
Abstract
In the current global warming phase, relict glacial areas are one of the most threatened ecosystems in the world. They are “cold-spots” of biodiversity and of great interest both from the ecological and conservation point of view. We investigated the biological communities (plants and arthropods) hosted by one of the southernmost European glaciers: Calderone Glacier, the last preserved glacier of the Apennines mountain chain (Italy). We analyzed supraglacial debris and the nearby moraine and we found a rather diverse and peculiar biodiversity, which includes also new species. Some arthropods, such as the springtail Desoria calderonis, are particularly sensitive to the presence of ice at microtopographic level. Among plants, only Arabis alpina caucasica is able to grow on the supraglacial debris, perhaps because of factors related to the seed germination and seedling survival on this habitat type. Calderone glacier, with its particular biogeographic location, is probably currently acting as one of the last refugia for rare and endemic cryophilic species within the Mediterranean Region during the ongoing warm period.
Keywords
Introduction
Climate warming will be one of the major threats to biodiversity over the next 100 years (Román-Palacios and Wiens, 2020). In cold areas, this phenomenon directly harms several species and habitats (First, 2019; Gobbi et al., 2021; Root et al., 2003), as high altitude ecosystems are particularly threatened (Cauvy-Fraunié and Dangles, 2019; Dullinger et al., 2012; McCain and Colwell, 2011). These ecosystems, especially in peripheral mountain areas, are cold-spot of biodiversity (Cauvy-Fraunié and Dangles, 2019), having contributed to the evolution of several endemic species due their role as microrefugia during glacial and interglacial periods (Muellner-Riehl, 2019; Steinbauer et al., 2016; Zhang et al., 2020).
In the current global warming phase, alpine glaciers at global scale are experiencing a severe reduction but also an increase of supraglacial stony debris, due to freeze-thaw processes delivering large volumes of debris from the surrounding slopes to the glacier surface (Azzoni et al., 2018; Janke et al., 2015; Kirkbride, 1993; Kraaijenbrink et al., 2017; Tielidze et al., 2020). The role of glacier forelands and supraglacial debris as potential habitat and refugium for high altitude organisms, such as plants (Caccianiga et al., 2011; Fickert et al., 2022; Losapio et al., 2021; Tampucci et al., 2015) and arthropods (Gobbi, 2020a; Hågvar et al., 2020; Valle et al., 2020) is increasingly recognized. However, it is still not clear how many cryophilic species – being both cold-adapted and hygrophilous (Deharveng et al., 2008; Fjellberg, 2010; Jurekova et al., 2021) – can live in glacial habitats and how much they are directly related to the presence of ice (Stibal et al., 2020). This information is crucial for estimating possible responses, of this extremely specialized biodiversity, to global change, especially in case of the complete glacial disappearance. Previous studies showed that a significant fraction of plant species living on glacier forelands are considered to be at risk of local extinction following glacier retreat (Losapio et al., 2021). In addition, supraglacial stony debris may host large populations of cold-adapted and/or endemic species of vascular plants (Tampucci et al., 2017), springtails (Collembola), spiders (Arachnida: Araneae) and ground beetles (Coleoptera: Carabidae) (Bernasconi et al., 2019; Buda et al., 2020; Gobbi, 2020a; Valle et al., 2020). Species able to live in glacial or periglacial habitats have also been documented within other taxa (e.g. Coleoptera Staphylinidae (Kapp, 2007), Chilopoda (Gobbi et al., 2020b)).
In Europe, the southern and peripheral mountain areas are widely known as biodiversity hotspots with a high rate of endemic species (Branda et al., 2010; Medail and Quezel, 1999). In these areas, ice masses are very small but still persistent, owing to high winter precipitation and avalanche accumulation, particularly in association with north-facing steep and narrow valleys (Giraudi, 2002; Grunewald and Scheithauer, 2010). However, the ongoing climate change is severely threatening these small ice bodies (Smiraglia and Diolaiuti, 2015). In these areas, glacier-linked biodiversity is unique and could be particularly threatened with extinction.
Calderone glacier is one of the southernmost glaciers in Europe (Grunewald and Scheithauer, 2010; Hughes, 2009), and the only relict glacier of the Apennines. It is located in the Gran Sasso massif in Central Apennines (Abruzzo, Italy) with Corno Grande (2912 m asl) being the highest peak of the entire Apennines and dominating the glacier. This confers to Gran Sasso massif an important biogeographical position, as confirmed by its abundance of endemites together with the nearby Majella massif, compared to other Mediterranean mountain regions (Conti and Bartolucci, 2016; Tammaro, 1971; Testi et al., 1996). Calderone glacier is almost completely debris-covered and has been defined as a glacieret, that is a small glacier body with no dynamicity and erosional activity and without an accumulation and an ablation zone (Smiraglia and Diolaiuti, 2015), with a negative trend in glacier area and mass over the past two decades (Pecci et al., 2008). This unique glacial system is thus near the definitive disappearance. No detailed information is available about organisms living on Calderone glacier, although its isolated position within the Mediterranean region (following the Apennine chain, the nearest glacier is Clapier in Maritime Alps, about 530 km away) might suggest an important ecological and biogeographic role for high mountain biodiversity.
This work aims to provide an overview of plant and arthropod communities hosted by Calderone glacier, investigating the species distribution on the glacier surface and in the surrounding habitats and its alpha and beta diversity. Our goals, specifically are: (1) to identify and functionally characterize supraglacial and proglacial communities of Calderone glacier; (2) to investigate the microtopographic drivers that affect species distribution (debris thickness and soil parameters); (3) to search for species directly related to the presence of ice, currently the most threatened by global warming.
Material and methods
Study area
Calderone glacier (42°28′’16.4″N 13°34′’01.4″E) is located in the Gran Sasso Massif, in the Central Apennines (Italy), surrounded by steep carbonate stonewalls and overlooking the Adriatic Sea with a North-East aspect (Figure 1). It is the only glacier present on the Apennines (Smiraglia and Diolaiuti, 2015). The Little Ice Age (LIA; mid 19th century; D’Orefice et al., 2000; Rovelli, 2006) moraine borders the tongue, very close or in contact with the glacier; the distance between the moraine base and the ice cannot be assessed, the area being covered by debris.
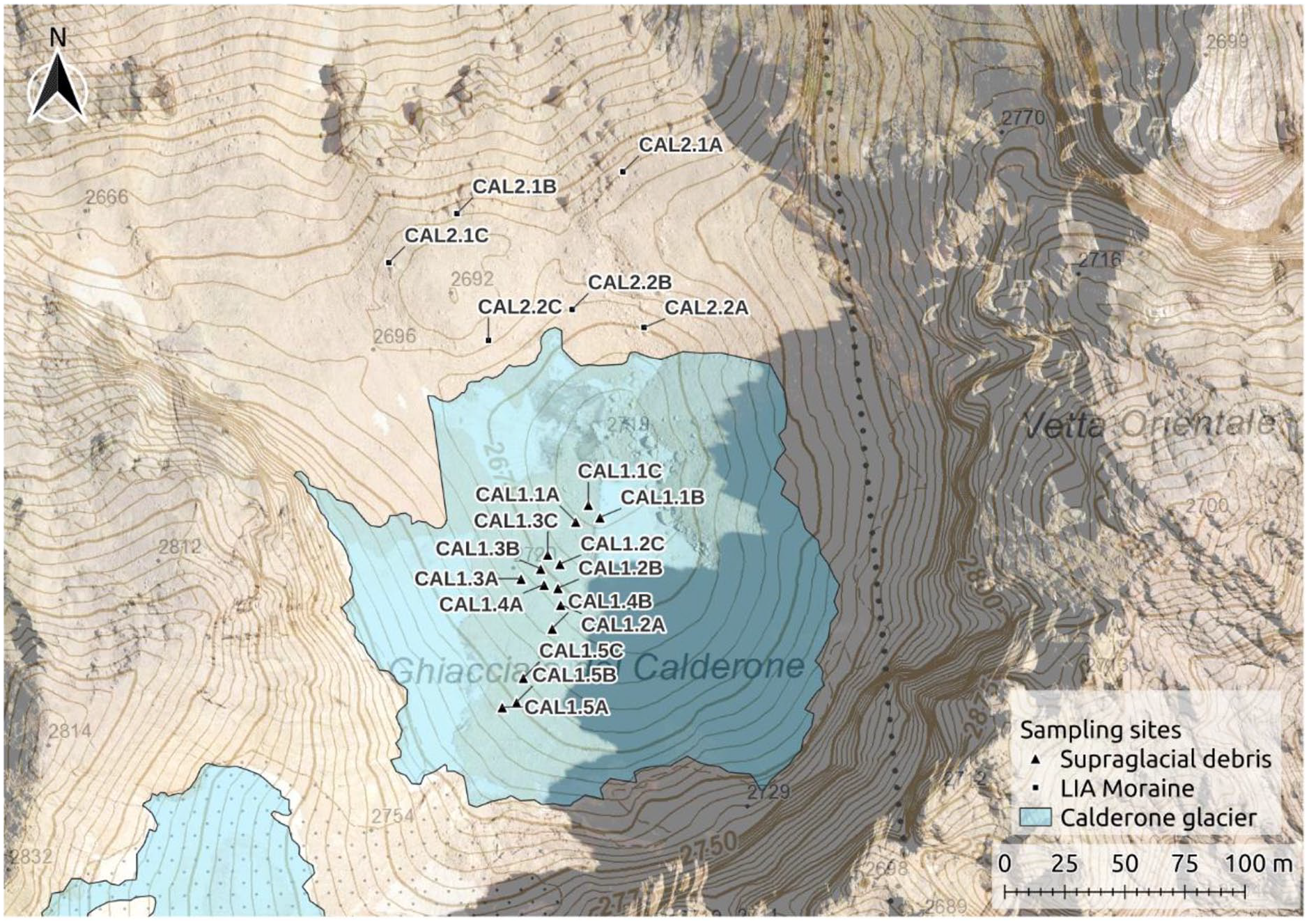
Calderone glacier and sampling design of the field work performed during July 2020.
Calderone glacier is fragmented into two ice masses, after the separation of the tongue (Lower Calderone) from the accumulation zone (Upper Calderone) occurred in 2000 (De Sisti et al., 2003; Pecci et al., 2001). In 2011 the total surface was 0.04 km2, with a reduction of 20% from 1981–84 and of 33% from 1959–62 (Smiraglia and Diolaiuti, 2015). The thickness of the ice was 15 m at the end of the 20th century (Smiraglia and Diolaiuti, 2015). The glacier is now almost completely covered by stony debris coming from the surrounding rock walls.
Sampling design
Two environmental units were selected: the supraglacial stony debris on the Lower Calderone (Habitat 8340; Council Directive 92/43/EEC, 1992) and the LIA moraine (Habitat 8120, Council Directive 92/43/EEC, 1992). Specifically, we placed 14 sampling points on supraglacial stony debris and 6 on the LIA moraine (Figure 1). Sampling points are spaced from each other by at least 2 m. In each sampling point:
Vegetation sampling was performed in a 2 m × 2 m plot and the plant species cover was reported as a percentage.
Pitfall trap was set out to sample ground-dwelling arthropods. A Pitfall trap consist of a plastic glass (diameter 7 cm) filled up with a non-toxic and frost-resistant solution of water and vine winegar (2:1) with salt and soap to catch and preserve the animals during the sampling period (Gobbi, 2020a). In order not to reduce the impact on such a small and delicate habitat (Lencioni and Gobbi, 2021) we performed only two sampling session in the period of maximum activity for arthropods (Gobbi, 2020a): 7–8 July/9–10 July 2020 (2 days of trap activity) and 9–10/29 July 2020 (20 days of trap activity).
Soil microarthropods were sampled using the flotation method (Marshall et al., 1994) on debris samples collected with a scoop at different depths (at 1, 10, 20 and 50 cm), in order to obtain a representative sample of the whole ground/soil profile, placed in a plastic glass (diameter 7 cm, height 8 cm), and screened for arthropods in a small basin filled with mineral water.
A soil sample of 200 g was collected at the surface where to measure soil pH (in 1:2.5 soil:water), organic matter (Walkley–Black method) and carbonate calcium content (Dietrich–Fruhling calcimeter).
Sub-surface ground temperature was recorded 10 cm below the ground surface for 1 year (9 July 2020–27 July 2021) every 2 h using iButton 1922 temperature probes. Fifteen probes were placed in the study area, 14 in the supraglacial stony debris in correspondence with the sampling points and one as control, on the external side of the LIA moraine (in correspondence with the sampling point CA2.1A). Moreover, to collect relative humidity (RH%) two Tinytag plus 2 dataloggers were placed 10 cm below the soil surface, one in the supraglacial stony debris (in correspondence with the sampling point CAL1.4A) and one on the internal side of the LIA moraine (in correspondence with the sampling point CAL2.2A). In order to estimate the max and the min temperature we calculated respectively the mean value of the max temperature from the 10 hottest and coldest days.
On supraglacial debris, thickness was measured for every sampling point, digging through the debris until the underlying ice was reached.
Collected organisms were identified, when possible, to the species level by experts (See Supplementary Material 8, available online also for nomenclature references). Adult specimens were identified; juvenile specimens were identified where possible: for springtails, pseudoscorpions (Pseudoscorpiones), harvestmans (Opiliones) and earwigs (Dermaptera). The spider Pardosa sp. and the springtail Sminthurides sp. were not identifiable at species level because all the available specimens were juvenile, but Pardosa sp. was clearly different from Pardosa cavannae. Deutherosminthurus sp., Hypogasrura sp. and Pseudisotoma sp. were not identifiable at species level because of the availability of only one or few specimens.
Collected specimens are preserved in ethanol in permanent collections now stored at: Museo civico di Scienze Naturali di Bergamo (Italy) (spiders, harvestmans), MUSE – Science Museum of Trento (Italy) (ground beetles, snout beetles (Curculionidae), earwigs, springtails, other taxa not identified), Museo civico di Storia naturale di Verona (pseudoscorpions, rove beetles with the exception of Coryphium atratum, preserved in Zanetti, 1984).
Dataset building
Data were ordered in a matrix associating each sampling point with location data (latitude, longitude, altitude, slope, aspect), soil data (stony debris thickness, pH, soil organic matter content and soil carbonate calcium content), vegetation data (values of vegetation cover), and fauna data (number of individuals). Arthropod abundances were transformed in activity density (ratio amongst the number of collected individuals and the days of trap activity) for pitfall traps (PT) and calculated as absolute number of specimens sampled for flotation method (FM).
Ground beetles, spiders, springtails, rove beetles, harvestmen, snout beetles, earwigs, pseudoscorpions, oribatids (Oribatida) were identified at species level; millipedes and other orders of mites were analyzed only at class level. All taxa with high dispersal ability (i.e. winged species) (Supplementarl Material 4, available online), were not included in the analysis but reported as “airborne arthropod fallout (namely aeroplankton).”
We assigned a binary factor (presence/absence) to each species in relation to its geographic and altitudinal range. Specifically, we used five chorological categories: endemic of Gran Sasso and Majella, endemic of Apennines, endemic of European Alps and Apennines, occurring on Southern European mountains and species with wider distribution (also if some chorotypes are included in others, we considered this trait as mutually exclusive, assigning to every species only the more restrictive category).
Based on their altitudinal distribution, the species were divided in three main groups: alpine species occurring above the treeline (about 1800 on average on Apennines), low alpine species occurring from 1000 m asl up to the treeline, “lowland” species with a wider altitudinal distribution.
The binary factor (yes/no) was used also to identify cryophilic species: we considered as cryophilic those species that are both cold-adapted and hygrophilous (Deharveng et al., 2008; Fjellberg, 2010; Jurekova et al., 2021). Thus, snow-related species were included in this category, being adapted to cold environments characterized by high humidity.
The loss of the ability to fly is also an important trait for high altitude species, since it reduces dispersal ability; for this reason, we divided the species in winged, brachypterous and apterous (not considering taxa not winged for phylogenetic reason: e.g. springtails). We grouped the species on the basis of feeding habits: predators, saprophagous, phytophagous, spermophagous, parasitoids, microorganism consumers, detritivores; the same species can occur in more than one group due the heterogeneity of its diet.
Information about these variables were found for plants in Bartolucci et al. (2018) and Pignatti (2017); for harvestmans in Phillipson (1960), Hågvar and Flø (2015) and Hillyard (2005) (We considered Mitopus sp. both predator and saprophagous because, although harvestmen are scavengers, altitude ones are known as predators); for oribatid in Hajmová and Smrš (2001) for pseudoscorpions in Gardini (2000); for spiders in Pantini and Isaia (2019), Pekár et al. (2021), Thaler (1986); for springtails in Bretfeld (1999), Gisin (1960), Mateos (2011), Pomorski and Kaprus (2007), Potapov (2001), Potapov et al. (2016), Thibaud et al. (2004); for earwigs in Costa (1882), Galvagni (1970) and Stoch (2000); for ground beetles in Brandmayr (2005), de Jong et al. (2014) and Pesarini and Monzini (2011); for rove beetles in Kapp (2007), Koch (1989) and Loebl and Loebl (2015); for snout beetles in Abbazzi and Maggini (2009). Wing morphology was detected by a stereomicroscope.
Data analysis
An principal component analysis (PCA) was performed on soil data to evaluate the association among all variables and to rule out from the subsequent analyses some of the auto-correlated variables.
Species richness was compared between the two environmental units, that is, supraglacial debris, and the LIA moraine. A T-test was used to assess the differences in species richness and functional composition between supraglacial habitats and the LIA moraine. For each functional category we estimated both the species richness and their incidence (number of species in the selected category divided by the total number of species sampled). For identifying typical plant and arthropod species of each environmental unit, we used Indicator Species Analysis (IndVal; Podani and Csanyi, 2010). To assess beta diversity for arthropod taxa we performed a permutation test of multivariate homogeneity of group dispersions (Anderson et al., 2006). The function betadisper in the ʻveganʼ package (Oksanen et al., 2019) was used to perform the PERMDISP analysis. This method consists in measuring the average distance from each observational unit to their group centroid and assessing variability in community structure. Average dissimilarity of sampling sites within environmental units (supraglacial debris, and the LIA moraine) was assessed using the Whittaker index (De Cáceres et al., 2013; Faith et al., 1987; Whittaker, 1952). To test for statistical differences in beta diversity between environmental units, 999 permutations were performed and tested using one-way ANOVA. We investigated the relationship between species richness and soil characteristics in the sampling point located in the supraglacial habitat using linear models (LM). The LMs were performed considering linear, quadratic and cubic terms and the model with the highest R2 was chosen. In the same way, a Poisson generalized linear model (GLM) was used to test the relationship between arthropod abundance (see Dataset Building) and soil characteristics: debris thickness, pH, soil organic matter content (g/kg) and soil carbonate calcium content (%). The species that showed less than three occurrences in the supraglacial sampling points were excluded. The GLMs were performed both with and without quadratic terms. For each species and each soil variable, the model with the highest D2 was chosen.
Data processing was performed with R 4.0.2 (R Core Team, 2020), using the following packages: ggplot2 (Wickham, 2016), tidyverse (Wickham et al., 2019), dplyr (Wickham et al., 2020), patchwork (Pedersen, 2020), ggpubr (Kassambara, 2020), vegan (Oksanen et al., 2019).
Results
The average annual (July 2020–July 2021) sub-surface ground temperature recorded on the supraglacial debris is 0.7°C; snow covered the glacier surface for 250 days, while average annual temperature on the LIA moraine was 1.3°C and snow cover lasted 146 days (Tables 1 and 2).
Summary table of climatic variables recorded on Calderone glacier.
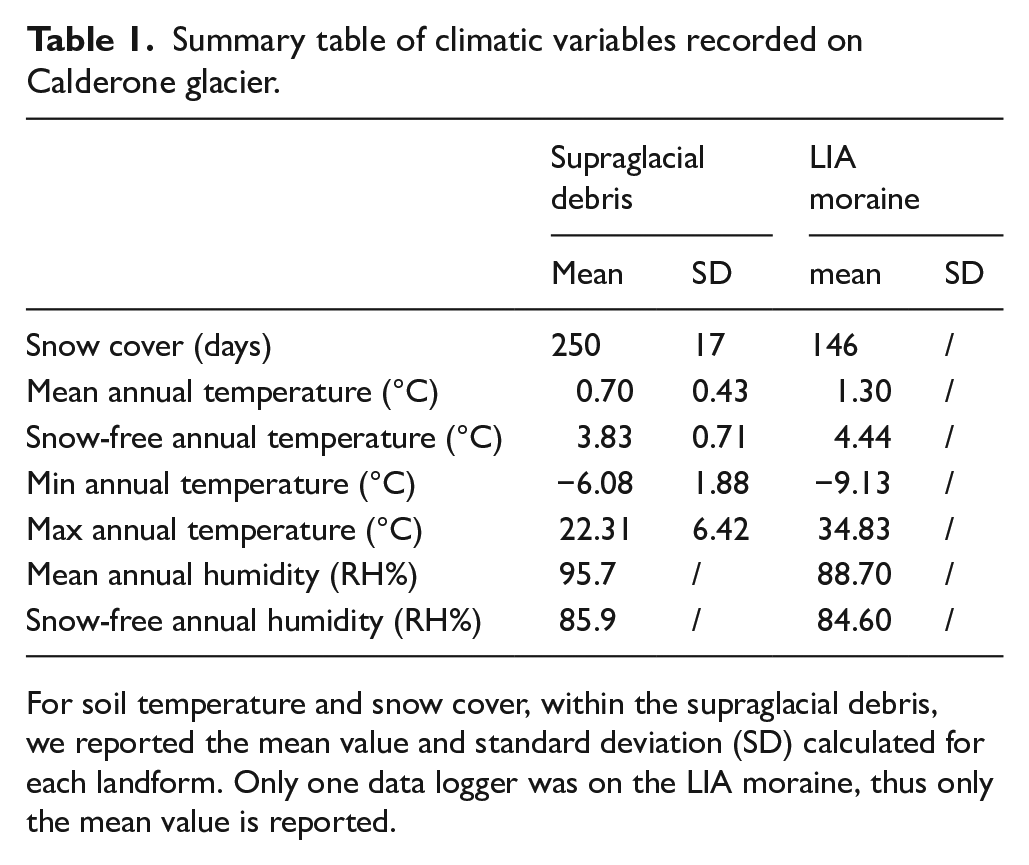
For soil temperature and snow cover, within the supraglacial debris, we reported the mean value and standard deviation (SD) calculated for each landform. Only one data logger was on the LIA moraine, thus only the mean value is reported.
Species found in the two environmental units with the two sampling methods.

Mean soil parameters of the two units. For all other traits and differences among sampling methods, see Supplemental Material 7, available online.
Ap: endemic of Apennines; GS-M: stenoendemic species of Gran Sasso and Majella.
p=plants, c=ground beetles, coll= springtails, sp=spiders, st= rove beetles, cr= snout beetles, op= harvestmen, ac= mites, ps= pseudoscorpions, dr= earwigs, dp= millipedes.
Cryophilic species.
The LIA moraine (organic matter content = 3.88 g/kg and pH = 8.28) showed a higher organic content than supraglacial debris (organic matter content = 1.68 g/kg and pH = 8.62) (Table 2). The sampled supraglacial debris had a mean thickness of 23 cm with a range from 8 to 40 cm. PCA (Figure 2) highlights that organic matter content and pH are the most important variables to characterize the glacier environment.
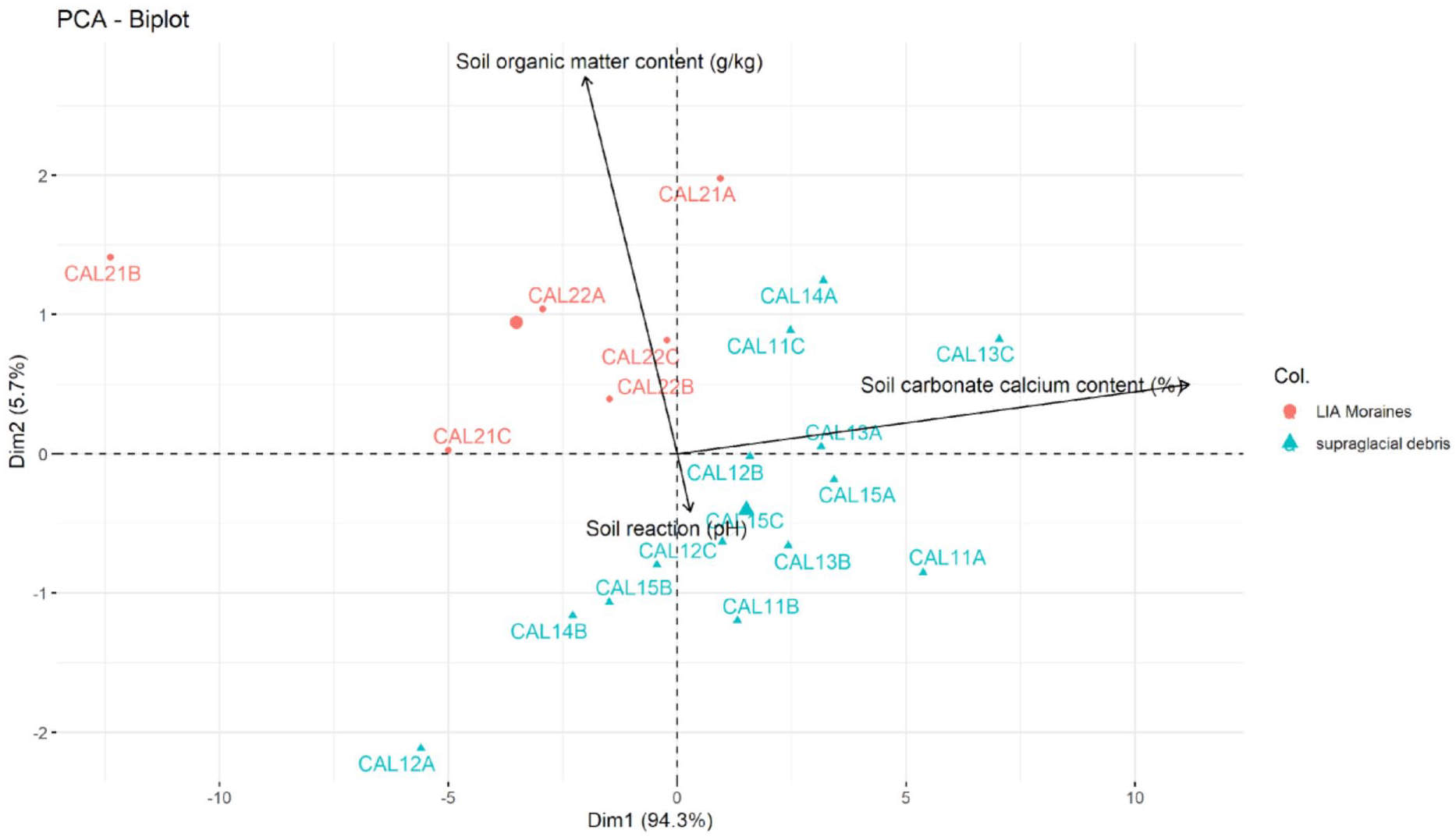
Principal component analysis (PCA) on soil data evaluates the association among all variables.
Identification and characterization of Calderone supraglacial and proglacial communities
Forty-five species were found among the studied taxa (Table 2), 37 arthropods and eight vascular plants. Six (13%, all arthropods) are steno-endemic (species with narrow distribution range) of Majella and Gran Sasso and six (13%, one plant and five arthropods) are endemic of the Apennines (Table 2). Five species (11%, all arthropods) are cryophilic: the ground beetle Nebria orsinii orsiniii, the rove beetle Aleochara meridionalis, the pseudoscorpions Neobisium fiscelli and N. osellai and springtail Desoria calderonis (Table 2). Airborne arthropod fallout is reported in Supplemental Material 4, available online. Among all arthropod species, 34 were sampled with the pitfall trap method and nine with the flotation method; three species were caught only with the flotation method (the springtails Pseudisotoma sp. and Protaphorura octopunctata and the rove beetle Liogluta roettgeni).
Comparing the biological communities of supraglacial debris and the LIA moraine, we found a strong difference in species richness of plants and arthropods (Tables 2 and 3). On supraglacial debris, we found only Arabis alpina caucasica (species richness for sampling ranges from 0 to 1, mean value 0.1), while on the LIA moraine also Cerastium thomasii, Hornungia alpina, Poa alpina, Saxifraga exarata, S. oppositifolia, Papaver alpinum alpinum, and Sedum atratum were observed (species richness in each sampling ranges from 0 to 5, mean value 2.7). Arthropods species richness (considering both pitfall trap and flotation methods) was even more significantly higher on the LIA moraine (p-value = 0.031) and ranges from 5 to 15 (mean value 9.3) while on supraglacial debris it ranges from 2 to 9 species (mean value 4.1) (Tables 2 and 3).
Differences in species composition between the two communities (mean values of species and the mean incidence) in relation to every functional trait respectively for supraglacial debris and LIA moraine.
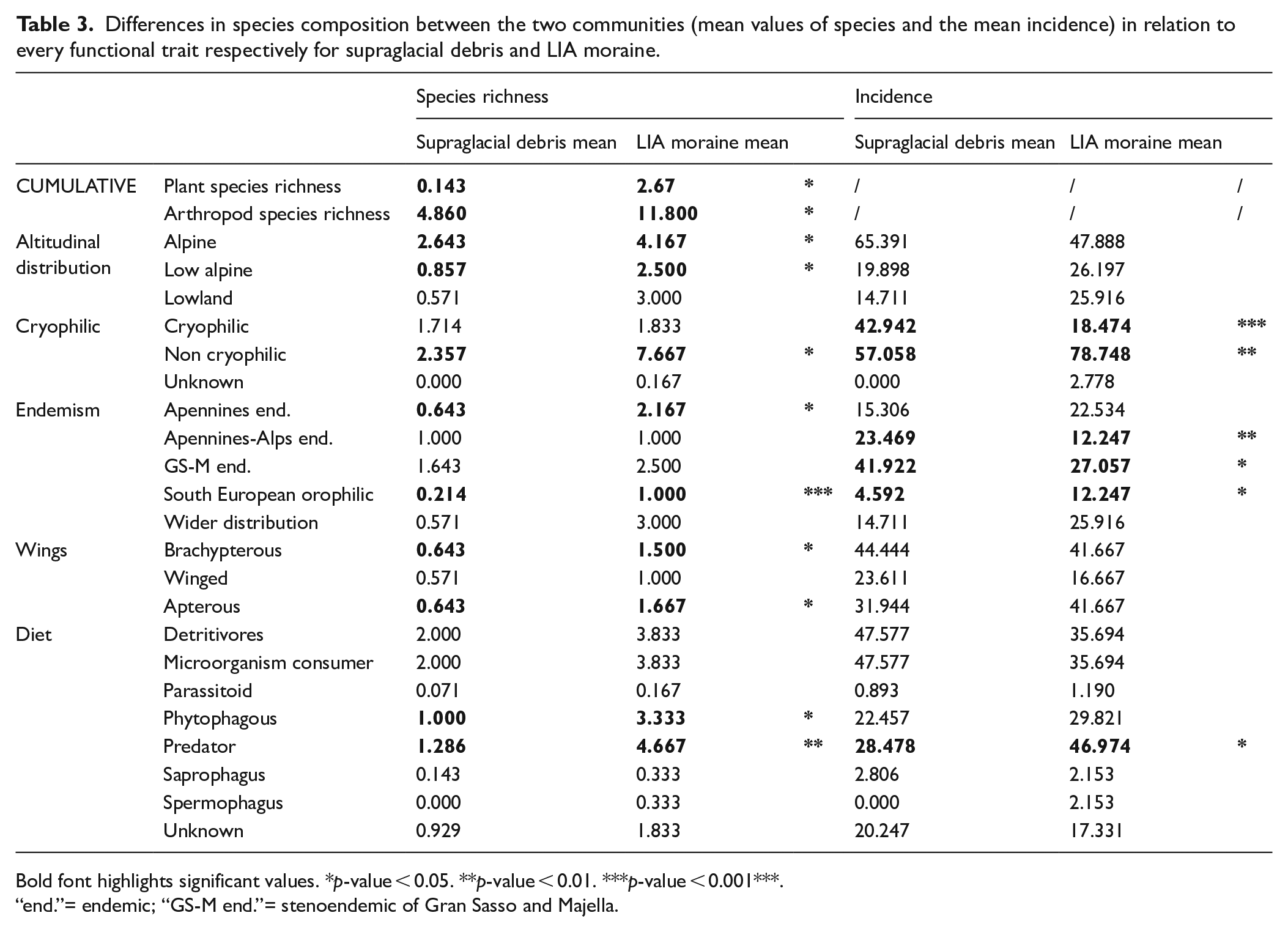
Bold font highlights significant values. *p-value < 0.05. **p-value < 0.01. ***p-value < 0.001***.
“end.”= endemic; “GS-M end.”= stenoendemic of Gran Sasso and Majella.
Analysis of beta-diversity showed how the species assemblages of each of the considered environmental units are different (Supplemental Material 1, available online, Figure 2). According to the IndVal analysis (Supplemental Material 5, available online, Figure 5), Desoria calderonis is the only arthropod indicator species of supraglacial debris; conversely, indicator species of the LIA moraine are the ground beetle Amara samnitica, the springtails Pseudachorutes palmiensis and Lepidocyrtus spp., the spider Pardosa cavannae, the snout beetle Donus oblongus and Diplopoda. Common species of the two environmental units are the ground beetle Nebria orsinii orsinii, the springtail Orchesella cf. frontimaculata, the spider Agyneta rurestris, the rove beetles Aleochara meridionalis, Anotylus nitidulus, Tachyporus nitidulus, and the earwig Forficula apennina.
Stenoendemic species of Majella and Gran Sasso are the most represented on both the LIA moraine and supraglacial debris, and are dominant on supraglacial debris (incidence respectively 27% and 42% in the two communities, p-value = 0.016; Table 3, Supplemental Material 2, available online). Endemic species of the Apennines showed the highest species richness on the LIA Moraine (p-value = 0.011). The incidence of endemic species of Alps and Apennines were significantly higher in the supraglacial debris (p-value < 0.01). South European orophilic species were significantly higher on the LIA moraine both considering the species richness and their incidence (p-value < 0.001, p-value = 0.043, respectively).
Considering altitudinal distribution, we found that the LIA community has a significantly (or almost significantly) higher species richness of alpine and low alpine species. On supraglacial debris there is a higher incidence (65%; p-value = 0.097) of alpine species and a similar incidence of mountain (20%; p-value = 0.162) and generalist (15%; p-value = 0.272) species (Table 3, Supplemental Material 2, available online).
The species richness of brachypterous and apterous species are higher on the LIA moraine (respectively, p-value = 0.01 and p-value = 0.026) (Table 3, Supplemental Material 2, available online).
The incidence of cryophilic species is higher on the supraglacial debris (p-value < 0.001), while non-cryophilic species are more incident on the LIA moraine (p-value = 0.001; Table 3, Supplemental Material 2, available online).
The feeding strategy distributions were different between LIA moraine and supraglacial debris (Table 3, Supplemental Material 3, available online): while on supraglacial debris detritivores and microorganism consumers were dominant (mean value of about 48%, corresponding to all the present springtails) and predators accounted for 28% (only Nebria orsinii orsinii, Agyneta rurestris, Pardosa sp. and Tachyporus nitidulus), the LIA moraine community was dominated by predators (mean of about 47%, p-value = 0.021: the ground beetles Nebria orsinii orsinii, Pterostichus morio samniticus, Amara samnitica, the spiders Pardosa cavannae, Agyneta rurestris, Oreonetides glacialis, Entelecara sp.n. Improphantes cf. improbulus, the rove beetle Quedius obscuripennis, the harvestman Mitopus sp. and both pseudoscoropions, Neobisium fiscelli, Neobisium osellai). Other feeding strategies were less represented, especially on supraglacial debris. The species richness of phytophagous was significantly higher on the LIA moraine (p-value = 0.039) including the snout beetles Dichotrachelus variegatus variegatus, and Donus oblongus, the springtails Orchesella cf. frontimaculata, Lepidocyrtus cf. lignorum, Lepidocyrtus cf. tellecheae, Tomocerus vulgaris and Deutherosminthurus sp. and the opportunistic oribatid Tectocepheus velatus velatus. On supraglacial debris the only phytophagous were the opportunistic Orchesella cf. frontimaculata and Tectocepheus velatus velatus and the common snout beetle Protapion trifolii. Saprophagus and spermophagus are not significantly different between the environmental units, since the only spermophagous was the ground beetle Amara samnitica, which is also a predator, on the LIA moraine. The only saprophagous was Mitopus sp., ubiquitous between the two environmental units. Parasitoids are represented only by the rove beetle Aleochara meridionalis, found in both environmental units with a single specimen.
Microtopographic drivers affecting species distributions on supraglacial debris
On supraglacial debris, the highest species richness was found in correspondence with the mean value of debris thickness, about 20 cm; Figure 3).
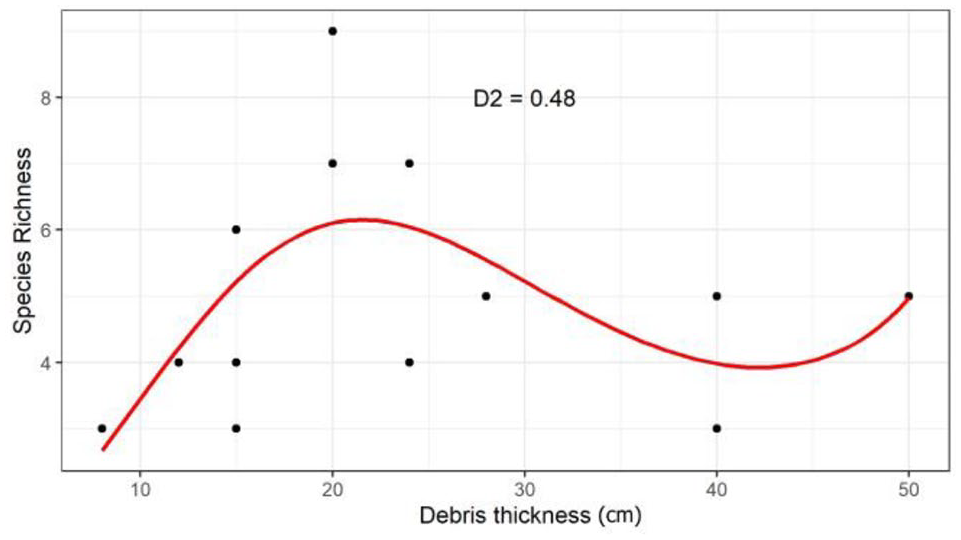
Generalized linear model testing the relationship between arthropod species richness (estimated with both pitfall trap and flotation method) and debris thickness (SDS) of supraglacial debris.
The springtail Orchesella cf. frontimaculata was positively related to debris thickness; this link is more notable with the pitfall trap method (PT: R2 = 0.5) than with the flotation method (FM: R2 = 0.44) (Figure 4). The springtail Desoria calderonis, on the other hand, is more abundant where the ice is closer to the surface; this result is more significant with the floatation method (FM: R2 = 0.64), than with the pitfall trap method (PT: R2 = 0.32). With the exception of the ground beetle Nebria orsinii orsinii, which seemed to slightly prefer a lower debris thickness (R2 = 0.12), the abundance of other organisms considered – Acari, the spider Agyneta rurestris and the earwig Forficula apennina – showed a slight positive relation to a thicker debris.
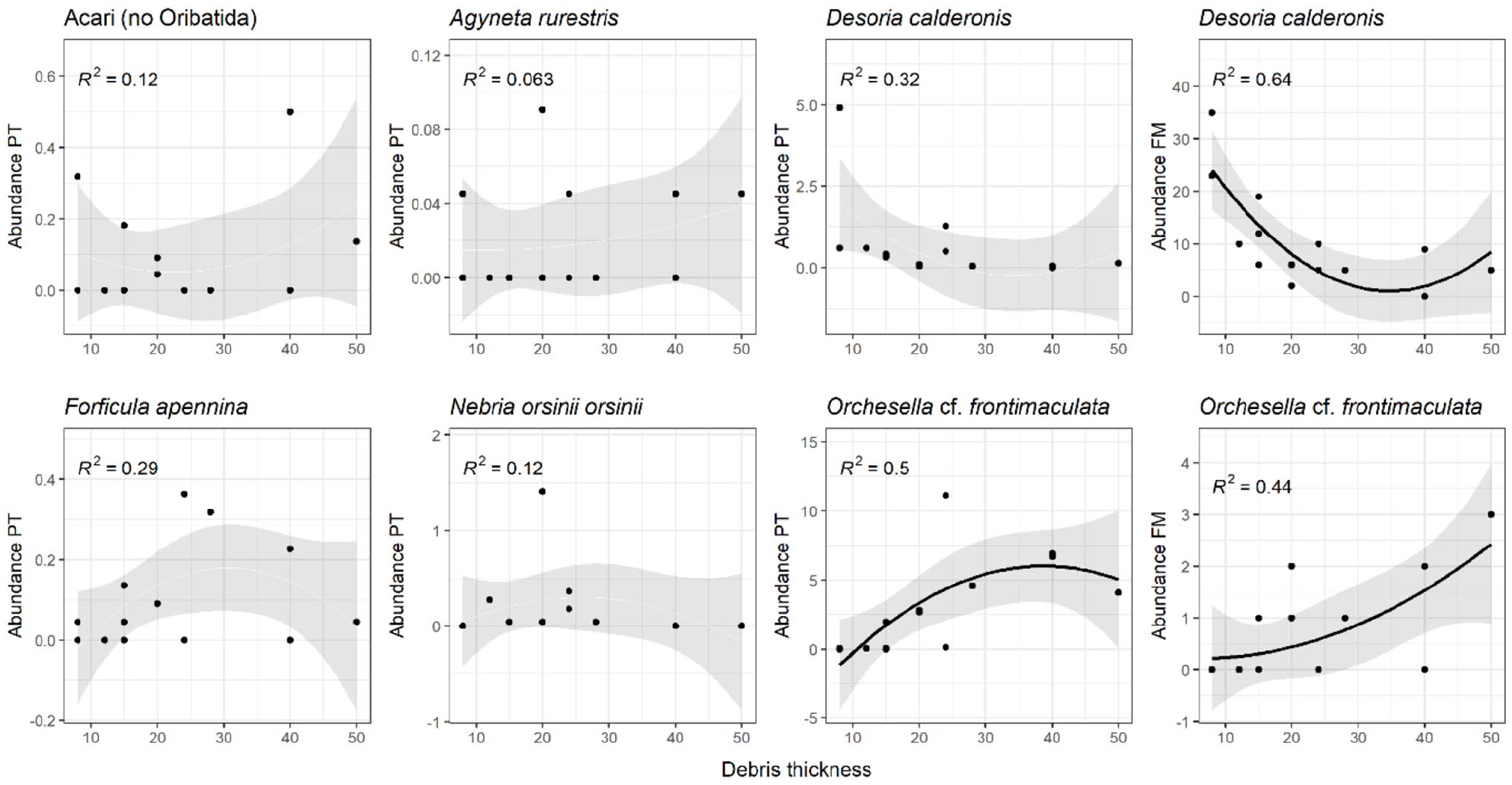
Linear models testing the relationship between species abundances and debris thickness (cm) on supraglacial debris. PT = samples collected with pitfall trap method and abundance calculated as activity density (AD), FM = samples collected with flotation method and abundance corresponding to absolute number of specimens collected. The standard error of the pattern is reported; the black line was reported only for significant models.
Only Acari abundance, excluding oribatids, (R2 = 0.29) seemed positively, although weakly, related to the abundance of organic matter (Supplemental Material 6.A, available online). Acari (without oribatids) abundance was also the most negatively related to pH values (R2 = 0.61) (Supplemental Material 6.B, available online). The abundance of spider Agyneta rurestris (R2 = 0.39) and of the ground beetle Nebria orsinii orsiniii (R2 = 0.69) are weakly negatively related to soil carbonate calcium content (Supplemental Material 6.C, available online).
Discussion
Peculiarities of supraglacial and proglacial communities
Even in such a limited space as the supraglacial and proglacial areas of Calderone glacier, biological communities are very well differentiated among the considered environmental units: the supraglacial debris and the LIA moraine (we summarize the most important features in Figure 5). The first discriminant between the two environmental units is the species richness, both plants and arthropods, which is markedly lower on supraglacial debris.
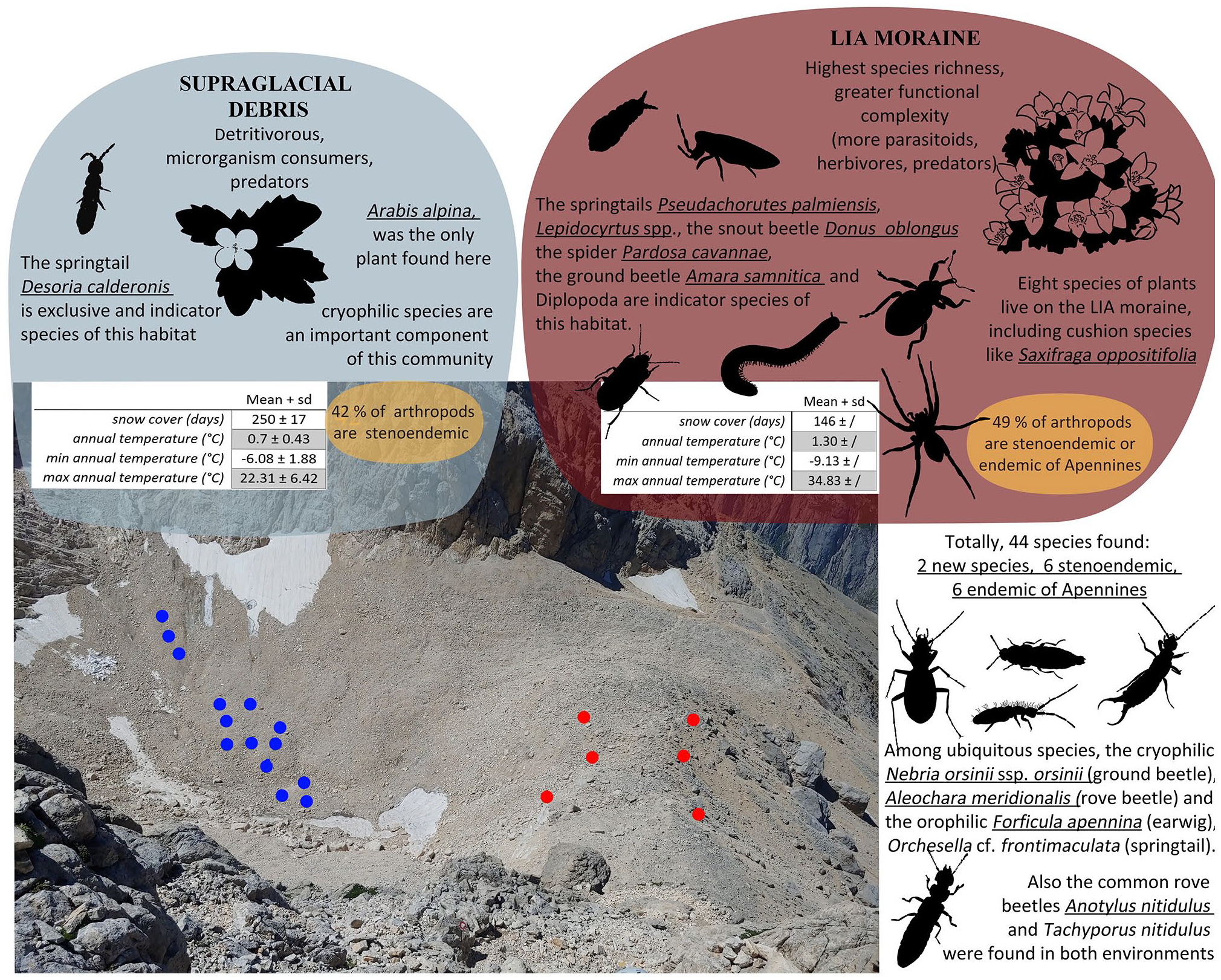
Graphical synthesis of the characteristics of the two communities. Blue and red points represent sampling points respectively on supraglacial debris and on the LIA moraine (their position is approximate, for their actual distribution see Figure 1). Taxa represented as characteristic of the two environmental units are those found with the IndVal analysis. Organisms are not in scale. Underlined species are those represented in the figure.
Cryophilic species – as observed in previous studies (Lamprecht et al., 2018; Valle et al., 2020) – are effective in distinguishing supraglacial debris from the LIA moraine species assemblages considering their incidence. However, their richness is similar between the two considered environmental units: we hypothesize that the moraine could be influenced directly by the proximity to the ice allowing most cryophilic species to live in this environment. Only the cryophilic springtail Desoria calderonis does not colonize the LIA moraine despite the proximity to the ice.
On the other hand, the LIA moraine hosts many arthropod species that are not strictly cold- or wet-loving, but characteristic of high-altitude open environments: here the trophic chain is more complex, characterized by the presence of a multiple trophic levels, including a significative incidence of herbivores. This can be explained by the differences observed in plant cover, organic matter content and stability between the LIA moraine and supraglacial debris.
Despite the potential intake of seeds from the surrounding environment (e.g. rock walls), only Arabis alpina caucasica grows on supraglacial debris, while on the nearby moraines eight plant species are present. This evidence suggests that not only the dispersal ability but also seed germination and seedling survival traits act as limiting factors in determining plant growth on supraglacial debris. Indeed, Arabis alpina shows a positive response to vernalization (i.e. the exposure of plants or seeds to low temperatures) (Wang et al., 2011). Vernalization and, more in general, seed dormancy can contribute to the maintenance of long-lived seed banks by ensuring that germination is either delayed until the onset of favorable conditions, or that the seed bank does not germinate all at once (Baskin and Baskin, 1998; Fenner and Thompson, 2005; Ooi et al., 2009). Thus, seed dormancy could be an adaptive strategy to survive in cold habitats (Schwienbacher et al., 2011). Moreover, Arabis alpina shows a high phenotypic plasticity (Andrello et al., 2016; de Villemereuil et al., 2018), perhaps owing to its high genetic diversity in the Mediterranean mountains (Koch et al., 2006). Indeed, as proof of its plasticity, this species is widespread over a wide altitudinal range, from 800 to 3100 m asl (Pignatti, 2017).
The comparable incidence of wing reduction or absence on moraine and supraglacial debris suggests that the reduced mobility in ground-dwelling fauna is a common and quite constant characteristic of organisms living at high altitude, specifically in glacial habitats (Gobbi et al., 2017). Reduced mobility is typical of species with a restricted distribution range, both geographical and altitudinal (Pizzolotto et al., 2014; Sattler et al., 2014): both environmental units, in fact, have an high incidence of endemic and stenoendemic species, the latter being even dominant on supraglacial debris. Concerning the altitudinal distribution range, we observed the same phenomenon: high altitude species show higher richness on the LIA moraine, but they are dominant on supraglacial debris.
Calderone glacier gives evidence of the “Collembola first principle” (Hagvar and Gobbi, 2022), being these pioneer organisms crucial for glacial habitat. They graze on biofilm (Hågvar and Pedersen, 2015) as well as on airborne organic matter (Ingimarsdóttir et al., 2014; König et al., 2011) and constitute a local source of food for all predators (Hågvar and Ohlson, 2013; Sint et al., 2019). Nevertheless airborne arthropod fallout found includes families of forestry and winged (i.e. long distance dispersers) beetles, indicating also “allochthonous” sources of preys relatively far from the glacier.
From the methodological point of view, the use of two sampling methods for arthropod was fundamental to obtain more complete data on the communities studied, whereas the communities reflected by the two methods singularly were very different. In particular, among springtails the flotation method caught the only euedaphic species found, Protaphorura octopunctata, well reflected the distribution of Desoria calderonis in the supraglacial debris and excluded a lot of strongly epiedaphic species like Lepidocirtus spp. and Tomocerus vulgaris, suggesting that this method might be very useful to study soil fauna of mineral soils (Marshall et al., 1994). Nevertheless, the pitfall trap method better reflects the general complexity of the two communities studied.
The importance of glacial refugia, “cold-spots” of biodiversity
Calderone glacier, despite its limited extension, proved to be a great coffer of exclusive biodiversity, since endemic species are a very important part of its glacial communities. We discovered two new species on Calderone glacier, the springtail Desoria calderonis (sampled during this field campaign and described in Valle et al., 2021) and the spider Entelecara sp.n. (sampled during this field campaign and currently under description by P. Pantini) taxonomically related to the species Entelecara media Kulczyński, 1887 Among known species, some remarkable taxonomic catches are the rare rove beetle Coryphium atratum, the spiders Improphantes improbulus – a glacial relict according to Thaler (1986) – and both pseudoscorpion species, as after the description by Callaini in 1985 for Abruzzo, there have been no more findings (Gardini, 2000).
In addition, considering the peculiarity and the isolation of Calderone glacier, the poor known springtail Orchesella cf. frontimaculata, described by Gisin (1960) on the basis of pigmentation, is probably a different and new species: pigmentation is not sufficient in distinguish this taxa and chaetotaxy could show a hidden biodiversity (Jordana and Baquero, 2005; Potapov and Kremenitsa, 2008; Smith, 2015). Similarly, Lepidocyrtus spp., as demonstrated by Zhang et al. (2018), could hide a great cryptic biodiversity in relation to habitat type. Thus, it would be necessary to investigate morphology, ecology and genetics of these entities in order to verify their taxonomic position (Frati et al., 2000; Porco et al., 2014; Potapov and Kremenitsa, 2008; Smith, 2015).
The richness of endemic and rare high altitude species, as well as the identification of new entities indicate how much this relict glacier is an important refugium in the current warm-stage period for high altitude biodiversity, a so called “cold-spot of biodiversity” (Cauvy-Fraunié and Dangles, 2019), with low absolute level of biodiversity, but extremely specialized and threatened. This is indicated especially by the presence of cryophilic species with endemic distribution range, as already observed for different taxa such as yeast and yeast-like organisms (Branda et al., 2010). Among such species, the best indicator of the ice presence is the springtail Desoria calderonis, showing a high sensitivity to the microhabitat, unlike other cryophilic organisms observed also outside, although close to the ice. We confirmed the link of Desoria calderonis to the ice, hypothesized in Valle et al. (2021), its narrow ecological niche and thus its role as indicator of cold thermal conditions and of the presence of sub-surface ice. The high specialization of cryophilic springtails (Collembola: Isotomidae) was already highlighted for different mountain regions, from the Rocky Mountains in North America (Fjellberg, 2010) to the Pyrenées in France (Lauga-Reyrel and Lauga, 1995; Najt, 1981), constituting a good indicator of permanent ice.
In the present global warming phase Calderone glacier is experiencing an intense phase of reduction (De Sisti et al., 2003; D’Orefice et al., 2000; Pecci, 2020) which will probably lead to its extinction if the trend of global change does not change. Giraudi (2000) outlined the past fluctuations of Calderone glacier through radiometric dating, demonstrating that it already almost disappeared during the Holocene Optimum. The resulting question is where and how these cryophilic organisms survived during such disappearance phases, a highly critical question in the present climatic phase. It is possible that residual small ice patches covered and protected by stony debris – which considerably reduced their ablation rate (Nakawo and Rana, 1999) – were persistent enough for the survival of these species. Another hypothesis, which does not exclude the first one, is that in this karstic area these organisms may have found a suitable refugium in caves or deep rock crevices where snow can accumulate and persist (Gobbi et al., 2021; Raschmanová et al., 2018). It would be important to test these hypotheses, by searching cryophilic species in proximity to subterranean habitats and performing genetic analysis at population level, in order to define where and what this past refugium was and to understand the fate of these peculiar species in the short-term climate scenario.
The small, vanishing glaciers such as Calderone glacier, located on relatively low mountain chains, could represent a reliable projection of the processes that might occur in the next future on the main glacial areas at higher latitude and altitude.
Conclusions
In two confined environmental units, located in a well studied geographic area of Europe, we found a rare and peculiar biodiversity of high altitude habitats and, even, two previously undescribed species. This biodiversity seems to be related to the biogeographic position of Calderone glacier and its role as a refugium for high-altitude species. In the current climate scenario of temperature increase, which threatens cold-adapted species (Seastedt et al., 2021), it is important to study the ecology of these environmental units in order to monitor glacial “cold-spots” of biodiversity, since an important part of this biodiversity is still taxonomically unknown, especially in peripheral mountains (Gobbi et al., 2021). Only in this way it will be possible to hypothesize also a monitoring and, possibly, a conservation strategy, as well as to forecast the global future patterns and the fate of glacial biodiversity.
Supplemental Material
sj-docx-1-hol-10.1177_09596836221096292 – Supplemental material for Biodiversity and ecology of plants and arthropods on the last preserved glacier of the Apennines mountain chain (Italy)
Supplemental material, sj-docx-1-hol-10.1177_09596836221096292 for Biodiversity and ecology of plants and arthropods on the last preserved glacier of the Apennines mountain chain (Italy) by Barbara Valle, Michele di Musciano, Mauro Gobbi, Marco Bonelli, Enzo Colonnelli, Giulio Gardini, Massimo Migliorini, Paolo Pantini, Adriano Zanetti, Emanuele Berrilli, Anna Rita Frattaroli, Davide Fugazza, Anna Invernizzi and Marco Caccianiga in The Holocene
Footnotes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.