
Abstract
Understanding long-term responses of subalpine forest-parklands to Holocene climate variability in local context is critical for better managing those ecosystems under future climate change. Available records suggest that western North American subalpine forest-parkland ecosystems responded to Holocene climate in various ways at different places. Here we present a Holocene record of upper montane forest-parkland dynamics from Silver Lake in Big Cottonwood Canyon, Wasatch Range, on the Great Basin’s eastern margin. Our results show that at the end of the Younger Dryas, Silver Lake was surrounded by open Pinus/Picea parkland mixed with Artemisia subalpine steppe. By ~9.0 cal ka BP Picea began to expand in response to early Holocene summer warming and enhanced winter moisture. Picea-dominated forests prevailed from ~8.0 to 5.0 cal ka BP, after which time a more open Picea parkland developed. By ~3.5 cal ka BP Pinus increased under cooling conditions, and Picea gradually rebounded to form the mixed conifer forest present today. The Silver Lake long-term record of moderate shifts within an upper montane ecosystem is consistent with other regional sites in the timing of major vegetation changes reflecting large-scale climatic forcings, but distinct in response to local factors including the importance of enhanced lake-effect snowpack and the absence of competing lower montane Pinus. These local factors may help account for the long-term stability of Picea-dominated forests in the area.
Introduction
Understanding variation in montane ecosystems over long time scales is key to effective management under conditions of increased anthropogenic disturbance and rapidly changing climate (Buma et al., 2019; Jackson and Hobbs, 2009). Montane ecosystems encompass substantial topographic and geologic heterogeneity, they endure manifold disturbance factors, and their constituent biota respond to dynamic climatic regimes and successional relationships in complex and often individualistic ways (Daubenmire, 1943, 1956; Fall, 1997; Schimpf et al., 1980; Whitlock and Bartlein, 1993). Montane ecosystem histories yield information about responsiveness, vulnerability, stability, and resilience to climatic change and other disturbance factors.
To contribute to a better understanding of upper montane ecosystem history in relation to long-term climatic variation in western North America, we present a Holocene paleovegetation record from Silver Lake, Big Cottonwood Canyon, Wasatch Range, Utah (Figure 1). Big Cottonwood Canyon is a critical watershed for Salt Lake City and the densely urban Wasatch Front, and provides a range of important regional ecological services. Future climate warming may adversely affect upper montane forest ecosystems worldwide (e.g. Notaro et al., 2012), with deleterious consequences to those services. A long-term perspective on upper montane ecosystem stability gives a point of comparison to consider in developing local management and conservation strategies (Brugger et al., 2018; Buma et al., 2019; Morris et al., 2015; Schwörer et al., 2017).
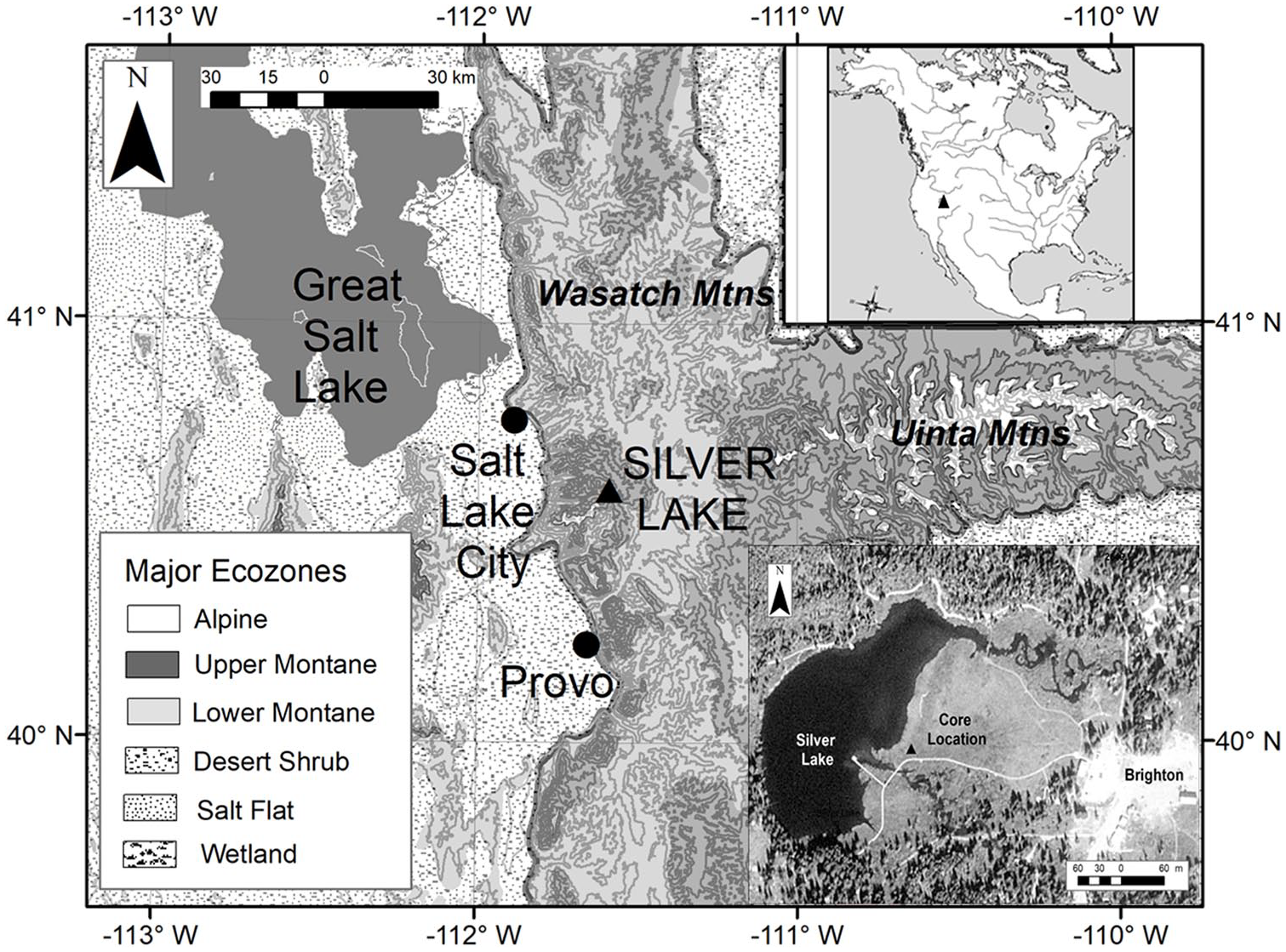
Location of Silver Lake in Big Cottonwood Canyon, Wasatch Mountains, Utah.
The Silver Lake record also fills a valuable position in tracking large-scale hydroclimatic gradients across western North America, bridging the transition between the Great Basin and the Rocky Mountains and Colorado Plateau (e.g. Anderson, 2012; Barron and Anderson, 2011; Hermann et al., 2018; Metcalfe et al., 2015; Mock, 1996; Shinker and Bartlein, 2009, 2010; Shuman and Serravezza, 2017; Thompson et al., 1993; Whitlock and Bartlein, 1993; Wise, 2010). Long-term histories of Great Basin subalpine forests (e.g. Madsen and Currey, 1979; Minckley et al., 2007; Morris et al., 2013, 2015; Shriver and Minckley, 2012; Thompson, 1984; Wahl et al., 2015) are not nearly so well known as Great Basin lowlands (e.g. Louderback and Rhode, 2009; Madsen et al., 2001; Mensing et al., 2004, 2008, 2013; Rhode, 2016; Thompson, 1992; Thompson et al., 2016; Wigand and Rhode, 2002), and certainly not as well-documented as the Rocky Mountains (e.g. Anderson et al., 2008, 2015; Benes et al., 2019; Briles et al., 2012; Brunelle et al., 2005, 2013; Fall, 1997; Fall et al., 1995; Feiler et al., 1997; Jiménez-Moreno and Anderson, 2012; Krause and Whitlock, 2017; Mensing et al., 2012; Minckley et al., 2012; Power et al., 2011; Vierling, 1998; Whitlock, 1993; Whitlock et al., 2012). The Silver Lake record contributes to a developing corpus of local ecological histories across these large spatial and elevational gradients.
Modern setting
Silver Lake is a small tarn (currently ~3.5 ha) located at 2660 m above sea level (asl) next to Brighton at the head of Big Cottonwood Canyon, ~30 km southeast of Salt Lake City. Its east-facing cirque basin lies on quartz monzonite mantled by glacial till (Atwood, 1909; Richmond, 1964). By ~15 cal ka BP glaciers in the canyon retreated to within ~2.5 km of Silver Lake and were likely gone by the Younger Dryas (YD) interval (Madsen and Currey, 1979; Quirk et al., 2018).
Silver Lake currently enjoys cool moderately dry summers and a long snowy cold season (1981–2010 National Climatic Data Center monthly normal [www.wrcc.dri.edu]: mean temperature January = −6.4°C, July = 14.8°C, annual = 2.7°C; mean precipitation December–February = 37.3 cm, March–May = 32.2 cm, June–August = 13.5 cm, September–November = 29.4 cm). Plentiful cold-season precipitation supports vegetation much like the middle Rocky Mountains (Allan, 1962; Arnow et al., 1980; Cronquist et al., 1972; Scott, 1995). In Big Cottonwood Canyon, Acer-Quercus forest/chaparral mixes with Artemisia-grass shrublands below ~2300 m asl (Allan, 1962). Further up-canyon at ~2700 m asl, Quercus chaparral mingles with lower montane conifer forest (Abies concolor, Pseudotsuga menziesii, Picea pungens). Populus tremuloides, Salix, Alnus incana var. tenuifolia, Betula occidentalis, scattered Picea, and other montane shrubs line canyon bottoms. Patchy stands of Picea engelmannii and Abies lasiocarpa grow on footslopes around Silver Lake. A Carex marsh with scattered Salix copses fills the basin’s east side; Salix scouleriana with some Alnus dominates creeks (Allan, 1962: 27). Dry upper slopes and ridges support Pinus flexilis stands and Artemisia-dominated steppe communities, while moist slopes support diverse herbaceous or shrubby meadows. A small alpine association dominated by Asteraceae and Poaceae inhabits the highest peaks >3200 m asl (Allan, 1962; Richmond, 1964).
Methods
We extracted a core from the meadow directly adjacent to Silver Lake (40.604°N. lat.; 111.588°W. long., Figure 1), using a 5 cm manual square-rod piston coring device (Wright et al., 1983). Lacking access to the lake itself, we positioned our coring close to the modern lake shoreline to obtain a record of undisturbed lake and pond sediments underlying modern meadow sediments. Coring reached 2.16 m, but the lowermost 90 cm of coarse sand was poorly recovered so we focused on the upper 1.26 m of retrieved sediments for this study.
Five radiocarbon dates from terrestrial macrofossils and peat were obtained from major stratigraphic breaks in the core and calibrated to calendrical ages (CALIB 7.1 using INTCAL13, Reimer et al., 2013) (Table 1). Plant macrofossils used for dates were identified with reference to modern specimens and published illustrated keys (Cronquist et al., 1972). These calibrated dates (as well as the surface, assumed to be the coring year in 2012) were used to fit an age–depth model consisting of sequential linear interpolations between each date (Figure 2). We used the BACON program (Blaauw and Christen, 2011) to estimate 95% confidence intervals around the resulting age–depth profile.
Radiocarbon dates used for the depth–age model of the Silver Lake core.
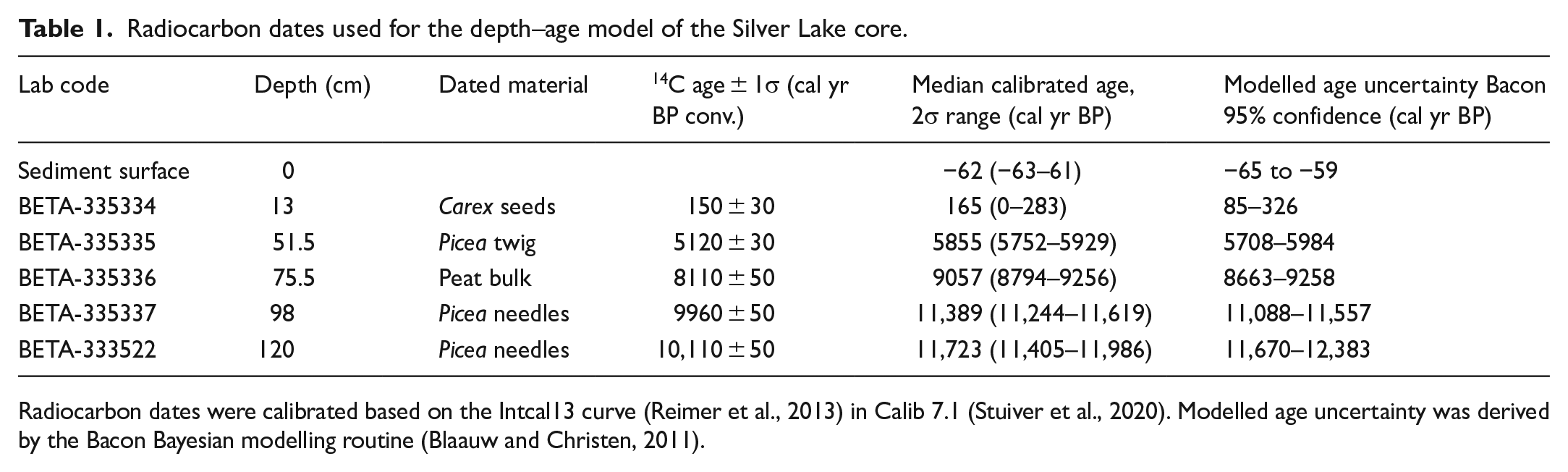
Radiocarbon dates were calibrated based on the Intcal13 curve (Reimer et al., 2013) in Calib 7.1 (Stuiver et al., 2020). Modelled age uncertainty was derived by the Bacon Bayesian modelling routine (Blaauw and Christen, 2011).
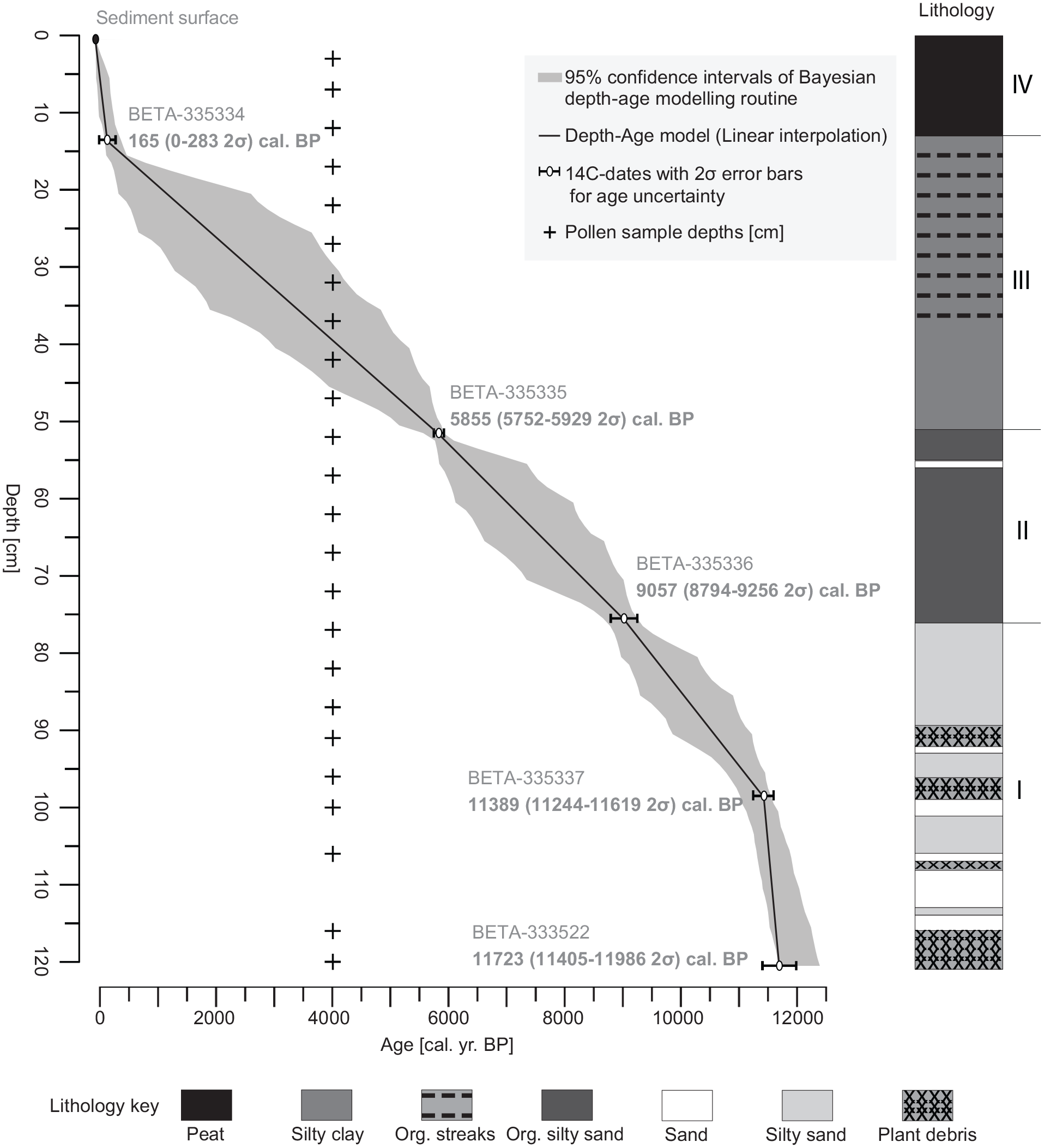
Silver Lake age–depth model. The age–depth model (linear interpolation fitted with CLAM; Blaauw, 2010) is shown as solid black line with a grey envelope delimiting the 95% confidence interval of the estimated ages obtained with the Bayesian age–depth modeling routine BACON. Five radiocarbon dates (white dots with 95% confidence intervals of the calibrated ages) and the sediment surface of the coring year in 2012 (black dot) were used to fit the age–depth model. Ages are shown in calibrated years before present (cal yr BP with 1950 = 0). Sediment accumulation rate (dashed line) is calculated based on the age–depth model. Right column shows major sedimentary units (I–IV).
Twenty-four samples spaced approximately 5 cm apart were taken for pollen and sediment analysis. Sequential loss-on-ignition (Dean, 1974) was used to estimate organic carbon, carbonate, and mineral fractions. Pollen samples were processed using standard treatments (Faegri et al., 1989). Lycopodium tracers were added to calculate concentrations and influx (Stockmarr, 1971). Pollen and non-pollen palynomorphs (NPP) were counted at 400×, identified using reference collections at the Desert Research Institute and in published guides (Kapp et al., 2000; Moore and Webb, 1978). Percentages were calculated based on the terrestrial pollen sum (TPS) including trees, shrubs, and non-aquatic herbs. TPS counts average 267 grains. Two samples in the lowermost coarse sands contain TPS counts of less than 200 grains which may affect the accuracy of proportional estimates of rarer pollen types; however, major trends (as discussed here) may be captured with minimum sums of as few as 50 items (Brugger et al., 2019; Heiri and Lotter, 2001). Counts of aquatic and marsh taxa (Cyperaceae, Typha/Sparganium, Potamogeton, Isoetes, and the microalgae Pediastrum and Botryococcus) are reported as percentages of the TPS. The low temporal resolution prevents a detailed interpretation of short-term fluctuations, but long-term multi-century to millennial vegetation shifts can be reconstructed.
Constrained hierarchical clustering of samples was conducted using CONISS (Grimm, 1987) based on Euclidean distances of square root-transformed terrestrial percentages. Pollen zones derived from clustering analysis were tested for statistical significance (p < 0.05) with the broken stick model (Bennett, 1996). Principal components analysis (PCA) was conducted on the covariance matrix of square root-transformed terrestrial percentages to identify major axes of variation among the samples (Birks et al., 2012; ter Braak and Prentice, 2004); results are provided in Supplemental Material.
Results
Age–depth model and sedimentary units
The age–depth profile is nearly linear across most of the core except for the uppermost modern meadow sediments (<14 cm depth) and the lowermost coarse sand deposits (>100 cm depth). The sedimentary record shows slow and steady accumulation rates after initial rapid infilling (Figure 2) with four distinct sedimentary units, showing no evident unconformities or significant breaks. This consistency across the core suggests that linear interpolations between dates are fully adequate for estimating ages from depths without requiring additional curve-fitting assumptions.
Sedimentary Unit I (~11.7–9.1 cal ka BP) is coarse quartzite sand interbedded with dark micaceous fine sand/silt lenses and Picea needle/twig mats. Organic content is low but increases in the macrofossil mats. Rapid initial deposition of coarse sand (~15 yr/cm) slowed to an average of ~104 yr/cm after ~11.4 cal ka BP.
Overlying Unit II (~9.1–5.8 cal ka BP) contains dark brown/black organic-rich clayey silt interpreted to be shallow lake/marsh muds. The lower unit is massive organic-rich mud while the upper part contains crumbly silt/clays with coarse plant fragments and roots indicating in-situ aquatic plant growth. The sedimentation rate slowed to an average of ~133 yr/cm.
Unit III (~5.8–0.2 cal ka BP) consists of silts and clays with low carbon-organic content, interpreted to be deeper lake/pan muds. Massive dark brown silty clays comprise the lower part (>~4.0 cal ka BP), with thin organic streaking above. Intact diatoms are abundant but roots indicating in situ plant growth are absent. Sedimentation slowed to an average of ~148 yr/cm.
Finally, Unit IV (~<0.2 cal ka BP–present) represents modern Carex wet meadow habitat with dark brown fibrous sedge peat, roots, humified plant matter and silty clays. General Land Office survey maps (AD 1871 and 1902) suggests the Carex meadow around the coring area was filling in historic times, following extensive local mining and logging (Allan, 1962).
Palynological record
We obtained two statistically significant pollen zones, Silver-1 and Silver-2 (Figure 3), and divided Silver-2 into distinct subzones, 2a and 2b (not significant at p < 0.05). The zonation is supported by the ordination analysis separating sample scores according to pollen zones (Supplemental Figure S1).
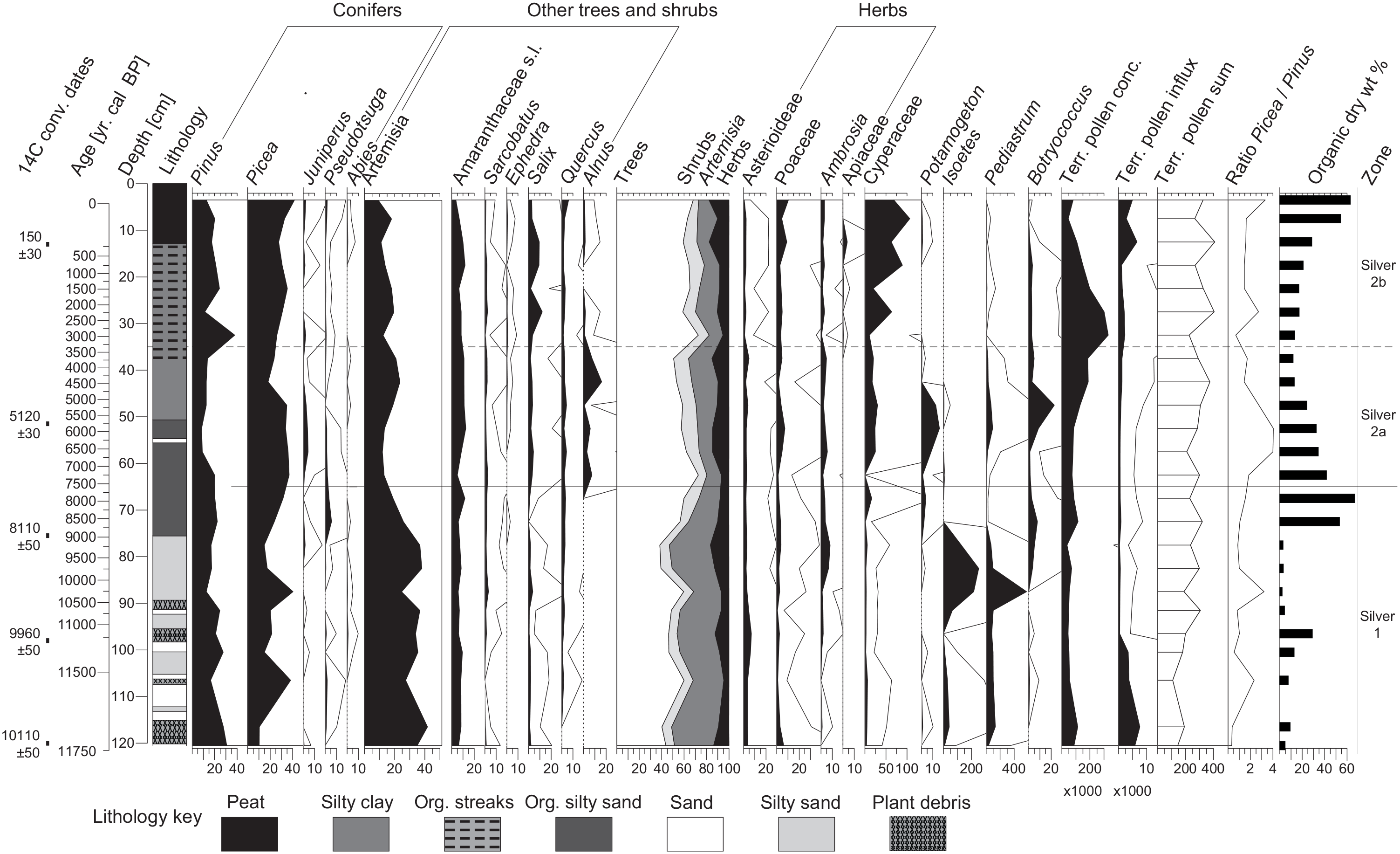
Silver Lake pollen diagram. From left: Selected pollen and non-pollen palynomorphs (NPP) shown as percentages based on the terrestrial pollen sum, terrestrial pollen concentrations and influx, the counted terrestrial pollen sum, ratio of Picea and Pinus pollen, and carbon-organic material as percent of total dry sediment weight (organic dry wt%). Thin lines in pollen abundances show 10× exaggeration. Cyperaceae was excluded from the terrestrial pollen sum as Carex dominates the local wetland.
In Silver-1 (~11.7–7.7 cal ka BP), Pinus (21% median), and Picea (20%) pollen dominate the tree taxa, while Artemisia pollen (35%) dominates overall. Artemisia pollen likely came from subalpine shrubs and herbs (e.g. A. tridentata var. vaseyana, A. arbuscula, A. scopulorum; Allan, 1962; Arnow et al., 1980) and probably lower elevation species as well (cf. Anderson et al., 2015; Fall, 1992; Thompson et al., 2016). Minor contributors include Salix, Asteraceae, and Poaceae, common in subalpine and alpine open habitats, as well as Amaranthaceae sensu lato (formerly referred to as “cheno-ams”) (~7%) and Quercus (~2%) pollen deriving from lower elevations (Allan, 1962; Cronquist et al., 1972). Aquatic taxa include abundant Pediastrum (~100% up to ~400% of TPS) and Isoetes (~42% median to ~250%) before ~9.0 cal ka BP, indicating freshwater dilute lake conditions (Doner, 2009). Botryococcus, Potamogeton, and Cyperaceae all rise after ~9.0 cal ka BP (sedimentary unit II), signaling a shift to a shallow-lake and marsh habitat.
Subzone Silver-2a (7.7–3.5 cal ka BP) spans much of sedimentary unit II and lower unit III. Picea is the dominant arboreal taxon (34%, declining to ~20% after ~5.0 cal ka BP), while Pinus (13%) reaches its lowest values. Artemisia (16%) declines as Picea increases, but after ~5.0 cal ka BP it rises again. Alnus (~7%) appears for the first time. Amaranthaceae pollen increases slightly (11%), as does Quercus (~3%). Herb pollen increases slightly (~11%), predominantly Asteroideae, Poaceae, and Ambrosia suggesting stable meadow and understory habitats. Wetland and aquatic indicators during Silver-2a include Cyperaceae (22%), Potamogeton (~13%), Pediastrum (~29% median), Botryococcus (~4%), and traces of Typha, Isoetes and Nuphar, indicating a diverse open water and marsh mosaic.
During Silver-2b (3.5–0 cal ka BP), Picea pollen remains dominant (31%) but Pinus increases (20%). Artemisia (14%) and Amaranthaceae pollen (8%) declines, while Quercus remains stable, rising only in the modern sample. Among herb taxa, Poaceae pollen increases with historic logging, grazing, and recreation (Allan, 1962). A Carex-dominated wetland develops (Cyperaceae ~64% of TPS), with Salix (4%) increasing and Alnus decreasing to very low values (~1%).
Discussion
The Silver Lake record spans the Holocene with a basal date of ~11.7 cal ka BP (2σ range ~12.0–11.4 cal ka BP) on Picea cf. engelmannii needles. Owing to the short core length and slow sediment deposition rates, the record is relatively coarse in resolution. Individual samples through much of the core typically average ~100–150 years of deposition and are spaced ~500–750 years apart. The Silver Lake record is therefore suitable for examining broad, multi-century to millennial scale changes in vegetation history, but not for higher-resolution comparisons.
The Silver Lake record reveals broad transitions in the expansion and maturation of upper montane parklands and forests, with moderate and mostly gradual shifts among a few major taxa including Picea, Pinus, Artemisia, and Alnus. By the end of the YD, highlands around the lake supported an alpine-subalpine Artemisia steppe with open Pinus/Picea parkland under a cold dry climate, a pattern consistent with many regional upper montane as well as lowland records (e.g. Louderback and Rhode, 2009; Louderback et al., 2015; Lundeen and Brunelle, 2016; Madsen and Currey, 1979; Madsen et al., 2001; Mensing et al., 2012; Morris et al., 2015; Munroe, 2003; Rhode, 2016; Thompson et al., 2016). Beginning ~9.0 cal ka BP, Picea parkland began to expand at the expense of Artemisia, indicating rising treeline and development of mature Picea-dominated subalpine forest. We interpret the Picea expansion to reflect early Holocene summer warming coupled with greater winter snowpack, consistent with many other records including Snowbird Bog in neighboring Little Cottonwood Canyon (cf. Broughton et al., 2008; Lundeen and Brunelle, 2016; Lundeen et al., 2013; Madsen and Currey, 1979; Power et al., 2011; Thompson et al., 2016).
Picea forests dominated around Silver Lake from ~8.0 to 5.0 cal ka BP, at the expense of Pinus. Alnus also became established, presumably expanding up-canyon. Both indicators suggest warming conditions, especially after ~7.0 cal ka BP. If the Picea/Pinus ratio reflects increasing moisture, as previously suggested for Snowbird Bog (Madsen and Currey, 1979), then ~7.0–5.0 cal ka BP would be the moistest interval in both records. However, this ratio is also influenced by temperature (Maher, 1963), which can lead to conflicting interpretations (Fall, 1997). The current climate at Silver Lake favors Picea engelmannii with moister and slightly warmer winters than is tolerated by Pinus flexilis (Thompson et al., 2015). We suggest that mid-Holocene Picea dominance was favored by sustained high snowpack (augmented by spring-fall lake-effect storms; see below) as well as winter warming. Thompson et al. (2016: 274) suggested the Great Salt Lake pollen record indicated cooler and moister conditions after ~7.2 cal ka BP, with further cooling after ~6.1 cal ka BP, but noted that this reconstruction fits northern sites better than Great Basin or Rocky Mountains records.
The increase of Alnus and Artemisia and decline of Picea after ~5.0 cal ka BP suggests increased representation of lower-elevation taxa and opening of the Picea forest. We suspect that local factors, such as changes to Silver Lake’s inflow associated with the deposition of sedimentary unit III at this time, may underly the pollen representation of vegetation change identified here, but additional studies are needed to ascertain the cause. Regional paleoenvironmental reconstructions between ~6.0 and 4.0 cal ka BP are quite variable with cooling trends, warming trends, pervasive drying, or a combination (e.g. Lundeen et al., 2013; Lundeen and Brunelle, 2016; Madsen and Currey, 1979; Mensing et al., 2012; Morris et al., 2015; Shuman and Serravezza, 2017; Thompson et al., 1993, 2016). One or more of these trends may be contributory at Silver Lake, but none are entirely persuasive.
After ~3.5 cal ka BP Pinus increased and remained high, while Picea gradually increased to form a patchy mixed conifer parkland. This trend may signal a downward shift of Picea treeline, followed by subsequent re-expansion (Fall, 1997; Maher, 1963). Late-Holocene cooling is widely observed in regional records (e.g. Lundeen and Brunelle, 2016; Munroe, 2003; Munroe et al., 2013; Reinemann et al., 2009; Thompson, 1984; Thompson et al., 2016). In the Great Salt Lake basin, this period is marked by downward expansion of Juniperus woodlands between ~3.8 and 2.6 cal ka BP (Madsen et al., 2001; Thompson et al., 2016; Wigand and Rhode, 2002). The last ~2.5 cal ka BP in the Great Salt Lake pollen record show subtle complexity with alternating droughts and cooler/wetter episodes (Thompson et al., 2016: 279–281; cf. also Adams and Rhodes, 2019; Benson et al., 2002; Mensing et al., 2008, 2013; Millar et al., 2019; Stine, 1994; Wigand and Rhode, 2002). The Silver Lake record is not sufficiently high-resolution to capture these shifts, and instead shows broadly stable vegetation during this period.
The Silver Lake record suggests that upper montane Picea-dominated ecosystems in the Wasatch Range were broadly stable during the Holocene, responding moderately and gradually to large-scale climatic trends. While the Silver Lake record is relatively coarse-grained in its temporal resolution, it is consistent with many regional records in the timing of response to large-scale climate change. We suggest two local factors were especially important in the general stability of local vegetation communities. First, the Wasatch Range is subject to a pronounced lake effect climatology from Great Salt Lake, which effectively amplifies cold-season snowpack especially in fall and spring (Alcott and Steenburgh, 2013; Alcott et al., 2012; Carpenter, 1993; Hostetler et al., 1994; Mock, 1996; Steenburgh et al., 2000), differentiating the region climatologically (Shinker and Bartlein, 2010). Localized lake-effect precipitation coupled with orographic influences moderates warm season drought and, we suggest, helps account for long-term persistence of Picea in the Wasatch Range.
The second factor is the lack of competing lower montane pine species (Pinus contorta, P. ponderosa), which are rare to absent in the region (Allan, 1962). In other settings, Picea is often outcompeted by lower montane conifers under drier or warmer conditions; once replaced, Picea fails to re-establish as a dominant (e.g. Louderback et al., 2015). We suggest the lack of these conifer competitors contributed to the long-term stability of upper montane Picea populations in the Wasatch Range. Persistence of Picea forests under future climate scenarios may be favored by lack of competition from lower montane Pinus, though they may suffer other threats such as devastating insect infestations or altered fire regimes (Minckley et al., 2012; Morris et al., 2013, 2015).
Future climate change will likely result in complex and unexpected modifications to upper montane ecosystems in the intermountain West (Mensing et al., 2012; Morris et al., 2015; Shinker and Bartlein, 2010). Our Silver Lake record suggests that these ecosystems often respond consistently in timing to major regional climate changes but may vary in direction, magnitude, and rapidity of response based on local climatic drivers, ecological thresholds, biotic competitors or predators, and other factors. Management of these ecosystems must be attentive to local factors as well as larger-scale modeled expectations; local paleoenvironmental records provide an important tool to anticipate biotic responses to these factors.
Supplemental Material
sj-pdf-1-hol-10.1177_0959683620972761 – Supplemental material for Holocene subalpine forest-parkland dynamics in Big Cottonwood Canyon, Wasatch Mountains, Utah, USA
Supplemental material, sj-pdf-1-hol-10.1177_0959683620972761 for Holocene subalpine forest-parkland dynamics in Big Cottonwood Canyon, Wasatch Mountains, Utah, USA by David Rhode, Lisbeth A Louderback and Sandy O Brugger in The Holocene
Footnotes
Acknowledgements
Thanks to the reviewers whose suggestions materially improved our interpretations and presentation; any shortcomings remain our own.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: We thank Alpine Archaeological Consultants, Desert Research Institute, and Swiss National Science Foundation SNSF Postdoc.Mobility grant P2BEP2_188180 for funding. Jonathan Grant, Anthony Macharia, and David Page assisted in fieldwork.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.