
Abstract
A combined analysis of sedimentological, molecular, and microfossil proxies was used to determine paleoclimatic and paleoceanographic variations and assess sediment transport pathways along the last c.a. 6300 years in a sedimentary record collected on the inner shelf of the southwestern Atlantic Ocean. The combined use of terrestrial and marine proxies enables a more complete environmental perspective of the La Plata River plume influence on the southwestern Atlantic shelf, regarding the relationship between climate, productivity, as well as hydrodynamic. Variations in marine productivity, lipid biomarkers of terrestrial plants, diagnostic geochemical indices, and sea-surface temperature point to three phases within the core. The first interval (6300–5200 cal yr BP) was characterized by low marine and terrigenous organic matter input, with pulses of phytodetritus indicated by benthic foraminifera. These features suggested dry climatic conditions over the southeastern portion of South America, with the influence of sediments from the adjacent continent and, on a smaller scale, transported from the Argentinian shelf due to the high relative sea level. The second interval (5200–2800 cal yr BP) was distinguished by a slight increase in marine productivity and terrigenous input, indicating a transition period with an increase of total rainfall influenced by an intensified South American Monsoon System (SAMS) and the modern El Niño-Southern Oscillation (ENSO) phenomenon. These features, associated with the changes in wind patterns and the lower relative sea level, suggest the onset of the influence of the La Plata River (LPR) plume over the southern regions of the Brazilian continental shelf for the last 6300 cal yr BP. The last interval (2800–890 cal yr BP) had the highest contents of marine and terrigenous proxies, suggesting less climatic variability, as the long-term intensification of the SAMS and a further increase in the amplitude of the ENSO resulted in the modern establishment and dominant influence of the LPR plume on the core site. Variations in sea-surface temperature up to 3.3°C were related to the influence of the LPR plume on the shelf off southern Brazil.
Introduction
The climatic conditions of the Holocene period (covering the last ~11,700 years) have been characterized with less variability compared to the late glacial period. However, on time scales of centuries to millennia, abrupt climate changes have also been recognized in this period (Bond et al., 1997; Mayewski et al., 2004; Wanner and Ritz, 2011). These events, coupled to orbital insolation variations scale, had significant impacts on the landscape, vegetation, and global climate, with chemical changes in the oceans (Curry et al., 1988; Miriyala et al., 2017), atmosphere (Riebeek, 2005; Steig, 1999), and terrestrial plant structures (Baker et al., 2001; Ganopolski et al., 1998; Mayle et al., 2000). Understanding the mechanisms of variability and finding patterns of climatic events in the geological record may reveal the sensitivity, frequency, and probability of future fluctuations.
Numerous studies have been conducted on the southwestern margin of the Atlantic Ocean to understand how variations in the climate and hydrodynamic conditions in the Holocene have determined the sources, transport, and deposition of sediments (de Mahiques et al., 2009; Gyllencreutz et al., 2010; Mathias et al., 2014; Nagai et al., 2009; Perez et al., 2018). The main oceanographic variations in the mid- to late-Holocene were associated with relative sea-level (RSL), riverine discharge, and the wind regime, with millennial oscillations in the sedimentary conditions resulting from the changes in wind-driven water masses (de Mahiques et al., 2009; Gaiero et al., 2004; Gilli et al., 2005; Gyllencreutz et al., 2010; Prieto et al., 2017; Razik et al., 2013). The main hydroclimatic changes over southeastern South America during mid- to late-Holocene were controlled by changes in insolation on an orbital timescale (e.g. Cruz et al., 2005) and North Atlantic sea-surface temperatures on a millennial timescale (e.g. Vuille et al., 2012). The role of ENSO, on the other hand, is largely discussed since mid- to late-Holocene records with an interannual resolution are scarce (e.g. Piovano et al., 2009; Razik et al., 2013).
The southeastern portion of South America has a modern seasonal rainfall pattern that is tightly associated with continent-ocean thermal contrasts that cause variations in regional atmospheric circulation (Vera et al., 2002, 2006a; Zhou and Lau, 1998). The South American Monsoon System (SAMS) is an important climatic component of this region and strongly affects changes in precipitation, including in the region of the La Plata River (LPR) drainage basin (Garreaud et al., 2009).
Regarding decadal-scale variations, climatic changes are a product of the overlap of various large-scale phenomena (Chiessi et al., 2009; Garreaud et al., 2009), such as the ENSO, Atlantic Multidecadal Oscillation (AMO), and Pacific Decadal Oscillation (PDO). On a centennial scale, changes in the intensity of the SAMS appear to be partly driven by variations in the temperature of the Northern Hemisphere, especially along the North Atlantic Ocean. These changes generate the latitudinal migration of the ITCZ, which, in turn, affects the quantity of upstream precipitation in the Amazon basin and causes a change in monsoon intensity (Bernal et al., 2016; Cruz et al., 2005; Vuille et al., 2012).
Sedimentological, physical, and biological evidence of a modern northward displacement of the cold and less saline waters originating from the LPR plume (“Plata plume water” – PPW) and transported by the wind-driven Brazilian Coastal Current (BCC) can be detected up to the 24°S parallel (Eichler et al., 2019; Möller et al., 2008; Nagai et al., 2014; Piola et al., 2000, 2005). The BCC and the related PPW displacement are the primary controlling feature of the seasonal variation in marine production (Ciotti et al., 1995) and the source of continental sediment in the southwestern Atlantic margin (Figure 1; de Mahiques et al., 2004, 2020; Nagai et al., 2014; Razik et al., 2015). The region is also influenced by the discharge of Brazilian southern coastal lagoons (i.e. Patos Lagoon) and the drainage of the Itajaí-Açu and Tijucas River River (Campos et al., 2008; Ciotti et al., 1995; Piola et al., 2008). The maximum northern extension of the BCC and associated PPW are related to years of low discharge due to the low rainfall in the LPR basin and S/SE winds associated with La Niña conditions (Campos et al., 2008). During El Niño events, despite the higher amount of rainfall in the LPR basin, a shift to offshore displacement and southward displacement of the PPW occurs due to the counter N/NE wind force (Figure 1; Gonsalez-Silvera et al., 2006; Möller et al., 2008; Piola et al., 2005). Therefore, wind stress plays a crucial role in the northward displacement of the BCC and transport of the PPW, while changes in river discharge are considered a secondary driving force (Möller et al., 2008; Piola et al., 2000, 2005, 2008). During the early- and mid-Holocene, before the establishment of the influence of the PPW on the southern Brazilian continental shelf, studies proposed that this area could have received sediments from the Argentinian shelf, although the driving features and timing are still discussed (de Mahiques et al., 2009; Gyllencreutz et al., 2010; Mathias et al., 2014; Razik et al., 2013). Moreover, several works described the spatial distribution and seasonal variations of the benthic foraminiferal assemblages and organic carbon flux in the southwestern Atlantic continental margin (Burone et al., 2011, 2013; Eichler et al., 2008, 2019), although little is known regarding the influence of the PPW on temporal fluctuations of productivity during the mid- to late-Holocene in the southern Brazilian continental shelf.
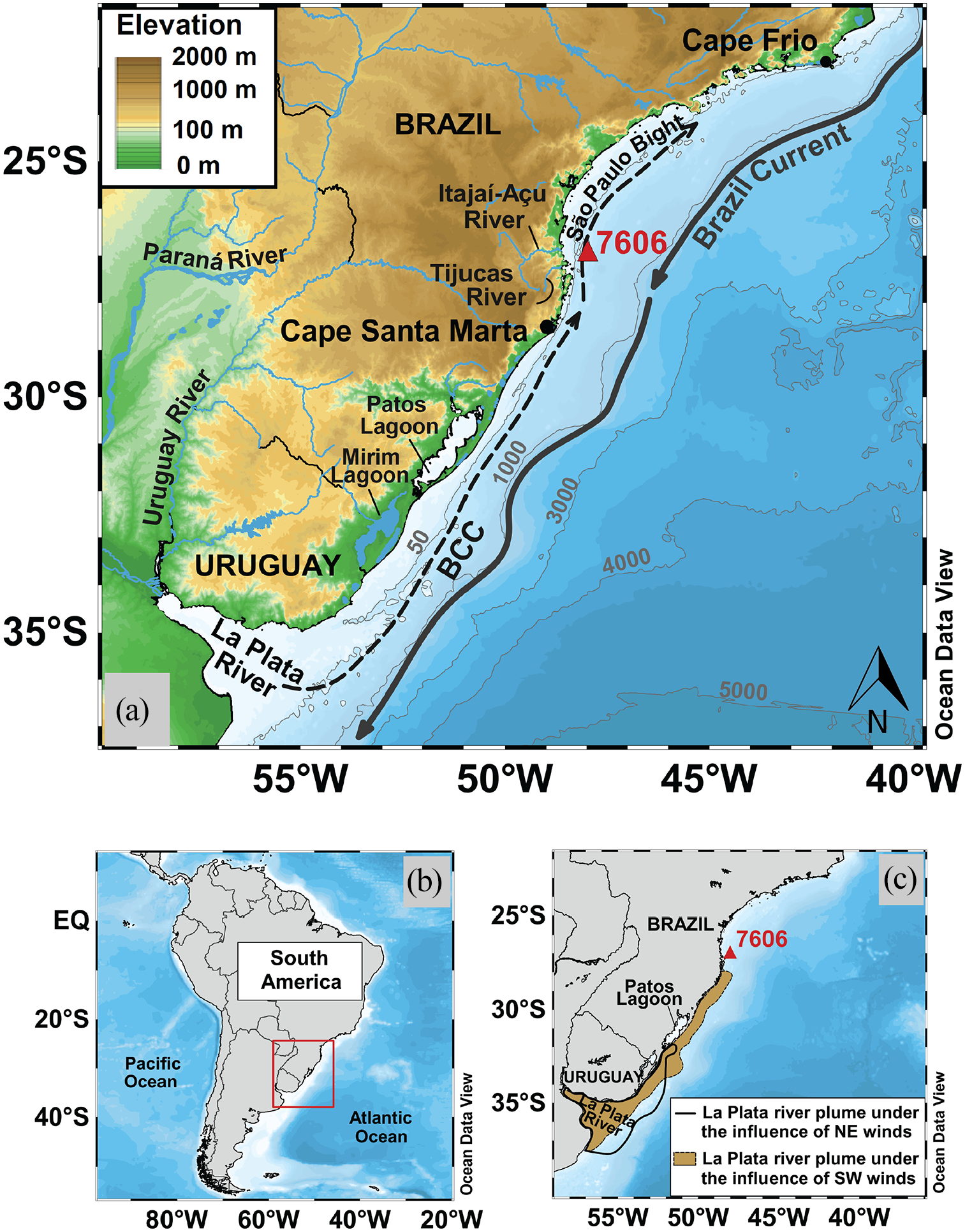
(a) Map highlighting the geographic position of the piston core 7606 (red triangle) and the main oceanographic features occurring near the study area. Brazilian Coastal Current (BCC) (dark gray dashed line) approximate northward spreading was obtained from de Souza and Robinson (2004), while the southward flow of the Brazil Current (dark gray solid line) was estimated from Campos et al. (1995) and Biló et al. (2014). Item (b) shows the location of the study area at Southeastern South America and (c) represents the Rio de La Plata plume displacement under the influence of Northeastern (NE) (black line contour) and southwestern (SW) (brown-shaded area) winds (Piola et al., 2000). Ocean bathymetry and land topography were acquired from GEBCO data, available at www.gebco.net. The coastline was obtained from GSHHS data (www.ngdc.noaa.gov). Rivers and lagoons’ geographic boundaries were obtained from the CIA World Data Bank II (www.evl.uic.edu/pape/data/WDB).
In this work, we provide a sedimentological, molecular, and microfossil paleoreconstruction study on the southwestern shelf of the Atlantic Ocean during the mid to late-Holocene in a marine sedimentary record. The use of reliable lipid biomarkers, such as n-alkanes, n-alkanols, and alkenones, together with benthic foraminiferal abundance and other foraminiferal indices, yield information about climate, hydrodynamic, and paleoprodutivity related parameters (e.g. sea-surface temperature and the input of terrestrial material to the ocean). This combined approach indicates environmental changes associated with paleoclimatic and paleoceanographic variability as well as the onset of the influence of PPW on the southern Brazilian continental shelf (Cape Santa Marta) for the last 6300 cal yr BP and its influence on the evolution of productivity in a shelf region with a relatively low adjacent river input but subjected to variations in terrestrial and marine contribution rates.
Oceanographic and climatic settings
The São Paulo Bight (Figure 1a) is a morphological feature of the SE Brazilian continental margin extending from Cape Santa Marta (28°30′S–49°00′W) to Cape Frio (23°00′S–42°00′W) (Zembruscki, 1979). This region is characterized by the presence of canyons, channels, and a slope ranging from 120 to 180 m in depth (Furtado et al., 1996; Zembruscki, 1979).
The drainage system of the São Paulo Bight flows toward the continent, with only small rivers reaching the adjacent coast. The two main rivers on the adjacent continent are the Itajaí-Açu River (average discharge of 230 m³ s−1) and the Tijucas River (average discharge of 24.4 m³ s−1) (Branco et al., 2009; Ça Schettini et al., 2010). The sediment input from the bordering continent is limited and modern sedimentation is likely restricted to allochthonous sources, especially from southerly fluvial components such as the LPR (de Mahiques et al., 2004; Razik et al., 2015). Additionally, de Mahiques et al. (2020) reported that the inner shelf region (to the 60 m isobath) is more influenced by the contribution of the PPW.
The variations in the seasonal precipitation over southeast South America are mainly related to the southward expansion and intensification of the SAMS, which in turn is controlled by tropical temperature and pressure gradients between ocean and continent (Vera et al., 2006b; Zhou and Lau, 1998).
During austral summer, the ITCZ is displaced to the south and the larger heat zone over South America moves toward the subtropics. With the higher-pressure gradient between the NW African high- and the South American low-pressure regions, the boreal northeasterly tradewinds, transporting the moisture to the Amazon Basin, are intensified (Vera et al., 2006b). These tradewinds intensify the South American Low-Level Jet (SALLJ) and transport moisture toward SE Brazil. By anticlockwise rotation of the South Atlantic high-pressure cell, the austral tradewinds also transfer moisture from the tropical Atlantic to the southeastern Brazilian coast. These winds contribute to higher convective variability of the South Atlantic Convergence Zone (SACZ) during austral summer and to seasonal maximum precipitation over the LPR basin (Garreaud et al., 2009; Marengo et al., 2004; Razik et al., 2013; Vera et al., 2006a).
The estuary of the LPR is formed by the confluence of the Uruguay River and Paraná River, the latter of which is the major tributary, accounting for approximately 73% of LPR water discharge (Depetris and Pasquini, 2007). The LPR is the second largest river in South America and its drainage area covers an area of approximately 3.1 × 106 km2, representing ~20% of the South American continent (Acha et al., 2008). The combination of the LPR outflow (23,000 m3 s−1) and Patos Lagoon discharge (1750 m3 s−1) mainly constitutes the cold, less saline (<26) PPW (Piola et al., 2000, 2008). In summer, periods of higher river discharge and the predominance of NE winds promote the displacement of the PPW toward the ocean. In contrast, winter conditions are marked by lower water discharge along with the dominance of SW winds, which cause a near coast northward displacement of the PPW (Figure 1c; Depetris and Pasquini, 2007; Piola et al., 2008).
Along the middle and outer shelves of the southwestern Atlantic continental margin, the southerly flow of the Brazil Current (BC) is a decisive feature of sedimentary processes (Campos et al., 2000; de Mahiques et al., 2002, 2004). The BC flow is destabilized by abrupt bathymetric changes due to the shelf limitation and the orientation shift (NE–SW to N–S) in the upper slope, resulting in the formation of meanders and vortices (Campos et al., 2013; Palma and Matano, 2009; Palma et al., 2008).
An upward vertical flux caused by clockwise cyclonic vortices is recognized near Cape Santa Marta and promotes upwelling events of the South Atlantic Central Water (SACW) at the shelf break. This feature injects cold, nutrient-rich waters into the euphotic zone, following the migration of meanders (Brandini et al., 2014; Sato, 2015). Over the austral summer, the dominance of upwelling events favored by northeasterly winds also promotes coastal vertical flux events and intensifies the continuous shelf-break upwelling process (Castelão et al., 2004; Palma and Matano, 2009). The co-occurrence of these two upwelling features generates a SACW uplift on the shelf near the BC (Campos et al., 2013; Castelão et al., 2004) accompanied by surface elevation caused by the offshore migration of the oligotrophic and relatively warmer water mass, Tropical Water (TW), due to the influence of NE winds (Brandini et al., 2014; Palma et al., 2008).
Materials and methods
Sampling
The core 7606 (506 cm sediment record) was collected from the inner shelf of Cape Santa Marta using a piston corer (26°59′16,8″S–048°04′33,6″W) (Figure 1) at a depth of 60 m during a campaign onboard the Professor W. Besnard research vessel in January 2005. The core was subsampled at 2 cm fractions and the samples were frozen for subsequent freeze-drying.
Age model
As well-preserved planktonic foraminifer tests were not available, the age-depth model was based on 11 AMS radiocarbon dates obtained for organic material. About 7 g of bulk sediment were collected at 50 cm intervals and used for AMS-14C dating at Beta Analytics Inc. (Miami, USA). All radiocarbon ages used to produce the age model of core 7606 have been previously published in de Mahiques et al. (2009). Although, in this work, the calibration Marine20 (Heaton et al., 2020) was used for all radiocarbon ages with the aid of the HTML Calib 8.2 software (Stuiver et al., 2020), considering δ13 C values of −26.0‰ and −19.0‰ as reliable end-members (de Mahiques et al., 1999). A regional reservoir age correction (ΔR) of 28 ± 52 was used (Alves et al., 2016). The BACON software, version 2.4.3 (Blaauw and Christen, 2011), was used for age-depth modeling.
Benthic foraminifera
Benthic foraminiferal species were analyzed considering intercalated samples resulting in a temporal spacing of approximately 100 years (approximately 6 cm intervals). Samples of 10 cc were sieved through a >63 µm mesh and then spread on a gridded tray. Grids were randomly examined until at least 300 individual foraminifera were identified. Faunal counts were multiplied to give values for the absolute abundance of each species per 10 cc. Species identification was based on specialized literature (e.g. Barbosa, 1998; Boltovskoy et al., 1980; Loeblich and Tappan, 1988).
The microhabitat of benthic foraminifera is predominantly determined by oxygen and nutrient availability. In the study region, the assemblage was used to infer changes in export productivity and bottom water oxygenation throughout the sedimentary record (García-Gallardo et al., 2017; Jorissen et al., 1995, 2007). Epifaunal taxa are more abundant in environments with nutrient limitations and well-oxygenated regions, whereas infaunal taxa are predominant in regions with high organic content in the sediment (Jorissen et al., 1995, 2007; Rodrigues et al., 2018).
The benthic foraminiferal high productivity index (BFHP) (equation (1)) and benthic foraminiferal oxygen index (BFOI) (equation (2)) were determined to identify periods of high organic carbon supply to the ocean floor and variations in interstitial water oxygen content, respectively (Kaiho, 1994; Martins et al., 2007).

in which P is the density of high-productivity species and T represents total abundance;

in which O and D are numbers of indicator specimens of oxic and dysoxic environments, respectively. Details are presented in the Supplemental Material.
The benthic foraminiferal Shannon species diversity index (H′) and Pielou’s evenness index (J′) were calculated based on Martins and Santos (1999). Higher values of H′ and J′ suggests higher species diversity and equitability, respectively.
Species classification (R-mode) was performed using a frequency matrix of representative benthic foraminifera (species that appear in at least 10% of samples and with abundance >2%) and the weighted pair group method with arithmetic mean (WPGMA) by hierarchical groupings with a Bray-Curtis similarity coefficient (Landim, 2011). All analyses were carried out using the Multivariate Statistical Package (MVSP) software, version 3.1 (Kovach, 1999).
Lipid biomarkers
Lipid biomarkers were analyzed considering intercalated samples based on time intervals of approximately 100 years (approximately 6 cm intervals). The extraction and cleanup steps as well as instrumental determination parameters are detailed in previous studies (Lourenço et al., 2016, 2017). Briefly, sediment sample aliquots (3 g) were spiked with a surrogate mixture (n-hexadecene, n-eicosene, and 5α-androstanol), extracted using an ultrasonic bath (3 × 15 mL n-hexane/dichloromethane (DCM) 3:1 (volume/volume) 30 min each), and concentrated in a rotary evaporator. The extracts were fractionated by adsorption chromatography using an open column containing silica (2 g) and alumina (1 g) into n-alkanes and alkenones using 8 mL of n-hexane followed by 6 mL of DCM/n-hexane (8:2, v/v) and into n-alkanols using 15 mL of methanol.
The n-alkanes and alkenones were analyzed by gas chromatograph with a flame ionization detector (GC-FID). The n-alkanols were dried and derivatized with BSTFA (bis (trimethylsilyl) trifluoroacetamide) and 1% TMCS (trimethylchlorosilane) for 90 min at 65°C and analyzed by gas chromatograph coupled to a mass spectrometer detector (GC-MS). Analytical standards were used for identifications and to create analytical curves. Quantification was based on the internal standard (IS) method, using n-tetradecene for n-alkanes and alkenones and 5α-cholestane for n-alkanols.
Procedural blanks were determined for each compound class and no peaks interfered in the target compounds analysis. The spiked blank analysis was also conducted and presented recovery rates of n-alkanes, n-alkanols ranging from 60% to 91% and 80% to 98%, respectively. Surrogate recovery ranged from 51% to 96%. The method detection limit was based on the standard deviation of seven blank replicates and ranged from 0.002 to 0.032 μg g−1 for n-alkanes and alkenones and 0.005–0.012 μg g−1 for n-alkanols.
Odd long-chain n-alkanes (C27, C29, and C31) and even long chain n-alkanols (C24OH, C26OH, C28OH, and C30OH) are components of epicuticular waxes from the leaves of vascular plants (Eglinton and Hamilton, 1967; Mudge and Norris, 1997) and were used in this study as proxies of terrigenous material input into the marine sediment.
The carbon preference index (CPI; the ratio of odd and even long-chain n-alkanes in the range of C24–C34) (equation (3)) can also be used as a proxy to evaluate major contributions of terrestrial plants to the organic matter of marine sediment (Bray and Evans, 1961; Bush and McInerney, 2013; Clark and Blumer, 1967; Herrera-Herrera et al., 2020).
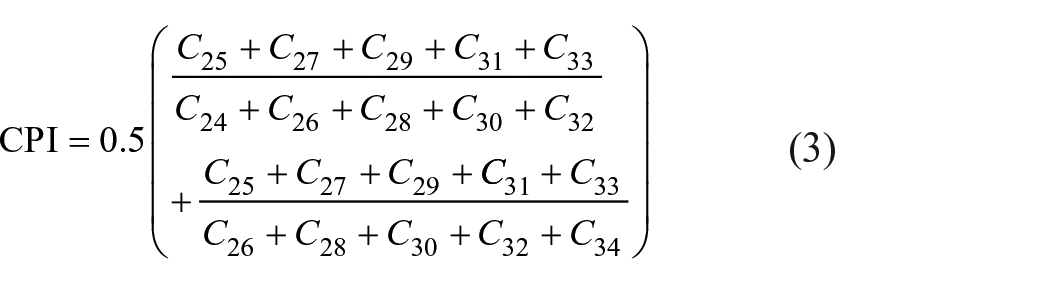
When exposed to high temperatures, plants tend to synthesize longer chain waxy constituents, which enables associating changes in chain-length in plant n-alkanes with environmental shifts of temperature and humidity in the region of the vegetation source (Poynter and Eglinton, 1991; Poynter et al., 1989). Therefore, changes in the average chain length of terrestrial n-alkanes (ACL27–33; equation (4)) and the predominance relation of C29 and C31 n-alkanes using the alkane index (AI) (equation (5)) can be used to infer changes in vegetation cover over distinct climatic intervals (Horikawa et al., 2010; Rommerskirchen et al., 2006; Schefuß et al., 2003). These indices may also be used to distinguish between terrestrial origins introduced by C3 plants (woody plants or tropical forest) and those produced by C4 plants (grasslands) (Badewien et al., 2015; Cranwell, 1973; Zhang et al., 2006). While C3 plants tend to produce higher amounts of C27 and C29, C4 grasses maximize at C31 and C33 (Damsté et al., 2011; Zech et al., 2010).
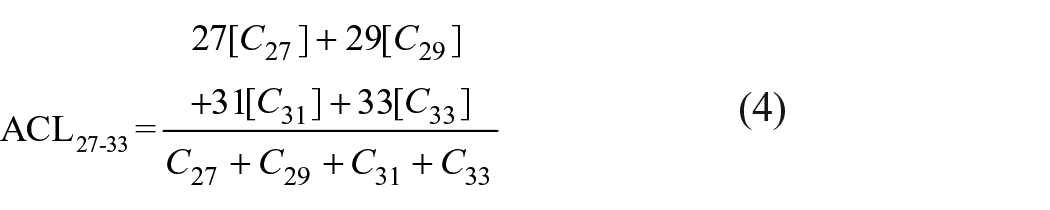
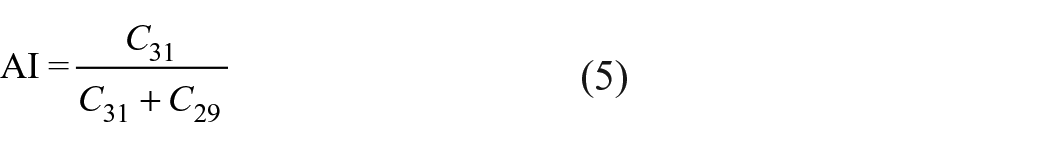
Sea-surface temperature (SST) values were estimated based on abundances of C37:2 and C37:3 alkenones (chromatographic signals) and the UK′37 index (SST-UK′37) (Brassell et al., 1986), using the global calibration given by Müller et al. (1998).
All the lipid biomarkers concentrations were normalized by TOC to avoid misinterpretation caused by variations in sedimentation rates (de Mahiques et al., 2020; Muri et al., 2004; Schulte et al., 2000).
Results
Age model
The age model of the core 7606 obtained in this study is relatively divergent from the age model proposed by de Mahiques et al. (2009). This difference is probably related to the use of a more recent and improved marine radiocarbon calibration curve in this work. de Mahiques et al. (2009), using the Marine04 calibration curve (Hughen et al., 2004), reported a time range from 860 to 7660 cal yr BP and sedimentation rates varying from 0.025 to 0.250 cm yr−1. In this work, using the Marine20 calibration curve (Heaton et al., 2020), the core 7606 represents a time range from 6300 to 890 cal yr BP and sediment accumulation rates ranged from 0.13 to 0.35 cm yr−1. From 6300 to 4000 cal yr BP sedimentation rate was relatively low. An increase is observed between 4000 and 3200 cal yr BP, with sedimentation rates remaining high until 2400 cal yr BP, where a decrease is observed. After 2000 cal yr BP an increase in the sedimentation rate is observed until the core top (890 cal yr BP) (Figure 2). The results of the 14C dating are shown in Table 1.
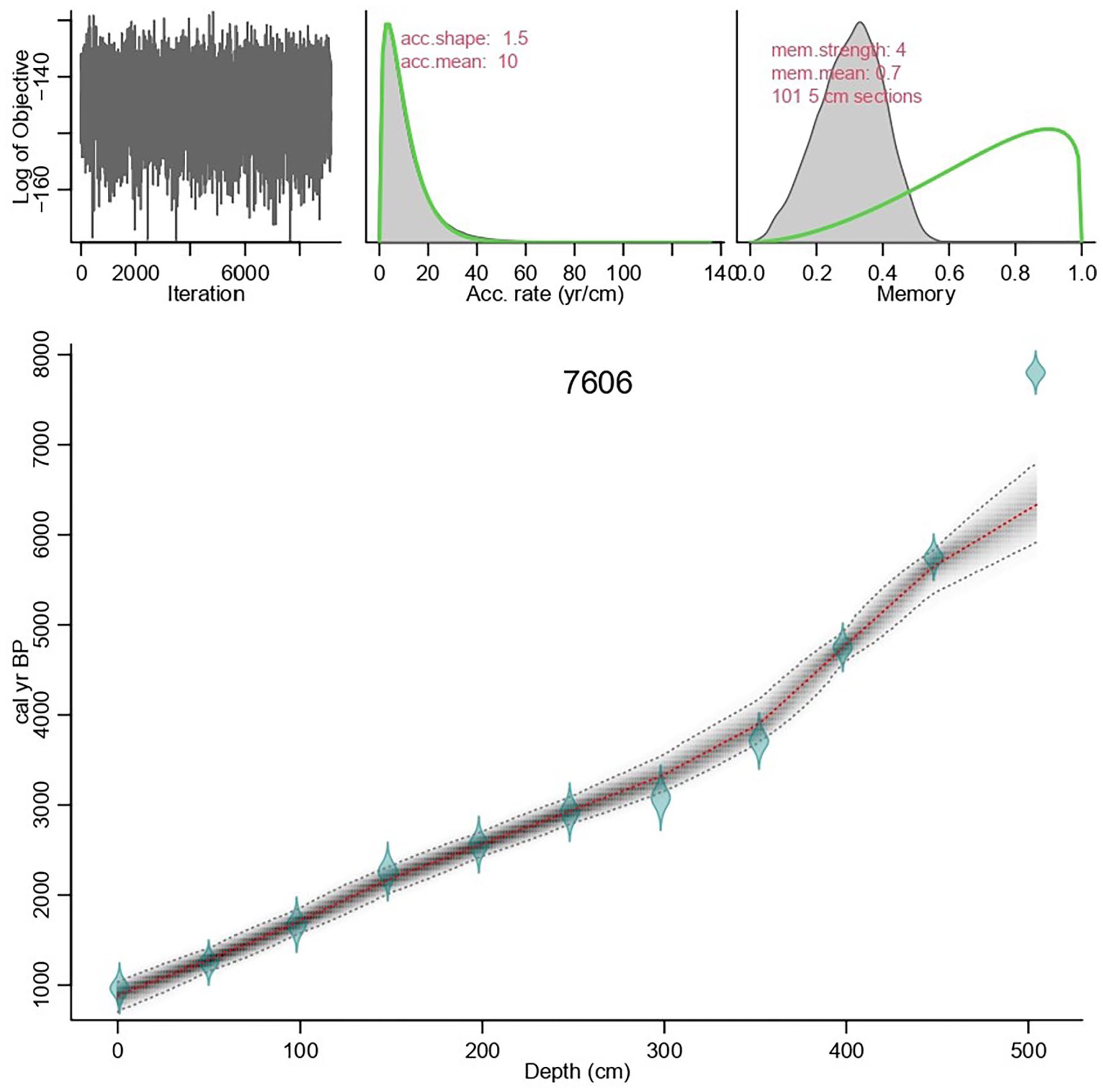
Age-depth model for Core 7606 produced by Bacon software (Blaauw and Christen, 2011). Upper graphs show Markov Chain-Monte Carlo (MCMC) iterations (left), accumulation rate (middle), and memory R (right). Green line and gray area indicate prior and subsequent distributions of each parameter, respectively. The bottom graph represents the age-depth model (gray) containing calibrated distributions of specific dates (blue). The 95% probability intervals are symbolized by the gray dashed lines and the weighted mean “best” fit curve (age for each depth) represented in red.
AMS radiocarbon dating results in organic matter of selected samples from core 7606.
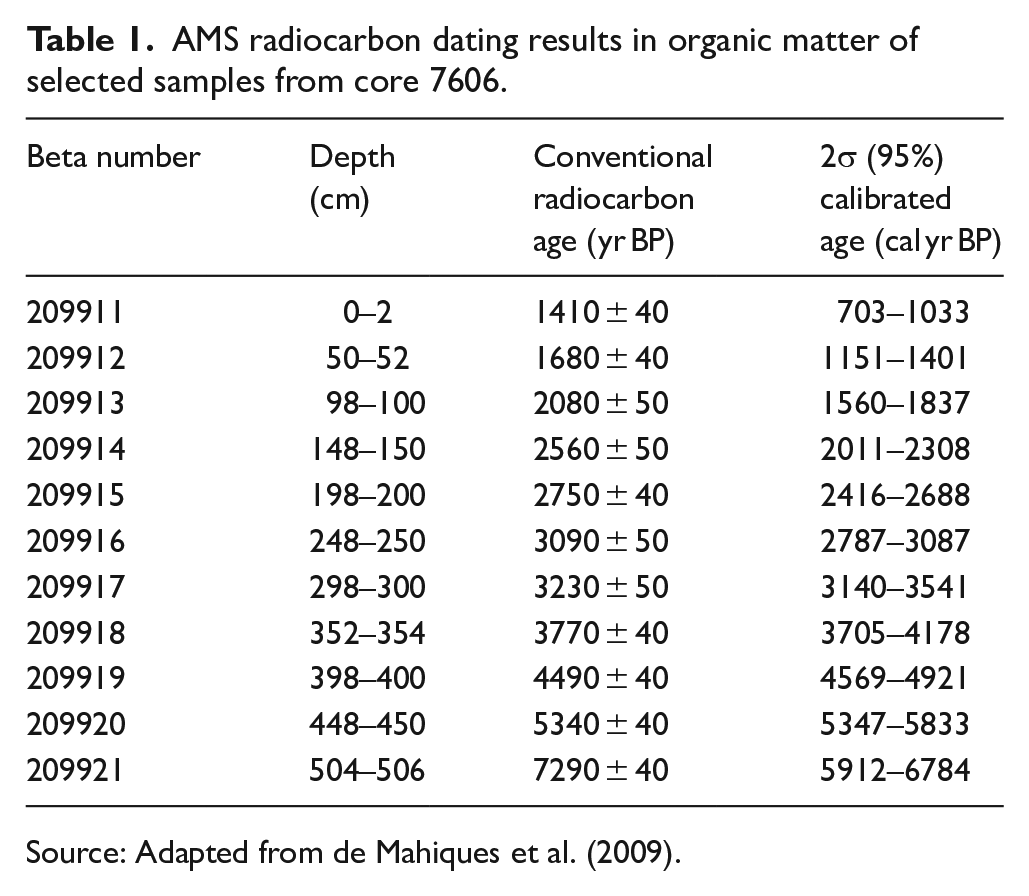
Source: Adapted from de Mahiques et al. (2009).
Bulk parameters and benthic foraminifera
The grain size and TOC results of the 7606 core were previously published by de Mahiques et al. (2009). In this work, we reanalyze these results as accessory data to the benthic foraminifera and lipid biomarkers proxies.
Fine sediments (silt + clay) were predominant throughout the core and ranged from 58% to 95%, with the silt fraction as the main component (77%–88%) (Figure 3a). Coarser sediments were more frequent in the period between 6300 and 5400 cal yr BP, followed by an increasing fine sediment trend, with the highest percentage after 2700 cal yr BP. TOC content ranged from 0.5% to 1.1%, with a mean of 0.7% ± 0.2, showing a continuous increase along with the core with higher content at the core top (Figure 3c).
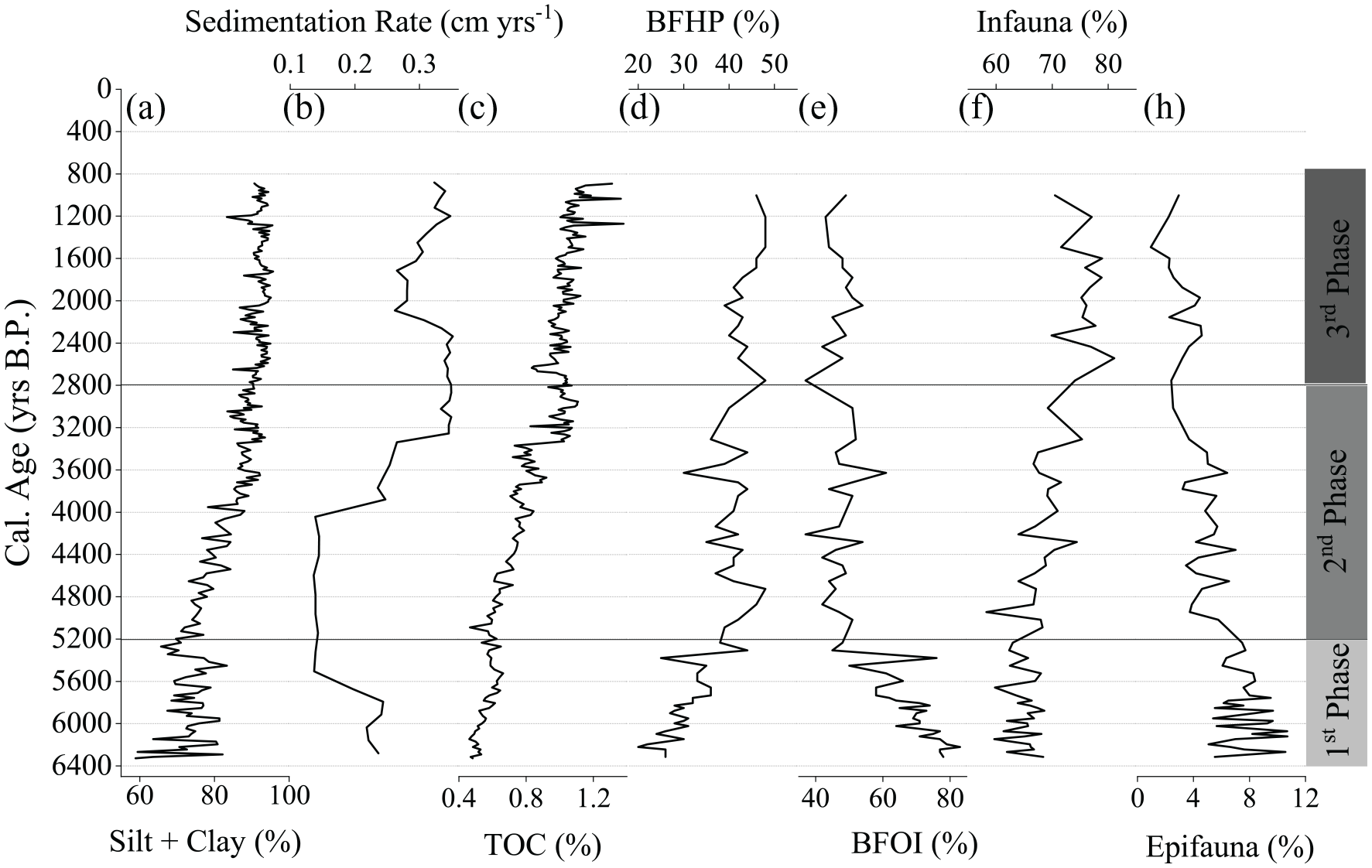
Along-core distribution of bulk parameters ((a) Silt + clay, (b) Sedimentation rates, and (c) TOC), (d) Benthic foraminifera indices (benthic foraminiferal high productivity (BFHP), (e) Benthic foraminiferal oxygen index (BFOI)), (f) Frequencies of epifaunal, (h) infaunal taxa. Silt + clay, sedimentation rate, and TOC results adapted from de Mahiques et al. (2009).
A total of 67 species and 39 genera of benthic foraminifera were identified in the samples from core 7606, with relatively high abundance of Globocassidulina subglobosa (8%–30%), Globocassidulina spp. (4%–22%), Uvigerina peregrina (4%–16%), Bulimina marginata (1%–13%), Bulimina spp. (2%–13%), and Buliminella elegantissima (0%–18%) (Figure 4). The results are comparable with data from previous studies conducted in the region (Burone et al., 2011; Eichler et al., 2008). These species are associated with a high, continuous flow of organic matter (B. marginata, Mojtahid et al., 2009, 2013), the episodic input of phytodetritus (G. subglobosa, García-Gallardo et al., 2017; U. peregrina, de Mello e Sousa et al., 2006), low to moderate oxygen depletion in the bottom and pore water (G. subglobosa; U. peregrina), well-oxygenated waters (Zarriess and Mackensen, 2010), and strong bottom currents (Mackensen et al., 1995; Schmiedl et al., 1997) and were used in the evaluation of BFHP and BFOI indices.
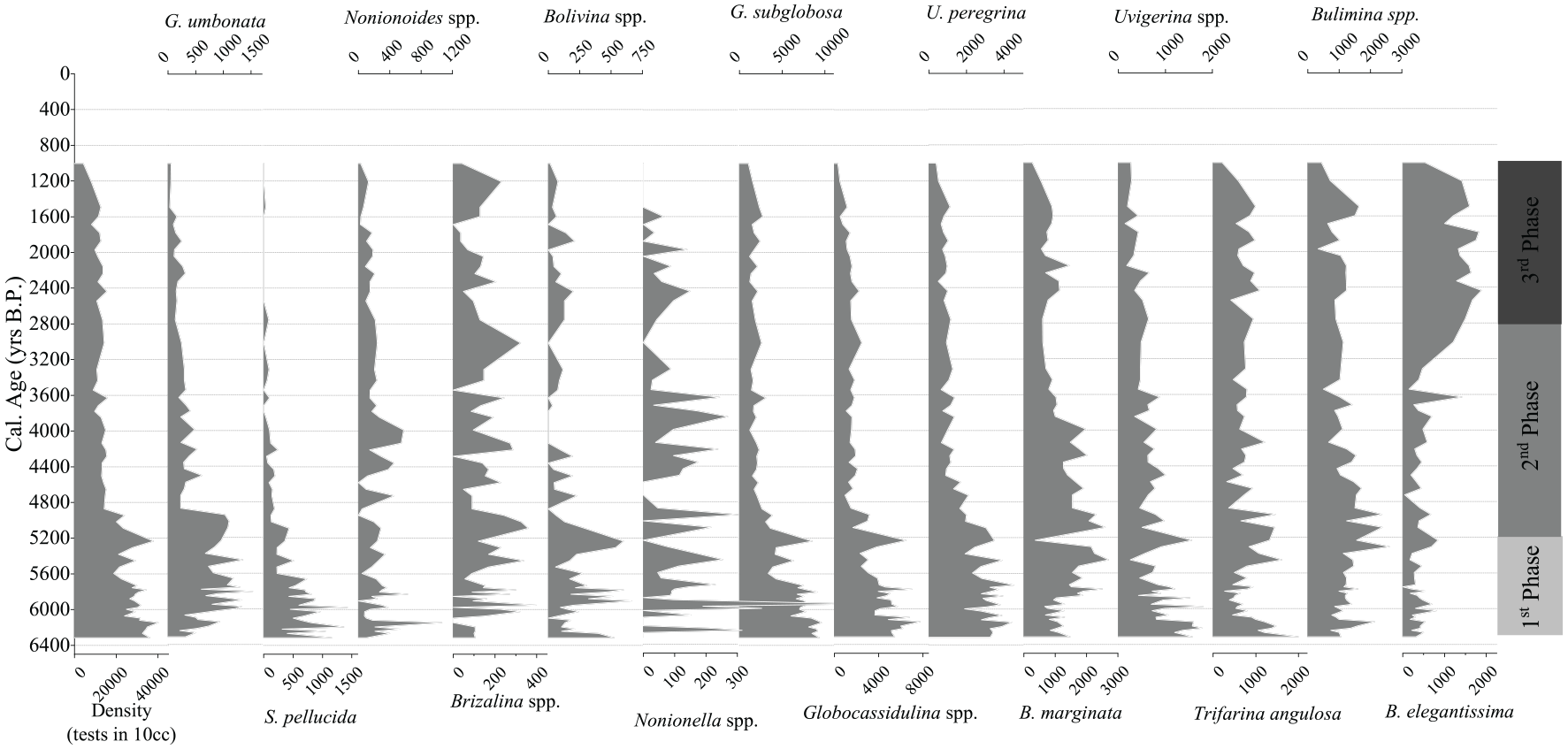
Along-core distribution of the density and the most abundant species of benthic foraminifera in the four main assemblages.
Only 10 out of the 67 identified species were not classified as epifaunal or infaunal due to a lack of bibliography. However, their relative contribution values do not exceed 2%. Among the classified species, 19 and 38 were considered epifaunal and infaunal, respectively (Martins et al., 2007). The distribution of benthic foraminifera revealed a general dominance of infaunal species (abundance varying from 58% to 81%), whereas epifaunal species did not exceed 11% (Figure 3f–h).
A gradual decreasing pattern of epifaunal species was found from 6300 to 890 cal yr BP, with the lowest contribution after 3500 cal yr BP (Figure 3h). The lowest relative abundances of infaunal species were found between 6300 and 5200 cal yr BP. The infaunal taxa exhibited a continuous increase between 5200 and 890 cal yr BP, with higher relative abundances after 2800 cal yr BP (Figure 3f).
The BFHP index ranged from 20% to 48% and exhibited a sharp increase between 6300 and 5200 cal yr BP. After this date, a slight increase was found throughout the core, with the highest values from 3000 to 890 cal yr BP (Figure 3d). An inverse trend was found for BFOI, ranging from 37% to 83%, with a strong downward trend from 6300 to 5200 cal yr BP. A slightly decreasing value is found from 5200 cal yr BP to 890 cal yr BP, with the interval between 5200 and 2800 cal yr BP exhibited higher values than after 2800 cal yr BP (Figure 3e). The high BFOI values indicate oxygen-rich waters, which can hinder the preservation of benthic foraminifera. Therefore, the BFHP for paleoproductivity should be used with caution. Analyses of the benthic foraminiferal assemblage were used to corroborate productivity results.
The combined >63 µm size portions were used in the R-mode cluster analysis, which revealed four main assemblages designated according to the dominant species: G. umbonata, Nonionoides spp., G. subglobosa, and B. marginata (Supplemental Figure S1).
The G. umbonata assemblage was composed of G. umbonata and Seabrookia pellucida. This assemblage occurred mainly near the core bottom and frequencies did not exceed 6% (Figure 4).
The Nonionoides spp. assemblage was composed of Nonionoides spp., Nonionella spp., Bolivina spp., and Brizalina spp. and also had species with low frequencies (<4%).
Brizalina spp. and Nonionella spp. exhibited a slightly decreasing trend throughout the core.
The G. subglobosa assemblage was composed of G. subglobosa, Globocassidulina spp., and U. peregrina. The highest relative abundances of these species were found between 6300 and 5000 cal yr BP (Figure 4).
The Bulimina marginata assemblage included B. marginata, B. elegantissima, Trifarina angulosa, Bulimina spp., and Uvigerina spp. Higher relative abundances were found between 6300 and 4000 cal yr BP for all species, except B. elegantissima, which had higher relative abundances in the last 3000 cal yr BP (Figure 4).
The lowest H′ and J′ ecological index values were found between 6300 and 5200 cal yr BP, although with an increasing pattern with the highest values at 4000 cal yr BP. After this date, a decreasing trend was also found between 3800 and 890 cal yr BP (Supplemental Figure S2).
Lipid biomarkers
The chain length distribution of n-alkanes and n-alkanols in core 7606 ranged from C12 to C35 and C14OH to C30OH, respectively. Both compound series exhibited unimodal distribution, maximizing at C29 or C31 for n-alkanes and C26OH or C28OH for n-alkanols.
Alkanes25–33/TOC ranged from 0.20 to 0.53 µg g−1/TOC and exhibited considerable variability, with higher concentrations between 5000 and 4200 cal yr BP and 3000 and 2600 cal yr BP (Figure 5b). Alkanols24–30/TOC exhibited presented the lowest values between 6300 and 5200 cal yr BP. At 5000 cal yr BP, the alkanols24–30/TOC values increased, remaining without substantial variability until 2800 cal yr BP. After this date, an increase in the alkanols24–30/TOC is observed until 890 cal yr BP. Alkanols24–30/TOC values ranged from 0.27 to 2.34 µg g−1/TOC, with higher concentrations after 2400 cal yr BP (Figure 5a). As the variations in TOC content in the sediment core were smaller than the variations in the concentrations of lipid biomarkers, the normalized TOC data were very similar to the non-normalized results. It can therefore be inferred that the preservation/degradation of organic matter induced by sedimentation may not have been the prevailing aspect regulating the deposition and accumulation of organic matter and that the flow of autochthonous/allochthonous marine and terrigenous organic matter played a more important driving role (Stein, 1990).
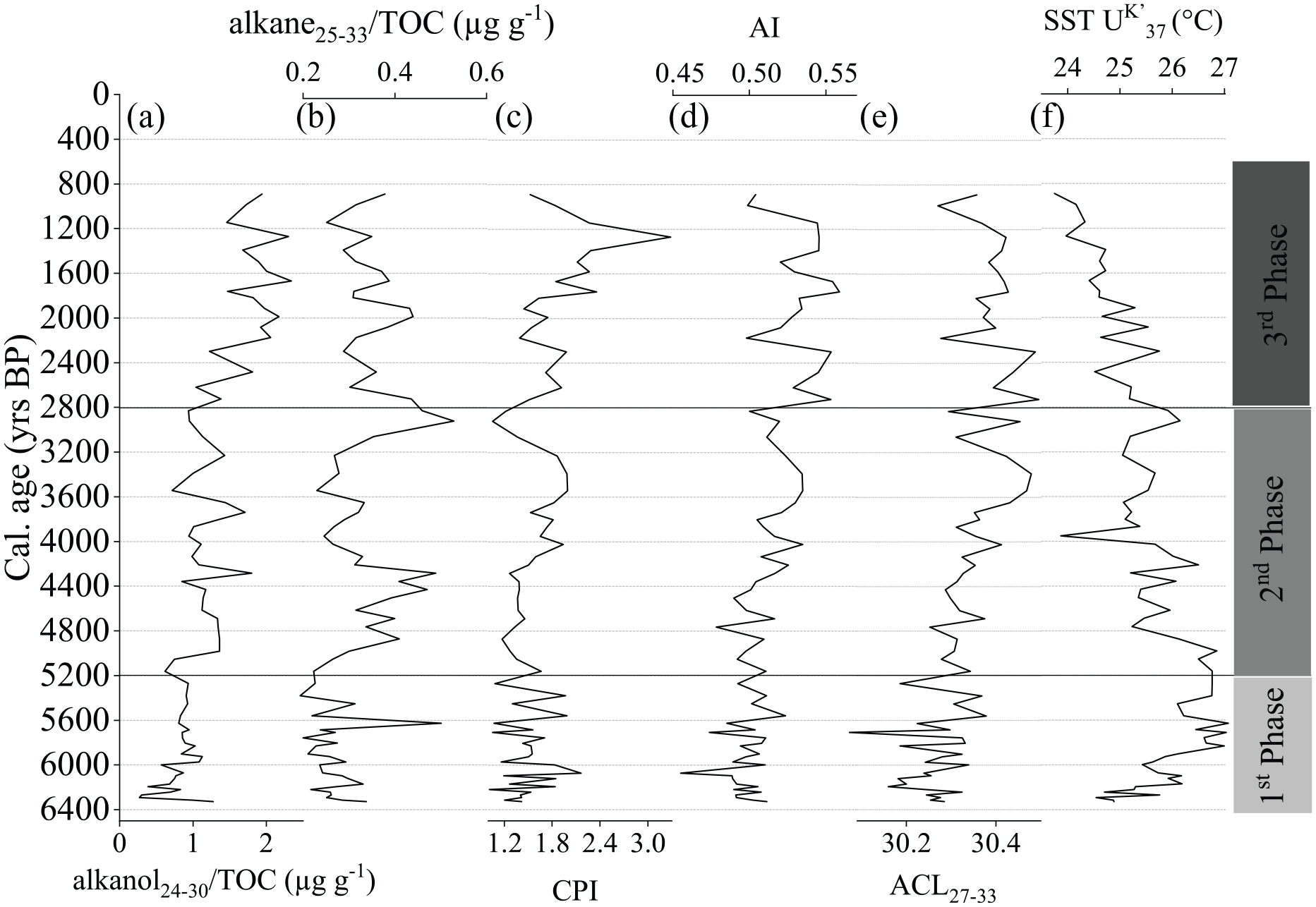
Along-core distribution of (a) alkanol24–30/TOC, (b) alkanes25–33/TOC, (c) Carbon preference index (CPI), (d) Alkane index (AI), (e) Average chain length (ACL27–33), (f) SST-UK′37.
CPI values ranged from 1.05 to 3.28. Relatively lower CPI values were found from 6300 to 4200 cal yr BP. This was followed by an increase, with the highest value reached at 1200 cal yr BP, indicating a continuous increase in terrigenous input between 4200 and 1200 cal yr BP (Figure 5c).
ACL27–33 values ranged from 30.07 to 30.41, with a mean of 30.32 ± 0.08. Despite the low range values within the core, a markedly increasing signature was found after 4000 cal yr BP, with higher values between 2800 and 890 cal yr BP (Figure 5e). AI values ranged from 0.49 to 0.55 and exhibited similar trends as those found for ACL27–33 (Figure 5d and e). Between 6300 and 5000 cal yr BP, AI values were ⩽0.5. An increasing trend was found from 5000 to 890 cal yr BP, with values >0.5. The highest AI values were found between 2800 and 890 cal yr BP, although showing a very slight decrease between 2200 and 890 cal yr BP. These results may indicate a shift in the regional input of terrestrial organic matter starting at ~5000 cal yr BP and its establishment at ~2800 cal yr BP.
SST-UK′37 fluctuated around 3.3°C throughout the core (Figure 5f). The period between 6300 and 5000 cal yr BP had the highest SST-UK′37 values, with an increase from 24.8°C to 26.7°C. A continuous decrease from 26.9°C to 23.7°C was found between 5000 and 890 cal yr BP, with a more markedly decreasing trend from 3000 to 890 cal yr BP.
Discussion
The molecular, micropaleontological, and sedimentological analyses performed in the present study enabled the recognition of three hydrographic phases for the southwestern Atlantic shelf during the mid- to late-Holocene (first phase: 6300–5200 cal yr BP; second phase: 5200–2800 cal yr BP; third phase: 2800–890 cal yr BP) (Figure 6). These results suggest the influence of synchronized variations in sea level, precipitation over the Paraná River drainage basin, and the northward displacement of the PPW by the BCC, transporting sediments from southern areas to the core 7606 region.
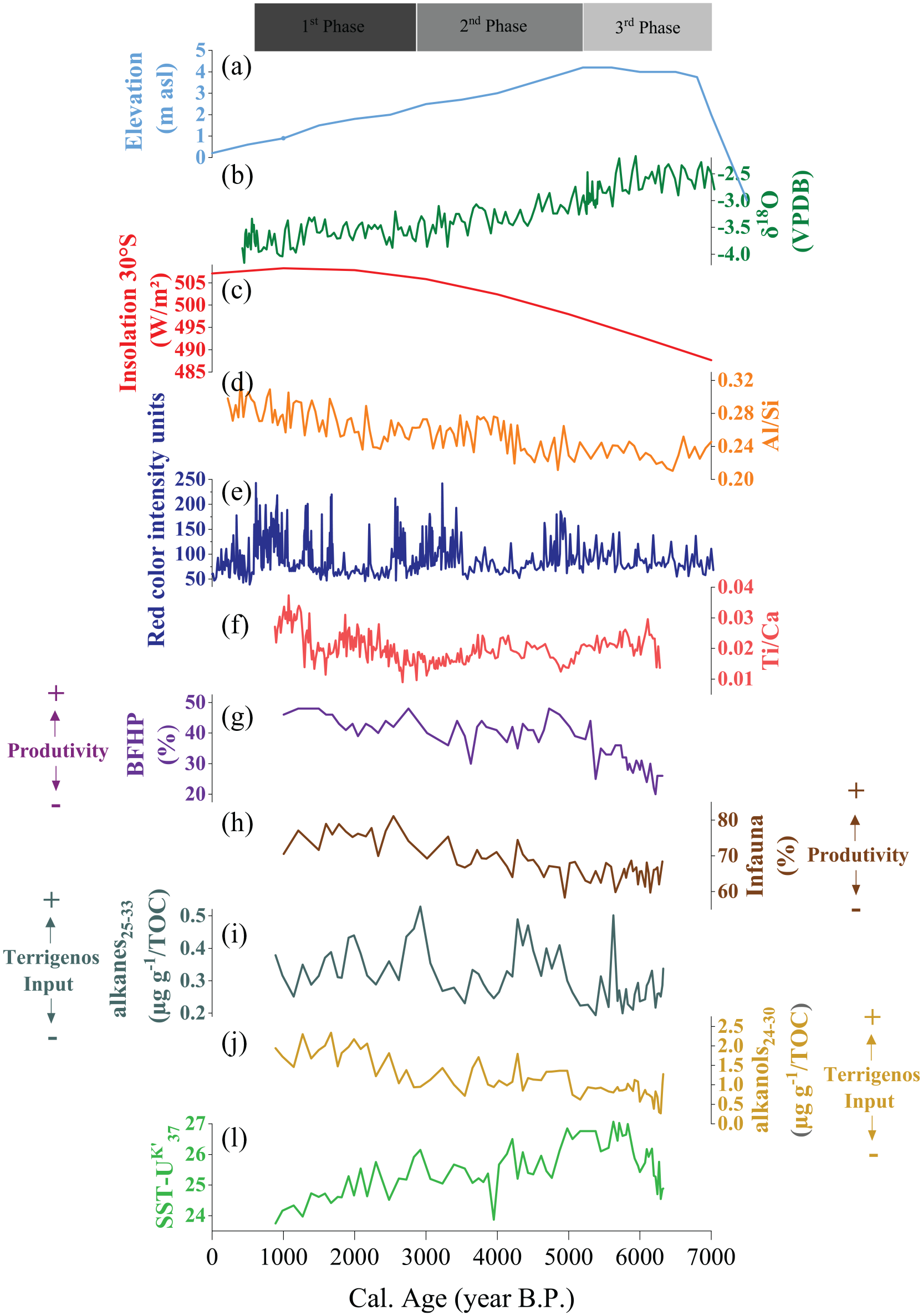
Proxy records of climatic and relative sea level changes in southeastern South America. (a) Relative sea-level (RSL) variation curve for La Plata River estuary (Prieto et al., 2017). (b) Stalagmite oxygen isotope record (δ18O) from Botuverá Cave (Bernal et al., 2016; Cruz et al., 2005). (c) Precession-driven February austral summer insolation (30°S) (Berger and Loutre, 1991). (d) Western South Atlantic, GeoB6211 core (Chiessi et al., 2010). (e) Pallcacocha Lagoon sediment red color intensity (25-point average plotted) (Moy et al., 2002) and (f) Core 7606 Ti/Ca ratio (de Mahiques et al., 2009); (g) benthic foraminifera high productivity index (BFHP). (h) Infauna. (i) Alkanes25–33/TOC. (j) Alkanols24–30/TOC. (k) Sea surface temperature from alkenones (SST-UK′37) for Core 7606.
Sea level
The variations in relative sea level (RSL) on the southeastern South America continental shelf throughout the Holocene played a crucial role in the variability of marine sediment transport and deposition, especially considering the extensive sediment discharge of the LPR estuary, its large size, and the smooth slope of the delta (de Mahiques et al., 2009; Gyllencreutz et al., 2010; Lantzsch et al., 2014; Prieto et al., 2017). Using 14C-calibrated ages, Gyllencreutz et al. (2010) re-plotted the RSL curve reported by Cavallotto et al. (2004) for the Holocene in the southwestern LPR. This curve presents shoreline transgression until 6500 cal yr BP and shows a maximum RSL of 6.5 m, followed by a discontinuous lowering pattern until the present. However, Prieto et al. (2017) proposed a distinct Holocene RSL curve using 52 calibrated 14C data from 36 different stations along the coast of the LPR estuary, indicating a maximum highstand reaching 4 m above the present RSL at 5800–5400 cal yr BP, followed by a gradual lowering toward the present (Figure 6a). This RSL curve is in agreement with the Holocene RSL curve proposed by Angulo et al. (2006) and de Freitas Toniolo et al. (2020) for the coastline of southern and southeastern Brazil as well as by Martínez and Rojas (2013) for the Uruguayan coast.
A relatively suppressed terrigenous signal (i.e. alkanol24–30/TOC, alkanes25–33/TOC, and CPI) was found in core 7606 between 6300 and 5200 cal yr BP (Figure 6h and i). This finding is probably related to the highstand of the RSL 4 m above the modern sea level, trapping sediment from the Paraná drainage sediments in the LPR estuary (Gyllencreutz et al., 2010; Prieto et al., 2017). During the maximum RSL, the sediments that deposited on the Brazilian continental shelf were likely derived from the Argentinean continental shelf and were transported to a northward position by a mixing of the BCC and the Patagonian Current (Mathias et al., 2014).
With the RSL lowering, a gradual change in the environmental dynamics is suggested by our data of terrigenous input (e.g. n-alkanols24–30/TOC, alkanes25–33/TOC, CPI, AI, and ACL27–33), marine productivity (e.g. BFHP and Infauna taxa) as well as surface paleotemperature (e.g. SST-UK′37). Our results indicate a progressive influence of the PPW established after 5200 cal yr BP and its dominant influence after 2800 cal yr BP. Fluvial sedimentation was reported in the south of the LPR mouth and attributed to the RSL lowering after 5000 cal yr BP (Laprida et al., 2007). Mathias et al. (2014), using magnetic particles, reported similar interval periods for the PPW influence on the São Paulo Bight (25°S).
Climate changes
The wind regime is a determinant factor of changes in the moisture conditions of the environment in southeastern South America (Behling, 1997, 1998; Bernal et al., 2016; Ledru et al., 1996, 1998; Razik et al., 2013; Wanner et al., 2008). During the mid- to late-Holocene, variations in the wind regime were related to oscillations in the location of the ITCZ and subtropical shifts caused by the wind gradient associated with front migration. Moreover, the intensity of the subtropical westerlies may have also exerted an influence and could be associated with different phases of the annular mode (de Mahiques et al., 2009). Most of the variations in the amount of total precipitation in southeastern South America during the Holocene have mainly been driven by the intensity of the South American Monsoon System (SAMS) over the last 9000 years (Bernal et al., 2016). Insolation also played an important role, particularly in the mid-Holocene, when the North Atlantic was warmer, possibly weakening the SAMS (Brierley and Wainer, 2018; Shimizu et al., 2020). An intensification of the rainfall regime in the southeastern portion of South America during the mid- to late-Holocene was also described by Bernal et al. (2016), which found a gradual decrease in δ18O and Sr/Ca since 7000 cal yr BP in the Botuverá Cave (Figure 6b) (27°13′S–49°09′W). The authors found an increase in the intensity of the SAMS throughout the Holocene, resulting in higher precipitation, with its long-term optimum stating at 4000 cal yr BP when a phase of greater summer insolation was found in the Southern Hemisphere. This trend is also consistent with that described by Cruz et al. (2007) in an analysis of monsoon precipitation records from the Amazon region after 4000 cal yr BP, supporting the migration of greater moisture to the south and southeast Brazil from distal sources (Cheng et al., 2013).
Our results show a relatively lower terrigenous input between 6300 and 5200 cal yr BP in the core 7606. The highstand of the RSL in this interval is probably the main cause of the low terrigenous signal, although, a drier and warmer climate was also found for Southeast Brazil between 7000 and 4000 cal yr BP (Bernal et al., 2016; Ledru et al., 1996). The multi-proxy data compilation of rainfall in South America for the mid-Holocene (Prado et al., 2013) and the analysis of the Paleo Model Intercomparison multi-model simulation results (Brierley and Wainer, 2018) indicate a relatively drier southeastern portion of South America together with the intensification of rainfall in the northern portion of the continent consistent with a warmer North Atlantic relative to the south.
Relatively lower AI and ACL27–33 values were found between 6300 and 5200 cal yr BP. Previous studies reported the presence of woody vegetation on the continent adjacent to our coring site (24°S) (Behling, 2002; Behling and Negrelle, 2001; Gu et al., 2017) as well as the presence of grasslands in the southern regions of South America (30°S) (Contreras and Zucol, 2019; Gu et al., 2018; Mourelle et al., 2017; Rodrigues et al., 2016). Therefore, if AI and ACL27–33 are considered representative of the C3 or C4 vegetation type (Kouli et al., 2012; Schefuß et al., 2003; Vogts et al., 2009), the relatively lower values of AI and ACL27–33 indicates the influence of a mixture of woody vegetation from the adjacent continent, possibly transported through the Itajaí-Açu River, which provided local sediments over the mid- to late-Holocene once estuarine infilling was accomplished (Hesp et al., 2009; Lessa et al., 2000), and from grasslands from southern regions fluvial sources.
Analyzing trace metals in core 7606, de Mahiques et al. (2009) reported the lowest terrigenous input in the interval from 5000 to 3000 cal yr BP (Figure 6f). According to the authors, these results were related to the weaker intensity of the Southern Westerly Wind Belt and probable lower moisture conditions in the southern portion of South America. However, the present study results for the interval between 5200 and 2800 cal yr BP indicate a relatively slightly higher input of terrigenous material compared to the previous phase.
Some studies reported a highly variable precipitation regime from 5000 to 3000 cal yr BP, with abrupt changes indicating strong events of increased precipitation correlated with cooling SST events in the North Atlantic throughout the Holocene (Bernal et al., 2016; Strikis et al., 2011). The SAMS intensified over the southeastern portion of South America and the onset of the modern ENSO conditions was established at 5000 cal yr BP, causing strong events of intense discharge over the Paraná River basin (Figure 6e; Bender et al., 2013; Mourelle et al., 2015; Moy et al., 2002; Rein et al., 2005). The outflow of the LPR, associated with the lowering of the RSL observed after 5000 cal yr BP, probably indicates the onset for the last 6300 cal yr BP of the influence of the PPW over the southwestern Atlantic margin and explains the relatively higher terrigenous influence in this phase. The large sediment load discharge during El Niño events and subsequent stronger S/SE wind-driven currents caused by La Niña induced the transport of the PPW to the southern Brazilian shelf (Chiessi et al., 2010; de Souza and Robinson, 2004; Gyllencreutz et al., 2010; Möller et al., 2008; Piola et al., 2005).
At 5200 cal yr BP, the increasing values of ACL27–33 started, and indicated a shift in the source of hinterland vegetation sediment, with a major contribution from grasslands or dry environment vegetation (Figure 5d and e). The greater contribution of grasslands also agrees with the reach of the PPW acting as a transport mechanism to the inner shelf off Cape Santa Marta, as a stronger expansion of the Atlantic lowland rainforest took place on the adjacent continent after 5500 cal yr BP (Coe et al., 2014; Gu et al., 2018) and the LPR drainage basin is characterized by the higher contribution of grassland vegetation (Gu et al., 2018; Mourelle and Prieto, 2012).
The alkanes25–33/TOC showed higher variability and relatively higher values between 4800 and 4200 cal yr BP and 3000 and 2700 cal yr BP, although no explicit explanation was found for these high values, intensified ENSO activities were reported for these periods (Figure 6e; Moy et al., 2002). Therefore, we may suggest that alkanes25–33/TOC in these intervals better represent intense ENSO events periods, which cause intense rainfall in the Itajaí-Açu and the LPR river basin. However, further studies with higher temporal resolution should be carried out to evaluate the alkanes-ENSO relation.
The increase in the lipid biomarker terrigenous data and diagnostic indices, especially the alkanols24–30/TOC and CPI, suggests a gradual increase in total rainfall coeval with the gradual drop in RSL and the influence of the S/SW wind regime over the southeastern portion of South America (Figure 6). These climatic and oceanographic features are detectable since ~5200 cal yr BP but are more evident between 2800 and 890 cal yr BP, although with less variability (Figure 6i and j).
After 2800 cal yr BP, in addition to the long-term intensification of the SAMS over the southeastern portion of South America (Bernal et al., 2016), the potential for westerly wind anomalies were amplified, causing a greater amplitude of the ENSO (Haug et al., 2001; Koutavas et al., 2006). The timing of the greater terrigenous influence (after ~2000 cal yr BP) observed in the alkanols24–30/TOC, alkanes25–33/TOC and CPI is consistent with the increased precipitation in southern Brazil and the Paraná River drainage basin for the past ca. 2000 years (Bernal et al., 2016; Cruz et al., 2009; Mourelle et al., 2017; Woodroffe et al., 2003) and since 1700 cal yr BP in Southern Uruguay (Bracco et al., 2005). Chiessi et al. (2010) also reported an increase in the terrigenous signal in this interval in the GeoB6211-2 marine sediment core (32°S) (Figure 6d).
A concomitant increase in both El Niño and La Niña activity was observed during the late-Holocene and suggests centennial-scale changes in ENSO activity driven by tropical insolation (Makou et al., 2010; Wanner et al., 2008). The AI and ACL27–33 exhibited stabilization. These results may indicate the modern conditions of wind and riverine input, the low sea level, and a further enhanced El Niño amplitude followed by S/SWE wind during La Niña events, which indicates the dominant influence of the PPW transported by the BCC on sedimentary processes on the inner shelf off southern Brazil.
Paleoprodutivity
The paleoproductivity dynamic between 6300 and 5200 cal yr BP was represented by low organic matter export to the seafloor, as suggested by the low BFHP values and TOC content, and high epifaunal relative abundance (Figure 3). These results are probably related to the absence of the PPW influence, the primary controlling feature of the seasonal variation in marine production, agreeing with the low terrigenous signal between 6300 and 5200 cal yr BP. This interval was also characterized by intense hydrodynamic and more oxygenated bottom water conditions (indicated by the high BFOI values), low indices of fine sediments, and higher G. subglobosa abundance (Figures 3 and 4). The strong bottom water current possibly hampered the deposition of fine sediments and the accumulation of organic matter in the core site region. Studies on the southeastern Brazilian margin associate the opportunistic G. subglobosa to oligotrophic environments with pulsed organic matter on the seafloor and well-oxygenated bottom waters in areas with a strong bottom current (Eichler et al., 2008, 2016; Paquette et al., 2016).
The interval between 6300 and 5200 cal yr BP also presents a relatively higher total abundance of benthic foraminifera, which at first should indicate a more productive environment. However, in this interval, only three species represent >50% of the total abundance of benthic foraminifera (G. subglobosa, U. peregrina, and Globocassidulina spp.). These results, in addition to the low diversity (H′) and equitability (J′), show the dominance of few species, especially G. subglobosa, know as being opportunistic and better adapted to more intense hydrodynamic conditions and sandy substrate, reached high densities in this period.
Studies carried out in oligotrophic regions indicated the benthic foraminifera species diversity and the TOC supply as related parameters (Gooday and Rathburn, 1999; Kurbjeweit et al., 2000). Although, in the presence of intense phytodetritus input, the increase in metabolic activity of foraminifera communities may influence their oxygen consumption and diversity (Corliss et al., 2009; Gooday, 2002). The dominance of G. subglobosa, the relatively high total abundance of benthic foraminifera, and low H′ and J′ values between 6300 and 5200 cal yr BP (Figure 4; Supplemental Figure S1) suggest intense phytodetritus input scenarios in this phase, similar to what was reported at Cape Frio (23°S) (Dias et al., 2018). This time interval also presented a relatively higher abundance of U. peregrina. Previous studies in the southwestern Atlantic margin indicated that U. peregrina can assimilate more refractory organic matter buried deeper in the sedimentary column (Burone et al., 2011) and would be related to the enrichment of nitrate caused by upwelling events (presence of SACW) in coastal areas near Cape Santa Marta (de Freitas Toniolo et al., 2020; Eichler et al., 2008).
The onset for the last 6300 cal yr BP of the PPW, transporting colder and nutrient-rich waters, as well as terrestrial organic carbon to the southern Brazilian continental margin, likely caused the relative increase in productivity from 5200 to 2800 cal yr BP. This is indicated by the higher infaunal foraminifera abundance, TOC content, BFHP index values (Figure 3), and relatively higher abundances of Uvigerina spp. (Figure 4). In addition, a higher relative abundance of B. marginata between 5200 and 4000 cal yr BP is also observed. These species are considered to be related to a greater supply of organic carbon derived from oceanic productivity (Donnici and Barbero, 2002; Martins et al., 2006, 2007). At the same time, the abundance of G. subglobosa and U. peregrina decreased. Dias et al. (2018) proposed that B. marginata and G. subglobosa compete for phytodetritus on the Cabo Frio shelf (~23°S) during periods of high food availability, leading to a lower abundance of B. marginata, which suggests that this species may exhibit greater resilience in periods of lower flux and/or changes in food quality than most other species. This time interval also had relatively weaker bottom water currents and oxygenation (e.g. low BFOI values and low G. subglobosa abundance), followed by an increase in H′ and J′ values. The weaker bottom currents enabled the deposition of finer sediments, causing a relatively more constant export of organic matter to the seafloor, which favored an increase in the diversity of benthic foraminifera.
The highest marine productivity occurred between 2800 and 890 cal yr BP, marked by the highest values of the BFHP index and infaunal species (~80%) (Figure 3). The benthic foraminiferal assemblages were similar to those of the previous period. However, an increase in the abundance of B. elegantissima stands out. B. elegantissima is generally associated with high organic matter in coastal environments and less intense hydrodynamic conditions (Bonetti, 2000; Burone, 2002; Sen Gupta et al., 1996).
This phase is marked by a slightly decrease of the H′ and J′ indices and the low total abundance of benthic foraminifera. These results may seem contradictory to the hypothesis that this interval shows relatively higher productivity. However, with the increase in productivity, other environmental parameters can be changed (e.g. oxygen concentration and degradation of organic matter). Therefore, many specimens from the foraminifera community would not be able to adapt to this new environment, as well as the competition, thus occurring both a decrease in absolute density, diversity, and equitability. These conditions are well evidenced in the dominance of some species that manage to survive in this environment, such as B. elegantissima. Martins et al. (2007) observed a similar fact on the Iberian continental shelf, where a strong increase in organic carbon flux caused a sharp decrease in oxygen concentrations. This period also had similar BFOI values and G. subglobosa abundance compared to previous phase values, as well as an increasing pattern of sedimentation rates and fine sediments (~95%). These higher productivity conditions are in agreement with the greater influence of the nutrient-rich PPW on the inner shelf off southern Brazil.
Sea-surface temperature
The period between 6300 and 5200 cal yr BP was the only phase in which SST-UK′37 presented an increasing trend. Despite the correlation between SST variations and the strengthening of the BC on the BC front (Chiessi et al., 2014) and considering our coring site in an inner shelf region (60 m isobath), is more likely that the higher SST-UK′37 values are related to the fact that no intrusion of the colder PPW occurred in this period.
Cordeiro et al. (2014) and Ceccopieri et al. (2018) reported that inshore and central regions (<120 m) of the Cape Frio shelf (23°S) reflect mean winter (July–September) alkenone based SST-UK′37, which also corresponds to the period of the maximum abundance of haptophytes found in the region (Rodrigues et al., 2014). In contrast, the upwelling of the SACW at Cape Santa Marta occurs during spring and summer due to the action of northeasterly winds (Campos et al., 2013). These divergent seasonal processes may explain the increasing SST-UK′37 values with the presence of brief upwelling events registered by benthic foraminifera species.
The decreasing trend in SST-UK′37 starting at 5200 cal yr BP also suggests the influence of colder water masses related to the northward transport of the PPW by the BCC in this period (Figure 6j). The maximum northwards displacement of the PPW occurs in the winter, influenced by the dominance of SW winds (Depetris and Pasquini, 2007; Piola et al., 2008). Therefore, as the alkenones are produced by species of haptophyte algae (e.g. Emiliania huxleyi and Gephyrocapsa oceanica) and these species have their maximum abundance in the winter in the southern Brazilian continental shelf (Rodrigues et al., 2014), the SST-UK′37 probably reflects a winter seasonal signal in the core site region, and are mainly influenced by the PPW after 5200 cal yr BP.
After 2800 cal yr BP, the SST-UK′37 values followed the decreasing pattern of the previous period, reaching the lowest values. The SST-UK′37 results of the core 7606 for the last 6300 cal yr BP are directly influenced by the PPW, which may explain the disagreement with other paleotemperature studies in the southwestern Atlantic margin (Nagai et al., 2020). In addition, recent studies also reported the influence of the PPW on paleotemperature variations in the inner shelf off southern Brazil (de Freitas Toniolo et al., 2020).
Conclusions
The combined approach focusing on lipid biomarkers and benthic foraminifera proxies in a marine sedimentary record revealed three distinct phases along the mid- to late-Holocene on the southwestern Atlantic shelf. In addition, this work also produced novel information about the relationship between the LPR terrigenous input and marine productivity. The variations were affected mainly by sea level, hydroclimate changes in paleoproductivity, and the input of terrigenous material.
Between 6300 and 5200 cal yr BP, the higher sea level, northward displacement of the ITCZ, and weaker ENSO amplitude resulted in a relatively low rainfall regime in the Paraná River drainage basin and the trapping of sediments in the LPR estuary. These events influenced the inner shelf off Cape Santa Marta, reducing the sedimentation rate and input of terrigenous material, which consisted of a mixture of sources from the adjacent continent and southern regions, possibly from the Itajaí-Açu River and the Argentinian shelf, respectively. Marine productivity in this period was also lower, with a probable influence of phytodetritus input pulses. The period between 5200 and 2800 cal yr BP represented a transition phase, with a relative increase in terrigenous input and marine productivity, a decrease in SST, and a shift in the regional terrigenous plant source. The relatively lower sea level, the long-term intensification of the SAMS, the intensification of ENSO events, and the increase in the frequency of cold fronts, and, consequently, S/SE winds indicate the onset for the last 6300 cal yr BP of the influence of the PPW on the southern Brazilian shelf. Greater terrigenous material input from southern regions and, consequently, greater marine productivity were observed after 2800 cal yr BP. This points to the establishment of the modern conditions of wind and river discharge, which presents the LPR as the dominant source of sediment on the inner shelf off Cape Santa Marta and its displacement by the BCC the main controlling feature of the seasonal variation in marine production.
Supplemental Material
sj-docx-2-hol-10.1177_09596836211041727 – Supplemental material for Mid- to late-Holocene analysis of the influence of the La Plata River plume on the southwestern Atlantic shelf: A paleoenvironmental reconstruction based on lipid biomarkers and benthic foraminifera
Supplemental material, sj-docx-2-hol-10.1177_09596836211041727 for Mid- to late-Holocene analysis of the influence of the La Plata River plume on the southwestern Atlantic shelf: A paleoenvironmental reconstruction based on lipid biomarkers and benthic foraminifera by Marcia Caruso Bícego, Felipe Rodrigues Santos, Poliana Carvalho de Aandrade Furlan, Rafael André Lourenço, Satie Taniguchi, Silvia Helena de Mello e Sousa, Renata Hanae Nagai, Ana Beatriz Leite Cavalcante, Rubens Cesar Lopes Figueira, Ilana Klein Coaracy Wainer and Michel Michaelovitch de Mahiques in The Holocene
Research Data
sj-xlsx-1-hol-10.1177_09596836211041727 – for Mid- to late-Holocene analysis of the influence of the La Plata River plume on the southwestern Atlantic shelf: A paleoenvironmental reconstruction based on lipid biomarkers and benthic foraminifera
sj-xlsx-1-hol-10.1177_09596836211041727 for Mid- to late-Holocene analysis of the influence of the La Plata River plume on the southwestern Atlantic shelf: A paleoenvironmental reconstruction based on lipid biomarkers and benthic foraminifera by Marcia Caruso Bícego, Felipe Rodrigues Santos, Poliana Carvalho de Aandrade Furlan, Rafael André Lourenço, Satie Taniguchi, Silvia Helena de Mello e Sousa, Renata Hanae Nagai, Ana Beatriz Leite Cavalcante, Rubens Cesar Lopes Figueira, Ilana Klein Coaracy Wainer and Michel Michaelovitch de Mahiques in The Holocene
Footnotes
Acknowledgements
The authors wish to thank the crew of the R.V. “Prof. W. Besnard”. The authors would also like to acknowledge the contributions of Dr. Fabienne Marret and Dr. Alix Cage who provided helpful comments on the manuscript.
Data availability statement
The data presented in this article will be available on the Pangaea repository.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Financial support was provided by the Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP) through grant 2003/10740-0, 2017/17763-2 and 2019/15252-1.
Supplemental material
Supplemental material for this article is available online
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.