
Abstract
Over the last few decades – at a range of northern sites – changes in tree-ring width and latewood density have not followed mean summertime temperature fluctuations. This discrepancy sharply contrasts an earlier correlation between those variables. As the origin of this inconsistency has not been fully deciphered, questions have emerged regarding the use of tree-ring width and latewood density as a proxy in dendrochronological climate reconstructions. I suggest that temperature is no longer the most limiting factor in certain boreal areas, which might explain the observed divergence.
Keywords
Tree-ring width and latewood density originating from trees in northern boreal forests have been used for decades as proxies for local fluctuations in summertime air temperature (Briffa et al., 1998; D’Arrigo et al., 2014). These records have the main advantages of being well distributed on land, having an annual resolution and going back several centuries (D’Arrigo et al., 2014). While a range of studies are still able to correctly implement this approach (Anchukaitis et al., 2013, 2017; Briffa et al., 2013; Büntgen et al., 2008), others were unable to perform this feat as they found that these proxies started to diverge from the actual temperature measurements during the second half of the 20th century (Briffa et al., 1998). This observed reduction in correlation between temperature fluctuations and tree growth could in some cases be assigned to a methodological bias, such as “end effects” and a large distance between tree locations and meteorological sites (D’Arrigo et al., 2008; Esper and Frank, 2009). In other studies, these observations remain unresolved (D’Arrigo et al., 2008), indicating that the response of boreal forests to temperature could be more complex than previously thought (D’Arrigo et al., 2014).
While a short divergence has been recorded early in the 20th century (Schneider et al., 2014), no large divergence event has been observed in the 600 years of dendroclimatic data prior to 1950 (Esper and Frank, 2009; Tingley and Huybers, 2013). However, even though one of the longest temperature records ranges from 1659 (Manley, 1974) up to the present, more reliable global temperature measurements only go back to 1850 (Brohan et al., 2006). As a result, an undetected, large divergence during that period cannot be fully discarded (Stine and Huybers, 2014), but is unlikely (Cook et al., 2004), making the currently observed divergence problem (DP) puzzling.
Over the years, several hypotheses that could elucidate the DP have been proposed, such as an increasing competition with other plants and herbivory insects, higher UV-B levels, changes in atmospheric CO2 concentrations, increases in acid depositions and tropospheric ozone (Briffa et al., 1998), arctic dimming (Kirdyanov et al., 2020) and temperature-induced summer droughts (Barber et al., 2000; D’Arrigo et al., 2014). While all aforementioned factors are able to influence tree growth (Barber et al., 2000; Briffa et al., 1998; D’Arrigo et al., 2014; Kirdyanov et al., 2020), their individual effects are not necessarily substantial (Jacoby and D’Arrigo, 1997). It is difficult to identify the exact cause of this divergence in a given are as it is likely that several covarying environmental factors lie at the basis of this problem (Wilson et al., 2007). For example, when temperature increases, nitrogen cycling could increase (Litton et al., 2020) or moisture stress might occur due to an increase in temperature driven evapotranspiration without an increase in precipitation (Babst et al., 2019; Barber et al., 2000; Jacoby and D’Arrigo, 1995, 1997), both affecting tree growth more than would be expected by fluctuations in temperature alone. Furthermore, the effect of multiple covarying stressors, such as drought and a temperature increase, might not simply be additive but exert an interaction effect (Correia et al., 2018). This tight coupling between different environmental factors makes it hard to disentangle the effect of individual driving factors for tree growth.
Liebig’s law of the minimum states that plant growth is limited by the most deficient environmental resource available, i.e., the most limiting factor (Liebig, 1840; Sprengel, 1828). This implies that only a limiting environmental factor can be inferred from historical tree growth (Stine and Huybers, 2017). As such, when scientists aim for dendrochronological summertime temperature reconstructions, they should only sample trees located in temperature limited forests (Esper and Frank, 2009). In most northern boreal forests, it is still assumed that temperature is the most limiting factor for tree growth, despite a globally observed loss of about 8.7 million km2 in temperature-limited tree growth area, predominantly located in North-America and Asia (Anchukaitis et al., 2017; Babst et al., 2019). Recent divergence or decoupling between tree growth and mean summertime temperature further emphasizes that the assumption of temperature being the most limiting factor for tree growth might no longer be valid, especially as northern latitudes are more sensitive to climate change driven temperature increases compared to other areas (D’Arrigo et al., 2014) due to polar amplification. As a result, it can be concluded that other factors than summertime temperature have – at least partially – been driving recent fluctuations in tree growth. Even though many forests are still responding positively to recent temperature changes, it has been stated that boreal forests are currently more limited in growth by nutrient deficiency (Jarvis and Linder, 2000) or a combination of light availability and temperature (Stine and Huybers, 2014) than mean summertime temperature alone.
There are two ways by which temperature can lose its function as most limiting factor, either (i) temperature increases, making it no longer the most limiting factor, or (ii) another environmental factor, such as light, nutrients or water availability, becomes more limiting. If the DP results from a climate change driven temperature increase, it might be concluded that the DP is anthropologically driven, explaining why it has restricted itself to most recent decades (D’Arrigo et al., 2014). Divergence as a result of temperature losing its function as most limiting factor can be investigated by plotting tree growth as a function of mean summertime temperature for a given species in a given area (Figure 1). If this plot illustrates a strong correlation between tree growth and temperature for lower temperatures, which is absent for higher temperatures, temperature has lost its function as most limiting factor and divergence is most likely the result of a temperature increase. In other words, another factor (e.g. light availability or water supply) can become more restricting when growth rate increases due to a temperature increase, thus effectively disabling temperature’s function as most limiting factor. This reduced responsiveness of tree growth to temperature fluctuations during warmer periods (Figure 1) can already be observed in published data (e.g. Figures 2 and 3 in Jevšenak and Levanič (2016)). As the environmental needs of different species vary, this model can shift to higher or lower temperature ranges; for example, while one species obtains a growth rate that is limited by light availability at 15°C in a given area, another species may have a lower light dependency and is only limited by the same amount of light at a growth rate obtained at 20°C. Even though the point at which temperature loses its function as most limiting factor differs, the general model underlying this hypothesis remains applicable.
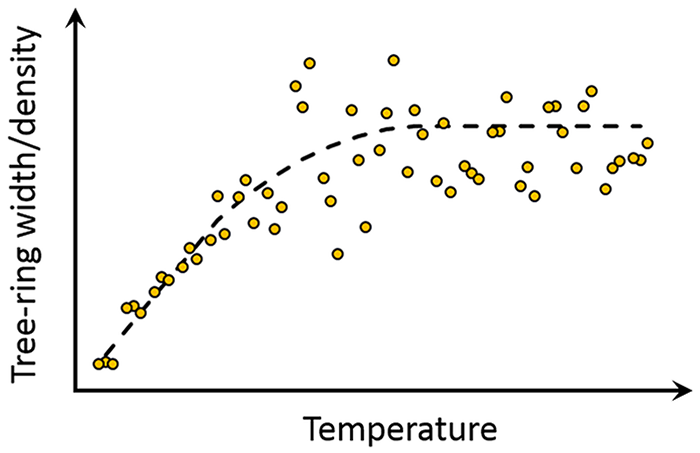
Hypothetical trend for tree-ring width and latewood density as a function of mean summertime temperature, illustrating a decoupling between both variables when mean summertime temperature increases.
A partial solution for temperature losing its function as most limiting factor lies in an adaptation of the currently used methodology. The classical approach for historical temperature reconstructions is based on averaged standardized tree-ring records, which only makes sense if tree growth is subjected to one single most limiting factor (temperature in the context of this paper) and noise, thus relying on Liebig’s law of the minimum (Stine, 2019; Stine and Huybers, 2017). However, additional local stressors for tree growth have been widely acknowledged (Fritts, 1976). As a result, when identifying trees that mimic temperature variability, temperature stress has to be sufficiently high, relative to other stressors, to invoke temperature-limited growth (Stine, 2019). When additional stressors intermittently limit growth, this approach based on averaged standardized tree-ring records should not be used (Stine, 2019; Stine and Huybers, 2017). While Barber et al. (2000) attempted to resolve this problem by calculating a climate index which depends on both temperature and precipitation during two consecutive years, Stine and Huybers (2017) took a more general approach by using a tree growth model which splits environmental stressors in a common growth factor (CGF; temperature in the context of this paper) and a local growth factor (LGF; any stressor at a scale smaller than the stand level), allowing one to overtake the other depending on climatic conditions (Stine and Huybers, 2017). In other words, this model allows for a change in most limiting factor (CGF or LGF), based on small scale environmental fluctuations. Applying this approach illustrated that records indicating less growth (smaller percentiles) were more likely to reflect a LGF, suggesting a preference for higher percentiles (i.e. faster growing trees) when attempting to reconstruct the CGF, for example, changes in mean summertime temperature in temperature limited areas (Barber et al., 2000; Stine and Huybers, 2017). These findings coincide with Cade et al. (1999) and Schröder et al. (2005), which suggest that higher percentiles are better suited to assess a CGF as they contain a stronger signal than mean reconstructions, especially when a large number of samples is available (Stine, 2019). This CGF-LGF based approach could significantly reduce the observed loss in correlation between temperature fluctuations and tree growth, i.e., DP. Stine and Huybers (2017), for example, were able to recover climate signals in 35 sites of their dataset that showed no signal when using averaged standardized tree-ring records by using this model. However, if temperature keeps increasing, other stressors will keep intermittently overtaking temperature as limiting factor for growth, reducing tree growth’s dependency on temperature until temperature completely uses its function as most limiting factor. In such cases, it becomes more likely for changes in a LGF (e.g. water availability) to limit growth rather than temperature, making a former LGF the new CGF.
Dendrochronological data can be used to assess changes in tree productivity throughout time, but might not be univocal when addressing historical fluctuations in mean summertime temperature, especially on a local or regional scale (Wilson et al., 2007). As a result, tree growth data expressing an unresolved, locally based DP should not be used in large composite databases, unless the observed DP can be identified as unique for this period (Wilson et al., 2016). Historical data inferred from tree ring width and latewood density measurements are ecophysiologically driven, allowing multiple environmental factors to influence these records. Questions regarding the validity of using dendrochronological data to reconstruct historical temperature fluctuations are valid as the most limiting factor – which is not necessarily temperature – has the strongest influence on tree growth. In order to unravel the causes for the DP, I suggest future research to focus on why temperature no longer is the most limiting factor for tree growth (i.e. (i) has temperature increased or (ii) has another environmental factor become more limiting) in a specific area and which environmental factors (e.g. water supply or light availability) has taken its place, potentially by using a model-based approach.