immediately after the disturbance (1.5) (Figure 5). SDI ranged from 1.5 to 2.5 for the whole peat core. Moreover, analyzing the functional traits variability (body size, biovolume, and aperture size), we also observed rapid changes within the disturbances phase (during the second half of the 20th century). This phase was simultaneous with a decrease in metabolism and a slight increase in body size (Figure 6). Before disturbance, the body size, biovolume, and aperture size increased, together with a decrease in aperture position, whereas during the disturbance, these traits highly varied. A similar relationship was recorded in a previous study (Marcisz et al., 2016). Different response of proxies to disturbances of Gorodetsky Moch peatland could be related to their sensitivity (Słowiński et al., 2017, 2018), while the value of the multiproxy approach is particularly important when analyzed profile is sampled in high-resolution, that allows to assess the timing of disturbance as well as the regeneration (Colombaroli et al., 2018).
Currently (D zone, 2000–2016 CE), despite the dry phase in the second half of the 20th century CE (C zone), which is caused most probably by digging drainage ditch, the peatland is moderately wet and most likely still retains its original characteristic. The relative abundance of Sphagnum increased together with wet indicator species of testate amoebae (A. flavum and H. papilio) in the top part. Obtained patterns indicate regeneration and confirm that degraded raised peatlands in Europe can sometimes regenerate. Moreover, our study provides another example of high indicative value of mixotrophic testate amoebae (represented by the metabolism trait). Loss of this group should be regarded an early warning signal of Sphagnum peatland disturbance, and their recovery is a promising signal of ecosystem resilience (Jassey et al., 2015).
Climatic background and a forecast for the future
It is also highly probable that climatic conditions impacted the development of the Gorodetsky Moch peatland during the last 300 years. Spinoni et al. (2014) reported that Russia was one of the regions most exposed to prolonged and severe droughts between years 1951 and 1970. This period precedes the disturbed layer and drainage of the Gorodetsky Moch peatland in the second half of the 20th century. The annual precipitation between 1951 and 1970 CE was between 382 and 863 mm, whereas the average annual air temperature was lower than those in previous and later years (Weather and climate, 2016). During 1946–2016 CE, the lowest rainfall (below 500 mm) was recorded in 1946 (439 mm), 1951 (382 mm), and 1959 (458 mm) (Supplementary Figure 1). Studies show that droughts can be the main factor affecting the peatland ecosystem (Ireland et al., 2012). The extended periods with low water table can contribute to changes in the structure of vegetation, especially among mosses (Malmer et al., 1994) such as Sphagnum, which are very sensitive to hydrological changes (Rydin et al., 2006). It was recently revealed that vegetation changes considerably when the water table drops below 11.7 cm in the long time scale (Lamentowicz et al., 2019b). However, it is unlikely that only climate change in such a short time lead to hydrological stress, vegetation composition shift and, as a result the collapse of mixotrophic testate amoebae on the Gorodetsky Moch peatland.
Despite the peatland recovered recently fires are a real threat to this ecosystem considering recent drainage, lower water levels on the peatlands, climate changes and climatic predictions. Global warming in recent decades has significantly increased the risk of the occurrence and spread of fires (Supplementary Figure 1). The effect of prolonged drought and high air temperatures was the reason of the summer fires of forest and peatlands in many regions of the European part of Russian Federation in year 2010 (Hansen et al., 2012; Gilbert, 2010; Shvidenko et al., 2011; Witte et al., 2011). Peat fires were noted mainly in the Moscow Oblast, but also, among others, in the Tver Oblast and Pskov Oblast. Drained peatlands such as Gorodetsky Moch are more susceptible to fires. It is estimated that about 3.8 million ha of peatland in Russian Federation territory (mainly in the European part) have been drained (Päivanen and Paavilainen, 1996). A forecasted increase in the intensity of fires in the current century in Europe (Flannigan et al., 2013) will potentially affect peatlands hydrology and carbon balance. Moreover, the peatlands can often be the only source of water for animals in the face of climate change and the growing hydrological crisis. Therefore, it is important to maintain peatlands in a proper hydrological state and restoring degraded peatlands.
Supplemental Material
Supplementary_file_20May – Supplemental material for Disturbance and resilience of a Sphagnum peatland in western Russia (Western Dvina Lakeland) during the last 300 years: a multiproxy, high-resolution study
Supplemental material, Supplementary_file_20May for Disturbance and resilience of a Sphagnum peatland in western Russia (Western Dvina Lakeland) during the last 300 years: a multiproxy, high-resolution study by Dominika Łuców, Mariusz Lamentowicz, Milena Obremska, Maria Arkhipova, Piotr Kittel, Edyta Łokas, Andrey Mazurkevich, Tomasz Mróz, Rik Tjallingii and Michał Słowiński in The Holocene
Footnotes
Acknowledgements
We would like to thank Mateusz Kramkowski (Institute of Geography and Spatial Organization, Polish Academy of Sciences, Poland) and Bartosz Kotrys (Polish Geological Institute, Pomeranian Branch in Szczecin), and Krzysztof Błaszczyk (Stanisław Sankowski Regional Museum in Radomsko) for their help during the fieldwork. We thank the anonymous reviewers and editor Frank Chambers for their comments and suggestions that allowed them to improving the manuscript. This study will be a contribution to the PhD thesis of Dominika Łuców entitled “Disturbances of Sphagnum peatlands in Central-Eastern Europe.”
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was supported by grants 2019/35/N/ST10/03492, 2018/31/B/ST10/02498, 2015/17/B/ST10/03430, and 2015/17/B/ST10/01656 from the National Science Center (Poland), and the Institute of Geological Sciences Polish Academy of Sciences (project: GORODIEC).
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.