
Abstract
Despite global and local evidence of significant precipitation changes since the Little Ice Age (LIA), their impact upon lake hydrology and surrounding vegetation has yet to be investigated in NW Anatolia. Moreover, the LIA, as a trigger to the social/political recession of the Ottoman Empire in the 17th century is expected to have these impacts. To address this important gap, we studied three cores from Lake Sünnet, a landslide-dammed high altitude lake with extreme sedimentation rates, by using a wide range of proxies including lithology, stable isotopes of O and C, diatoms, pollen, and ostracods. The LIA timespan of AD 1510–1750 is represented by poorly preserved diatom flora and scarce ostracod fauna that collectively suggest very shallow aquatic conditions. The period AD 1640–1710 within the LIA is standing out with higher Abies and total herb pollen percentages that denotes considerably cooler conditions. During and soon after this cold and dry period precisely overlapping the Maunder Minimum, the currently deep Lake Sülüklü in close vicinity was a dry lowland where tall trees were growing. Progressive warming/lake level rise in Lake Sünnet after the AD 1750s is suggested based on a continuous diatom record with increasing planktonic share, increase in diverse and warm demanding arboreal cover, and a positive shift in δ18O of the lake muds. Following the relatively dry/cold period of AD 1800–1850 (Dalton Minimum) when the lake level was stationary and Artemisia cover became the most expanded of the whole record, a relatively warmer climate has dominated the area until the end of the 20th century.
Introduction
Paleoclimatological studies in the last several decades have shown that the Little Ice Age (LIA), traditionally placed between AD 1450 and 1850, was a global climate phenomenon and the coldest of the last 4000 years, though some warmer spells also intervened (Kreutz et al., 1997; Mann et al., 2009; Matthews and Briffa, 2005). The strong co-variance between cosmogenic isotopes (10Be, 14C) and climate proxies in oceanic and ice cores of the time suggest that the change in solar insolation was the main factor for the LIA (Bard et al., 2000; van Geel et al., 1999). Nevertheless, later studies showed a stronger temporal connection between the LIA and volcanic eruptions (Brönnimann et al., 2019; Crowley et al., 2008; Hegerl et al., 2003; Owens et al., 2017). Regardless the reasons, the Atlantic circulation system was influenced by the LIA cooling such that the cooler polar glacial waters continually penetrated the more southern realms and caused ice ages particularly in NW Europe and the Balkans (González Trueba et al., 2008; Hughes, 2009; Mörner, 2010; Zasadni, 2007).
Although the LIA appears as a northern hemispheric cold phenomenon, its precipitation pattern and underlying climatic drivers are highly complex. For instance, a temperature drop of 2°C deduced from the Meso-American marine records is associated with an important precipitation increase as demonstrated by the terrestrial pollen records (Richey et al., 2009). This fact was correlated with the intensified meridional sub-tropical airflows (del Socorro Lozano-García et al., 2007). The LIA in East Africa is characterized by a spatial complexity of alternating centennial dry and wet periods (Russell et al., 2007). In the East Mediterranean, the LIA cooling was accompanied by a significant drought (Kaniewski et al., 2011), whereas the Caspian Sea region in the north was wetter and experienced higher sea levels (Kroonenberg et al., 2007; Leroy et al., 2011). Similarly, most of the proxy records in northeast Asia confirm considerable precipitation increase or peaked instability during the LIA cooling as a result of the southern shift of the Intertropical Convergence Zone (Chen et al., 2019; Rowan, 2017; Yan et al., 2015). Consequently, it is believed that understanding the spatial and temporal dynamics of the LIA could provide important insights regarding ongoing global warming (Free and Robock, 1999; Keller, 2009; Mörner, 2015).
Although numerous studies detected the LIA in Anatolia (Figure 1a), the high-resolution, well-dated records are confined to several varve and tree-ring data from central and western Anatolia, as well as stalagmite and lake records in central, NW, and SW Anatolia (Danladi and Akçer-ön, 2018; Dönmez et al., 2021; Göktürk et al., 2011; Jones et al., 2006; Ocakoğlu et al., 2016a; Touchan et al., 2007; Ülgen et al., 2012). Based on different proxies, these studies demonstrate significant changes in different climate parameters in particular stages of the LIA most likely due to solar variation. Nevertheless, the relative impact of these changes upon the natural environment are poorly understood or even contrasted across Anatolia. For example, Lake Sülüklü in NW Anatolia was completely dry during most of the LIA, consistent with pollen records indicating dry conditions in the surrounding region (Ocakoğlu et al., 2016a; Ülgen et al., 2012). On the other hand, there is an indisputable impact of human beings in addition to climatic shifts on the natural environment in this period in central Anatolia (England et al., 2008). Moreover, paleoclimate data, especially from the boundaries of the different climate belts might be advantageous in recording the hydrological and environmental signals because climate change is associated with shifting biome boundaries through time (Garamvölgyi, 2013; Jia et al., 2019). In this regard, Lake Sünnet is selected for this study because it is located between the arid Central Anatolian and humid Black Sea regions (Figure 1a). Consequently, high-resolution multi-proxy records especially at the conjunction of different climate belts are still required in order to have a wider and clearer climatic and environmental picture of Anatolia.
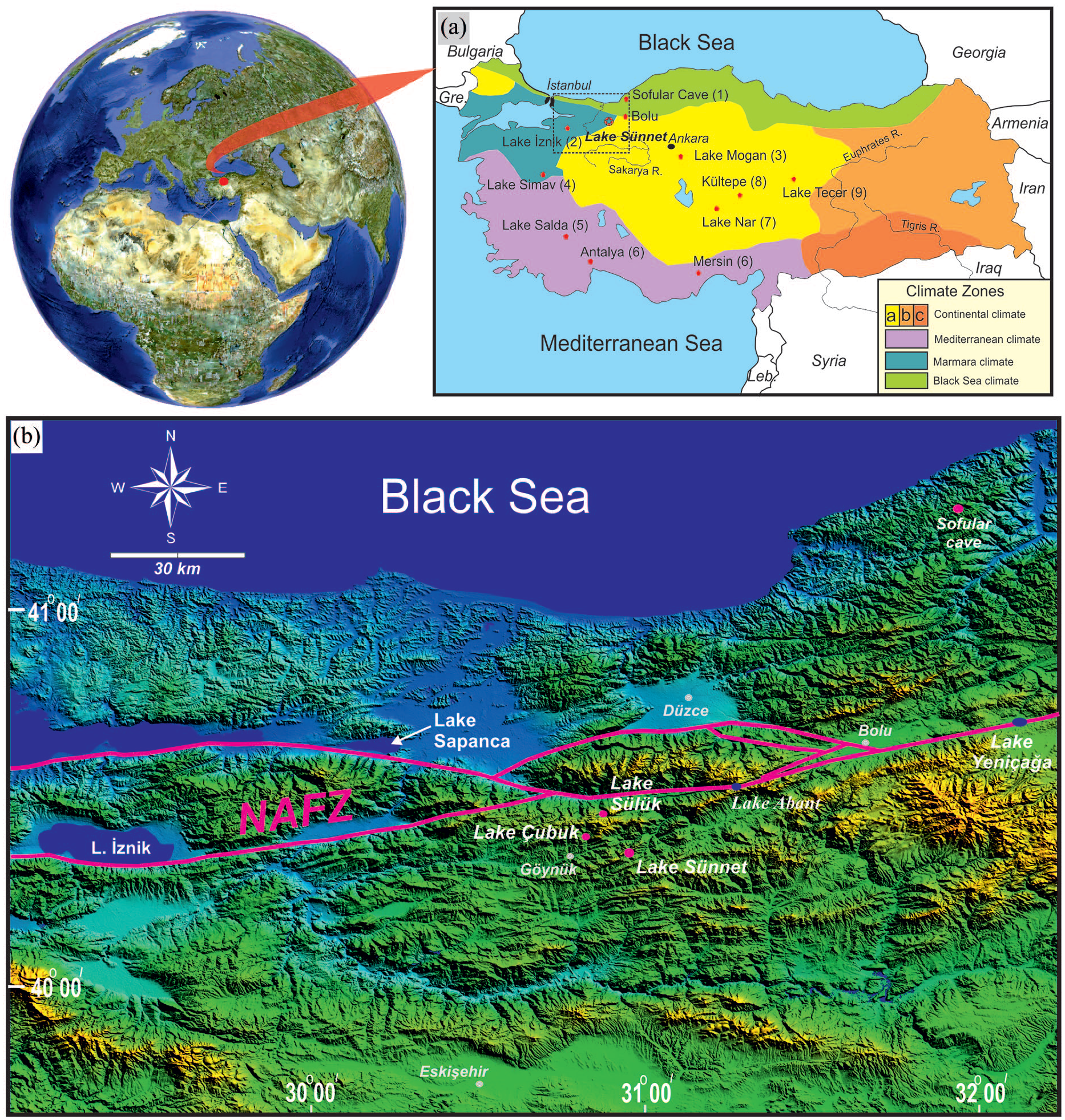
(a) Climate zones in West Anatolia (Atalay and Morton, 1997) (a, b, and c are Central, Eastern, and Southeastern Anatolian sub-zones of continental climate) and location of the Little Ice Age records mentioned in the text: 1 – Sofular Cave (Göktürk et al., 2011), 2 – Lake İznik (Ülgen et al., 2012), 3– Lake Mogan (Oybak-Dönmez et al., 2021), 4– Lake Simav (Ocakoğlu et al., 2019), 5– Lake Salda (Danladi and Akçer-ön, 2018), 6– South Anatolian tree-ring records (Touchan et al., 2007), 7– Lake Nar (Jones et al., 2006), 8– Kültepe (Şenkul et al., 2018), 9– Lake Tecer (Kuzucuoğlu et al., 2011), Gre: Greece, Leb: Lebonan, R: River. (b) Topographic outlines of NW Anatolia including Lake Sünnet and some other records mentioned. NAFZ: North Anatolian Fault Zone.
To contribute to this ultimate objective, Lake Sünnet, a landslide-dammed lake, was selected, where the accumulation rate is extremely high and aridity is pronounced. Based on a previous bathymetric survey (Ocakoğlu et al., 2011), we collected three cores from different parts of the lake and carried out different multi-proxy analyses, ranging from stable isotopes to diatoms, pollen and ostracods. The results were correlated with the tree-ring record of the nearby Lake Sülüklü and the high-resolution Black Sea and Central Anatolian stalagmites and lake records, in order to reveal the environmental and climatic history of the area.
Climate and vegetation
Lake Sünnet (40°25′17″N′, 30°57′24″E) is a high-altitude (1044 m) lake in NW Anatolia and lies at the conjunction of three widespread climate zones: the central Anatolian dry continental climate, the cold and humid Black Sea climate, and the warm and humid Marmara climate (Atalay and Morton, 1997) (Figure 1a). The long-term mean precipitation and evaporation data from the nearby Bolu Meteorological Station (538 and 827 mm respectively) reveal the significance of aridity in the area. The area has a prominent difference in mean temperature between summer and winter (18.7°C and 1.6°C respectively) (Meteoroloji Genel Müdürlüğü, 2020). Accordingly, Lake Sünnet and the vicinity are classified as a semi-humid region according to the De Martonne aridity index (Meteoroloji Genel Müdürlüğü, 2020). The study site is located in the southern part of the montane forest belt of northern Turkey, which belongs to European-Siberian phytogeographic region and is dominated by black pine trees under semi-continental climatic conditions (Quézel and Barbéro, 1985). On the northern exposures of the montane forest belt, which is about 50 km from Lake Sünnet area, fir, Abies nordmanniana and beech, Fagus orientalis occur in large numbers under humid climatic conditions (Atalay, 1983). The location of Lake Sünnet is also close to the steppe area of central Anatolia in the south which is characterized by a diverse herb vegetation cover and relatively dry climate (Quézel and Barbéro, 1985). Due to significant aridity, Pinus nigra sp. pallasiana (black pine) is prevalent in the area. Lake Sünnet also shelters a rich aquatic flora that is represented mainly by Alisma lanceolata (water plantain) and pondweed species such as Potamogeton berchtoldii, P. gramineus, and P. luceus. These are accompanied by Eleocharis palustris, Epilobium hirsutum, Galium rivale, Lycopus europaeus, and several other species that prefer shoreline and very shallow lakes (Seçmen and Leblebici, 1997). The modern extent of meadow and agricultural areas accounts for 15% of Lake Sünnet’s watershed. Dry-farming of mainly barley and wheat is the only agricultural activity at higher altitudes, while arboriculture (mainly with apple, peach, and pear) and vegetable production (with potato, lettuce, and cabbage) are common in valley floors. A previous study by Ocakoğlu et al. (2011) demonstrated that the modern lake bottom sediments are dominantly composed of Pinus nigra pollen up to 87%.
Bathymetry of Lake Sünnet
Lake Sünnet is located in a narrow river valley that is deeply incised into Mesozoic limestones, 15 km to the east of the Göynük Town (Bolu) (Figures 1b and 2). Earlier studies demonstrated that the lake was formed by two landslides that were triggered most probably by a mega-earthquake at about 8 kyr ago (Abdüsselamoğlu, 1959; Hoşgören and Ekinci, 2004; Ocakoğlu and Tuncay, 2019; Ocakoğlu et al., 2011). Two creeks (Gölbaşı and Karaköy creeks) in the south flow into the lake (Figure 2). The lake sediments belonging to the more humid Middle Holocene period are preserved as terraces at both sides of the creeks. Two prominent deltas are developed at the subaqueous prolongation of the creeks (Figure 2a). The frontal part of both deltas is shaped by complex channel morphology. The delta front of the better developed Karaköy Delta is defined by a 10 m-wide channel that becomes narrow and indistinct toward the delta top. The valley margins beneath the lake, far from the deltas are as steep as 60°, however, the slope of the landslide dam under water is less steep. The previous detailed bathymetric investigation revealed many circular pockmarks of unknown origin as large as 15 m wide and 10 m deep (Figure 2b). Lake Sünnet has a surface area (AL) of 0.19 km2 and receives surface water from a watershed (AW) of 12,567 km2 (AL/AW = 0.015). Previous water budget estimations show a very short (only several years) residence time for the lake (Erayık, 2011; Ocakoğlu et al., 2011).
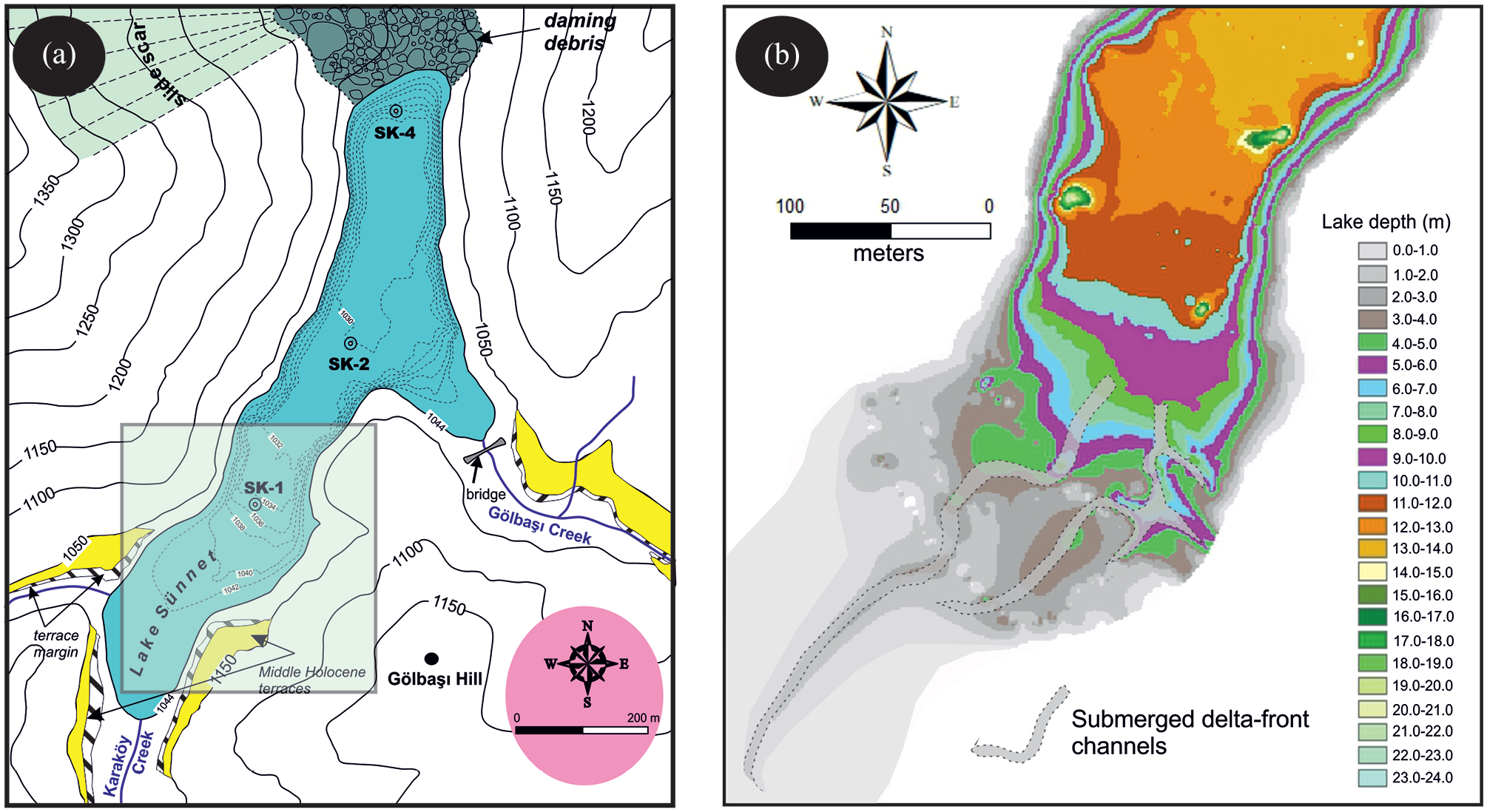
(a) Morphological outlines of the study area. SK-1 to SK-4 mark the location of the studied cores. (b) Complex delta-front morphology of the southern delta (rectangle in (a)) (redrawn from Ocakoğlu et al. (2011).
Modern hydrology and ecology
According to monitoring research conducted between 2005 and 2007, the lake level changed as much as 6 m between dry and wet seasons (Balcı, 2008). Accordingly, the pH of the lake water varies between 7.2 and 8.2, defined by sub-saline (0.20‰–0.29‰) and slightly alkaline conditions throughout the year. The same study shows very little dissolved oxygen (<2 mg/L) below 6 m water depth. Because the salinity changes between wet and dry seasons are very small, the main reason for the extreme lake level changes has been attributed to leakage from the landslide dam rather than evaporation (Ocakoğlu et al., 2016b).
There is a limited richness of living ostracod species in Lake Sünnet due to its mesotrophic-eutrophic nature compared to the average Turkish lakes (Külköylüoğlu et al., 2010). Among the nine living and five sub-fossil ostracod species observed, Limnocythere inopinato is the most abundant in both dry and wet seasons while Ilyocypris bradyi is encountered only in wet seasons. The rest of the species are specific to only dry months. Diatom species on the lake bottom dominantly consist of the planktonic species Cyclotella ocellata and the benthonic Gyrosigma acuminatum in a lesser amount, both of which indicate oligohaline to alkaline conditions (Ocakoğlu et al., 2011).
Methodology
The coring operations in Lake Sünnet were carried out using a Livingstone corer on a 4 m × 4 m size modular plastic platform in 2009. The retrieved cores were halved, logged, and sampled for multi-proxy analysis in a lake-side laboratory. The other half of the core was preserved in cool conditions at +4°C for potential future analysis in the Sedimentology Laboratory at Eskişehir Osmangazi University. Stable isotopes, Total Organic Carbon (TOC), Total Carbonate (TCO3), and Magnetic Susceptibility (MS) analyses were done at 2 cm intervals in core SK-2 while pollen, diatom, and ostracod analysis were carried out in two cores (SK-2 and SK-4) at 4 cm intervals. Due to similar results, only the data from core SK-4 are given in the text, but data from core SK-2 are also fully available in Ocakoğlu et al. (2011). Pollen analyses were obtained on two separate slides for each sample. The main steps for pollen slide preparation include subsampling of 1 cm3 of sediment, removal of calcium carbonate and humic acids by using 10% HCl and 5% KOH, respectively, in a heated water bath at 100°C for 30 min, sieving, treatment with 6 mL of 40% HF in water bath at 100°C for 15 min for removing silica, staining with basic fuchsin, and mounting in glycerine gel. A soldering iron was used to move pollen grains on slides for critical identification. Reference collections in the Palynology Laboratory at Hacettepe University (Turkey) and some milestone references (Faegri and Iversen, 1989; Moore et al., 1991) were used to identify pollen. The calculation sum is based on total land pollen (TLP) excluding aquatics, spores, and unknown or unidentifiable pollen, with a minimum sum of 509 grains. As Pinus pollen is over-represented in all samples, which is over 80%, it is excluded from the sum in zonation to enable a better assessment of the past vegetation. Tilia software (Grimm, 2005) is used for the zonation by cluster analysis (CONISS). A square root transformation was applied before the zonation.
Diatom samples were boiled in acid, as explained in greater detail elsewhere (Round et al., 1990). Krammer (2000) and Lange-Bertalot (2001) were considered in the determination of diatom species in the studied samples. In each slide, a total of 300 diatom frustules are identified and counted. Forty grams of each sample were processed for ostracod studies. The samples were reacted with dilute (5%–10%) and hot (70°) hydrogen peroxide for about 12 h (and 24 h if necessary). After that, the residuals were left to dry naturally. The dried materials were sieved using a set consisting of 1000, 500, and 250 μm mesh. The valves were picked under an Olympus SZ60 binocular microscope (see Tuncer, 2020 for the taxonomic studies and more detailed information). All materials are stored in the Paleontology Laboratory at Hacettepe University. Total organic carbon (TOC) and total carbonate (TCO3) measurements were done by loss on ignition tests. The powdered and dried samples were first heated at 550°C for 5 h for their organic matter, and a further 2 h at 950°C for their CO3 content (Santisteban et al., 2004). The equations of TCO3 = 1.36 * LOI950 and TOC = LOI550/2 by Heiri et al. (2001) were applied for the transformation of weight loss to TOC and TCO3 respectively. Stable isotopes of O and C were measured from bulk mud at the Iso-Analytical Laboratory (UK) with precisions of 0.2 and 0.1 per mil respectively. Magnetic susceptibility measurements were carried out on powdered and dried samples by using a Bartington MS2B sensor on a mass-specific base. Lastly, the radiocarbon dates were analyzed from charcoal except for one sample at Georgia University (USA). The radiocarbon ages were calibrated by CalibRev8.1 software using the Intcal20 database (Stuiver et al., 2022).
Description and correlation of the cores
Three cores were studied to shed light upon the environmental and paleoclimatological changes in Lake Sünnet. Core SK-1 was collected from 10 m water depth at the delta front while core SK-2 was retrieved from 13 m water depth at the lake center (Figure 3). The last core (SK-4) stands at 10 m deep at the toe of the gently sloping landslide dam in the north (Figures 2a and 3). From the delta top to the deeper part of the lake, an obvious decrease in grain size, and a color shift from dark brown to gray/olive green occurs (Figure 3). Core SK-1 is made of the alternation of thicker (<7 cm) dark brown coarse sand with thinner (2–3 cm) lighter brown silt and mud. An almost sand-free interval between 180 and 150 cm is distinct in this core. In core SK-2, light brown and gray-green muds alternate at 3–5 cm intervals. Two thicker dark brown mud intervals occur at 80–70 cm and below 160 cm. Some intervals in the core frequently comprise pine needles and charcoal fragments. At the base of the SK-4 core (164 cm) horizontal reed swaths and overlying small-sized mollusk shells occur. Upsection, the alternation of gray and bluish muds with occasional algal carbonate tubes occur between 120 and 60 cm. The core is predominantly composed of the alternation of black and brown mud in the upper 40 cm.
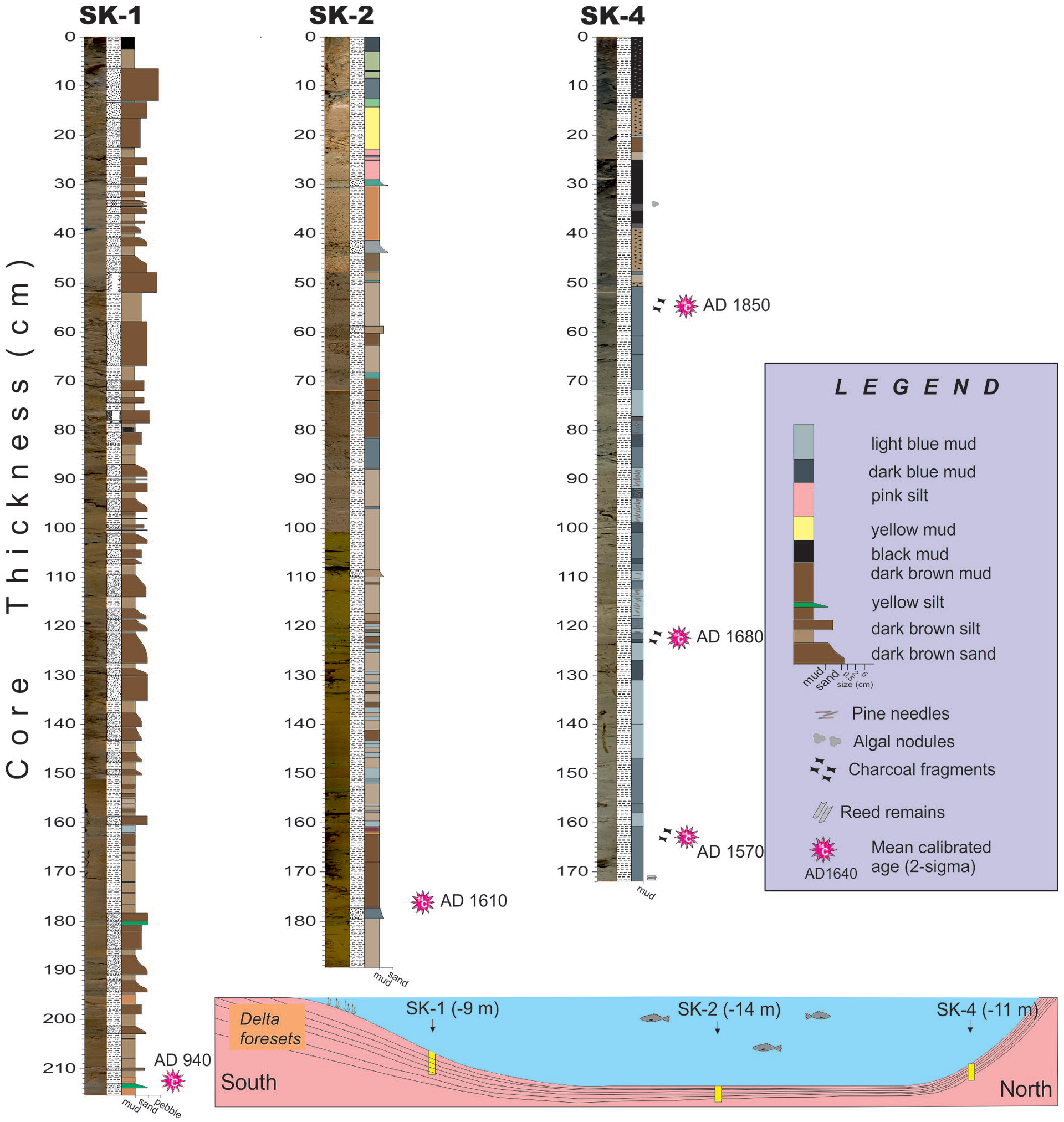
Lithology and correlation of the three cores from Lake Sünnet. The inbox shows the bottom morphology and location of the cores across a longitudinal profile.
Chronological frame
Five radiocarbon samples were collected to determine the chronological frame of the studied cores. The radiocarbon data and the calibrated ages are shown in Table 1. The age from bulk mud at the bottom of the delta front sands in core SK-1 dates back to AD 940. Due to the potential older carbon transported from the watershed, this age is suggested to be biased to an older age. The mean calibrated age of a plant fragment close to the bottom of core SK-2 is AD 1611. Core SK-4 is dated by three radiocarbon samples from charcoals and is modeled by using Bchron R software (Haslett and Parnell, 2008; Parnell et al., 2008; Figure 4). The model output shows more than 100 years of uncertainty in a 95% confidence interval throughout the record. Following the mean two-sigma calibrated ages, the base of the core sees AD 1545, and an average accumulation rate of 3.4 mm/year. As a result, the average time resolution of the proxies applied in this study varies between 5 and 10 years.
Radiocarbon data for the samples from Lake Sünnet cores.

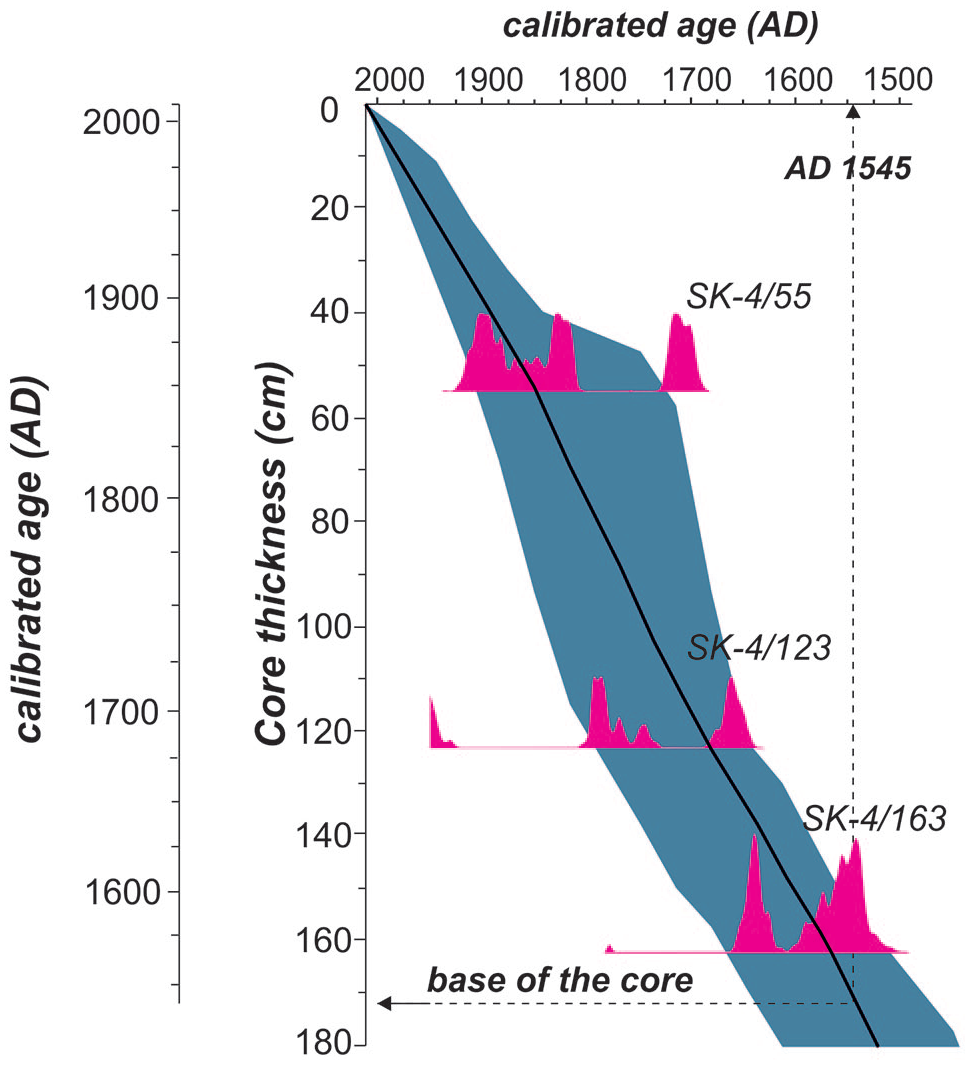
Age-depth model with two-sigma probability curves of core SK-4.
Multi-proxy results
Pollen record in core SK-4
This pollen record comprises a total of 64 palynomorph types from the 42 samples including 14 arboreal plants, 35 herbaceous plants, 8 aquatics, and 7 ferns. The arboreal taxa are mainly represented by Pinus (average 90.3%), Fagus (average 21.3%), Quercus (average 14.5%), Abies (average 6.17%), Carpinus (average 5.4%), and Juniperus (average 6.6%) (Figure 5). Other woody plants, such as Alnus, Betula, and Ulmus also occur, albeit in relatively low frequency. The biotic diversity of herb pollen in the assemblage is high. The most abundant herbs in decreasing amounts are Poaceae, Plantago coronopus type, and Ranunculaceae (Figure 5). The main aquatic taxa is Myriophyllum spicatium making an extraordinary peak of 41% at 118 cm. Cluster analysis and changes in pollen percentages allow us to identify four zones and several sub-zones in the pollen assemblage of core SK-4 (Figures 5 and 6).
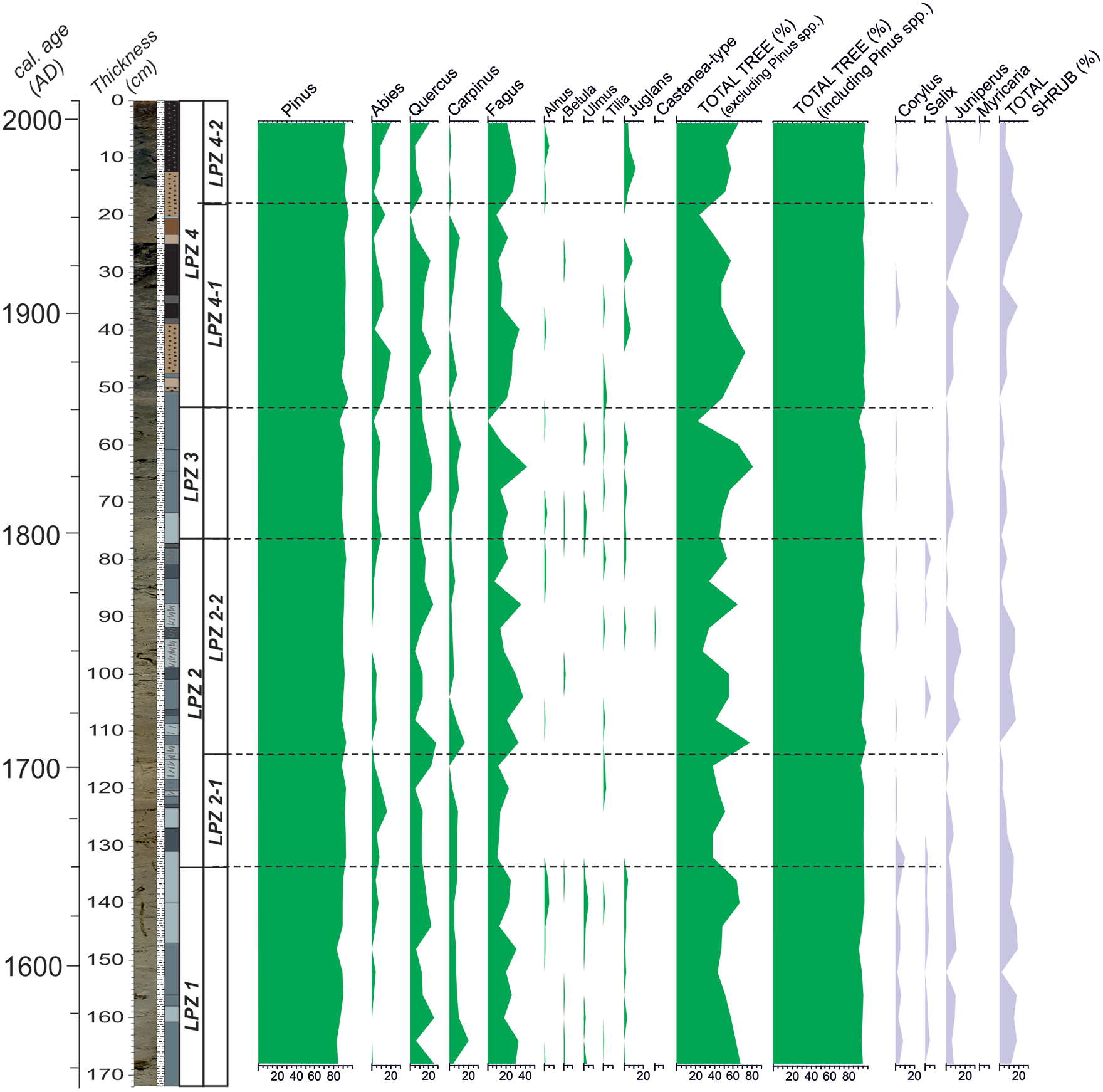
Tree and shrub pollen of core SK-4. The X-axis is the percent abundance.

Herb and aquatic pollen of core SK-4. The X-axis is the percent abundance.
LPZ1 (168–133 cm)
This zone is represented by a high abundance of broad-leaved taxa including Fagus (12.2%–32.9%), Quercus (6.8%–26.2%), and Carpinus (3.7%–10.3%), followed by a moderate quantity of Juniperus (0%–10.9%) and Corylus (1.7%–7.9%) (Figure 5). The amount of Abies goes up from the bottom to the end of this zone. Herbaceous taxa are characterized by the predominance of Poaceae (1.8%–13.2%) and Plantago coronopus type (0%–6.9%). The pollen of Scrophulariaceae peaks (10.5%) at 152 cm. The percentage of aquatic species Myriophyllum spicatum ranges from 0% to 9.5%.
LPZ 2-1 (133–114 cm)
This is marked by an increase in arboreal taxon Abies (3.2%–15.6%), non-arboreal taxon Poaceae (5.0%–22.6%), and Plantago coronopus type (2.5%–13.4%) (Figure 5). The wetland taxa Myriophyllum spicatum (7.6%–45.2%) and Myriophyllum verticillatum (0%–14.5%) simultaneously gradually increase while a decline occurs in the abundance of drought-tolerant Juniperus (0%–7.6%) (Figure 6). The amount of arboreal taxa, such as Quercus, Carpinus, and Fagus, is still comparable to LPZ1. The pollen of Alnus, Betula, Ulmus, and Juglans firstly appear, though rare, in this zone. Saussurea type and Asteraceae Liguliflorae attain their maximum values of 15.3% and 8.0%, respectively. This also applies to the high abundance of ruderal taxon Centaurea (1.5%–5.0%). Herbaceous taxa Succisa, Ericaceae, Fabaceae, Trifolium montanum type and Stachys type appear in this zone, albeit in small amount.
LPZ 2-2 (114–76 cm)
This is characterized by high quantities of Quercus (5.1%–27.7%) and Fagus (8%–3%). Juniperus pollen is abundant (up to 15.5%) at the beginning but quickly drops in the rest of the zone (Figure 5). The Carpinus curve displays a similar trend. Poaceae (0%–11.8%) declines compared to the previous zone. A sudden drop in the amounts of Myriophyllum spicatum (no more than 7.9%) is also apparent. Centaurea shows a rising trend, reaching a maximum value of 10.3% at 88 cm in the middle of the zone, and keeps steady until the top of the zone. The Ranunculaceae abundance (0%–11.6%) is the highest in the lower part, diminishes upward and peaks at the top of this zone as Artemisia (5.3% at 80 cm) starts to appear simultaneously (Figure 6).
LPZ 3 (76–53 cm)
This zone is characterized by the co-dominance of trees (mainly Quercus and Fagus) and herbs (mainly Poaceae, Artemisia, and Plantago coronopus). The palyno-flora is very diverse and contains many rare taxa belonging to trees and herbs. The most significant change across the LPZ 2-2/3 boundary is a remarkable rise in Abies, Poaceae, and Artemisia. The curves of Quercus and Poaceae make two peaks at 64 and 56 cm. The amounts of wetland taxon Myriophyllum spicatum along the zone are much reduced compared to the previous zone (Figure 6).
LPZ 4-1 (53–18 cm)
The dominant pollen of the zone are Abies (2.4%–16.0%), Quercus (0%–22.1%), Fagus (9.4%–34.1%), and Juniperus (0%–17.3%). Other trees such as Carpinus (0%–8.0%) and Juglans (0%–9.5%) are of secondary significance and are accompanied by minor Alnus (0%–2.4%), Betula (0%–2.3%), and Tilia (0%–4.0%) amounts. Herbs are still diverse and are represented by the constant Poaceae (4.0%–15.0%) and increasing amounts of Plantago coronopus type (0%–9.4%) and Scrophulariaceae (0%–7.2%). Cerealia-type increases, yet still fluctuates. Polygonum aviculare type (9.4% at 20 cm) climaxes at the end of this subzone. Spores are also diverse and are characterized by minor quantities of Botrychium, Lycopodium, and Ophioglossum (Figure 6).
LPZ 4-2 (18–0 cm)
Frequent occurrence of Fagus, represented by the amount between 20.5% and 30.5%, characterizes this sub-zone (Figure 5). Abies and Quercus are proportionally stable, whereas Juglans peak (12.1%) at 12 cm, along with a decrease in Carpinus (0%–2.2%). Juniperus shows an overall decrease from 11.3% to 2.1% along the sub-zone. Alnus is scarcely represented (max. 5.3%).
Diatom record in the core SK-4
The most abundant and continuously represented diatom species and the abundance of planktonic and benthonic diatom species are shown in Figure 7. The most significant aspect of this record is the sparse occurrence of diatoms (generally benthonic) below 90 cm. Nevertheless, they occur in every level above 90 cm. Moreover, the planktonic diatom abundance gradually rises to 80% at the top of the core. Between 173 and 90 cm, only five samples contain diatoms. Three of them are represented by benthonic species of Hantzschia amphioxys in minor abundance. In the other three samples, several planktonic species (Aulacoseira granulata and Cyclotella meneghiniana) are represented by a limited number of individuals (Figure 7). At 90 cm, the abundance of benthonic diatoms is the highest of the whole record. Up in the core, diatom species Eunotia arcus, Gomphonema intricatum, and Gyrosigma acuminatum occur at almost all stratigraphic levels. Cyclotella bodanica, a species that prefers oligotrophic clean water (Cox, 1996), forms two peaks at 70 and 30 cm (Figure 7). It also occurs at the top of the core in high abundance. We also studied the cores SK-1 and SK-2 for their diatom content (Ocakoğlu et al., 2011), but these data are not included herein due to the similarity of the diatom flora and the space limitations. In core SK-1, samples below 176 cm are either barren or comprise only of Hantzschia amphioxys. Similarly, in core SK-2 there is no diatom preserved below 160 cm (i.e. AD 1500–1600), while only several intervals between 160 and 120 cm exhibit limited benthonic diatom species. Similar to core SK-4, the continuous diatom occurrence is observed above 120 cm. These correlative discontinuous diatom records in the three cores may have significant implications regarding the past lake-level changes.
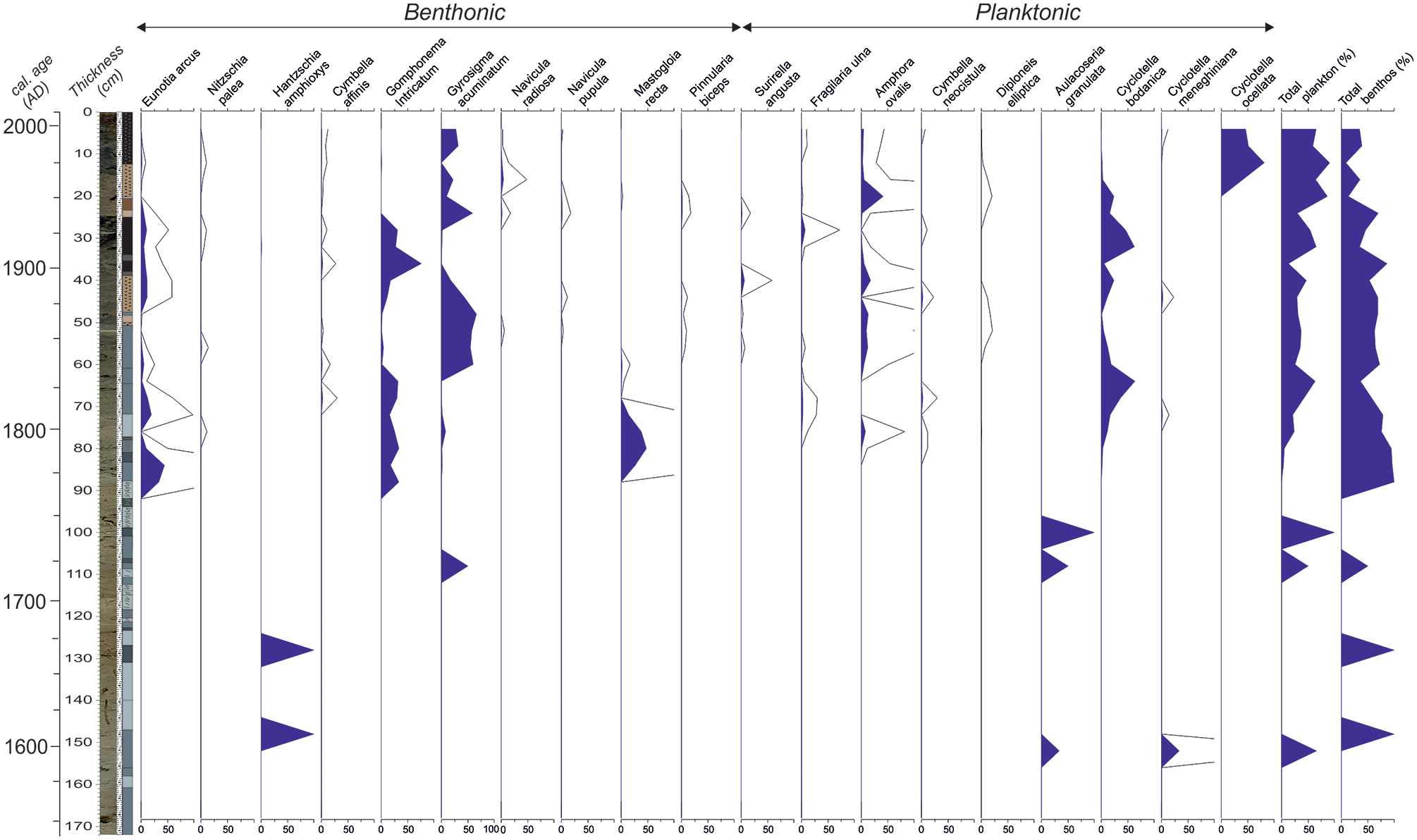
The major diatom taxa in core SK-4. The X-axis is the percent abundance.
Ostracod fauna assemblage
Thirteen ostracod taxa (Cypridopsis vidua, Potamocypris cf. fulva, Prionocypris zenkeri, Psychrodromus olivaceus, Candona candida, Neglecandona neglecta, Pseudocandona sp., Cyclocypris ovum, Cypria sp., Ilyocypris bradyi, Ilyocypris hartmanni, Ilyocypris inermis, and Limnocythere inopinata) belonging to 11 genera were determined (Figure 8). Grossly saying, the abundance of ostracods is low (<10 valves) below 60 cm and very high (up to 70 valves) albeit with some fluctuations above 60 cm (Figure 8). The main contributor to this higher abundance is Neglecandona neglecta. Ilyocypris bradyi and Cyclocypris ovum are the only species continuously represented along with the core though in limited abundance (<5 valves). The summary of ostracod fauna and their environmental demands in informally divided core segments are given below.
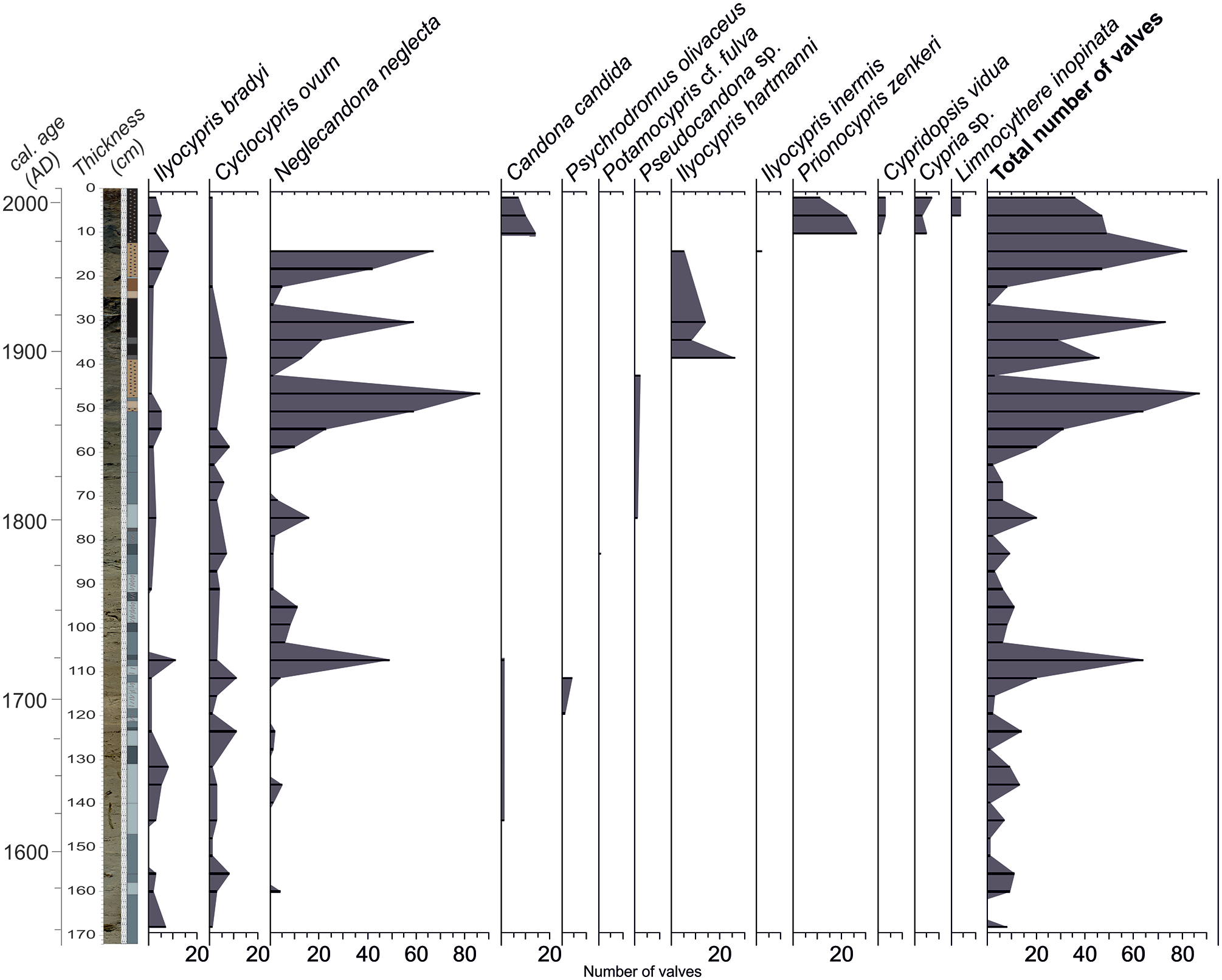
Vertical distribution and abundance of the ostracod taxa in core SK-4.
168–124 cm
The abundance and diversity of ostracods are low (Figure 8). Here, the widespread and highly tolerant species C. ovum and N. neglecta (found even in drought conditions), which can also tolerate the low oxygen and freshwater-mesohaline (up to 6.4‰) conditions (Meisch, 2000), are present. Especially, C. ovum, associated with Charophytes, indicates a very shallow and phytal water body (Meisch, 2000). Despite its low abundance, the presence of rheophilic I. bradyi may indicate surface water recharge pulses in the lake basin. The oligothermophilic N. neglecta along with I. bradyi may point out the prevalence of the relatively cooler water conditions. Owing to the highly tolerant species, the possibility of temporary desiccation cannot be ignored in this interval. The fact that many of the samples (164, 152, 148, 140, and 128 cm) are barren (0 and/or 1 valve) in terms of ostracods, and the abraded surface of some of the ostracods available from this interval supports this interpretation (Figure 8).
120–108 cm
The lower parts of the interval comprise P. olivaceus, a spring-dweller species (Meisch, 2000) in low abundance (Figure 8). However, the number of total ostracod valves dramatically increases in the upper part (Figure 8). Adult and juvenile valves of N. neglecta and I. bradyi are generally observed together. Freshwater mollusks (Pisidium-type pelecypods and planispiral gastropods) also increase at this interval.
104–64 cm
The abundance of ostracods relatively diminishes at this interval (Figure 8), only widespread and highly tolerant species are present. Ostracods along with mollusks and charophytes that keep represented at that interval may indicate phytal, very shallow, and freshwater-oligohaline conditions.
60–16 cm
The population of ostracods begins to increase at the base of this interval and reaches the maximum at 48 cm of the core where adults and juveniles of N. neglecta are dominant (Figure 8). Afterward, the population of ostracods dramatically diminishes at 44 cm (Figure 8). At the following level (40 cm), I. hartmanni becomes the dominant species with its first occurrence along with the core. The ecology of I. hartmanni is not well-known. Külköylüoğlu et al. (2018) reported this species from creeks, troughs, and streams. It might demonstrate very shallow and relatively warmer water bodies.
12–0 cm
The ostracod fauna is replaced by a different and relatively diverse assemblage (Figure 8). The first occurrences of P. zenkeri, C. vidua, Cypria sp., L. inopinata, and abundance of C. candida characterize this interval and separate it from the lower parts. The dominant species P. zenkeri is an oligothermophilic species and mainly lives in slow-flowing streams (Meisch, 2000). P. zenkeri and accompanying taxa show that the water recharge to the basin became effective and very shallow (littoral), permanent, phytal and freshwater-oligohaline lacustrine conditions revived temporarily. In the uppermost part of the core, nodes on the L. inopinata (polithermophilic) valves are frequent that may mark increasing alkalinity (McCormack et al., 2019). Furthermore, the ostracods in this interval (P. zenkeri, C. vidua, L. inopinata, C. candida, and I. bradyi) were recorded alive in Lake Sünnet between May 2005 and September 2007 (Külköylüoğlu et al., 2010).
Magnetic susceptibility (MS), TOC, and TCO3
MS and TCO3 curves display contrasting trends (Figure 9). From the base up to 70 cm, the variations in the MS curve are minimal and gradual. Following a peak at 50 cm, a significant decrease occurs at 40 cm. The gradual increase up to 20 cm is followed by relatively reduced MS values in the uppermost 10 cm. The TCO3 curve shows relatively higher values (43%–50%) from the bottom to 95 cm. A gradual decrease until 45 cm is interrupted at 40 cm, but later it increases to 20 cm and reaches the minimum (13%) of the whole record, followed by a quick recovery. The TOC curve displays the minimum value (1.6%) between 160 and 130 cm. This period is also characterized by more positive δ18O and more negative δ13C values. A similar low-TOC interval occurs at 115–80 cm. Following the highest value of the whole record (4%), TOC gradually decreases until 40 cm. After the last peak between 40 and 30 cm, the TOC curve gradually decreases until the top of the core.
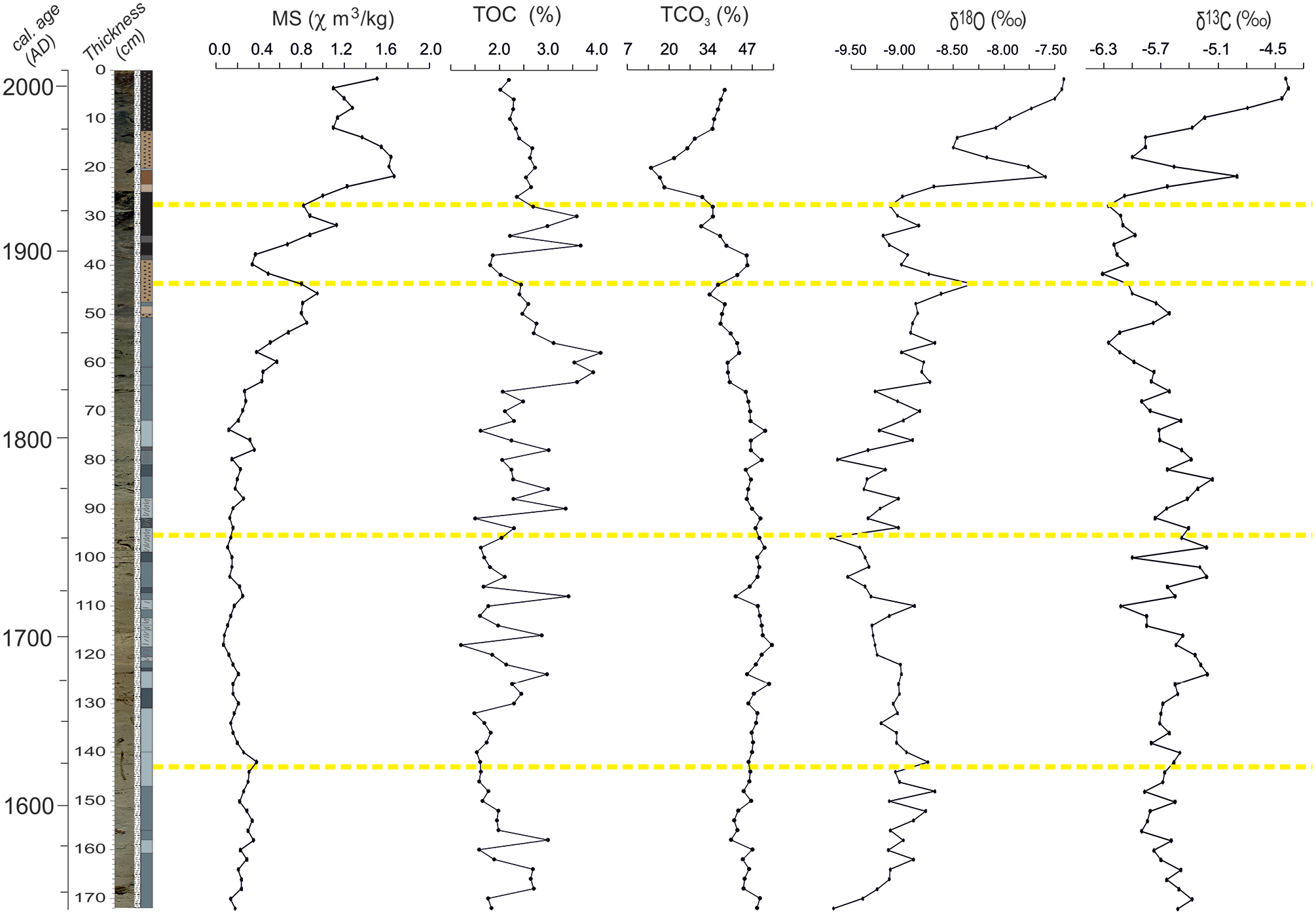
Physico-chemical proxies measured in core SK-4. Yellow guidelines are the boundaries of trends seen in some of the proxies mentioned in the text.
δ18O and δ13C
The lowest (−10.3‰) and highest (−7.5‰) δ18O values are seen at the base and the top of the core respectively (Figure 9). Three centennial cycles are superimposed on higher frequency cycles with magnitudes of 0.25‰. The age-depth model implies that the latter might correspond to 11 years-long sunspot cycles (Garric and Huber, 2003). These cycles are not observed in the uppermost 30 cm. The highest value in the larger cycle occurs at 145 and 45 cm. The uppermost cycle starts at about 30 cm with a value of −9.25‰. After a secondary peak at 25 cm, the curve gradually shifts to more positive values in modern times. The δ13C curve fluctuates between −6.3‰ and −4.5‰ and is inversely correlated with the δ18O curve below 45 cm. A similar trend is seen upsection. In this curve, the high-frequency fluctuations with magnitudes of 0.3‰ are noticeable below 60 cm, but quickly disappear upward (Figure 9).
Interpretation of the proxy data
Before AD 1750
Diatoms provide important insights in terms of past limnological conditions owing to both environmental demands of individual species and the ratio of planktonic to benthonic species (Battarbee et al., 2001; Laird and Cumming, 2008; Wolin and Duthie, 1999; Wolin and Stone, 2010). In this regard, the absence of diatoms (only Hantzschia amphioxys, Auoceria granulate, and Cyclotella meneghiniana) or their limited diversity below 90 cm in core SK-4, below 176 cm in core SK-1, and below 160 cm in core SK-2 is a result of unfavorable conditions for diatom preservation (excessive salinity and very shallow water) (Flower and Ryves, 2009; Reed, 1998). Cyclotella meneghiniana is an indicator of higher salinity (Juggins, 2011). Reed et al. (2012) similarly relate this species to higher salinity (3.0–3.5 g/L) in Anatolian lakes. Hantzschia amphioxys, is a cosmopolitan species of different environments, and one of the most common soil diatoms in the world (Foets et al., 2020). On the other hand, a frequent yet limited number of ostracods and well-preserved pollen in this diatom-barren interval, invalidates the complete desiccation of the lake for extended periods. These data imply that the lake was generally very shallow at the core location which presently is 3 m above the deepest part of the lake (Figure 3).
Other proxies provide some insight into the change of temperature and/or precipitation in this period. For instance, the pollen record displays three zones/subzones that are characterized by the fluctuations in the pollen of broad-leaved trees (Quercus, Carpinus, Fagus, and Corylus) and herbs between 180 and 168 cm. LPZ 1 with higher broad-leaved tree pollen and less herbaceous pollen represents a relatively warm and humid climate (Figures 5 and 6). The δ18O of the lake muds, which is a measure of the precipitation/evaporation balance in lake sediments (Jones and Neil Roberts, 2008; Jones et al., 2006), displays a positive shift due to increased summer evaporation during a warmer climate. Although one can suggest significant isotopic bias due to the potential detrital carbonate input, the widespread extent of siliciclastics in the lake watershed implies the contrary. Furthermore, a comparative isotope study of the Middle Holocene terraces of Lake Sünnet demonstrate a strong isotopic covariance between bulk mud and selected mollusk shells, confirming the predominantly autogenic origin of carbonates (Ocakoğlu and Kapan-Yeşilyurt, 2014). This interval is also typical of a less diverse and limited ostracod association. Among them, C. ovum prefers very shallow phytal environments. Furthermore barren levels and abraided ostracod valve suggest repeated exposure. As a result, we interpret relatively higher lake levels and increased evaporation under warm and humid conditions.
The subsequent pollen zone (LPZ 2-1) is formed of higher herbaceous and Abies pollen and less broad-leaved tree pollen, which indicates a relatively cold and dry climate. Dry conditions are reflected in the ongoing more positive δ18O values at this interval. The culmination of aquatic pollen toward the top of this zone strongly implies the prevalence of very shallow water conditions. It seems that the lower temperature helps preserve of organic matter. The occurrence of P. olivaceus, a spring-dweller ostracod species, at the top of this interval implies springs around the shallow lake. LPZ 2-2 represents a weak swing to relatively warm and humid conditions where broad-leaved tree pollen increases and herb pollen decreases. In this interval, the negative shift at 95 cm in the δ18O graph and the following recovery represent cold and subsequently warmer conditions.
Between AD 1750 and 1850
Diatoms from core SK-4 consistently appear after 90 cm (i.e. AD 1750). In addition, the planktonic/benthonic (P/B) diatom ratio, as a paleo-bathymetric indicator, shows that the lake level started to increase in a fluctuating manner after AD 1750. After this time, Cyclotella ocellata and Cyclotella bodanica species are the most abundant. Having planktonic nature, these species are often seen in higher abundance in deep lakes. Cyclotella ocellata is widely found in the littoral zone of lakes and running water environments (Czarnecki and Blinn, 1978; Krammer and Lange-Bertalot, 1991). According to Reed et al. (2012), the species generally characterizes freshwater conditions (salinity between 2.6 and 2.9 g/L) in Anatolian lakes while C. bodanica is rather oligotrophic and common in clean waters (Cox, 1996). Foged (1981) indicated that the species is oligohaline and can be found in neutral pH conditions. Therefore, the interval between 90 and 0 cm in core SK-4 represents clean and oligotrophic freshwater conditions with slight alkaline properties. The lower part (90–50 cm) of this interval comprises two pollen zones based on the subtle changes in the amount of broad-leaved and Abies pollen. The upper LPZ 3 zone includes higher Abies, Poaceae, and Artemisia, altogether demonstrating a relatively cold and dry climate. This interval includes a small number of ostracod valves with high environmental tolerance. The frequent occurrence of mollusks and charophytes in the same interval strongly indicates shallow, phytal, fresh-to-oligohaline aquatic conditions.
After AD 1850
Above 50 cm (AD 1850), remarkable changes occur in the appearance of sediments and in proxies. The color of sediments shifts from gray to brown-black alternations. Total herb pollen apparently decreases while Cerealia-type pollen displays a remarkably unsteady increase (LPZ 4-1). The contemporaneous increase in the pollen of Quercus, Fagus, and Juglans implies a warmer and humid climate. In this and subsequent pollen zones, the total aquatic pollen make two weak peaks that closely match with sediment color change and positive shifts in δ18O at the AD 1890s and 1950s. In these relatively dry periods framed in the modern warm/humid episode, the number of ostracod valves also increases. After the AD 1960s (uppermost 20 cm of the core), the δ18O curve shifts to the most positive values of the whole record as a result of increasing drought. This drought is synchronous with a slight shoaling (increase in benthonic diatoms), replacement of very different and diverse ostracod assemblages, and a remarkable peak of Cerealia-type pollen.
Discussion
The number of studies on the nature of the LIA and its environmental/societal impact in Anatolia is gradually increasing, although high-resolution multi-proxy records are still scarce. Among others, the isotope and pollen records of Lake Nar (England et al., 2008; Jones et al., 2006) and isotope record from Sofular Cave (Göktürk et al., 2011), pollen and diatom records of Lake Çubuk (Ocakoğlu et al., 2016a), isotope and geochemical records from Lake Salda (Danladi and Akçer-ön, 2018), tree-ring record from south Anatolia (Touchan et al., 2007), and pollen record from Lake Mogan (Oybak-Dönmez et al., 2021) are all noticeable (Figure 1a). In this regard, the Lake Sünnet record in NW Anatolia is distinguished for the multi-proxy approach applied therein and its higher resolution owing to the high sedimentation rate in a landslide-dammed setting. These positive aspects allow obtaining a more complete picture of the LIA when integrated with the previous studies in NW Anatolia.
Lake Sünnet exhibits unusual bimodal diatom preservation. The period between AD 1545–1750 is mainly characterized by the absence of diatoms except for distinct horizons; while, after AD 1750, the diatom record becomes continuous. We attribute the scarcity or absence of diatoms to very shallow and alkaline lake conditions. This barren interval is associated with overall higher δ18O and TCO3 and lower TOC values, and a distinct ostracod fauna that collectively confirms drier conditions. The lowstand fan morphology in relation to the modern deep lake bottom supports very shallow lake conditions at this time (Figure 2b). However, especially the pollen record enables the distinction of a relatively warm and humid period before AD 1630 and a cooler yet dry period between AD 1630–1710. This latter interval comprises the Maunder Minimum with short time shifts before and after (Figure 10). The climax of this global event happens in the 1670s and was essentially characterized by the extremely low solar activity driven mainly by volcanic explosions (Owens et al., 2017).
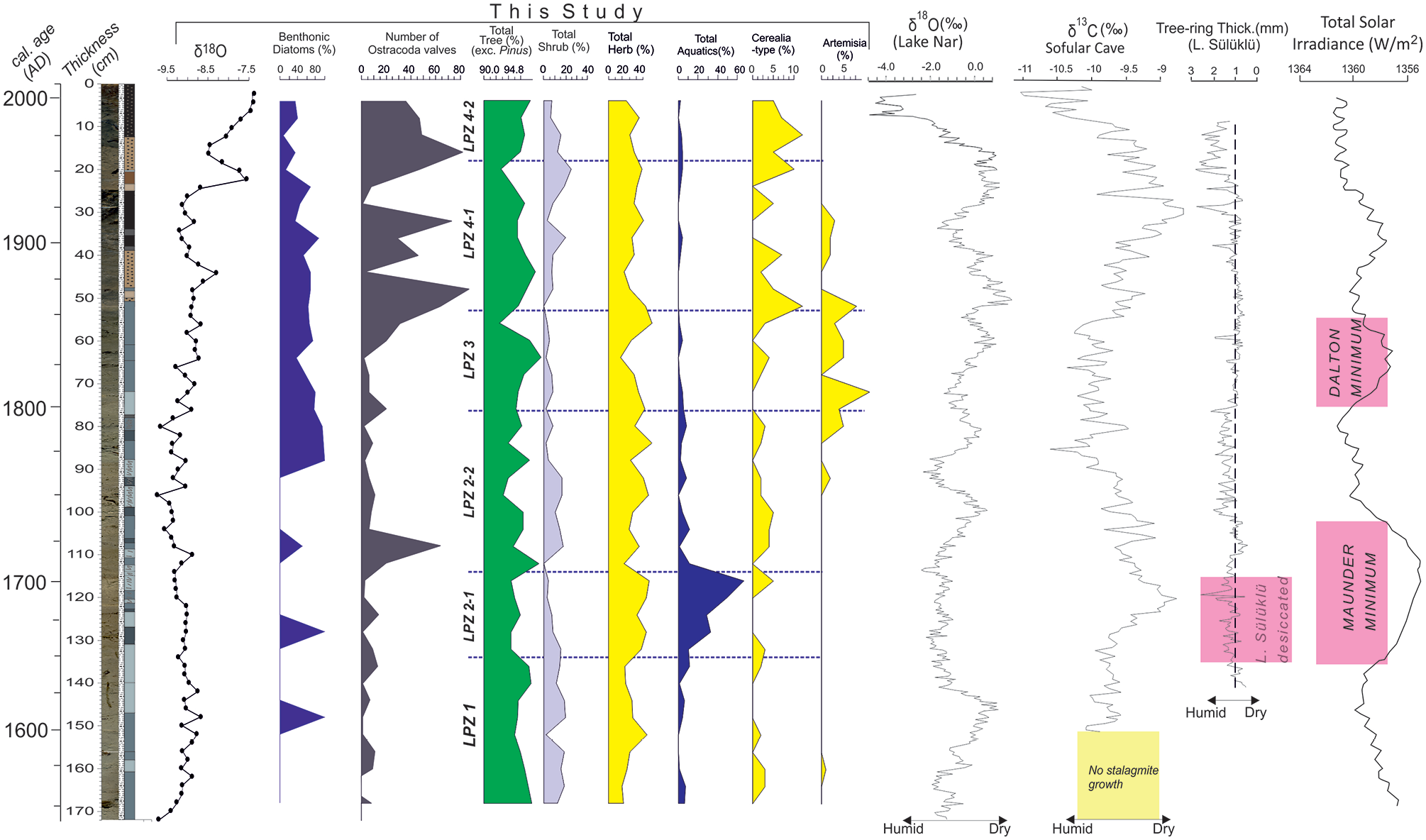
Selected proxies from core SK-4. Tree-ring data from Lake Sülüklü (redrawn after Aytuğ and Kılıç, 1986), δ18O record in Lake Nar (Jones et al., 2006) and δ13C data from Sofular Cave (Göktürk et al., 2011), and Total Solar Irradiance curve (Egorova et al., 2018) are also shown.
Three lacustrine archives in NW Anatolia confirm the LIA drought postulated here. Ülgen et al. (2012) demonstrated a cold and dry episode between AD 1300 and 1925 by using geochemical proxies and the minimal occurrence of Mediterranean xerophytes in Lake İznik (Figure 1a). The low resolution of their record does not allow further climatic subdivisions. The nearby Lake Çubuk with almost identical lake area/watershed area metrics to Lake Sünnet started to shoal after AD 1500 and experienced the shoalest conditions around AD 1700 that match with the Maunder Minimum (Ocakoğlu et al., 2016a). The composite tree-ring studies carried out on the submerged erect dead trunks and the living trees around Lake Sülüklü (Figure 1b) revealed that the lake area was completely desiccated and hence suitable for tree growth between AD 1645 and 1703 (Aytuğ and Kılıç, 1986), again closely matching with the Maunder Minimum (Figure 10). The subsequent rise in precipitation should have resulted in the resumed lake development and the submersion and death of lowland vegetation. It is surprising that the thickness of tree-rings during the Maunder Minimum in Lake Sülüklü is relatively high (Figure 10). This fact may imply that the LIA drought in this part of Turkey is associated with a radical decrease of fall and winter precipitation as tree-ring thickness basically records the May–July humidity (Touchan et al., 2007).
The stalagmite δ13C record from Sofular Cave (Göktürk et al., 2011) displays an overall positive shift the summit of which matches with the Maunder Minimum and desiccation of Lake Sülüklü (Figure 10). The authors suggest that the more positive δ13C values in this record basically represent decreased spring-to-fall precipitation. Similarly, the LIA record in Lake Simav in W Anatolia (Figure 2a), even though poorly dated, is represented by noticeably dark brown lake muds with a remarkable herbaceous pollen peak mainly of Asteraceae, Chenopodiaceae, and Caryophyllaceae grossly between AD 1400 and 1700 (Ocakoğlu et al., 2019). As to the annually resolved tree-ring record in SW Anatolia near Antalya and Mersin (Figure 1a), Touchan et al. (2007) demonstrated that the period AD 1591–1660 was one of the driest periods experienced in the last millennia. Similarly, Ca-proxy for the drought in Lake Salda from the same region (Figure 1a) points out a dry period between AD 1580 and 1700 (Danladi and Akçer-ön, 2018). These two facts confirm that the LIA drought expanded through the Mediterranean coast in the late 17th century. In Lake Nar to the east, the LIA started almost suddenly at AD 1400 with a positive shift of δ18O and continued with some warm spells until the 1900s (Jones et al., 2006). The authors suggest that the summer rainfall at this time increased as a result of teleconnections with Indian Mansoon and the winter precipitation was adversely impacted due to the North Sea-Caspian Sea climate pattern. Palynological investigation of the Lake Nar sediments revealed that the LIA climate shift has no discernable impact on vegetation and land use. On the contrary, human activity has been the main driver in the last 2000 years (England et al., 2008). Two records with comparatively lower resolution in the further east display a contrasting climate pattern for some unknown reasons during the LIA. Lake Tecer record (Figure 2a) experiences evaporate/aragonite free deposition implying a relatively cool and humid period between AD 1450 and 1750 (Kuzucuoğlu et al., 2011). A contemporaneous increase in arboreal pollen near Kültepe is similarly interpreted as the prevalence of more humid conditions during the LIA (Şenkul et al., 2018).
The exit from the cold and dry LIA conditions seems to have occurred differently in NW Anatolian records. Between AD 1750 and 1825, the Lake Sünnet proxies indicate increasing lake depth, warming, and subsequent cooling. This period is typical with increased spring-to-fall precipitation (i.e. more negative δ13C) in the Sofular record (Figure 10). Lake Sülüklü tree-ring record confirms an early (AD 1750–1800) humid period followed by a dry one (Figure 10). Interestingly, the Dalton Minimum (AD 1800–1850) match precisely with the weak plant growth in Lake Sülüklü and overwhelming appearance of Artemisia in Lake Sünnet (Figure 10).
From the 1850s to the early 1900s, the Lake Sünnet records indicate the steady lake level and rather warmer conditions. Moreover, the widespread impact of humans over vegetation became evident by increased Cerealia-type pollen. However, the total tree pollen and that of individual trees were not changed considerably likely due to the still reduced human population. At this time, a progressive precipitation decrease in the Sofular record matches with the weak tree-ring development in Lake Sülüklü (Figure 10). Warming and more humid conditions in Lake Salda started in AD 1835s and continued up to date (Danladi and Akçer-ön, 2018). As to Lake Nar, a shift to more humid conditions occurred by the 1850s, interrupted during AD 1900–1950, and then resumed heading up to date (Jones et al., 2006). Since the 1920s, the water level in Lake Sünnet has been rising steadily parallel to the increasing precipitation (negative shift of δ13C) in Sofular Cave and the thicker growth rings in Lake Sülüklü (Figure 10). Nevertheless, the progressive increase of the δ18O record in Lake Sünnet is regarded as the indication of the worsening summer drought.
The LIA cooling/drought in different parts of Anatolia discussed above has a considerable impact upon the Ottoman Society. Kuru (2019), using agricultural and demographic data from the west and central Anatolia of the time, demonstrated a social boom (i.e. expansion of agricultural fields and the resultant population increase) as a response to warm and humid climate during the reign of Suleiman the Magnificent until AD 1580s. The appearance of the LIA drought overlaps with a significant shortage in agricultural production, large-scale land-use change, and widespread social unrest known as the Celali Rebellion in the early 17th century (Griswold, 2010). White (2013) asserted that the Celali Rebellion marks the onset of century-long social unrests that were fueled by a background dry climate and were terminated with the decisive retreat of the Ottoman Army from the Balkans. The last wave of this prolonged drought extending from Greece to Anatolia culminated in AD 1685–1687 (White, 2013). Conversely, Roberts (2021) suggested that the unexpectedly heavy impact of the relatively short (AD 1594–1609) drought leading to the Celali Rebellion in central Anatolia, is rather related to the higher vulnerability of the Ottoman Empire due to excessive geographic expansion. The end of the LIA (early 18th century, beginning of LPZ 2-2 in this study) that is climatically characterized by warmer and dryer conditions with extreme weather in the Mediterranean region resulted in the rise of commercial agriculture and the fall of mobile pastoralism (Duffy, 2020). Much later, two decadal drought events in AD 1842 and AD 1869, the latter of which is clearly distinguishable in the δ18O record in Lake Sünnet, similarly caused important social unrests (Erler, 2012).
Conclusion
We studied the paleoclimatological history of NW Anatolia for the last 500 years thus including the Little Ice Age by using proxy data of three cores collected from different parts of Lake Sünnet and correlated the findings with previous data across Anatolia. Landslide-damming origin of the lake in mountainous highlands allows us tracking of climatic changes in 5 to 10-year resolution in different proxies. Accordingly, the period AD 1510–1750 of the LIA, as demonstrated by scarce diatom and ostracod fauna, is represented by shoal aquatic conditions. AD 1640–1710 interval in this period is characterized by a pollen association (LPZ 2-1) reflecting drier and colder climatic conditions. The nearby Lake Sülüklü was completely desiccated and became a site of tree growth in this time period. During this period, perfectly matching with one of the lower solar insolation periods known as Maunder Minimum, the Sofular stalagmite δ13C record suggests an important decline in spring and fall precipitation. Since the SW, S, and central Anatolian lake and tree-ring records, along with the Lake Sülüklü and other lake records from NW Anatolia, demonstrate a contemporaneous drought, we conclude in an Anatolian-wide LIA drought. Historical records and evaluations agree on the vital impact of the century-long discontinuous drought upon the onset of the recession of the Ottoman Empire by the earliest 17th century. The proxy data from Lake Sünnet demonstrates a gradual shift from harsher to relatively humid and warm conditions after the 1750s. Interrupted by a colder arid period between AD 1800 and 1850, this gross climatic trend has continued until the end of the last century with some fluctuations in precipitation.
Footnotes
Acknowledgements
We are grateful to Prof. İsmail Ömer Yılmaz (Middle East Technical University, Turkey) and Osman Kır (Geologist, MSc.) for their contribution to the core description and the sample preparation respectively. We also appreciate Prof. Mehmet Serkan Akkiraz (Dumlupınar University, Turkey) who kindly drew the TILIA pollen diagram and helped with its interpretation. Clay Campbell (University of Kansas) did the grammar edit of the manuscript. Prof. Suzanne AG Leroy and another anonymous referee greatly contributed by reviewing the manuscript.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The Commission for Scientific Research Projects of Eskişehir Osmangazi University funded this study with a grant number 200915009.