
Abstract
Alkaline fens are important Natura 2000 habitats, which harbor many endangered plant species. Alkaline fens are formed in areas with groundwater discharge and usually developed in a natural way in the early Holocene. We radiocarbon dated the base of three peat deposits from spring fens along the flanks of the ice-pushed ridge near the village of Ootmarsum to find out when and why peat-forming vegetation started to grow. We cored a sequence in the Mosbeek Valley for detailed paleoecological analyses of micro- and macrofossils. To our surprise, we found strong evidence for human impact during the 13th and 14th centuries AD as the triggering factor for starting organic colluvial accumulation and peat growth at sites where natural springs are present. This shows that this fen is not a relic, but results from changes in land use. Human actions were: (1) deforestation causing increased run-off and reduced evaporation on the plateaus by the vegetation, resulting in increased seepage in the valleys, (2) intensification of agriculture, trade routes, and paired erosion, which formed colluvial deposits and sediment fans that hampered fast run-off water, (3) increased back and groundwater levels after the foundation of watermills; four lowering water levels due to intensification and reorganization of water use by new watermills, and (4) head cut erosion and spring erosion after privatization and cultivation of common pastures after the mid 19th century. This means that cultural-historical changes in the landscape were much more important for alkaline fens than expected.
Keywords
Introduction
Alkaline fens (EIONET, 2015; Provincie Overijssel, 2017) are important habitats for biodiversity. They are characterized by small sedge and brown moss communities, and rich in rare plant species (Šefferova Stanova et al., 2008). Alkaline fens belong to the most threatened habitats in Europe and have become very rare in most of the EU countries (Grootjans et al., 2021). They are protected by the European Habitat directive (European Union, 1992). In the Netherlands, alkaline fens are very rare, and they belong to the most diverse habitat types in the country (Joosten et al., 2017).
Alkaline fens develop on permanently waterlogged soils with calcareous water and small water level fluctuations, often in areas with groundwater discharge (Grootjans et al., 2021; Šefferova Stanova et al., 2008). The development of alkaline fens is basically a natural process, and many fens probably started in the early Holocene (Dobrowolski et al., 2019; Grootjans et al., 2015; Kossler and Strahl, 2011; van der Hammen and Wijmstra, 1971; Van Geel et al., 1980). Human influence on the origin of fens and peat growth is usually low, although some authors (Czerwiński et al., 2021; Lamentowicz et al., 2013; Moore, 1987) concluded that some mires can be regarded as man-made habitats. Anthropogenic deforestation in the early Middle Ages locally led to increased groundwater discharge and often resulted in the development of alkaline fens (Grootjans et al., 2021), sometimes even in a former Sphagnum fuscum bog (Hájková et al., 2012).
Paleoecological studies can contribute to the reconstruction of human impact on past environments (e.g. Behre, 1981). The study of pollen and spores allows reconstructions of trends in the changing intensity of human impact on the landscape, while macrofossils like seeds and mosses mainly show the local vegetation development of the sampling site. In the present study we performed a paleoecological reconstruction of the Mosbeek fen sediment and peat. The Mosbeek fen is one of the few remaining rather well developed alkaline fens in The Netherlands (Ootmarsum region; Figure 1: Provincie Overijssel, 2017). The main research question of our study is: when and why did Holocene organic accumulation and peat growth near springs start along the flanks of the ice-pushed ridge of Ootmarsum?
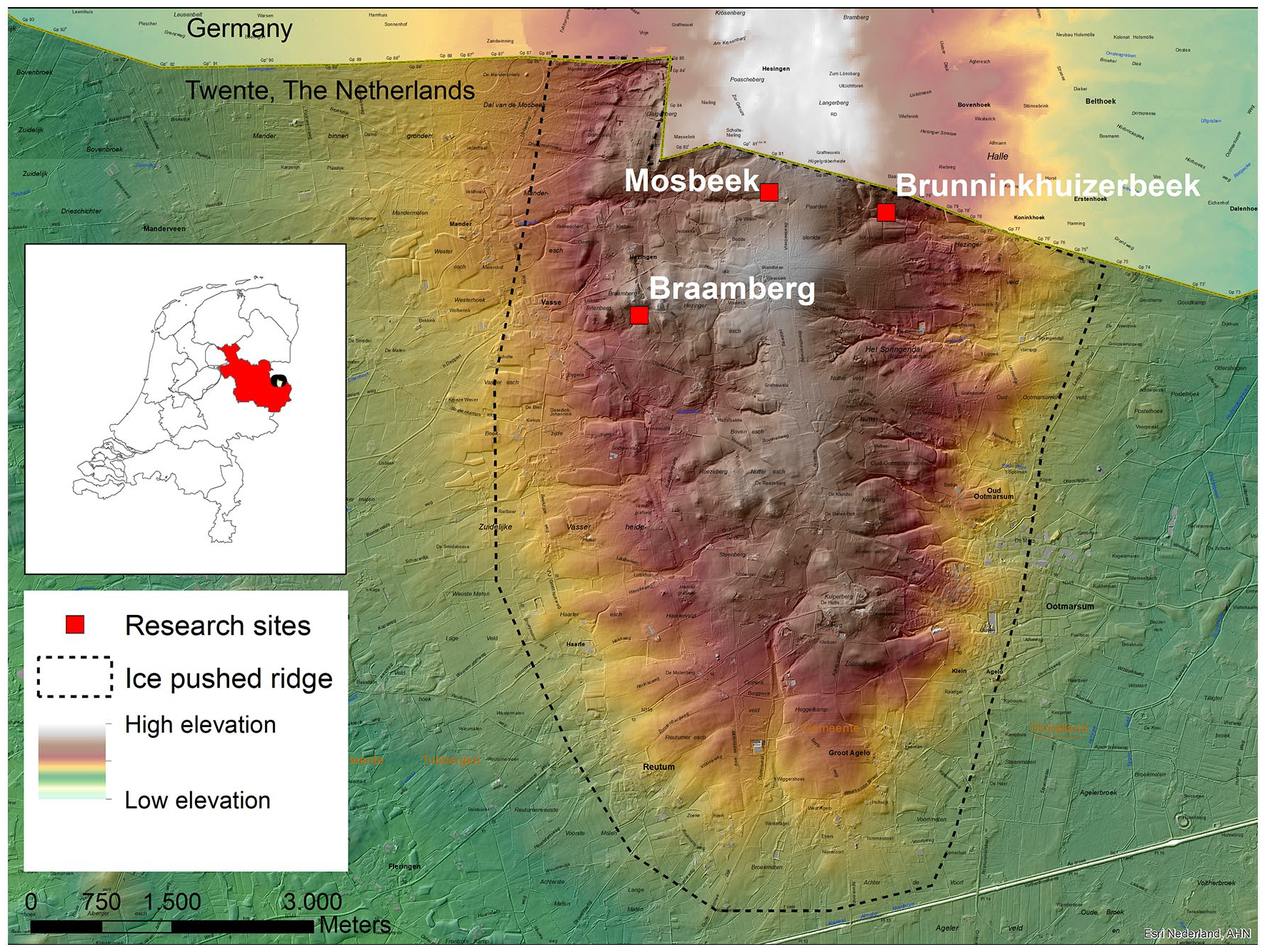
Global boundary of the ice pushed ridge in the Ootmarsum region and the research sites. Map source: Esri Nederland, Jan Willem van Aalst.
Study area
Some aspects of geology and geomorphology of the Ootmarsum region
The Mosbeek study area is situated on the ice-pushed ridge of Ootmarsum in the Twente region, between the villages of Ootmarsum and Vasse (Figure 1).
The landscape of the ridge shows relatively small-scale forests, heathland, meadows, and arable land, which were difficult to reclaim and cultivate in the past.
The stratigraphy of the ice-pushed ridge shows skewed, folded, and pushed up Tertiary and early Pleistocene deposits.
The oldest sediments outcropping at the surface are clays of Tertiary age (48–3.6 million years ago). Pushed-up fluvial deposits date from the Pliocene-Pleistocene transition period and come from the present area of the Elbe and Weser rivers. The Tertiary and Pleistocene deposits were ice-pushed during the Saalian glaciation and were forested during the Eemian. During the Weichselian glacial period the ridge became partly eroded by wind, water and solifluction, and various valleys were formed. Weichselian coversands and Holocene colluvia in the study area are generally thinner than 2 m (Beerten, 2014). The permeability of the skewed Tertiary clay layers is low, while the permeability of Pliocene and Pleistocene sands is high, which leads to the presence of local springs in the ice-pushed ridge (Figure 1). Groundwater is discharged from the springs permanently or periodically, and one of the most important spring areas, the source of the Mosbeek Valley, even developed into an alkaline fen, which is protected by the European Habitat Directive (European Union, 1992). In this spring area, peat was formed, which we used as a paleoecological archive for a reconstruction of the local and regional vegetation history.
Until recently, different factors for peat growth in the ice-pushed ridge area have been mentioned, but historical or paleoecological data to test hypotheses about the chronology and environmental triggers for peat growth were missing. Archeological data show that colluvial deposits were formed from the Neolithic to the Late Medieval Period (Groenewoudt, 1994; Scholte Lubberink, 1998; van Beek, 2009; van der Velde, 2011). The formation of colluvial deposits in stream valleys was enhanced in the Late Medieval Period by intensified arable land use and artificial water table fluctuations, necessary to get energy for several watermills (Smeenge, 2020). We focus on the chronology and vegetation development of starting peat growth near the spring area of the Mosbeek Valley, and the possible relation with human impact.
At present, the vegetation in the Mosbeek fen area consists of a gradient of Natura 2000 habitat types, from alkaline fen in the actual springs and brooks, to Molinia meadows on base-rich soils, species-rich Nardus grasslands and wet heaths with Erica tetralix in the surroundings (Provincie Overijssel, 2017). The spring fen is classified as alkaline fen in the Natura 2000 habitat system, but calcium levels of the groundwater are relatively low (Kooijman et al., 1994). In the upper 2 m of the spring soil, calcium values ranged between 29 and 35 mg/l, which makes the fen sensitive to local acidification.
In 2016, the herb layer within the spring area was dominated by Rhynchospora alba, Carex panicea, and Juncus acutiflorus (Table 1). Plant species such as Pinguicula vulgaris, Parnassia palustris, Molinia caerulea, and Erica tetralix were also present. The moss layer was dominated by Scorpidium scorpioides and Bryum pseudotriquetrum. Sphagnum species such as S. magellanicum are absent from the actual spring area today but were present in the 1940s (Kooijman et al., 1994). The same holds for Scorpidium revolvens , which was also known from the 1940s, but disappeared in later years.
Vegetation relevé sampling location 2016. Nomenclature is according to van der Meijden (2005) for vascular plants and van Tooren et al. (2007) for bryophytes. Cover values are estimated according to Braun-Blanquet (1932).

Information on human impact such as the year of building of watermills, the geographical location, and effects like erosion and sedimentation because of hydrological interventions was obtained from historical sources, such as archives about ownership, old maps, cadastral data, micro-toponyms, and oral history (Smeenge, 2020). Today there are two watermills in the area: Bels watermill and Frans watermill, but according to Slicher van Bath (1944), Hijszeler (1970), and Hagens (1979) there were two other watermills along the Mosbeek Valley in the past. Smeenge (2020) reconstructed the former locations of these additional watermills based on the stream gradient (Figure 2), cartographical data, old field names, and oral history.
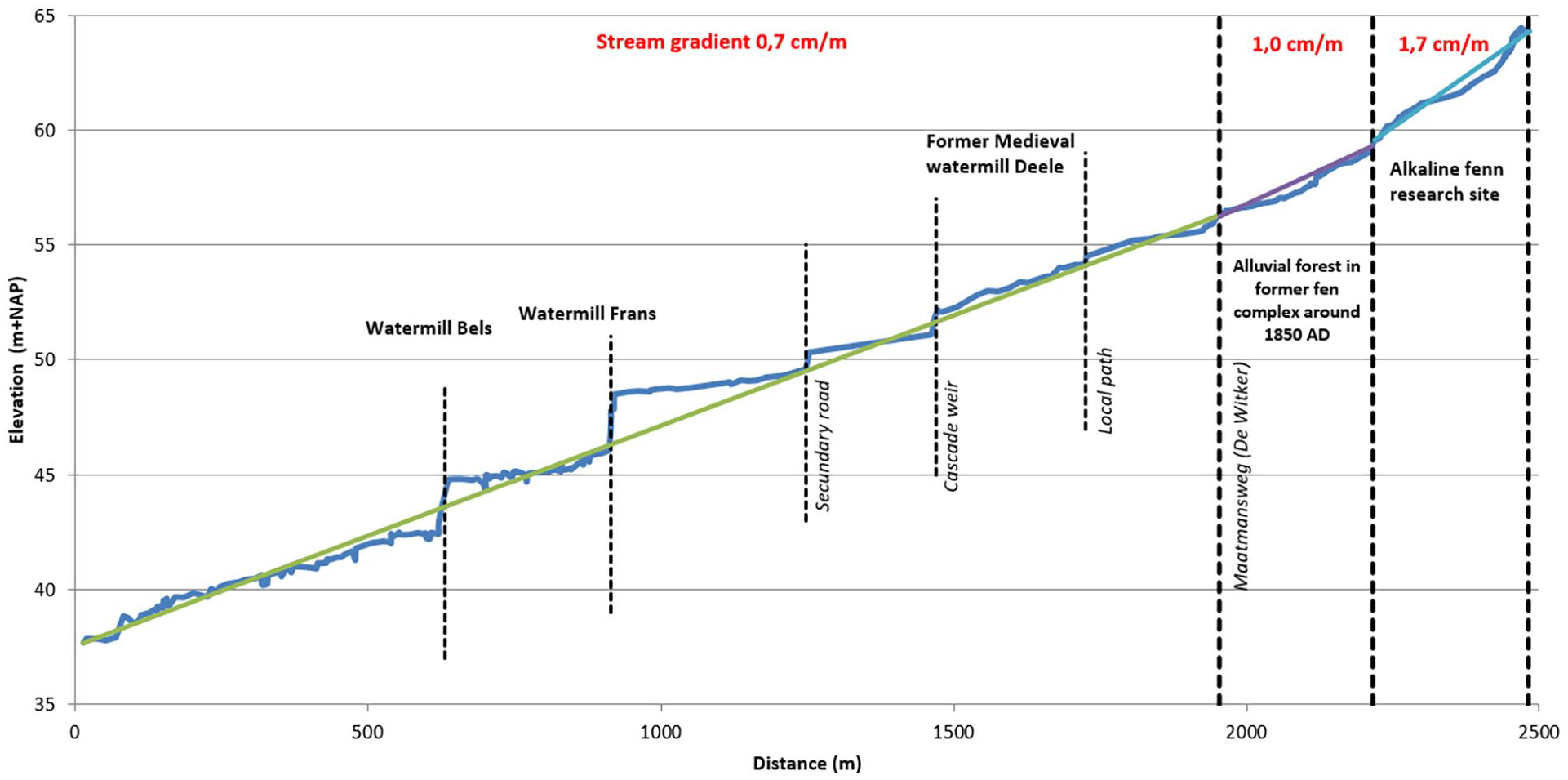
Stream gradient of the rivulet in the Mosbeek Valley. The research site is the spring area of the stream. Cascades are formed by obstructions in the stream bed.
To obtain paleohydrological information, the bottom level of the rivulet in the Mosbeek Valley was measured at 547 points, from the Mosbeek spring area until 2500 m downstream. The stream gradient shows zones of erosion (relatively steep intervals) and zones of sedimentation (relatively flat intervals) related to the presence of watermills (Figure 2). The profile also shows that the oldest watermill along the Mosbeek, the Deele watermill, probably built during the 13th–14th century AD, was located much closer to the Mosbeek spring area than the present ones (Hagens, 1979).
Material and methods
Sampling, radiocarbon dating, and age modeling
Samples for radiocarbon dating were taken at the base of organic accumulation for the Braamberg, Brunninkhuizerbeek, and Mosbeek sites (Figure 1). Seeds and leaf fragments were selected for radiocarbon dating. Radiocarbon dates (Tables 2 and 3) were calibrated using OXCAL, version 4.2.4 (Bronk Ramsey, 1995). Chronologies were modeled using a Bayesian approach using the rbacon package (version 2.5.7) in R (Blaauw and Christen, 2011). In addition, the sampling year AD 2016 was assigned to the surface of the core and the forest plantations around AD 1858 to the Pine pollen rise. After calibration, the modeling procedure of Bacon takes account of the entire probability distribution of each dated level while creating robust chronologies including estimations of age uncertainties.
Results C-14 dates of the bases of three cores.

Results C-14 dating of the Mosbeek sequence.

Based on the presence of peat-forming alkaline fen vegetation at the surface, and considering the thickness of the organic deposit, our Mosbeek site was selected for a detailed investigation. We used a Wardenaar profile sampler (Wardenaar, 1987) and obtained a 76 cm long sequence. The upper 71 cm of the core consisted of peat. The upper 25 cm was partly mineralized. Below 71 cm the sediment was colluvial sandy material. Obtaining sediment below 76 cm was not possible because of flowing wet sand. The material was transported to the laboratory, where 16 sub-samples were taken for the study of microfossils and, from the same depths, 16 samples for macrofossil analysis.
Three samples (at 76, 36, 26 cm depth) were taken for radiocarbon dating of the Mosbeek sequence, based on the stratigraphical information. Microfossil preparation was according to Faegri and Iversen (1989) and macrofossil samples were prepared following Mauquoy and van Geel (2007). Figures 3 and 4 respectively show the microfossil diagram and the macrofossil diagram. The diagrams were constructed using TILIA (Grimm, 1991) and TGView. Calculation of microfossil percentages were based on a pollen sum including pollen of trees, shrubs, and upland herbs. Percentages of taxa not included in the sum (aquatic taxa, ferns, mosses, fungi, and other non-pollen palynomorphs to the right of the pollen sum column) were also expressed on that pollen sum. The number of pollen grains per sample was between 350 and 400. In the pollen diagrams, the exaggeration factor is ×5.

Mosbeek microfossil diagram.

Mosbeek macrofossil diagram.
Pollen diagrams from lake sediments and peat deposits generally reflect a regional picture of the vegetation, while macrofossil diagrams mainly show remains of plants that were living nearby or strictly local at the place where the sequence was cored. We should consider that the extent of Mosbeek fen was once only 4 ha, whereas nowadays the remaining part is even smaller (ca 1 ha). It is situated in the uppermost part of the valley and therefore – apart from plant remains of strictly local origin – some macrofossils from plants growing at the higher and drier soils above the valley mire are also to be expected. Especially the lower half of the sampled core consisted partly of wind-blown and water-transported sand particles, probably deriving from eroding cultivated soils in the nearby upstream area. This “mix factor” may complicate the interpretation, especially of rare taxa.
Results
Radiocarbon dating
The sampling sites are valley bottoms with a colluvial base, mainly followed by Carex peat. Remarkably, the start of organic accumulation of the sampled cores (Braamberg, Brunninkhuizerbeek, and Mosbeek; Table 2) occur since the Late Medieval Period (between ca 1270 and 1400 AD). The Mosbeek fen started during the 13th/14th century AD, while local peat growth dominated from ca 1500 AD on.
Three calibrated radiocarbon dates from the Mosbeek sequence (Table 3) were used for the Bayesian model approach as shown in Figure 5. Considerable irregularities in the probability distribution of the calibrated radiocarbon dates at 26 and 36 cm depth are caused by strong fluctuations in the atmospheric radiocarbon content during the Little Ice Age (Mauquoy et al., 2002; Reimer et al., 2020). The estimated date for the historical known start of pine plantations (privatization common grounds around 1858 AD; Demoed, 1987) is shown in Figure 5 at 20 cm depth and the sampling year (surface of the core: AD 2016) have also been considered. It is evident that the start of the Mosbeek sequence was in the Late Medieval Period.
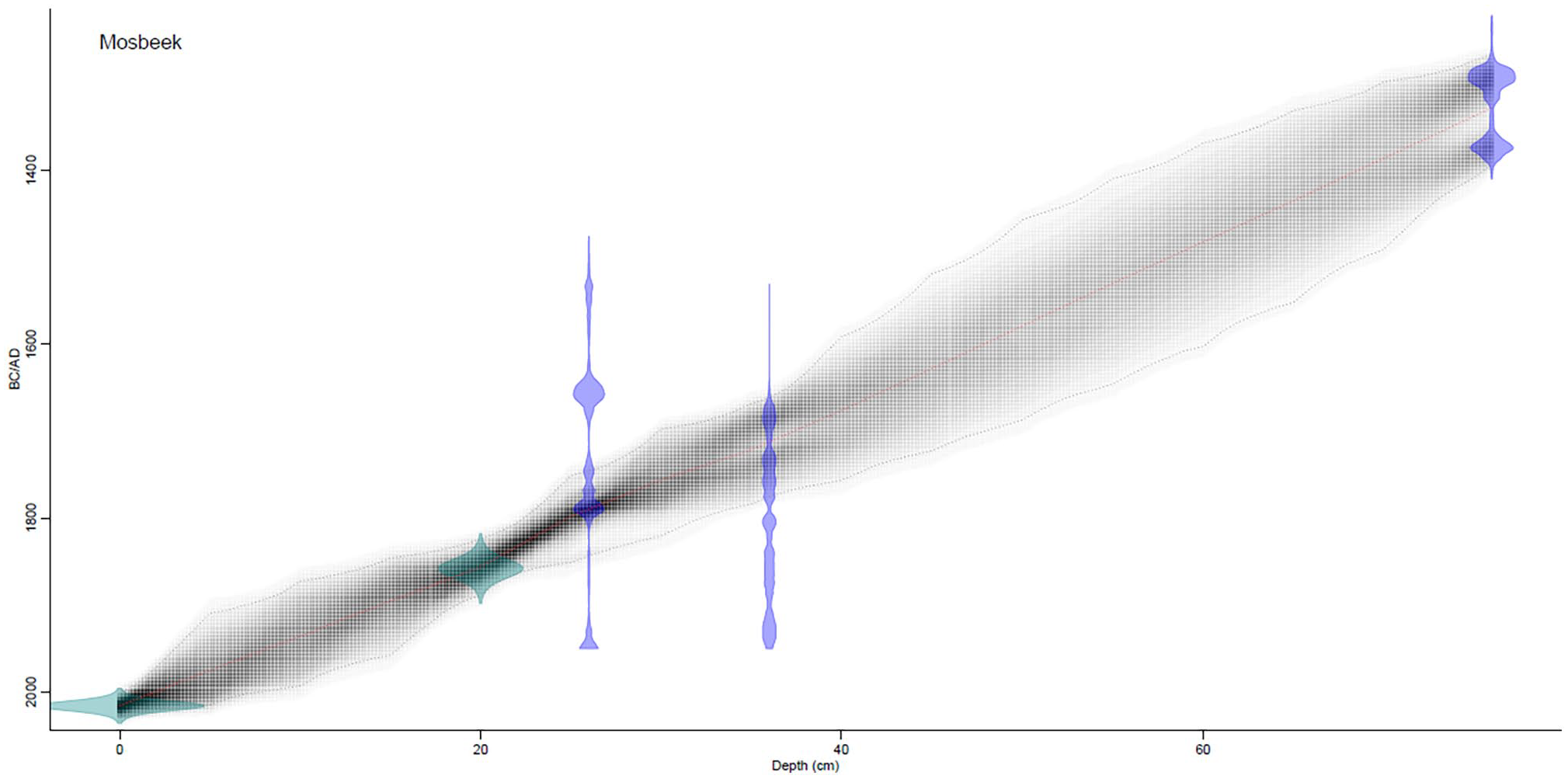
Age-depth relation based on the Bacon model results (Blaauw and Christen, 2011). The confidence intervals on the graph are at 95% results.
Regional vegetation history as based on the Mosbeek core
The Mosbeek sequence encompasses the last ca 700 years, regarding the radiocarbon date at the base of the core. For the description of the regional vegetation development we distinguished two pollen assemblage zones (A and B), with a zone boundary at 22 cm depth (Figure 3).
Zone A (76–22 cm; ca 1330–1860 AD)
The presence of Fagopyrum pollen already in the lower part of zone A further supports a Late Medieval (or later) start of the base of the Mosbeek sequence (but see de Klerk et al., 2015 for exceptional earlier dates of Fagopyrum). In zone A, sand was an important sediment component, which points to a dynamic landscape with upland erosion and sedimentation of eroded sand in the valley. Ascospores of the Sordaria-type point to the nearby deposition of herbivore feces (van Geel and Aptroot, 2006).
Alnus dominates the pollen record in the lower 45 cm of the core, but alder may be over-represented. The presence of Lonicera pollen in all samples of the lower 45 cm of the core probably indicates that an alder carr was present in the valley at a short distance from the sampling site. These pollen grains are relatively big and produced in low quantities as it is an insect pollinated species. The lower percentage of Alnus pollen at 36 cm may represent temporary forest cutting and fast regrowth of the alder carr. Other tree species in the region were Betula, Corylus, and Quercus. Dominant herbaceous taxa, representing a cultural landscape, were Poaceae and Ericales. Other human impact indicators were Asteraceae, Centaurea cyanus, Fagopyrum, Persicaria maculosa, Plantago species, Rumex acetosella, and spores of Anthoceros punctatus and Phaeoceros laevis (Koelbloed and Kroeze, 1965). Secale cereale and other cerealia show an increase in the upper part of zone A.
Zone B (22–0 cm; ca 1860–2016 AD)
The peat from 22 cm depth to the top of the core represents the last ca 160 years.
Alnus shows a steep decline, pointing to felling of alder in the valley areas, while the rise of Pinus and Picea reflects the historically known regional plantations. The curve of Sordaria-type ascospores points to an increased population density of herbivores in the surroundings of the fen and a slight increase of Urtica indicates nitrogen-rich sites in the landscape.
Local vegetation succession of the Mosbeek core
We distinguished four zones for the local vegetation succession. Estimated quantities of sand volumes (vs organic material in the macrofossil samples) were also indicated in the macrofossil diagram.
The description and interpretation of the local vegetation succession is mainly based on the macrofossil record (Figure 4) and to a minor extent upon the microfossils (Figure 3).
Zone 1 (76–53 cm; ca 1330–1550 AD)
The sand in the sediment of zone 1 points to a dynamic landscape with upland erosion and sedimentation of eroded sand in the valley by wind and water transport. Large numbers of Juncus seeds may have arrived with the sand.
We found species pointing to a local spring, where permanently running water was present. The invertebrates present (Ostracods, Plumatella, Sialis, Simocephalus, and Trichoptera) require an aquatic environment. Potamogeton coloratus nowadays is one of the rarest Potamogeton species in the Netherlands. It needs relatively cool water with a high bicarbonate level and low amounts of ammonium and phosphate (Weeda et al., 1991). A base-rich environment is also reflected by Sagina nodosa and Montia fontana, together with Selaginella selaginoides, which is now extinct in The Netherlands and the brown mosses Scorpidium scorpiodes and Bryum pseudotriquetrum (Schaminée et al., 1995). Micro-environments were complex because there were also acidifying mini habitats with Sphagnum species (species of the section Cuspidata, S. squarrosum, S palustre, and S. papillosum, the latter pointing to local hummocks). Vascular plants included Molinia caerulea, Erica tetralix, and Calluna vulgaris. Nearby the sampling site Persicaria mitis, and various Juncus species played a role, especially in transitional zones toward the relatively high and dry mineral soils outside the valley.
Zone 2 (53–28 cm; ca 1550–1730 AD)
The taxa that indicate open water in the interval of zone 1 disappeared and from ca 1550 AD onward, the accumulation of organic material, and thus the formation of peat started. The relative contribution of sand in the sediment declined and lower numbers of Juncus seeds were recorded. The increase of Scorpidium scorpioides points to a continued availability of base-rich groundwater and most of the base-rich spring fen taxa of zone 1 continue their presence. Scorpidium revolvens is also a species of base-rich conditions, although it may indicate a transition toward more acidic conditions. Sphagnum magellanicum and Viola palustris point to the co-existence of hummocks dependent on rainwater. The presence of Glyceria maxima, Lychnis flos-cuculi, Carex disticha, and Stellaria graminea indicate local eutrophication, perhaps due to grazing.
Zone 3 (28–16 cm; ca 1730–1890 AD)
The spring fen taxa have disappeared, although rich-fen species such as Scorpidium revolvens, Carex oederi, C. flacca, and Sagina nodosa are still present in the lower part of this section. However, the peat mainly consists of Sphagnum section Cuspidata and Acutifolia, which indicate increasing acidification. This is supported by the presence of Molinia caerulea, Rhynchospora alba, Polytrichum commune, and Erica tetralix.
Zone 4 (16–0 cm; ca 1890–2016 AD)
In this zone, base-rich species reappeared. Carex flacca and C. otrubae indicate a continuation of the availability of base-rich water, together with Riccardia chamedryfolia and Plagiomnium affine. Sphagnum tenellum is a species of mesotrophic to oligotrophic conditions. Grazing and mineralization of the upper peat layer caused eutrophication, as witnessed by the presence of Lycopus europaeus and Cirsium palustre.
Discussion and conclusions
The results show that the Mosbeek fen started during the Late Medieval Period, as a result of human impact. During that period the land holdings of the Bishop of Utrecht, local nobility, and monasteries expanded. As a result, upland areas were deforested and converted into fields, while alder carrs in stream valleys were deforested to create meadows (Noomen, 1990; Spek, 2004, 2014; Spek et al., 2010; Woolderink, 2014; Smeenge, 2020).
Deforestation, an increase of arable land and intensive traffic of a Hanseatic trade route on a bordering cart track (see right side in Figure 6) triggered erosion followed by colluvial deposits in valley bottoms (Figure 7; Bruns and Weczerka, 1967; Smeenge, 2020). The natural runoff in the valleys was slowed by sediment fans perpendicular to the valley (De Vries, 2016; Scholte Lubberink, 2008; Smeenge, 2020). But when forest was converted into short vegetation, the net precipitation surplus also increased due to reduced evaporation. So increased sedimentation of eroded sand, in combination with reduced evaporation in the surrounding landscape led to a rise in the groundwater levels in the valley (Dolman and Moors, 1994; Hájková et al., 2012; Hoogendoorn, 1992; Spek, 2004; Straelens and Mohren, 2010).
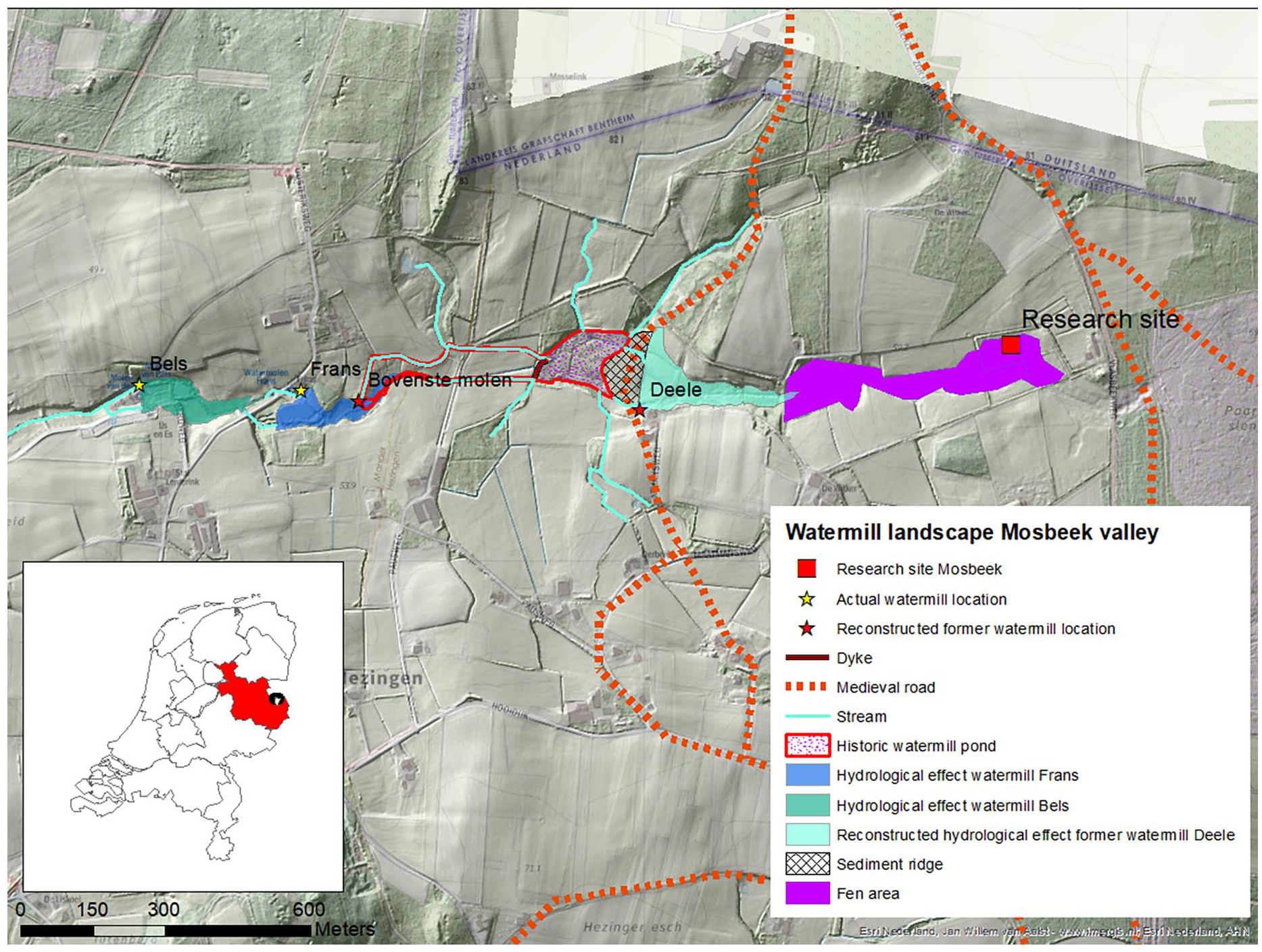
Reconstructed effects of watermills in the valley. The Deele Mill was the first one that was replaced in the 17th century by the Bovenste molen followed by the Bels and Frans watermills. The last two are the only remaining watermills in the valley.
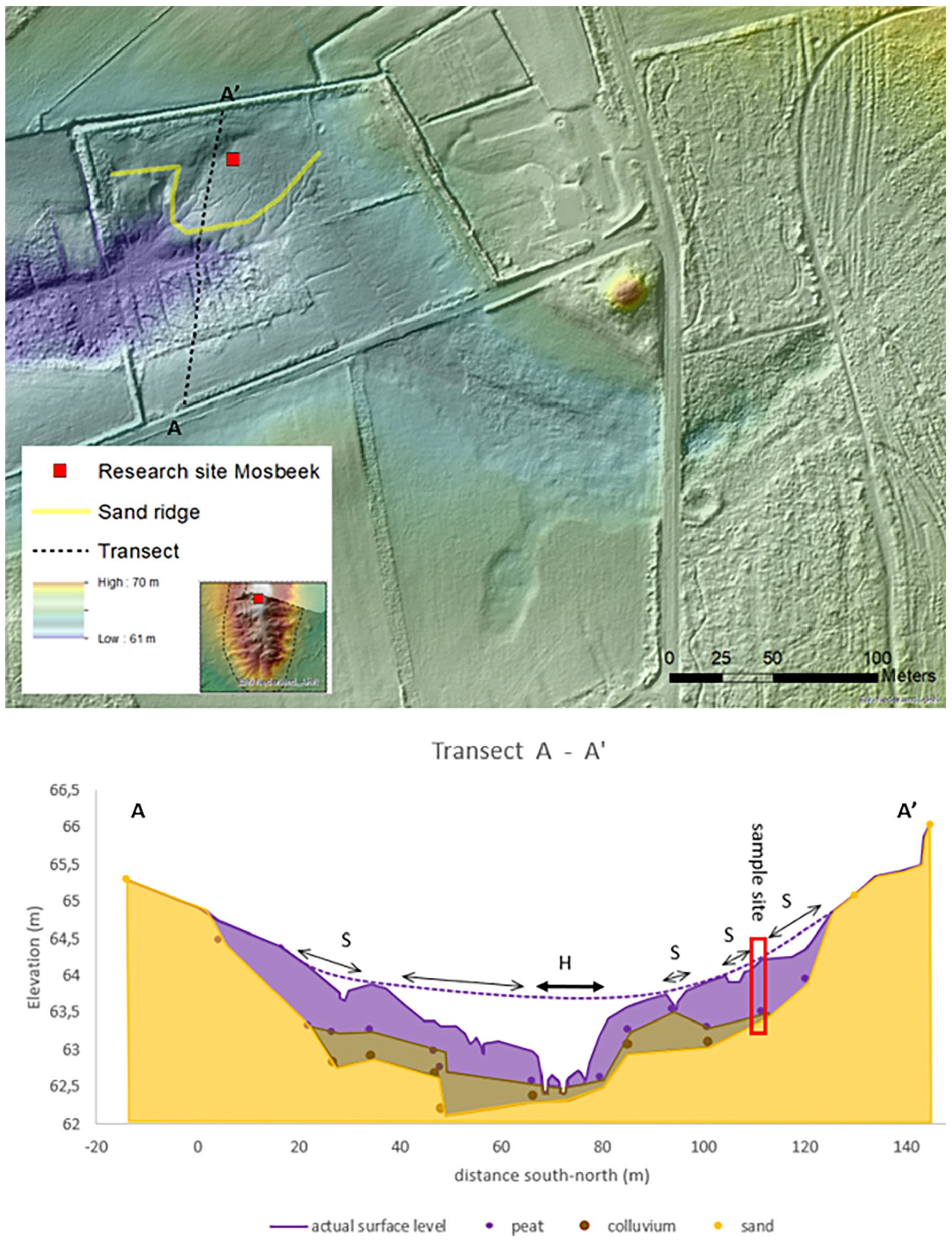
Transect of the “flow through mire” with the stratification and main drainage that led to head cut erosion (H). Several smaller gullies and former ditches led to spring erosion (S). Based on the height of the surface level of the fen area we assumed that much of the peat in the mire disappeared due to erosion forms (see dotted line at the surface level).
A more continuous flow of surface water through the valleys stimulated the Bishop of Utrecht to organize the building of watermills. Those mills were built on sediment fans to make the back-up of water as easy as possible. The accumulation of water in reservoirs increased waterlogging and a rise of groundwater levels in the valleys (Smeenge, 2020). The reservoir upstream of the Deele Mill probably served till 1676 AD as a “cork in the bottle” for the Mosbeek fen and reduced its drainage (Figure 7). It is not clear when exactly the mill was built in the 13th–14th century, but together with deforestation and higher groundwater levels, it likely played an important role in the start of the peat formation (Figure 6).
The paleoecological analysis of our Mosbeek sampling site points to an open vegetation from the start of the recorded sequence. The local environment during the first few centuries was aquatic, and carbonate rich. Water levels further increased with the construction of the Medieval watermills. The deposition of the sandy sediments in the valley was also linked to deforestation and increase of arable land with intensification and extension of agriculture. Also, the Mosbeek springs were close to a major trade route (Smeenge, 2020), and probably used by drinking animals, which trampled the soil. In the Mosbeek sequence we observe after ca 1770 AD a lowered input of sand. The relatively steep height gradient and a high groundwater pressure caused a development of a “flow through mire” (Joosten et al., 2017). Because the buffer capacity of the spring water is relatively low, the macro-remains of the mire represent both hollows, characterized by species that need base-rich conditions (Sagina nodosa; Scorpidium scorpiodes; Bryum pseudotriquetrum), and hummocks with species preferring more acid ( Rhynchospora , Viola palustris; Erica tetralix) conditions (e.g. Kooijman and Hedenäs, 1991; Weeda et al., 1988). Low buffer capacity of the spring fen is supported by the presence of Scorpidium revolvens, which generally grows in intermediate fen types (Kooijman and Hedenäs, 1991). This species has become extremely rare in the Netherlands, and the Mosbeek fen was one of the last places where it was found (Kooijman, 1992). The present complex Mosbeek vegetation has also become rare in the Netherlands as well as in other European countries (Joosten et al., 2017).
A major change in the local vegetation succession at 28 cm depth toward acid conditions (transition zone 2 to zone 3) can be explained as the effect of declining contact of the vegetation with mineral-rich groundwater and increased influence of nutrient-poor rainwater. Based on historical records the Deele Mill was replaced by several new downstream watermills between 1676 to 1711 AD (Figure 6). This probably caused a drop in the groundwater level and appearance of rainwater indicator species in the spring area. The minimum age of the radiocarbon date at 28 cm depth reflects best with these historical ecological events (ca. 1730 AD).
Based on the ecohydrological conditions, acidification due to stagnation, which is common in peat forming ecosystems, did not occur (Joosten et al., 2017). A major change was caused by “flow through mire” the privatization of the common fields around 1858 AD. Different farmers received a piece of the mire and dug out several ditches to improve the pasture quality (Figure 7). However, the central part of the fen was not drained, thanks to an enclosing sandy ridge. Therefore, peat continued to grow in this core area. Due to mineralization of peat and spring erosion that led to gully forming in the surrounding part of the mire, base-rich species were found in zone 4 (Figure 4) again.
Most records used to assess ecosystem changes are based on short-term ecological information, spanning only a few decades. However, Willis and Birks (2006) highlight the potential of paleoecological records in conservation biology, including application to biodiversity maintenance. With paleoecological methods we can reconstruct successions in a longer-term perspective. With our reconstruction of the vegetation succession and changing environmental conditions of the Mosbeek site we provide a longer temporal perspective to address conservation issues of base-rich fens (compare Faber et al., 2016). The awareness of links between culture-historical changes in the landscape and the start of peat growth may be a useful tool for present-day and future nature management. Nowadays, human impact on fen ecosystems is usually negative, as increased drainage and groundwater extraction for drinking water and irrigation leads to lowering of water levels and decrease in groundwater discharge. However, this study shows that human impact can also have positive effects. The Mosbeek fen was not a relic from the early Holocene, but resulted from changes in land use during medieval times.
Footnotes
Acknowledgements
The authors thank Landschap Overijssel, the State Forest Service Overijssel, and the Steggink family for permission to take samples and for funding radiocarbon-dating. The water authority Vechtstromen owes thanks for detailled measurements of the rivulet Mosbeek. Annemarie Philip prepared the microfossil slides and Stefan Engels kindly prepared diagrams. Louis Lansink, Fons Eysink, and Loekie van Tweel-Groot kindly helped with the site selection.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.