
Abstract
During the Ceramic Age (500 BCE–1500 CE), Lesser Antilles rice rats (Tribe Oryzomyini) made up a significant portion of the diet of Caribbean islanders. Archaeological excavations across the archipelago resulted to the discovery of large quantities of remains from to these now extinct taxa. It offers a unique opportunity to investigate the past biogeography of this taxon of high cultural and ecological importance. We have studied 1140 first lower molars originating from 40 archaeological sites across eleven islands of the Lesser Antilles archipelago using two-dimensional geometric morphometric approaches to establish spatiotemporal patterns relying on phenotypic variations. This study identified three morphological groups, present in all chrono-cultural periods, that were geographically restricted and consistent with published ancient mitochondrial DNA clusters. These three geographically-separate groups likely represent three distinct genera of rice rats. The first group includes specimens from the North of the archipelago (Saint-Martin, Saba, Saint-Eustatius, Saint-Kitts, and Nevis) and likely referable to as Pennatomys sp.; the second, occurring in the South (Martinique), is assigned to Megalomys desmarestii; and the third corresponds to specimens from the center of the Lesser Antilles (Antigua, Barbuda, Marie-Galante, and Guadeloupe) and likely corresponds to Antillomys sp. These oryzomyine morphotypes are present during all studied periods and support an older presence of these rodents in the region. Our results are congruent with ancient DNA studies that favor the hypothesis of a natural introduction of the group in the archipelago before settlement of human populations. Moreover, the observed phenotypic homogeneity and stability over the 2000 years of Pre-Columbian occupation suggests that rice rats were not part of long-distance inter-island exchanges by humans. Instead, rice rat human consumption was likely based on in-situ hunting of local populations.
Keywords
Introduction
The Lesser Antilles are an oceanic archipelago in the Caribbean, located between two biogeographic features, the Anegada passage off the Anguilla bank (Jany et al., 1990), and Koopmans’ Line off the Grenada Bank (Genoways et al., 2010) (Figure 1). The Lesser Antilles show a low diversity of terrestrial organisms associated with a high rate of endemism common to many island ecosystems (e.g. Baker and Genoways, 1978; Bond, 1999; Hedges, 1999; Ricklefs and Bermingham, 2007) which are profoundly influenced by both modern and past human activity (Whittaker and Fernández-Palacios, 2007). The first attested human settlement in the Lesser Antilles dates to the fourth millennium BCE (Bonnissent et al., 2014) in Saint-Martin. Archaeological records confirm the ancient introduction and translocation of continental plants and animals that are still present today, including manioc (Manihot esculenta), maize (Zea mays), papaya (Carica papaya) (Newsom and Wing, 2004; Pagán Jiménez et al., 2005), dogs (Canis familiaris), and agouti (Dasyprocta sp.) (Bonnissent, 2008; Giovas et al., 2012, 2016; Wing, 2001a). Lesser Antillean rice rats (tribe Oryzomyini) are considered to be endemic to the archipelago, as the timing of their arrival in this area has been estimated via molecular clock analysis to the Late Miocene (6.814–6.303 Mya – Brace et al., 2015). However, the oldest secure evidence of rice rats recovered from the paleontological record is contemporaneous with the earliest human occupations of the archipelago (Steadman et al., 1984). Rice rats are abundant in archaeological assemblages and show clear evidence of consumption (cutting and burning marks) (Grouard, 2007), though are now extinct across the Lesser Antilles, with the last living specimen recorded during the mid-late 19th century (Allen, 1942; Ray, 1962). Traditional reasons for small mammal extinction on islands include the introduction of competing species (MacPhee and Flemming, 1999) such as rats (Rattus rattus and R. norvegicus) or new predators like the cat (Felis silvestris) (Henderson, 1992), the Small Indian mongoose (Herpestidae, Urva auropunctata) (Grouard, 2001; Henderson, 1992; Horst et al., 2001), and the raccoon (Procyonidae, Procyon lotor) (Louppe et al., in press), or overhunting (Ray, 1962; Steadman et al., 1984; Trouessart, 1885), along with the transformation of the landscapes and deforestation (Boudadi-Maligne et al., 2016). Yet the specific causes of rice rat extinction in the Lesser Antilles still remains unclear.

Map of the Lesser Antilles archipelago. The seven islands investigated are highlighted and followed by the number of archaeological rice rats (tribe Oryzomyini) teeth analyzed. The number of archaeological sites studied are in brackets. Information about the sites can be found in Table 1. Grey shadows indicate the geological banks exposed during the Pleistocene (Hedges, 2001; MacPhee and Flemming, 1999; Pregill et al., 1994). Dashed lines in the North mark the Anegada passage (Jany et al., 1990) and in the South Koopmans’ Line (Genoways et al., 2010).
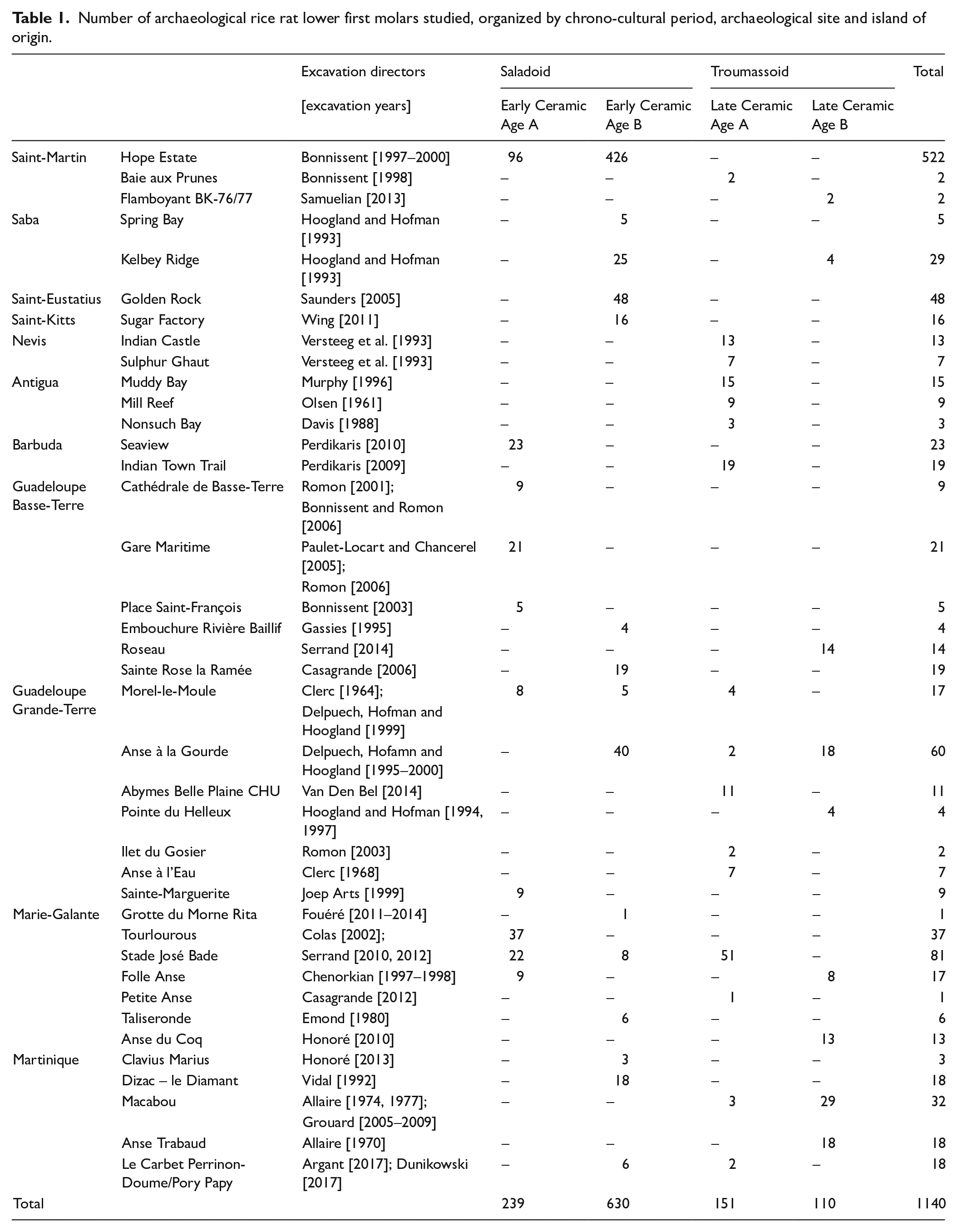
Number of archaeological rice rat lower first molars studied, organized by chrono-cultural period, archaeological site and island of origin.
Pre-Columbian societies relied primarily on marine resources such as fish and mollusks (Grouard, 2010; Wing and Wing, 1995) but terrestrial mammals, including oryzomyines, were also consumed as evidenced by the presence of butchery and burning marks on recovered remains (Grouard, 2004, 2010; Newsom and Wing, 2004; Wing, 2001a, 2001b). Rice rats were an important part of the human diet throughout the Ceramic Age. Analysis of zooarchaeological collection from the site of Hope Estate (Saint-Martin) show that rice rats comprise 54% of the total number of all identified vertebrates (NMI = 748/1384; Grouard, 2004). Extensive evidence for anthropic inter-island exchange in the Lesser Antilles has been evidenced for example dogs, humans, raw materials, and artefacts (i.e. Bonnissent, 2008, 2013; Fitzpatrick, 2015; Hofman et al., 2006, 2007, 2008; Hofman and Hoogland, 2011; Knippenberg, 2007; Laffoon et al., 2013, 2015, 2016; Stouvenot and Randrianasolo, 2013). It is still unclear whether rice rats were part of this network. Anthropic translocation has been proposed for the group (LeFebvre and deFrance, 2014) and examplified by the presence of South American rice rats of the genus Zygodontomys in the island of Carriacou (Giovas, 2018; Mistretta, 2019).
Rodents have colonized a remarkable number of islands worldwide, either by natural dispersal (e.g. Fabre et al., 2013; Jansa et al., 2006), as stowaway like the black rat (Rattus rattus; MacPhee and Flemming, 1999; Vigne and Valladas, 1996) or intentional transportation like the domestic guinea pig (Cavia porcellus; Kimura et al., 2016; LeFebvre and deFrance, 2014; Lord et al., 2018). Morphological similarities between rodent populations can be used to explore the type of dispersal (e.g. Cucchi et al., 2014; LeFebvre et al., 2019) along with the impact of environmental factors on their diversity (e.g. Maestri et al., 2018).
In this study we have used tooth identification as the basis of our analysis. Teeth have an advantage over other skeletal elements in that they are often well preserved, are recovered in large numbers in the archaeological record and known to be taxonomically informative (Cucchi, 2009; Darviche et al., 2006; Darviche and Orsini, 1982; Renaud et al., 1996; Van Dam, 1996). Consequently, teeth, especially their size and shape have been used to study past rodent populations and their relationship with human societies (e.g. Cucchi et al., 2014; Hulme-Beaman et al., 2018b; Valenzuela-Lamas et al., 2011).
Because Lesser Antilles rice rats have no direct modern relatives and because morphological evolution of insular populations can happen very fast (Millien, 2006) eventually leading to inter-island radiation (e.g. Kadmon and Allouche, 2007), the current systematic and taxonomy of the group is still not fully established. Three genera are currently recognized in the Lesser Antilles archipelago (Brace et al., 2015; Machado et al., 2014) (Figure 1): Pennatomys has been described in the North of the archipelago (Saint-Eustatius, Saint-Kitts, and Nevis) (Turvey et al., 2010), Megalomys (Trouessart, 1885) in the South (Martinique, Saint-Lucia, and Barbados) (Forsyth Major, 1901; Friant, 1941; Miljutin, 2010), and Antillomys (Brace et al., 2015) in the center (Guadeloupe, Antigua, and Barbuda) (Barbotin, 1970; Brace et al., 2015; Hopwood, 1926; Pregill et al., 1994). These three genera are supported by high genetic divergence (560 bp of the Cytochrome b gene) and according to ancient genetic data Megalomys and Pennatomys are sister clades, only distantly related to the genus Antillomys (Brace et al., 2015). The present study aims to assess the archaeobiogeography of the oryzomyines taxa in the Lesser Antilles during the Ceramic Age and explore whether spatio-temporal variation in their morphology can be explained through human translocation. Results were compared to published ancient DNA clusters in order to gain in the understanding on the group taxonomy.
Materials
Chrono-cultural division
The pre-Columbian archaeology of the Lesser Antilles is divided into two main periods: the Early and Late Ceramic Ages, each further divided into an early and a late phase. The Early Ceramic Age corresponds to the Saladoid cultures and it is divided into an early phase A (ECA) dating from c. 500 BCE to 400 CE (Early Cedrosan Saladoid and Huecan Saladoid), and a late phase B (ECB) from c. 400 to 600/800 CE (modified Cedrosan Saladoid, Late Saladoid, and Saladoid with Barrancoid influences) (Bérard, 2012; Fitzpatrick, 2015; Hofman et al., 2007, 2008). The Late Ceramic Age corresponds to the Troumassoid cultures and is divided into an A phase (LCA) dated from 600/800 to c. 1200 CE, and a B phase (LCB) dated from c. 1200 to 1500 CE. Contrary to the Early Ceramic, the two phases of Late Ceramic were characterized by a geographic division between the North and the South of the archipelago (Crock and Petersen, 2004; Mol, 2006; Rouse and Faber Morse, 1999; Siegel, 1989). The LCA is characterized by the Northern Troumassan Troumassoid and the Southern Suazan Troumassoid, while the LCB correspond to the Marmoran Troumassoid in the North and Troumassan Troumassoid in the South.
Studied specimens
A total of 1140 archaeological first lower molars (M1, either isolated or enclosed in the mandible) belonging to adult specimens with limited wear were analyzed. When teeth were not found isolated, only hemi-mandibles were recovered and could not be left and right paired to form complete jaws based on tooth wear abrasion or stratigraphic information, as a consequence it is possible that both the right and left M1 of some specimens were included in the analyses. Specimens originated from 40 archaeological sites spanning across the pre-Columbian Ceramic Age in eleven islands throughout the archipelago (Figure 1; Table 1). Specimens were assigned to chronological cultural phases based on primary publications and excavation reports, and according to investigators’ opinion (Table 1; Supplemental Material SI.1, available online).
Methods
Morphometrics
Photographs of the occlusal view of the lower first molar were taken using a LEICA 76 APO macroscope and a non-distorting objective, with a 1.25× or 1.6× magnification and Leica Microsystem LAS software (V4.8). A sliding semi-landmark based geometric morphometric approach was employed to quantify the size and the shape of the teeth. On each photograph, the two-dimensional coordinates of one landmark, placed at the junction between the most posterior point of the metaconid and the external buccal edge of the tooth were recorded, along with 65 equidistant sliding semi-landmarks localized along the external edge of the tooth (recorded clockwise) (Figure 2) using TPS Dig2 (Rohlf, 2004). All photographs and measurements were taken by the same person (M.D.). Coordinates were superimposed using a generalized Procrustes analysis (GPA) (Goodall, 1995; Rohlf and Slice, 1990), with the semi-landmarks allowed to slide following the Procrustes distance minimization criterion. Analyses of size were based on the log-transformed centroid size, and investigation of shape performed on the Procrustes residuals (coordinates after superimposition).
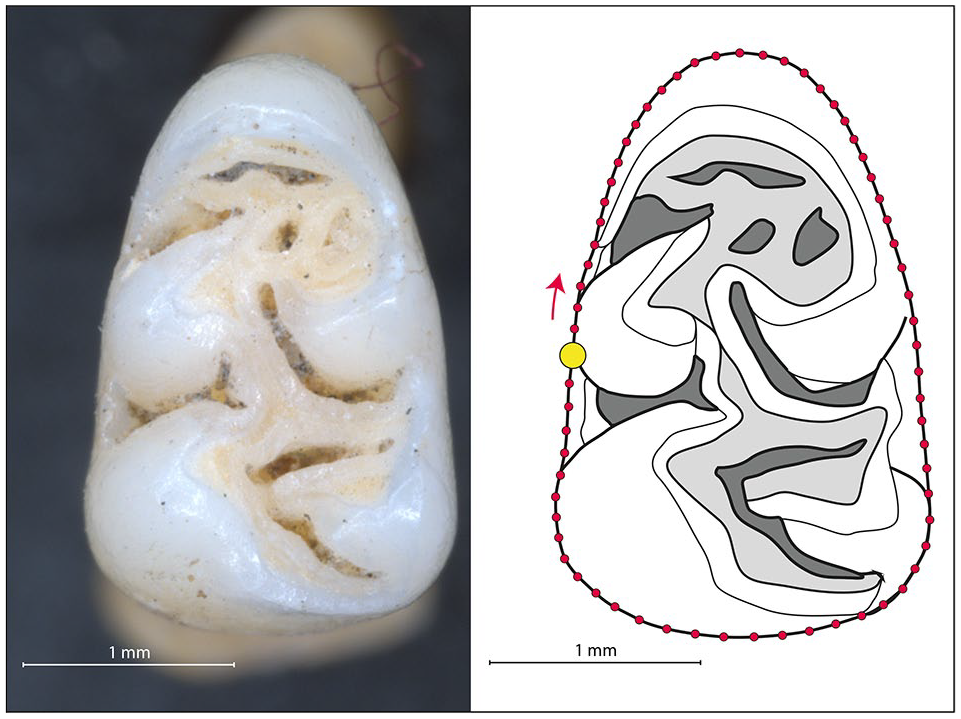
Left: Occlusal view of a first lower molar of a Lesser Antillean rice rat (specimen SRA-Guadeloupe-HE-089-D from Hope Estate, Saint-Martin). Right: Geometric morphometric protocol including one landmark (large yellow dot) and 65 sliding semi-landmarks along the outside curvature of the tooth, recorded in a clockwise direction.
Statistics
Prior to analyses, the existence of size or shape clusters at each archaeological site, island, island bank, and overall was examined using a Gaussian Mixture Modelling for Model-Based Clustering (R package “mclust,” Scrucca, 2016). This approach determines, based on a maximized loglikelihood approach, the number of clusters (i.e. groups) present in the dataset, without a priori knowledge.
Centroid size differences between populations from different sites, periods, islands, and island banks were tested with Kruskal-Wallis and Wilcoxon tests and visualized by boxplots. In pairwise comparisons, p-values were adjusted using a Benjamini-Hochberg multi-comparison procedure (Benjamini and Hochberg, 1995).
The shape variation was examined through the application of principal component analysis (PCA), before testing differences between groups through multivariate analysis of variance (MANOVA). Because of the large number of variables compared to the relatively low number of specimens per group, a dimensionality reduction of the data was performed (Baylac and Friess, 2005; Evin et al., 2013). Canonical variate analysis (CVA) and MANOVAs were performed on the firsts PCA scores that maximized the between-group discrimination quantified by leave-one-out cross validation (Baylac and Friess, 2005; Dobigny et al., 2002). CVA was used to quantify and visualize the group differences (Albrecht, 1980; Gittins, 1985; Russell et al., 2000). The discrimination power of the CVA was quantified by the mean cross-validation percentage (CVP) and the 90% confidence interval obtained from 100 CVAs performed on resampled same-size datasets (Evin et al., 2013). This approach randomly sub-samples the largest groups to the size of the smallest group, thus removing effects of unbalanced sample size which is common and largely inevitable in bioarchaeology (Evin et al., 2013). Cross-validation percentages were calculated for size, shape and form (size + shape). In addition to CVA, phenotypic dissimilarity between groups were assessed using both Mahalanobis and Procrustes distances. Distances were computed only for groups of at least 10 specimens and the repeatability of the topologies was estimated by bootstrap with 1000 replicates. Procrustes distances between group mean shape was obtained following Nagorsen and Cardini (2009) and Mahalanobis distances were obtained from resampled datasets with equal number of specimens per group. For both approaches the number of resampled specimens match the number of specimens in the smallest group. The majority-rule consensus and mean branch lengths topologies were computed as unrooted Neighbor-Joining (NJ) networks (Friesen et al., 2007; Saitou and Nei, 1987) on which the percentage of trees in which each observed node grouping appeared has been reported (i.e. bootstrap supports). All analyses were performed in R version 3.5.3 (R Core Team, 2019), with the “ape” (Paradis and Schliep, 2018), “Morpho” (Schlager, 2013) and “Geomorph” (Adams et al., 2018) packages.
Results
None of the clustering analysis on size and shape revealed the presence of multiple groups. As a consequence, the composition of each archaeological site was considered homogeneous. Despite the fact that the number of teeth per site and chrono-cultural period was maximized by including right and left teeth, statistical analyses were highly constrained by the number of specimens per site and chrono-cultural period. As a precaution, a subset of analyses were performed for teeth of only one side (left) of the mandible and provide congruent results though based on much less groups (for homogeneity groups with less than 10 specimens were excluded). Analyses performed in this study were constraint by available archaeological data which range from an island with only one site occupied during one period (e.g. Saint-Kitts) to islands occupied during all periods on multiple sites (e.g. Martinique).
Main archaeobiogeographic pattern
Analysis of the populations from each site and each cultural time period revealed three geographically structured clusters (Figures 3 and 4).
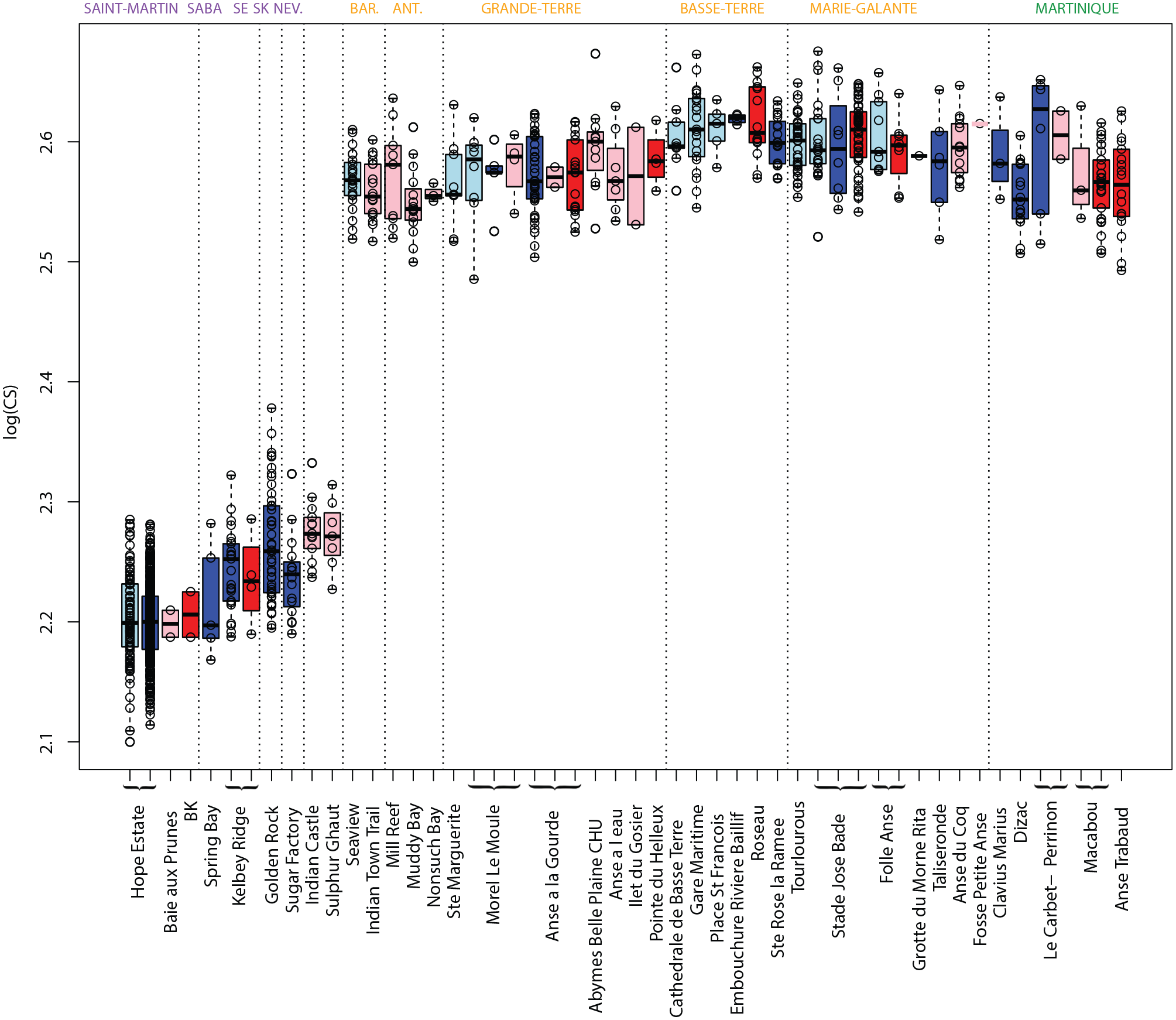
Size variation between rice rat populations of the Lesser Antilles. Boxplots of the log-transformed centroid size of the lower M1. Specimens were grouped by island, site and chronological occupation phase. In light blue: Early Ceramic Age A; dark blue: Early Ceramic Age B; pink: Late Ceramic Age A; red: Late Ceramic Age B. SE: Saint-Eustatius; SK: Saint-Kitts; NEV.: Nevis; ANT.: Antigua; BAR.: Barbuda. The islands names are colored according to the three shape clusters identified (Figure 4) (purple: Northern islands; yellow: Central islands; green: Southern island).
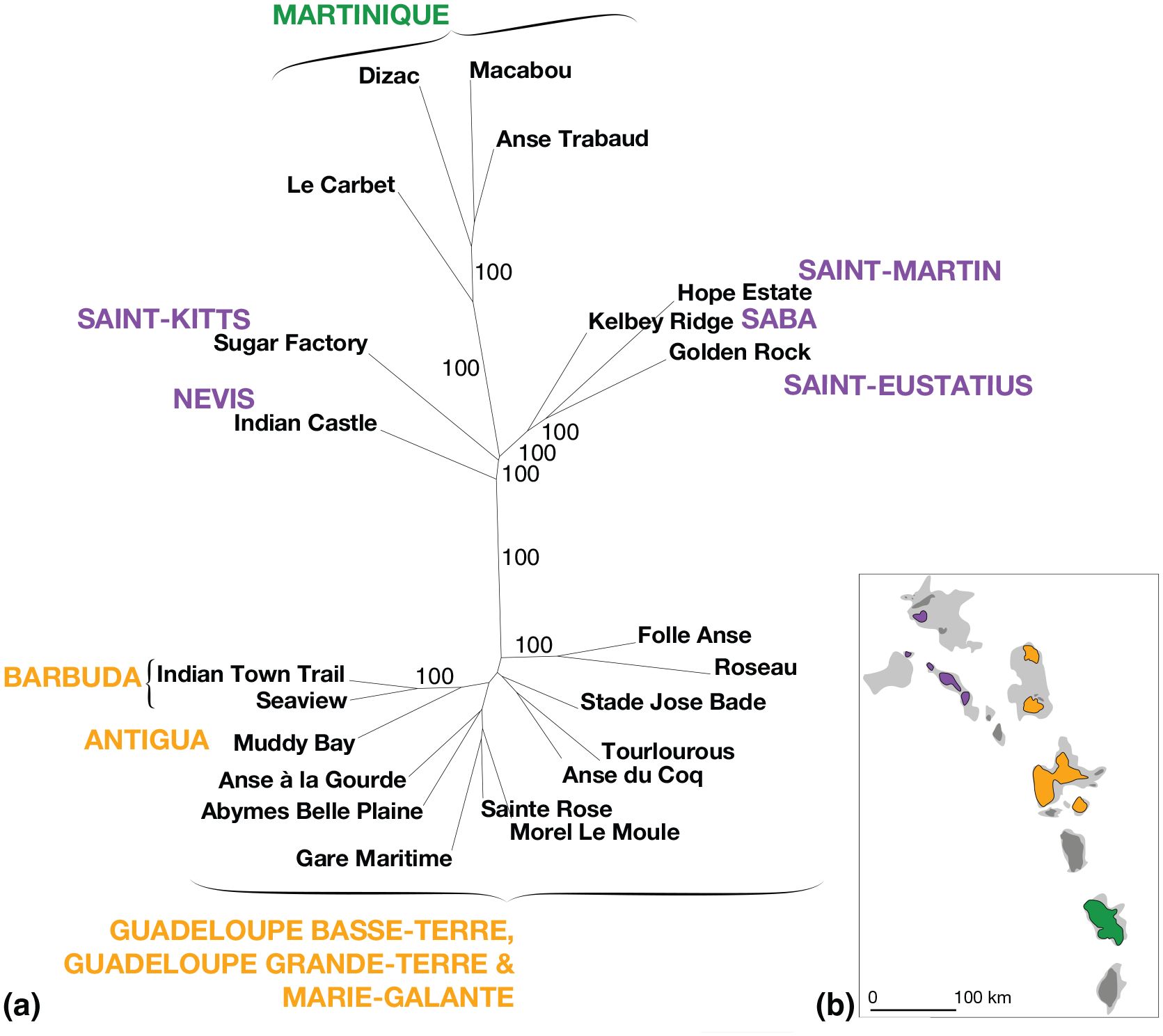
(a) Phenotypic relationship between sites across the Lesser Antilles. Mean branch lengths neighbor-joining network of the bootstraped Mahalanobis distances (1000 replicates) with mention of the bootstrap percentages above 50%. Only sites with more than 10 specimens were included. Island names are colored by their geographic and morphometrical cluster attribution (purple: Northern islands; yellow: Central islands; green: Southern islands). (b) Map of the studied islands colored by the three shape clusters identified.
Molar centroid size analysis revealed two main clusters, perfectly discriminated, with the specimens from the Northern islands (Saint-Martin, Saba, Saint-Eustatius, Saint-Kitts, and Nevis) showing much smaller teeth than any other specimens from across the archipelago (Figure 3).
Shape analysis revealed differences between the populations (F(46, 2238) = 105.91, p ⩽ 2.2e−16). While Procrustes distances do not provide a resolved network topology (most bootstrap values are <50%) (Supplemental Material SI.2, available online), Mahalanobis networks (Figure 4, Supplemental Material SI.2, available online) revealed three perfectly supported clusters (100% bootstrap values) that are geographically structured. The first cluster corresponds to the northern islands (Saint-Martin, Saba, Saint-Eustatius, Saint-Kitts, and Nevis), the second to the central islands (Barbuda, Antigua, Grande-Terre, and Basse-Terre of Guadeloupe, and Marie-Galante) while the third correspond the southern island of Martinique. However, when working at the island scale (Supplemental Material SI.2, available online) the split between Martinique and the central islands is not supported, but because of the relatively long length of the Martinique branch and the high size differences between the two geographic clusters (Figure 3) they were considered as valid morphometric and geographic entities. The paired leave-one-out cross validation between the three geographic clusters is high (94.4%; CI: 92.9–95.9%) confirming their morphometrical distinction. Moreover, when molar form is analyzed, combining size and shape data, this percentage reaches 99.7% (CI: 99.3–100%).
Regional spatio-temporal patterns
Variation within the three clusters was then investigated at different geographical and chronological scales by looking at the differences between island banks (during past low-stand sea levels occurrences), between islands and sites, and through the different cultural time periods.
The five northern Islands
Populations from the five northern islands differ in both size (X2 = 147.5, df = 4, p ⩽ 2.2e−16; CVP = 35% (CI: 28.8–43.8%); Supplemental Material SI.3.2, available online) and shape (F(44–2528) = 11.8, p ⩽ 2.2e−16; CVP = 51.1% (CI: 41.3–58.8%); Supplemental Material SI.3.1–2, available online) (Figure 5). The island structuring of the populations observed in the networks (Figures 4 and 5) is also evidenced by pairwise comparisons (Supplemental Material SI.3.2, available online). The differences between islands exceed the differences between cultural time periods despite the small number of populations available for comparison. Only Saint-Martin is represented by more than one period with sufficient specimens for a diachronic comparison. One site (Hope Estate) was occupied during the ECA and ECB, and the corresponding specimens differ only slightly in shape (F(4, 517) = 4, p = 3e−3; CVP = 55.7% (CI: 50.5–59.9%)) and not in size (W = 22,975, p = 0.3; CVP = 49.1% (CI: 46.8–53.2%)). On the other hand, differences between islands during the ECB are highly significant in both size (X2 = 106.3, df = 3, p ⩽ 2.2e−16; CVP = 39.7% (CI: 29.7–48.4%); Supplemental Material SI.3.2, available online) and shape (F(24–1533) = 15.5, p ⩽ 2.2e−16; CVP = 57.8% (CI: 40.9–65.7%); Supplemental Material SI.3.1–2, available online).

Phenotypic relationship between sites of the Northern islands. Mean branch lengths neighbor-joining network of the bootstraped Mahalanobis distances (1000 replicates) with mention of the bootstrap percentages above 50%. Site names are colored by chronological occupation phases (light blue: Early Ceramic A; dark blue: Early Ceramic B; pink: Late Ceramic A; red: Late Ceramic B). Only sites with more than 10 specimens were included.
Martinique
In Martinique, the populations differ in both size (X2 = 19.5, df = 5, p = 0.0015; CVP = 23.5% CI: 5.6–38.9%)) and shape (F(85, 355) = 2.2, p = 4e−7; CVP = 27.9% (CI: 11.1–44.4%)). Specimens from the Early and Late Ceramic differ in shape (F(14, 64) = 3.9, p = 8e−5; CVP = 71.7% (CI: 64.8–79.6%); Supplemental Material SI.4.1–2, available online), but not in size (W = 707, p = 1; CVP = 35.2% (CI: 3.6–51.9%), Supplemental Material SI.4.2, available online). The specimens from the ECB populations do not differ in size (X2 = 4, df = 2, p = 0.1) or shape (F(16, 36) = 1.7, p = 0.1). During the LCB, there was no observed size difference, (W = 257, p = 0.94), but shape varied (F(8, 38) = 2.6, p = 0.02; CVP = 64.5% (CI: 55.6–75%)).
The five central islands
The populations from the central islands differ in both size (X2 = 105.4, df = 4, p < 2.2e−16; CVP = 32.6% (CI: 28.1–37.8%)) and shape (F(100, 1536) = 4.4, p ⩽ 2.2e−16; 44.7% (CI: 38.5–51.2%)). Only populations from Barbuda and Guadeloupe Basse-Terre are clustered by islands (only one population was studied from Antigua) contrary to the populations from Grande-Terre and Marie-Galante (Figure 6).

Phenotypic relationship between sites of the central islands. Mean branch lengths neighbor-joining network of the bootstraped Mahalanobis distances (1000 replicates) with mention of the bootstrap percentages above 50%. Site names are colored by chronological occupation phases (light blue: Early Ceramic A; dark blue: Early Ceramic B; pink: Late Ceramic A; red: Late Ceramic B). M.G.: Marie-Galante; B.T.: Guadeloupe Basse-Terre; GT: Guadeloupe Grande-Terre. Only sites with more than 10 specimens were included.
Specimens from the two islands of the Barbuda bank (Antigua and Barbuda) differ from each other in shape (F(7, 61) = 3.1, p = 0.007; CVA = 65.6% (CI: 59.3–72.2%); Supplemental Material SI.5.1, available online) but not in size (W = 454, p = 0.2). Specimens from the three archeological sites from Antigua (Supplemental Material SI.5.1–2, available online), all attributed to the LCA, differ in shape (F(16, 36) = 2.1, p = 0.03; CVA = 41.4% (CI: 22.2–66.7%)) but not in size (X2 = 1.49, df = 2, p = 0.47). On Barbuda, the specimens from the two sites (one ECA, one LCA) do not differ in their molar size (W = 170, p-value = 0.2) nor shape (F(3, 38) = 0.3, p = 0.9).
Thirteen sites were analyzed from the Guadeloupe Bank (Basse-Terre + Grande-Terre, Table 1; Supplemental Material SI.5.1–2, available online). On average, specimens from Basse-Terre have larger molars than those from Grand-Terre (W = 6398, p = 5.4e−11, CVP = 71.1% (CI: 68.8–73.6%)) and differ slightly in shape (F(15, 169) = 4.2, p = 1e−6; CVP = 50.2% (CI: 41.6–58.4%). On Grande-Terre, the specimens from the different sites do not differ in size (X2 = 5.5, df = 6, p = 0.5), but differ in shape (F(78, 588) = 1.4, p = 0.02; CVP = 28.4% (CI: 14.3–50%)). During the ECA the populations do not differ in either size (W = 41, p = 0.7) or shape (F(5, 11) = 1.4, p = 0.3), nor do they differ during the LCA (size: X2 = 2.2, df = 4, p = 0.7; shape: F(36, 64) = 1.2, p = 0.2). However, specimens attributed to the Early and Late Ceramic differ in shape (F(6, 103) = 4.2, p = 0.0008; CVP = 67.8% (CI = 64.5–71.9%) but not in size (W = 1317, p = 0.3).
Similarly to the ones from Grande-Terre, Basse-Terre populations differ in shape (F(115, 240) = 1.4, p = 0.02; CVP = 22.8% (CI: 12.3–33.5%)), but not in size (X2 = 4.9, df = 5, p = 0.4).
Finally and again similarly, in Marie-Galante, the populations do not differ in size (X2 = 6.2, df = 9, p = 0.7) but differ in shape (F(66, 816) = 2.03, p = 6.1e−06); CVA = 24.6% (CI: 16.1–33.9%); Supplemental Material SI.5.1–2, available online). Specimens attributed to the Early and Late Ceramic differ in shape (F(12, 143) = 3.31, p = 0.0003; CVP = 63% (CI: 59.5–66.3%), but not in size (W = 2646, p = 0.2). We detect no differences between the ECA and ECB (size: X2 = 1.1, df = 1, p = 0.29; shape: F(16, 66) = 1.1, p = 0.4) whereas specimens differ in shape between the LCA and LCB (F(2, 70) = 5.5, p = 0.006; CVP = 65.1% (CI = 50–81.3%)), but not size (W = 323, p = 0.3; Supplemental Material SI.5.1–2, available online).
Discussion
Three morphometric clusters, corresponding to three biogeographic units, were identified from the molar size and shape variation: one in Martinique in the south, one in the center islands of the archipelago (Barbuda, Antigua, Guadeloupe, and Marie-Galante), and one in the Northern islands (including Saint-Martin, Saba, Saint-Eustatius, Saint-Kitts, and Nevis). Morphological distances between island cannot be explained by the geographic distances between them since some geographically close islands are distant morphometrically (e.g. Antigua (central cluster) is 83 km far from Nevis (northern cluster), and 106 km far from Basse-Terre (central cluster)).
This geographic variation greatly outweighs temporal variation, with only slight differences in molar morphology linked to cultural time periods within each geographical cluster. Our results thus support long term biogeographic patterning of morphological variation over the span of ca. 2000 years.
Morphological diversity and rice rat taxonomy
By only working on the first lower molars only a small portion of the entire phenotype of the organism is quantified. However, teeth are the most abundant rodents remains in archeological or paleontological sites, and they are widely used to infer the systematic, taxonomy and spatio-temporal patterns in this group (e.g. Hulme-Beaman et al., 2018a; Stoetzel et al., 2017). Tooth morphology is the result of both an adaptive and a heritability component (Bader, 1965; Polly and Mock, 2018; Renaud et al., 2006). Given the absence of genetic and morphometric data on exactly the same specimens, which make it impossible to draw definitive conclusions on the taxonomy of rice rats, the strong biogeographic patterning observed in tooth morphology could nonetheless be used to infer the diversity of Antillean rice rats and discuss taxonomical attributions. Our results reveal three distinct morphotypes likely corresponding to the three genera previously described in earlier morphometric and ancient DNA research (Brace et al., 2015; Turvey et al., 2010). The Martinican archaeological specimens (89 specimens) likely correspond to Megalomys desmarestii (Fischer, 1829), the only species described from the island (Friant, 1941; Trouessart, 1885) and for which only four specimens have been molecularly analyzed (Brace et al., 2015). Megalomys luciae (Forsyth Major, 1901) specimens from Sainte-Lucia were not included in this study. However, specimens from Barbuda, where Megalomys audreyae was previously described (Hopwood, 1926), were found to be morphologically closer to specimens from the central islands, including Grande-Terre and Antigua, where Antillomys rayi was genetically identified (Brace et al., 2015). The slight morphometric differences revealed by our analyses (407 specimens) between the central islands may correspond to intra-specific variation within Antillomys rayi although the possibility that our sample includes several distinct species (with morphometrically closely similar first lower molars) could not be excluded. Finally, the third cluster including specimens from the Northern islands (644 specimens) likely corresponds to Pennatomys, with Pennatomys nivalis described from the Saint-Kitts Bank (Turvey et al., 2010). Brace et al. (2015), based on the Cytochrome b mitochondrial gene, observed some genetic differentiation between islands across the Saint-Kitts Bank, with 2% of genetic divergence between specimens from Nevis and Saint-Kitts, and 5–7% divergence between these specimens and those from Saint-Eustatius. These molecular analyses are in perfect agreement with our morphometrical data (specimens from Nevis and Saint-Kitts being morphological closer than those from Saint-Eustatius) and suggest intraspecific or inter-specific variation. Our results suggest that the two geographically-distant island groupings, Martinique in the South and the Northern islands, are phenotypically more similar to each other, than to those of the geographical closer central islands. This is consistent with molecular data showing a sister relationship between Megalomys and Pennatomys, while Antillomys is more distantly related (Brace et al., 2015). Further research on this insular group taxonomy should seek to combine morphometric analysis with ancient DNA of the same specimens, in order to confirm the taxonomy of all investigated populations.
Influence of ecological factors within clusters
Rice rats from the Northern islands had very small molar size compared to the rest of the archipelago. A similar pattern has been observed in the Anolis lizards from Saint-Martin which particularly small size has been linked to the presence of competition (Brown and Wilson, 1956; Losos, 2000; Losos and Ricklefs, 2009; Roughgarden, 1995). Accordingly, the size of the oryzomyines from Saint-Martin and the other northern islands might be caused by the presence of a competitor species (e.g. Amblyrhiza inundata identified in the Anguilla bank during the Pleistocene; McFarlane et al., 2014) or by the existence of peculiar, but yet to be determined, local conditions which impacted species size on these islands.
In the present study comparison between chrono-cultural periods was greatly limited by the number of archaeological sites available for each island and period. Despite these limitations, we detected some differences in shape, but not in size, between diachronic populations, more especially between the Early and Late Ceramic in Martinique, Marie-Galante, and Guadeloupe Grande-Terre. Diachronic differentiation between archaeological rodent populations have been identified, in other species, using a similar methodology (e.g. Cucchi et al., 2014; Hulme-Beaman et al., 2018b) and has been interpreted as the result of a variety of causes including human landscape modification, climatic change, or a combination of both causes.
In the case of oryzomyine, their past and current distribution ranges on the South American continent indicates that climatic change has affected their distribution (Vázquez-Domínguez et al., 2020; Vickery et al., 2016). In the Lesser Antilles archipelago, a climatic change is registered around 900–1000 CE, and coincide to the transition between the Early and Late Ceramic Age. This change is linked with a transformation from a wet to a dryer and stormier climate across the archipelago (Beets et al., 2006; Bertran et al., 2004). In between these two periods, Pre-Columbian populations also experienced societal changes, identifiable in material culture and habitat (Fitzpatrick, 2015; Hofman et al., 2007; Siegel et al., 2015) and bioarchaeological studies point to a transformation in the management and consumption of resources (Grouard, 2004, 2007, 2010; Wing, 2001a).
If these elements are to be considered, further investigations are required to define the impact of ecological factors on insular rice rat morphology, which is particularly challenging when dealing with extinct taxa.
No strong evidence for human-mediated dispersal
A proposed northward river-based connection between the Amazon basin and the Caribbean Sea during the late Miocene (Wilkinson et al., 2010) may have facilitated broadly simultaneous overwater dispersal of many South American mainland vertebrates to the Lesser Antilles (review in Brace et al., 2015). For instance, the genus Leptodactylus (Anura, Leptodactylidae) is considered to have colonized the Lesser Antilles by dispersal across water in the mid-Cenozoic (Camargo et al., 2009; Hedges and Heinicke, 2007). According to genetic data the multiple colonization events of oryzomyine in the archipelago date to the same period (late Miocene) (Brace et al., 2015), despite their absence in the paleontological records (Grouard, 2015). Our results show only slight phenotypic changes through archaeological chrono-cultural periods, between 500 BCE and 1500 CE, as well as the continuous presence of three clearly defined geographic clusters supporting the hypothesis of an ancient arrival of the group in the archipelago.
Prior to their extinction, the rice rats of the Lesser Antilles lived close to human settlements and crop areas (Allen, 1942; Pinchon, 1967; Trouessart, 1885; Wing, 2001b). This tendency to commensalism was probably established since the first human occupation in the archipelago. Indeed, zooarchaeological studies highlight their importance in the subsistence economy of local human population throughout the entire Ceramic Age (Grouard, 2004, 2010; Wing, 2001b) and thus long-term interactions between these rodents and the Pre-Columbian populations. On the other hand, in the north of the archipelago the morphometric variation is well structured per island, a pattern that favor a hypothesis of no (or at least not strong) inflow from external rice rats populations that would have disrupted or smoothed out the inter-island phenotypic structure. In the center, the differences between island phenotypes is less structured, and the possibility of inter-island connections cannot be completely excluded. However, study of additional specimens and archaeological sites would be necessary to confirm these hypotheses. As the prevailing morphometric variation is geographically structured through time we suggest an absence of long-distance connection between islands from the different clusters, and thus an absence of human transport of oryzomyines from one island to another.
Moreover, rice rats fail to meet 5 of the 10 criteria proposed for classifying species as impacted by humanly-mediated translocations (Giovas, 2019; Heinsohn, 2003, 2010), and two of them could not be applied to this study. Even though the presence of oryzomyines is lacking in the paleontological and archaeological faunal records (criteria 2) prior to the Ceramic Age where they abruptly appear (criteria 3) (Grouard, 2015), they likely colonized the Lesser Antilles during the late Miocene (Brace et al., 2015). Rice rats show commensal tendency (criterion 5) on the continent (Guilday, 1972; Guilday and Parmalee, 1965; Vickery et al., 2016) and are found in zooarchaeological contexts in association with a plethora of naturally dispersed species (criterion 8) consumed by humans (Grouard, 2004, 2010; Newsom and Wing, 2004; Wing, 2001b), as well as with the agouti, another rodent potentially introduced (Allen, 1942; Newsom and Wing, 2004) and managed (Govoni and Fielding, 2001; Hardouin, 1995) (criterion 7). At the scale of the archipelago, their distribution is wide (criterion 4), their phenotypic and molecular differentiation is geographically cohesive (criterion 1) and inter-island differences persist through time (criterion 6, applied here to morphometric data). Finally, the criteria based on comparisons with historically documented species introduction (criterion 9) and diachronic reconstruction of invasion frontiers (criterion 10) cannot be applied due to data limitation. Overall, the evidence from previous analyses alongside our current research favors a non-anthropic dispersion of the group into the archipelago, or if contacts have existed, newly human-introduced populations from one island to another have not persisted and were not detected.
Conclusion
This study significantly contributes to our knowledge and understanding of the past diversity of the tribe Oryzomyini in the Lesser Antilles archipelago. Over a thousand specimens of these now extinct taxa were analyzed using geometric morphometrics, allowing the first exploration of their archaeobiogeography. The rice rats show diverse but very homogenous molar morphotypes defined by geographic parameters, which remain largely stable for the 2000 years of the Ceramic Age in the Lesser Antilles archipelago. The persistence of this strong archaeobiogeographical phenotypic pattern throughout the Ceramic Age supports a scenario of a pre-human dispersal of Oryzomyini species. While rice rats were a component of human diet throughout the Ceramic Age, our overall results do not support an initial hypothesis of human-mediated transport of rice rats between and within the three geographic entities, despite the evidences of transportation of cultivated plants, domesticated animals and raw materials. Additional studies combining geometric morphometric and ancient DNA analyses on the exact same specimens would allow for the consolidation of the archaeobiogeography, taxonomy and systematics of these extinct endemic rodents, and better identify the environmental adaptations that might have driven the establishment of rice rat populations across the archipelago.
Supplemental Material
sj-pdf-1-hol-10.1177_0959683620972785 – Supplemental material for Archaeobiogeography of extinct rice rats (Oryzomyini) in the Lesser Antilles during the Ceramic Age (500 BCE–1500 CE)
Supplemental material, sj-pdf-1-hol-10.1177_0959683620972785 for Archaeobiogeography of extinct rice rats (Oryzomyini) in the Lesser Antilles during the Ceramic Age (500 BCE–1500 CE) by Marine Durocher, Violaine Nicolas, Sophia Perdikaris, Dominique Bonnissent, Gwenola Robert, Karyne Debue, Allowen Evin and Sandrine Grouard in The Holocene
Supplemental Material
sj-pdf-2-hol-10.1177_0959683620972785 – Supplemental material for Archaeobiogeography of extinct rice rats (Oryzomyini) in the Lesser Antilles during the Ceramic Age (500 BCE–1500 CE)
Supplemental material, sj-pdf-2-hol-10.1177_0959683620972785 for Archaeobiogeography of extinct rice rats (Oryzomyini) in the Lesser Antilles during the Ceramic Age (500 BCE–1500 CE) by Marine Durocher, Violaine Nicolas, Sophia Perdikaris, Dominique Bonnissent, Gwenola Robert, Karyne Debue, Allowen Evin and Sandrine Grouard in The Holocene
Supplemental Material
sj-pdf-3-hol-10.1177_0959683620972785 – Supplemental material for Archaeobiogeography of extinct rice rats (Oryzomyini) in the Lesser Antilles during the Ceramic Age (500 BCE–1500 CE)
Supplemental material, sj-pdf-3-hol-10.1177_0959683620972785 for Archaeobiogeography of extinct rice rats (Oryzomyini) in the Lesser Antilles during the Ceramic Age (500 BCE–1500 CE) by Marine Durocher, Violaine Nicolas, Sophia Perdikaris, Dominique Bonnissent, Gwenola Robert, Karyne Debue, Allowen Evin and Sandrine Grouard in The Holocene
Supplemental Material
sj-pdf-4-hol-10.1177_0959683620972785 – Supplemental material for Archaeobiogeography of extinct rice rats (Oryzomyini) in the Lesser Antilles during the Ceramic Age (500 BCE–1500 CE)
Supplemental material, sj-pdf-4-hol-10.1177_0959683620972785 for Archaeobiogeography of extinct rice rats (Oryzomyini) in the Lesser Antilles during the Ceramic Age (500 BCE–1500 CE) by Marine Durocher, Violaine Nicolas, Sophia Perdikaris, Dominique Bonnissent, Gwenola Robert, Karyne Debue, Allowen Evin and Sandrine Grouard in The Holocene
Supplemental Material
sj-pdf-5-hol-10.1177_0959683620972785 – Supplemental material for Archaeobiogeography of extinct rice rats (Oryzomyini) in the Lesser Antilles during the Ceramic Age (500 BCE–1500 CE)
Supplemental material, sj-pdf-5-hol-10.1177_0959683620972785 for Archaeobiogeography of extinct rice rats (Oryzomyini) in the Lesser Antilles during the Ceramic Age (500 BCE–1500 CE) by Marine Durocher, Violaine Nicolas, Sophia Perdikaris, Dominique Bonnissent, Gwenola Robert, Karyne Debue, Allowen Evin and Sandrine Grouard in The Holocene
Supplemental Material
sj-pdf-6-hol-10.1177_0959683620972785 – Supplemental material for Archaeobiogeography of extinct rice rats (Oryzomyini) in the Lesser Antilles during the Ceramic Age (500 BCE–1500 CE)
Supplemental material, sj-pdf-6-hol-10.1177_0959683620972785 for Archaeobiogeography of extinct rice rats (Oryzomyini) in the Lesser Antilles during the Ceramic Age (500 BCE–1500 CE) by Marine Durocher, Violaine Nicolas, Sophia Perdikaris, Dominique Bonnissent, Gwenola Robert, Karyne Debue, Allowen Evin and Sandrine Grouard in The Holocene
Supplemental Material
sj-pdf-7-hol-10.1177_0959683620972785 – Supplemental material for Archaeobiogeography of extinct rice rats (Oryzomyini) in the Lesser Antilles during the Ceramic Age (500 BCE–1500 CE)
Supplemental material, sj-pdf-7-hol-10.1177_0959683620972785 for Archaeobiogeography of extinct rice rats (Oryzomyini) in the Lesser Antilles during the Ceramic Age (500 BCE–1500 CE) by Marine Durocher, Violaine Nicolas, Sophia Perdikaris, Dominique Bonnissent, Gwenola Robert, Karyne Debue, Allowen Evin and Sandrine Grouard in The Holocene
Supplemental Material
sj-xlsx-1-hol-10.1177_0959683620972785 – Supplemental material for Archaeobiogeography of extinct rice rats (Oryzomyini) in the Lesser Antilles during the Ceramic Age (500 BCE–1500 CE)
Supplemental material, sj-xlsx-1-hol-10.1177_0959683620972785 for Archaeobiogeography of extinct rice rats (Oryzomyini) in the Lesser Antilles during the Ceramic Age (500 BCE–1500 CE) by Marine Durocher, Violaine Nicolas, Sophia Perdikaris, Dominique Bonnissent, Gwenola Robert, Karyne Debue, Allowen Evin and Sandrine Grouard in The Holocene
Footnotes
Acknowledgements
We would like to acknowledge the Guadeloupe (including Saint-Martin) and Martinique Departments of Archaeology (Regional Office of Cultural Affairs, French Ministry of Culture), Antigua and Barbuda National Parks and the Barbuda Research Complex (BRC), and the Florida Museum of Natural History in Gainesville (FLMNH) who provided the archaeological specimens studied. We acknowledge the morphometric platform of the MNHN, Paris (UMS OMSI). We would also like to acknowledge Alex Chepstow-Lusty for language editing, and Carly Ameen and Ashleigh Haruda, as well as Pierre-Henri Fabre and Pierre-Olivier Antoine from the GAARAnti project (ANR-17-CE31-0009) for discussions and suggestions. We also warmly thank Andrea Cardini and all the anonymous reviewers who gave helpful comments on earlier versions of the manuscript.
Author contributions
AE and SG conceived the ideas; DB, KD, SG, SP, and GR conducted the fieldwork and collected the material, KD and SG identified the archaeological bones; MD acquired the data; MD and AE performed the analyses; MD, AE, SG, and VN interpreted the results and MD led the writing with assistance from AE, SG, and VN.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Financial support for this research was granted by the ATM Blanche program of the MNHN, and the PEPS ECOMOB grant of the CNRS, INEE.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.