
Abstract
Integration of palaeobotanical (spores, pollen, phytoliths and non-pollen palynomorphs) and organic geochemical proxies, such as stable isotopes of organic carbon (δ13C) and n-alkanes, for studying the evolution and palaeoenvironmental conditions of an archaeological site are rare in India. The evolution of a protohistoric-historic site at Erenda, situated in the eastern coastal region of India, has been studied by using multiple palaeobotanical and organic geochemical proxies assisted with AMS radiocarbon dates. The excavated site lies above Late Pleistocene–Early Holocene Sijua Formation. The absence of anthropogenic evidence in the Sijua Formation likely indicates inhabitable conditions in nearshore/estuarine marshy conditions. The earliest human settlements at the excavation site begin during the first millennium BCE after the initiation of habitable conditions along the coast. The presence of fungal spores and the dominance of C4 phytolith morphotypes indicate prevailing warm and humid climatic conditions and proximity to a freshwater body. The δ13C signature and n-alkane composition indicate the use of C4 grass for the construction of the mud and clay-built huts. The settlers most likely used to consume wild or domestic variety of rice, as evidenced by the presence of bilobate scooped morphotypes. The site was partly abandoned, covered with C3 and C4 vegetation and used as a dumping ground after 663 ± 92 BCE. This implies that people continued to live in the area but possibly moved to a nearby site while using the excavated site as refuse.
Keywords
Introduction
The proto-historic phase is a cultural phase that connects prehistory to the historical period with a major shift towards the settled way of life in a pre-literate context. In South Asia, this phase is placed in between the mid- and late-Holocene period and the seventh to first millennium BCE (Allchin and Allchin, 1982; Chakrabarti, 1993; Dhavalikar, 1997; Jayaswal, 2010; Misra, 2001; Paddayya, 2005, 2011, 2013; Pal, 2006; Pokharia, 2008; Saxena et al., 2006; Singh, 2002; Tewari et al., 2002, 2003, 2006; Verma, 1998). The proto-historic phase is divided into Neolithic, Chalcolithic and Iron-age. The Neolithic phase is defined by the complete absence of archaeological evidence of metal use. The Chalcolithic and Iron-age are marked with the presence of copper-bronze artefacts and evidence of Iron technologies, respectively. Over 4000 sites of the proto-historic phase, including the Indus valley civilization, have been reported in South Asia. However, the discovery of such a site in coastal eastern India is rare.
Recent explorations have unearthed several archaeologically significant sites in coastal eastern India that dates from proto-historic to the medieval period (e.g. Basak et al., 2014; Gangopadhyay et al., 2017). The discovery of the proto-historic phase in Erenda, a small town situated in the coastal region of eastern India (Figure 1), has attested to the presence of early settlers and offered a new cultural perspective of late-Holocene human settlement in coastal India. The site offers a unique opportunity to study the adaptation strategy of the early settlers that resulted due to the changes in the palaeoenvironmental conditions.
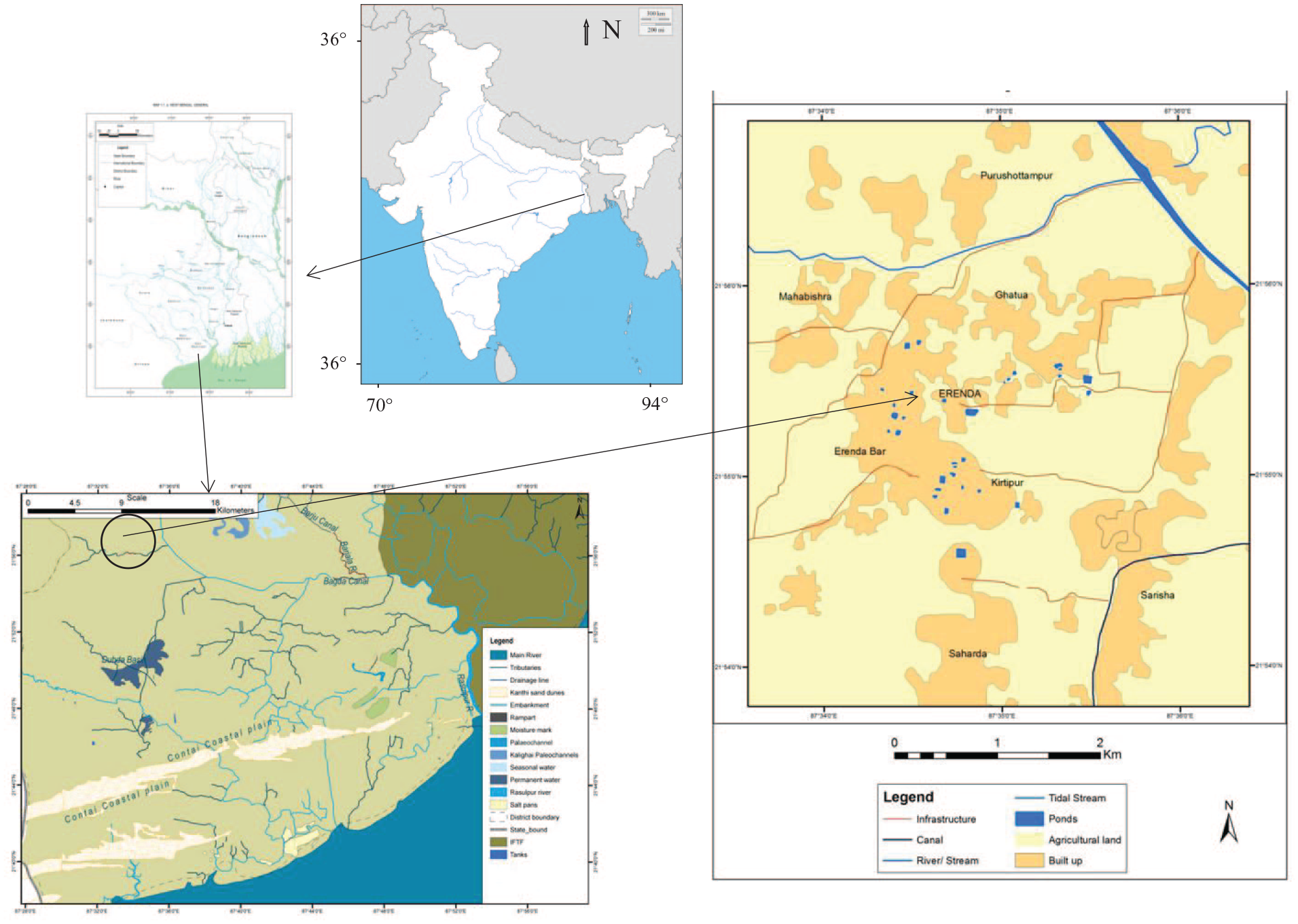
Location of the excavation site at Erenda in West Bengal, India.
Evidence of copper and iron made objects indicates the practice of using metal tools and ornaments by settlers in Erenda. The excavated potteries, dated between 950 BC and 600 BCE (Gangopadhyay et al., 2017), are marked by the presence of Black and Red ware, black slipped ware and varieties of redware. The potteries include various forms of bowls and jars, and vases used for storage. Inhabitants possibly lived in simple huts, consisted of earthen floors with wattle and daub wall and thatched roof. Burnt fragments of the walls have been recovered during excavations. With distinct colour, form and composition, the floors are distinguishable from the natural layers over which they are constructed. Other than these cultural features, the excavated items include faunal, and macro and micro botanical remains. The palaeo-botanical remains have been studied under a microscope and are reported here. With the development of advanced organic geochemical methods, the evidence of palaeoenvironmental conditions have been accentuated worldwide but has received less attention in the South Asian context. Here we present a meaningful application of the organic geochemical methods from a site in coastal eastern India and the first-ever attempt in the Indian state of West Bengal.
Palaeobotanical and geochemical analyses of prehistoric sediments from archaeological sites have gained importance in recent years to solve problems related to the subsistence practices, human-plant relationship and socio-economic developments (e.g. Arobba et al., 2014; Day, 2013; Dunseth et al., 2019; Wu et al., 2012). Microscopic palaeobotanical remains, including spores, pollen, phytoliths and non-pollen palynomorphs (NPPs) such as fungal and algal remains, serve as an excellent indicator of past climate, ecology and vegetation at archaeological sites (e.g. Ghosh et al., 2006; Lu et al., 2007; Reitz and Shackley, 2012; Revelles et al., 2016). Similarly, organic geochemical proxies, such as total organic carbon (TOC) to total nitrogen (N) ratio (C/N), stable isotopes of organic carbon (δ13C) and lipid biomarkers have been used in archaeological research (Aichner et al., 2018; Boutton et al., 1998; Eckmeier and Wiesenberg, 2009; Stolpnikova et al., 2020). The proxies are primarily used for identifying the source (e.g. terrestrial vs aquatic plants) of organic matter (OM) and for gleaning information on the processes affecting the production and preservation of the OM (Chmura and Aharon, 1995; Fleming et al., 1990; Melillo et al., 1989; Thornton and McManus, 1994). The C/N ratio, for example, in algae (5–8) differs from vascular terrestrial plants (>15) (Meyers, 1994). Moreover, the C3 (δ13C = −34‰ and −22‰) and C4 (δ13C = −16‰ and −10‰) plants show distinctly different δ13C values because the C3 and C4 plants follow separate photosynthetic pathway (Cerling and Harris, 1999; Cernusak et al., 2013; Diefendorf et al., 2010; Kohn, 2010; Farquhar et al., 1982, 1989). The δ13C values of C3 terrestrial plants (<22‰) and algae (−32‰ to −12‰) differ because C3 terrestrial plants use atmospheric CO2, and algae mostly use dissolved CO2 as the source of carbon. Further, the broad range of δ13C in aquatic plants depends on the δ13C of dissolved inorganic carbon. Hence, the δ13C composition coupled with the C/N ratio is used to delineate the sources of the OM, especially to identify the relative contribution from terrestrial versus marine, and C3 versus C4 vegetation derived OM preserved in sediments (Cai et al., 2015; Careddu et al., 2015; Das et al., 2008; Li et al., 2016; Ramaswamy et al., 2008).
Normal alkanes (n-alkanes) are straight-chain aliphatic hydrocarbons (Eglinton and Hamilton, 1963, 1967) that are reported in palaeosols, fluvial, lacustrine and marine sediments (e.g. Handley et al., 2008; Quenea et al., 2004; Sachse et al., 2004; Schefuß et al., 2003; Smith et al., 2007). Due to the lack of functional groups in n-alkanes, the biomolecules survive thousands of years in sedimentary geological records (Eglinton and Logan, 1991; Huang et al., 1995; Otto et al., 2005; Peters et al., 2005). Long-chain n-alkanes (n-C21–n-C37) are derived from epicuticular leaf wax of terrestrial plants and are often used as a terrestrial plant biomarker (Bush and McInerney, 2013). Leafy vegetation, in particular, is considered as the primary source of n-C27–n-C29 (Cranwell et al., 1987), whereas land grasses and small shrubs are considered as the source of n-C31 (Maffei, 1996). The n-alkane composition of phytoplankton and bacteria is dominated by short-chain compounds (n-C15–n-C20) (Blumer et al., 1971; Cranwell et al., 1987; Giger et al., 1980), where bacteria generally exhibit an even carbon number (n-C18 and n-C20) predominance (Han and Calvin, 1969). Submerged and floating aquatic plants are characterised by medium-chain (n-C21–n-C25) n-alkanes (Cranwell, 1984; Ficken et al., 2000).
Interestingly, the combination of palaeobotany, stable isotopes and n-alkanes has rarely been used in interpreting the ecological constraints and subsistence practice of pre-and proto-historic cultures in Indian archaeology (Ghosh et al., 2006). To the best of the authors’ knowledge, no published literature using a blend of palaeobotany, organic geochemistry and archaeology is available from a site in eastern India. Hence, the objectives of the study were to reconstruct the past environment and ecological conditions prevailing during the human occupation at the excavated site in Erenda by using δ13C, n-alkanes and microscopic plant remains, and to correlate the ecological and cultural matrix of early farming societies. The proto-historic phase of the site, as indicated by the excavated artefacts, has been validated by Accelerator Mass Spectroscopy (AMS) radiocarbon dating.
Excavation site and geology
Erenda
The excavated mound in Erenda (N 21°55′4.8″; E 87°34′ 42.4″) is located 30 km inland from the present coastline of the East Medinipur District of the Indian state of West Bengal (Figure 1). The archaeological mound lies ~1.7 m above the surface and covers an area of approximately 25 × 40 m2. Temple of a local goddess, Basuli, is located on the top of the mound. The occurrence of diagnostic pottery of the proto-historic phase in West Bengal, such as Black and Red Ware, along with other medieval wares, is recorded on the surface and in the exposed sections of the mound. Sculptures of the medieval period have also been recovered from the northwest corner of the mound. The habitation deposit, extended towards the northwest direction, is disturbed by present-day agricultural activities (Basak et al., 2014; Gangopadhyay et al., 2017).
Geological setting
Erenda is located in the palaeo-delta of Subarnarekha River (Figure 1). The drainage primarily comprises a series of man-made tanks and canals. These canals are likely remnant of past tidally affected channels, which later have been anthropogenically modified. The excavation site lies on the juncture of Late Pleistocene–Early Holocene Sijua and Middle Holocene Panskura Formations, and is covered with Mid-Late-Holocene tidal sediment lying ~5–10 m above the present-day sea level (Ghosh and Guchhait, 2015; Roy and Banerjee, 1990; Vaidyanandan and Ghosh, 1993). Calcrete nodule bearing Sijua Formation, consisting of fine to medium sand and deposited under estuarine conditions, has a vast expanse in the Western half of the Bengal Delta. Niyogi (1975) assigned an age of 175–275 Ka to Sijua Formation, but no published radiocarbon date of Sijua Formation is available (Shah and Chatterjee, 2021). A thick (~1–1.3 m) and dark to light brown silty clay deposit of Panskura Formation (dated between 4810 BP and 789 BP; Roy and Chattopadhyay, 1997) unconformably lies above the Sijua Formation. The silty clay interval contains archaeological layers defined as sedimentary units with similar artefactual remains and belonging to the same chronological horizons. The silty clay interval is most likely deposited under low energy fluvial/fluvio-tidal conditions. The areal spread of the silty clay is associated with the regression of the sea and deposition by rivers and tidal creeks. The base of the silty clay interval is virgin and indicates a phase before the first colonisation at the site.
Material and methods
Excavation at Erenda followed the principle of archaeological stratigraphy (Harris, 1997), and was conducted by laying square grids (6 m × 6 m) all over the site and subsequent excavation of three grids as ‘trenches’, namely ZA1, A1 and A2. Identification and differentiation of archaeological layers were based on the lithological characteristics. The layers essentially connoted individual strata as they could be observed in a horizontal section cut into the soil. The method of identifying archaeological stratification followed the laws of superimposition as originally conceived in geology. Changes in the artefacts, if any, were used as markers where changes in lithology were not visible. Cultural features, such as floors and garbage pits, were embedded in the natural soil. Layers of varying thickness were equated with cultural periods, initially based on artefacts or relative dating and thereafter through absolute dating by using the AMS radiocarbon method (Table 1). In trench ZA1, layers 4 and 5 culturally belonged to the protohistoric and early historic period. The layers 1, 2 and 3 lying above layers 4 and 5, and culturally belonged to the early medieval, medieval and modern periods, respectively. In trench A1, layer 3 belonged to the protohistoric phase, whereas the early historic phase was sandwiched between the layers 2 and 3. The layers 1 and 2 belonged to the cultural period of modern to early medieval periods.
AMS radiocarbon and calibrated age of charcoals collected from the Trench ZA1 and A1 (Figure 3). The radiocarbon ages were calibrated using the calibration curve CalPal 2007 Hulu, included in the CalPal software (2007 Version 1.5) (Weninger et al., 2007).

AMS radiocarbon analysis was performed at IUAC in India.
AMS radiocarbon analysis was performed at Palaeo Labo Co. Ltd in Japan.
Microbotanical analysis
Six soil samples (DG 20, 19, 18, 17, 16 and 6) for the spore, pollen, non-pollen palynomorphs (NPPs) and phytolith analyses were collected from the western wall of the trench A1 (Figures 2 and 3). The conventional non-destructive method was used following the protocol of Faegri and Iversen (1950) to extract the NPPs. Subsequently, the standard acetolysis method for clearing the cellulosic content of the pollen wall was used following the protocol given by Erdtman (1954). The samples were boiled with a 10% potassium hydroxide (KOH) solution, followed by hydrofluoric acid (HF) treatment and finally acetolysis with a mixture of acetic anhydride and concentrated sulphuric acid (H2SO4) (9:1 ratio). The extracted palynomorphs were stored in water and studied under the light microscope (Zeiss Axioskop 2) at ×450.

(a) Trench A1 layer 17. (b) Trench A1 North Section layers. (c) Trench ZA1. Eastern Section wall. (d) Trench ZA1. Sample collection area from Eastern Section.

(a) Stratigraphic layers. (b) AMS 14C ages and sampling intervals of organic geochemical and palaeobotanical studies in the trench ZA1 and A1 at the archaeological site in Erenda.
Phytoliths were extracted from soil samples following the modified protocol of Pearsall (2000). Dried, homogenised and sieved samples (2 g) were treated with concentrated nitric acid (HNO3) and hydrochloric acid (HCl) (1:1 v/v). Hydrogen peroxide (H2O2) was used for removing OM prior to density separation using cadmium iodide (CdI2) + potassium iodide (KI) with (sp. gr 2.3). The extracted phytoliths were mounted on slides using polyvinyl alcohol and DPX (Di-N-Butyl Phthalate) and were studied and photographed using a Zeiss Axioskop 2 microscope. Over 200 phytolith morphotypes were counted for each sample. The phytoliths were identified and classified following the classification of Fredlund and Tieszen (1994); Lu and Liu (2003); International Code of Phytolith Nomenclature (ICPN) 1.0, Madella et al. (2005); Solomonova et al. (2019); ICPN 2.0, ICPT: Neumann et al. (2019). The following grass phytolith indices were calculated.
a. Humidity-aridity Index (Iph %)
The Iph (Diester-Haass et al., 1973) was calculated by using the following equation.

Iph measured the dominance of Chloridoideae grasses over Panicoideae grasses. Chloridoideae grasses usually grew in warm and dry climatic or edaphic conditions, whereas Panicoideae grasses were adapted to warm and humid conditions with the availability of high soil moisture. Therefore, a xeric climatic condition was indicated by higher Iph values and the lower values indicated a mesic condition.
b. Water Stress Index (Fs %)
The Fs (Bremond et al., 2005) directly measured the transpiration rate or water stress and was represented by the following equation.

n-Alkane and stable isotope analyses and radiocarbon dating
Soil and sediment samples for organic geochemical analyses were collected from the eastern wall of layers 3, 4 and 5 of the trench ZA1 (Figure 3), which was adjacent to the trench A1 from where palaeobotanical samples were collected. The samples were dried in the air, homogenised using a mortar and pestle, and sieved through a −200-mesh sieve. The TOC and total N were analysed using a Euro EA3024 elemental analyser (EA). The carbon isotope analyses were performed by using an Isotope Ratio Mass Spectrometer (IRMS) coupled to the EA. Samples for TOC and δ13C analyses were treated with HCl acid to remove carbonates. Results were expressed in delta (δ) per mille (‰) notation with respect to the Vienna-PeeDee Belemnite standard as the calibration standard. The results of the C isotope analyses were calculated using the following equation:
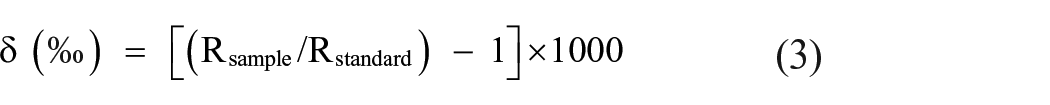
where Rsample for C was the 13C/12C ratio of the sample, Rstandard was the 13C/12C ratio of the calibration standard.
IA-R001 (wheat flour, δ13CV-PDB = −26.43‰) was used as the reference material. Check samples (IA-R001, IA-R005 (beet sugar, δ13CV-PDB = −26.03‰) and IA-R006 (cane sugar, δ13CV-PDB = −11.64‰)) were used for quality control. The check samples were calibrated against IAEA-CH-6 (sucrose, δ13CV-PDB = −10.43‰), which was an inter-laboratory comparison standard distributed by the International Atomic Energy Agency (IAEA), Vienna. Blanks and internal standards were run in between every eight samples. The precision of the analyses (10 replicated standard samples) was 0.3% for C, 0.03% for N and 0.06‰ for δ13C. The duplicate analysis was performed in 20% of the samples. The reproducibility of duplicate analyses was 0.1‰.
Charcoals, carefully collected from the layers 5, 4 and 2 in the trench ZA1 and layer 3 in the trench A1, were used for age determination using Accelerator Mass Spectroscopy at the Inter-University Accelerator Centre (IUAC) facility in New Delhi, India (Naskar et al., 2018) and the Palaeo Labo Co. Ltd in Japan (Table 1).
Correlation between trenches
Correlation of the archaeological layers of the two adjacent trenches, such as ZA1 and A1, was established by using digs, which were measured from a known benchmark (5 m in case of Erenda) above the mean sea level (MSL). Hence, digs in different trenches but lying at the same depth from the benchmark (5 m above the present MSL assumed during the excavation) belonged to the same cultural level. The AMS radiocarbon dating of charcoals collected from the digs in different trenches was further used to establish the inter-trench correlation.
Results
AMS radiocarbon dates and age of the site
The AMS radiocarbon dates (Table 1) testified that the human settlers occupied the site in Erenda from the beginning of the first millennium BCE (proto-historic period) until the middle of the first millennium CE. Hence, the proto-historic communities in Erenda represent perhaps one of the earliest human settlements in coastal east India. Cultural signatures, especially artefacts including pottery, metal and bone tools, were recovered from the layers 5 and 4. Dark brown silty clay and carbonate composition of the layers indicated living and working areas (therefore labelled as floors), and were characterised by earth rammed with potsherds. Remains of wattle and daub, particularly in layer 4 and partially in layer 3 in the trench ZA1, were observed. The occurrence of pottery, terracotta figurines and lamps, belonging to the early historic (third century BCE to the third century CE) and early medieval (fourth or fifth century CE) phases, above the layer 4 in the trench ZA1 were found mostly as dumps instead of well-defined cultural features. The non-cultural materials (ecofacts), excavated from the dumps, included vertebrate (both terrestrial and aquatic) bones and shells of invertebrates (e.g. molluscs) and were used as food resources.
Pollen, NPPs and phytolith
The identifiable palynoflora (Figure 4), along with the affinity and environment, were presented in Table 2. Presence of pollen grains of Casuarina sp., Clerodendron sp., Borassus sp., Brassicaceae, Fabaceae, Anacardiaceae, Cheno-Amaranthaceae, Poacaea and spores of Pteris sp., Ceratopteris sp., Polypodiaceae were observed in the studied samples. In addition, the NPPs recovered were algal cyst, Concentricystes and fungal spores.
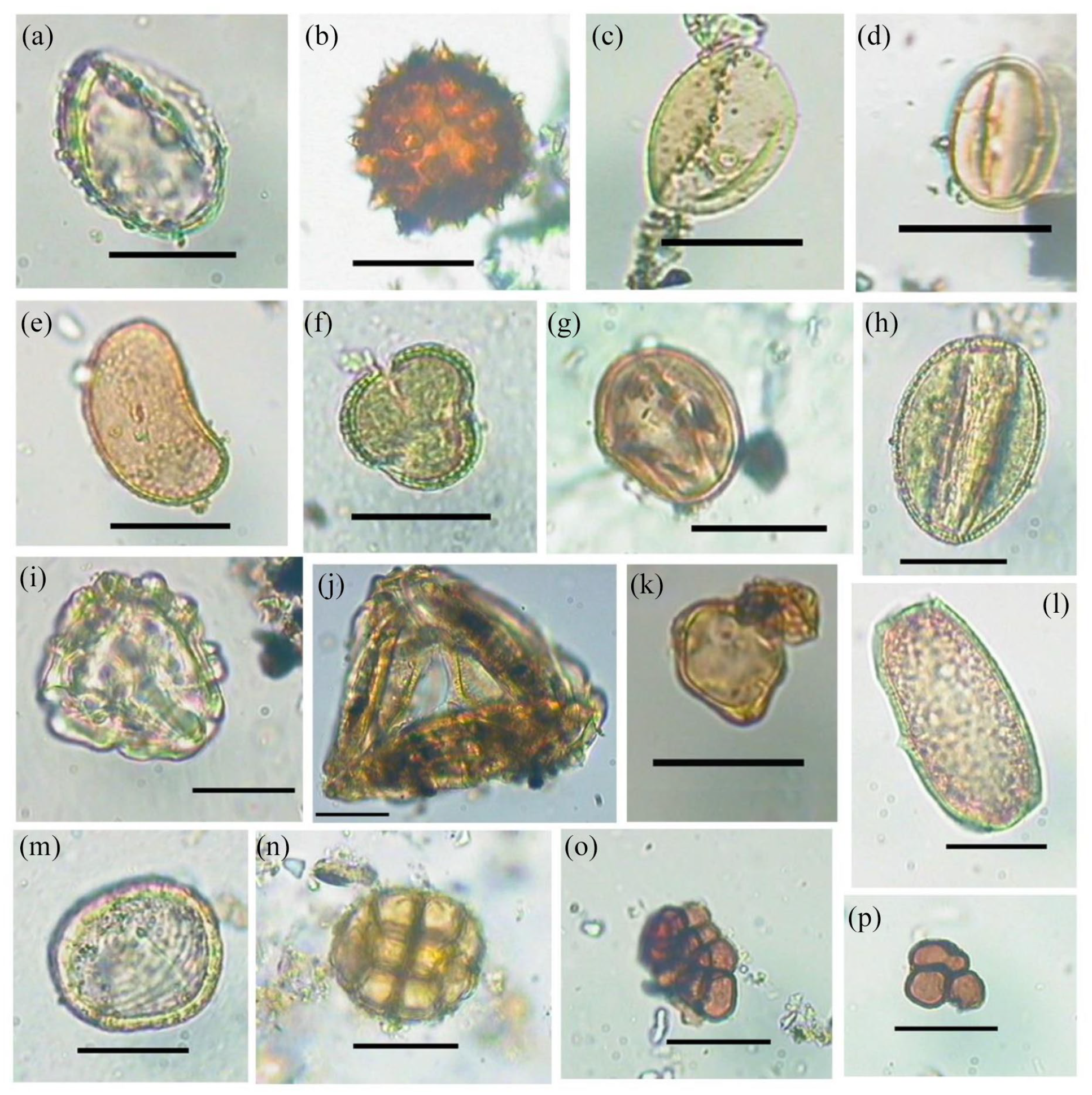
Palynoflora in soil samples from the trench A1 recovered from the archaeological site in Erenda. (a) Borassus sp. (b) Asteraceae. (c) Poaceae. (d) Fabaceae. (e) Polypodiaceae. (f) Brassicaceae. (g) Anacardiaceae. (h) Clerodendron sp. (i) Pteris sp. (j) Ceratopteris sp. (k) Casuarina sp. (l) Algal cyst. (m) Concentricystes. (n) Acacia sp. (o, p) Fungal spores (Scale = 10 µm).
Details of palynotaxa recovered from the soil samples from trench A1 in Erenda.

Twenty-one phytolith morphotypes were recovered from the studied samples (Figure 5). Among the recovered phytoliths, there were eleven different types of grass silica short cell (GSSC) phytoliths having ecological significance (i.e. bilobate, bilobate trapezoidal, bilobate scooped, cross, polylobate, saddle, saddle plateaued, saddle tall, saddle collapsed, rondel and trapezoid) whereas, the rests are non-diagnostic (i.e. acute bulbosus or silicified hair cell, blocky, epidermis, elongate cell, hair cell base, stomate, tower and tracheary). The non-grass phytolith morphotypes were grouped under a single category. The water stress marker phytolith (bulliform flabellate) was also recovered from the soils (Figure 5).
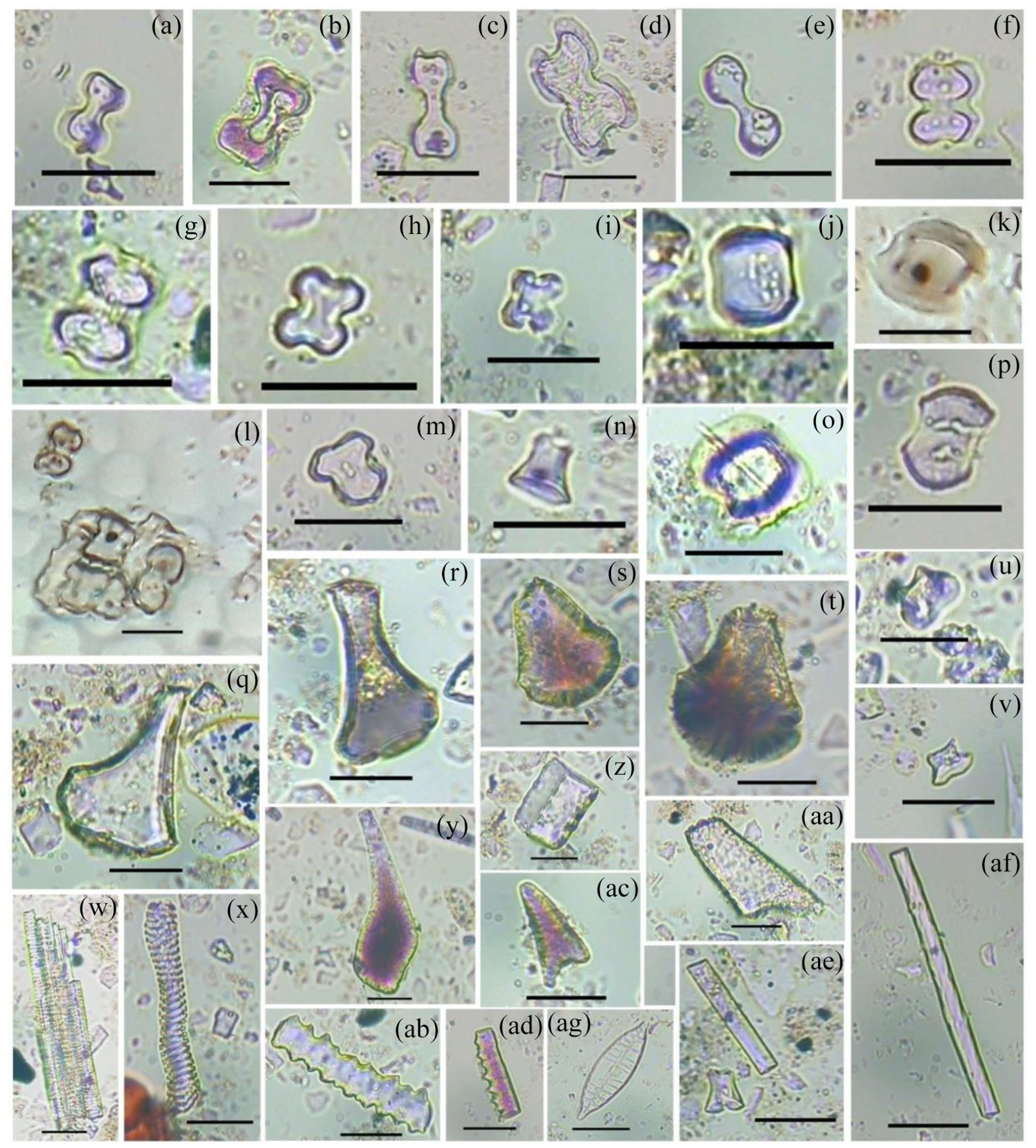
Light microscopic images of some phytolith morphotypes. (a–e) Bilobates. (f, g) Bilobate scooped. (h, i, m) Crosses. (j) Saddle. (k, o) Saddle plateaued. (l) Epidermal plate with attached short cell phytolith morphotypes. (n, u, v) Rondels. (p) Saddle tall. (q–t, y) Bulliform flabellate. (w, x) Tracheary. (z) Blocky. (aa, ac) Acute bulbosus. (ab, ad–af) Elongate cells. (ag) Diatom (Scale = 10 µm).
It was observed that the samples were characterised by a low content of woody dicot morphotypes (~7%) and relatively higher of Poaceae phytoliths (~90%), which was dominated by bilobates (~30%). Significant representations of other GSSCs, such as crosses (~4%), saddles (~3%), rondels (~7%), trapezoid (~3%) and saddle plateaued (~2%) were also noticed (Figure 6). Trapezoid and rondel types are known to be mainly derived from C3 plants in higher frequencies, whereas bilobates, crosses, polylobates and saddles from C4 grasses (Bremond et al., 2008). Other C3 indicating morphotypes recovered from the studied samples were bilobate trapezoid, bilobate scooped, saddle plateaued, saddle tall and saddle tall (Biswas et al., 2016). Recovery of bilobate scooped suggested the presence of the genus Oryza (rice) which may be of wild or domesticated variety (Piperno, 2006; Prasad et al., 2011).

Frequency of phytoliths recovered from the soil samples from the trench A1 in Erenda, along with the trends of two indices (Iph and Fs) applied on the samples and C3 and C4 morphotypes.
Subsequently, The C4 phytolith showed an overall dominance (32%–43%) but varied in frequencies throughout the trench. The abundance of C4 morphotypes was higher (39%–43%) in samples DG 20, DG 19 and DG 18 (Figure 6) in the trench A1. The sampling depths of DG 20, DG 19 and DG 18 in the trench A1 were correlated to the layer 5 of the trench ZA1 (Figure 3). The abundance of C4 morphotypes decreased (32%–33%) in samples DG 17 and DG 16 (Figure 6) collected from the depths equivalent to the bottom and top of the layer 4 in the trench ZA1 (Figure 3). The overall abundance of C3 morphotypes was low and show minute fluctuations with depth (Figure 6). The frequency of C4 morphotypes was high (~37%) in the uppermost sample DG 6. The observation based on the grass phytolith indices was consistent with the reported presence of both C3 and C4 plants in the Lower Ganga Plain (Basu et al., 2015). Low Iph (2%–11%) and Fs (10%–18%) values suggested a mesophytic environment with high humidity and an overall low evapotranspiration rate, respectively (Bremond et al., 2005, 2008) (Figure 6).
TOC, C/N and δ13C
The TOC showed a near-constant value of approximately 0.2% in Sijua Formation, varied between 0.3% and 0.4% from layers 5–4, and showed relatively higher values (0.4%–0.7%) in layer 3 and above. The C/N ratio varied between 8 and 10 in Sijua Formation, followed by a sharp increase to 13 in layer 5. The C/N ratio decreased upwards, was nine at the boundary between layers 4 and 3, and ranged between 9 and 11 within and above layer 3. The δ13C became positive upward in Sijua Formation (−17‰ to −19‰), and attained the highest positive value (−16‰) in layer 5 followed by a negative excursion in layer 3 and above (−18‰ to −20‰).
n-Alkanes
Ratios of sedimentary long-chain n-alkanes were calculated for understanding the source of the OM (e.g. Das et al., 2009). The aquatic macrophyte n-alkane proxy (Paq = (C23 + C25)/(C23 + C25 + C29 + C31)] expressed the relative proportion of n-alkanes derived from non-emergent macrophytes (n-C23 and n-C25) and emergent macrophytes plus terrestrial plants (n-C29 and n-C31). The Paq was 0.4 in Sijua formation and 0.6 in layer 5. The Paq was zero in the rest of the intervals (Figure 7).
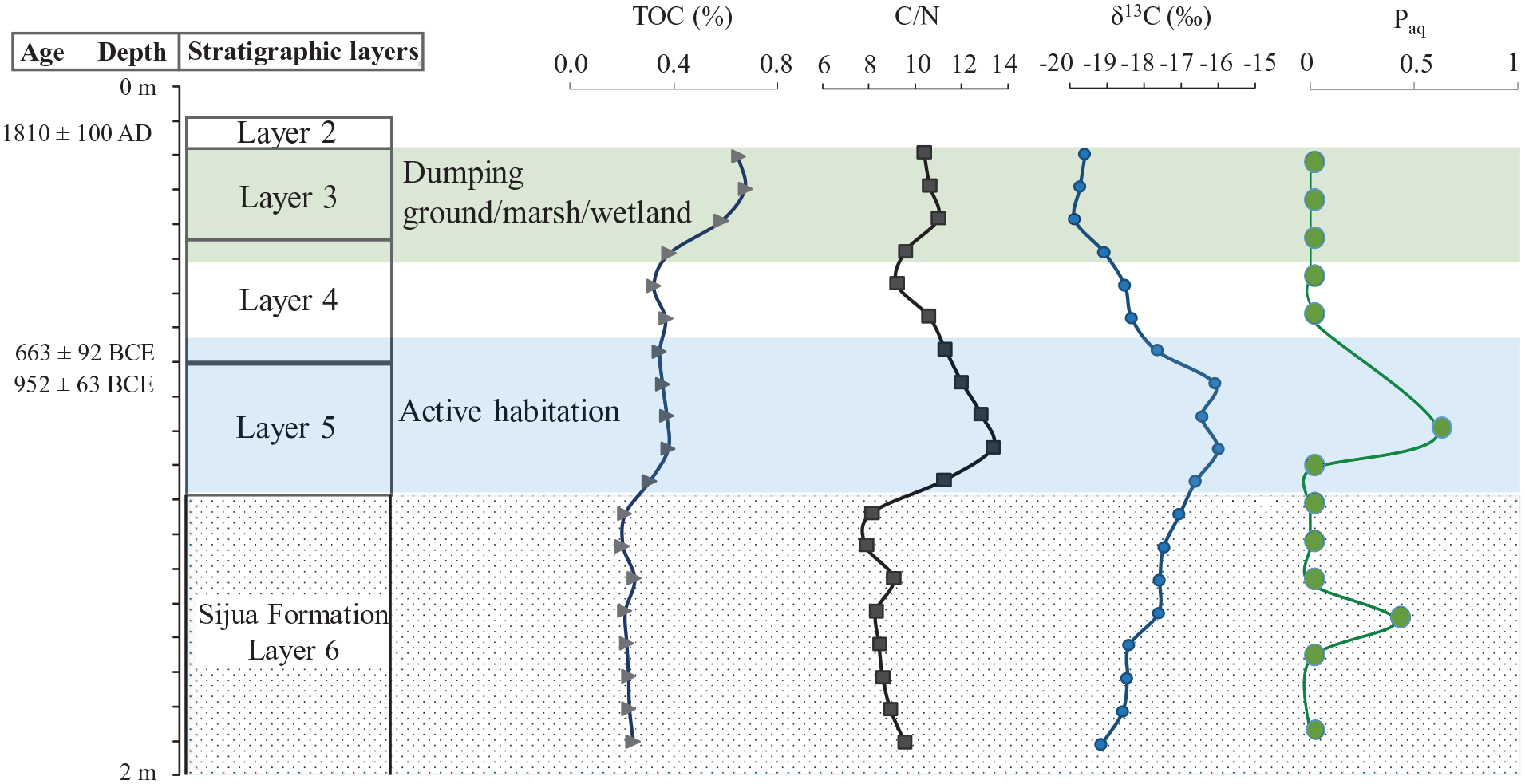
Stratigraphic layers, AMS 14C ages and depth profile of TOC, C/N, δ13C and Paq of trench ZA1 in Erenda.
Discussion
Palaeobotanical interpretation of climate and environmental conditions in Erenda
The recovery of the pollen grains having an affinity with the family Brassicaceae, Fabaceae, Anacardiaceae along with spores of Pteridaceae and Polypodiaceae predicts that a tropical and mesophytic condition prevailed at the site. The presence of spores of Ceratopteris and Concentricytes suggests the presence of a freshwater body near the vicinity of the settlement. Nonetheless, the presence of pollen grains of Casuarina sp., Borassus, along with other monosulcate pollen having an affinity with Palmae and Cheno-Amaranthaceae indicate the closeness of the archaeological site to the coast. Good numbers of dispersed fungal spores and the dominance of C4 phytolith morphotypes (i.e. bilobates and crosses) in the samples also indicate tropical warm and humid climatic conditions (Biswas et al., 2016; Bremond et al., 2008; Dilcher, 1965). This observation is consistent with the presence of saddle tall and saddle collapsed mostly found in Bambusoideae grasses that also indicate humid climate conditions prevailed during the phase of active human occupation at the excavated site in Erenda. Our observation is further supported by the reported prevalence of humid tropical climate at the Chalcolithic site of Pakhanna in Bankura District of West Bengal (approx. 200 km away, Chattopadhyaya et al., 2005; Ghosh et al., 2006). The recovery of saddle plateaued, which are found mostly in Phragmites grasses, is consistent with the spore-based interpretation on the presence of freshwater bodies nearby the settlement because Phragmites, a C3 Arundinoideae grass, grows only in and around swampy areas or freshwater bodies.
TOC, C/N and stable isotope source signature of OM
Organic matter in estuarine and coastal sediments represents a heterogeneous and complex mixture of compounds derived from multiple sources, including brackish/marine and freshwater phytoplankton, terrestrial and aquatic plants (Cai et al., 2015; Lamb et al., 2006; Schreiner et al., 2013). The C/N ratio (8–10) and δ13C values (−17‰ to −19‰) in the Sijua Formation (Figure 7) indicate marine phytoplankton and marine particulate OM (POM) as sources of the sedimentary OM and a tidal flat depositional environment (Figure 8).
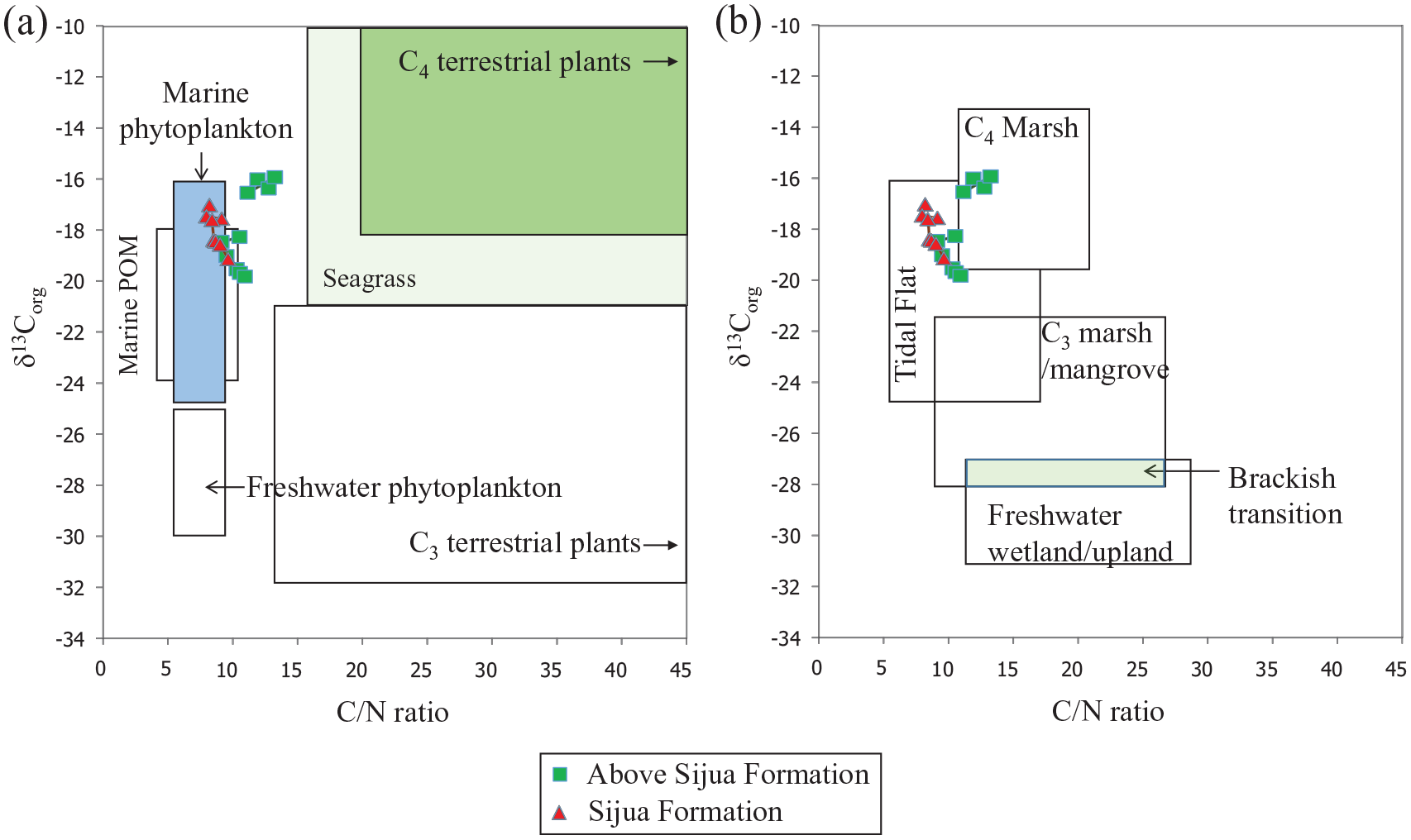
The range of δ13C versus C/N values displaying ranges of (a) organic matter sources (marine and freshwater phytoplankton, C3 and C4 terrestrial plants and sea grass) and (b) tidal flat, C4marsh, C3marsh or mangrove, brackish transitional and freshwater/upland depositional environments (Khan et al., 2015), and the δ13C versus C/N values of soil and sediment samples excavated from the archaeological site in Erenda.
The δ13C values show a positive trend (−20‰ to −16‰) above the Sijua Formation. Positive δ13C values (−16‰ to 17‰) in the layer 5 (Trench ZA1) fall within the range of OM produced by C4 plants (−16‰ to −10‰; average −13‰; Basu et al., 2015; Cerling and Harris, 1999). The stable isotope-based interpretation is substantiated by the increased abundance of C4 morphotypes, that is, bilobates (~31.9%), cross types (5.4%) and saddle (2.43%) in samples DG 20, DG 19 and DG 18 from the trench A1 (Figures 3 and 6). A dominant supply of OM from C4 vegetation at intervals where the evidence of human settlement becomes prominent and the floors of mud-built hut start appearing indicates the practice of using C4 plants, mixed with clay, as a construction material of huts by the settlers. This is because the presence of regular habitations at places with foliage dense enough to be reflected in the δ13C signal is quite improbable. Grasses, found in the marshy wetlands of eastern India, are used by the villagers for the construction of huts even today. A similar practice has likely been followed in the proto-historic periods. The occurrence of a large patch of clay belonging to the Sijua Formation at the anthropogenic floor in trench ZA1 indicates the use of the clay as a construction material. The choice of the Sijua clay is justified by its calcium carbonate composition that exerts more durability to the floor of the hut. The presence of charcoal along with burnt clods of clay with grass impressions below the floors indicate anthropogenic clearance of the foliage using fire prior to the construction of the huts. The transformation of the excavation area from an active habitation to a dumping ground is reflected by the reoccupation of C3 vegetation as evidenced from a drop in C/N ratio (10–11) and lighter δ13C values (−20‰) above the layer 4 (Figures 3 and 7). The decreased abundance of C4 morphotypes, that is, bilobates (~26%), crosses (~2%) and saddle (~2%) and increasing dominance of the C3 grasses morphotypes, mainly bilobate scooped types and rondels at corresponding layers of the trench A1 (Figures 3 and 6) is consistent with the stable isotope-based interpretation.
n-Alkane biomarker signature of OM source
The Paq shows distinct values for terrestrial plants (Paq < 0.1), and helps to distinguish emergent macrophytes (Paq = 0.1–0.4) from submerged/floating macrophytes (Paq = 0.4–1.0) (Ficken et al., 2000). In Erenda, the Paq indicates the presence of emergent macrophytes (Paq = 0.4) in the Sijua Formation, and submerged/floating macrophytes (Paq = 0.6) derived OM at the cultural interval of the layer 5 where the evidence of human settlement, and presence of mud-built hut using C4 vegetation, started appearing. Interestingly, the Paq indicates an aquatic source of the OM and the δ13C values (−16‰ to −10‰) and presence of C4 phytoliths indicate a dominant supply of C4 vegetation derived OM in the layers where the evidence of human settlement and presence of mud-built hut started appearing. This supports our inference that the inhabitants at the site used C4 grasses for building the huts.
Landscape evolution and protohistoric human settlement in the mid-late-Holocene period
The absence of anthropogenic evidence in the Sijua Formation indicates inhabitable conditions in nearshore/estuarine tidal flat conditions. Notably, human settlement in Erenda, at the beginning of the first millennium BCE, follow a post 2 ka regressive event recorded in coastal eastern India (Sarkar et al., 2009). In general, sea level is considered to have remained stable since c. 3000 BCE, although Hazra et al. (2001) have reported transgressive events at c. 970 ± 100 BCE and c. 1400 BCE along the eastern coast. However, the authors did not provide any information on the dating technique used to establish the date of the events. Farooqui and Vaz (2000) have reported sea-level regression following the major transgressive event at c. 4700 ± 110 BCE along the east coast of India. Perturbations in tidal range and intensity between 1150 and 370 BCE are inferred from radiocarbon-dated tidal to brackish water bivalves from Pulicat Lagoon in the Indian state of Andhra Pradesh on the eastern coast and are linked to sea-level changes (Vaz and Banerjee, 1997). The authors further observe a drop in sea level during the Little Ice Age (1700–1750 CE). We hypothesise that the proto-historic human settlements in Erenda are closely linked to the regressive events when the area became relatively habitable, and early settlers constructed huts of mud and aquatic grass plastered walls with floors made of local clay. The presence of a freshwater body near the vicinity of the settlement, as indicated by the spores of Ceratopteris and Concentricystes, might act as a source of drinking water and encouraged the proto-historic human to settle in Erenda. The habitation area of the site was later transformed into a dumping ground and eventually covered by terrestrial C3 plants and C4 grasses as evidenced by the phytolith distribution (Figure 6) and n-alkanes and stable isotope signatures (Figure 8). However, settlers continue to live in the area, as evident from the finds of pottery and other objects belonging to the early medieval period.
Conclusions
The organic geochemical study reveals that the proto-historic people started living at the site during the regressive episode. A prevailing warm and humid climatic condition during the human occupation and proximity to a freshwater body has been inferred from the presence of fungal spores and the dominance of C4 phytolith morphotypes. The settlers most likely used to consume the wild or domestic variety of rice, and used C4 aquatic grass for mud and clay-built hut construction. The site later transformed into a dumping pit and covered with vegetation. Palaeobotanical evidence and soil OM characteristics represent contemporary cultural activities and reflect the environmental and ecological conditions prevailing during the human occupation at the site. The palaeobotanical and organic geochemical study at the proto-historic site in Erenda is unique in terms of the coastal archaeology of India.
Footnotes
Appendix 1
Acknowledgements
The Department of Archaeology, University of Calcutta is acknowledged for support in conducting the excavations. The ASI is acknowledged for giving the license for excavation. The authors also thank Susanta Lahiri for helping in radiocarbon dating. S.N Rajaguru, and Sujit Dasgupta are acknowledged for reviewing an earlier version of the manuscript. Monalisa Rakshit is acknowledged for providing the map. Suggestions from two anonymous reviewers improved the manuscript.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The authors gratefully acknowledge the financial help of the University of Calcutta and IUAC through projects granted to KG for the study.